The floodplains of Amazonia's Rio Negro Basin are covered with 101000 km2 of seasonally flooded woody vegetation (Melack & Hess Reference MELACK, HESS, Junk, Piedade, Wittmann, Schöngart and Parolin2010). This habitat is home to a rich tree flora, exceeding 100 species in a single hectare (Ferreira Reference FERREIRA1997). It sustains endemic invertebrates (Adis et al. Reference ADIS, ERWIN, BATTIROLA, KETELHUT, Junk, Piedade, Wittmann, Schöngart and Parolin2010) and a high diversity of fish. Fruits and seeds have been found in the stomachs of 79 fish species in the Rio Negro, some of which are almost exclusively frugivorous (Goulding et al. Reference GOULDING, CARVALHO and FERREIRA1988).
Contrary to intuition, Amazonian black-water floodplain forests are susceptible to ground fire during their annual terrestrial phase. Low dry-season rainfall and floodplain forest fires are coincident with El Niño (e.g. 1926), with the warm phase of the Tropical Atlantic north–south sea-surface temperature gradient (2005, 2010) or with both sequentially (1997–1998). These drought extremes may become more frequent and severe in the future (Cox et al. Reference COX, BETTS, COLLINS, HARRIS, HUNTINGFORD and JONES2004, Reference COX, HARRIS, HUNTINGFORD, BETTS, COLLINS, JONES, JUPP, MARENGO and NOBRE2008).
Tree mortality from fire is much higher in black-water floodplains than in adjacent upland forest when the same ground fire penetrates into both forest types (Nelson Reference NELSON2001). Post-fire regrowth is slower in the floodplain and reburning may arrest succession (Flores et al. Reference FLORES, PIEDADE and NELSON2013). In seasonally flooded forest of low height and open canopy in the upper Rio Negro, the litter layer becomes dry and flammable after a few rainless days (Kauffmann et al. Reference KAUFFMANN, UHL and CUMMINGS1988, Uhl et al. Reference UHL, KAUFFMANN and CUMMINGS1988).
It is important to measure the fuels in black-water floodplain forest and how these may change based on decomposition, depth of annual inundation and soil texture because this gives us an idea of how fires are generated, why tree mortality from fire is high and how fires may increase in the future. We tested three hypotheses: Leaf decomposition is slower in seasonally inundated forest compared with adjacent upland forest (H1); Fine-fuel biomass (litter + associated root mat) is higher in the floodplain forest (H2); Within the floodplain forest, the length of annual inundation (H3a) and the soil sand content (H3b) are positively correlated with fine-fuel biomass.
To test H1, we compared leaf decomposition rates using litter bags placed on the floor of non-flooding upland forest and on a submerged forest floor at high-water stage in a backwater of the Rio Negro. Submerged bags were at two different depths: <1.5 m and 4–6 m. Oxygen saturation at the study site for these two depths is 40–150% and 0–40%, respectively (Aprile & Darwich Reference APRILE and DARWICH2009). We used Clitoria fairchildiana Howard as our standard leaf, an Amazonian tree species used as a standard in other litter decomposition studies (Luizão & Schubart Reference LUIZÃO and SCHUBART1987, Luizão et al. Reference LUIZÃO, LUIZÃO and PROCTOR2007). Its leaves are of medium size (73 ± 21 cm2). Specific leaf area is typical for a tropical rain-forest tree (132 ± 30 cm2 g−1) (Poorter & Rozendaal Reference POORTER and ROZENDAAL2008). Values are mean ± 1 SD, n = 31.
We prepared 240 numbered litter bags of 30 × 30 cm using 1-mm nylon mesh with 20 holes of 10-mm diameter to allow access by invertebrates. Each bag received about 4 g of air-dried whole leaves. We recorded the initial mass of each bag's contents with a precision of 0.01 g. Using small weights, 80 bags were submerged on the litter layer of the inundated floodplain forest at the shallow, oxygen-rich depth. Another 80 bags were submerged on the litter layer below the thermocline where dissolved oxygen is depleted. A third set of 80 bags was placed on the litter layer of nearby upland forest. All bags were placed in the field on 27 May 2010, 1 mo before the end of the rising-water phase, to take advantage of a nearly stable high-water level over the following 11 wk. This minimized the need for repositioning the submerged bags to maintain their depth (done only once). Late May is the end of the rainy season, so decomposition in the upland forest was during a period of low rainfall.
We retrieved a subset of 16 bags from each of the three treatments every ~16 d. Weight loss from decomposition was determined by washing off sediment, oven drying at 60 °C and correcting the weight to an air-dried equivalent. Litter was discarded after weighing and undisturbed litter bags were retrieved at each collection date. To further maintain independence between observations over time, we had allocated the litterbags across four different sites per treatment. The experiment was ended at the fifth collection on day 81 as the Rio Negro began to drop rapidly.
Litter decomposition over time is described by an exponential decay model (Barlöcher Reference BARLÖCHER, Graça, Barlöcher and Gessner2007):
 \begin{eqnarray}
M_t = M_0 e^{ - kt}
\end{eqnarray}
\begin{eqnarray}
M_t = M_0 e^{ - kt}
\end{eqnarray}
 \begin{eqnarray}
{\rm In}\,[M_t ] = {\rm In}\,[M_0 ] - kt
\end{eqnarray}
\begin{eqnarray}
{\rm In}\,[M_t ] = {\rm In}\,[M_0 ] - kt
\end{eqnarray}
Conveniently, k for each treatment is the slope of equation 2 when fitted to that treatment's data. Variance of ln[Mt] is zero at t = 0, so we used data from the first date of litterbag retrieval onward. The 95% confidence intervals were used to compare the slopes of the three individual regressions. Exploration of the data showed much smaller residuals for one of the regressions, so we could not compare slopes by testing pairwise for interaction between treatment and time (t).
To test H2, we compared fine fuel stocks between flooded and upland forests. Fine fuel on the forest floor is all dead organic material under 2.5 cm thickness and located above the mineral soil. These rapidly drying fuels (Kauffmann et al. Reference KAUFFMANN, UHL and CUMMINGS1988) were divided into two layers, the root mat mixed with humus above the mineral soil and the litter above the root mat. We sampled 29 sites during the low-water stage of the seasonally flooded forest and 28 sites in the upland forest. These were spread across a radius of 20 km from the mouth of the Cuini River (63°08′W 0°45′S), a black-water tributary of the middle Rio Negro. Seasonally flooded forest sites were all located on alluvial deposits. At each site we established a 200-m-long sampling line following a single elevation. At five equally spaced points along the line we measured the thickness of each of the two fine-fuel layers in a soil pit. At one of the five points we collected both fuel layers under a surface area of 20 × 20 cm. All but the finest live roots were removed and the remaining mostly dead biomass was oven-dried at 105 °C and weighed. Linear regressions were developed between litter layer thickness and its dead biomass and between root-mat thickness and its dead biomass. Separate regressions were obtained for the upland and for the floodplain biomass samples. These were used to estimate dry mass from fuel-layer thickness at the four points per transect where we only measured thickness.
We tested H3 within the floodplain sites using a multiple linear regression. The independent variables were the height of the high-water mark on a tree trunk at each sampling line (an indicator of submergence duration) and the sand content (%) of the upper mineral soil. Total fine-fuel load was the dependent variable.
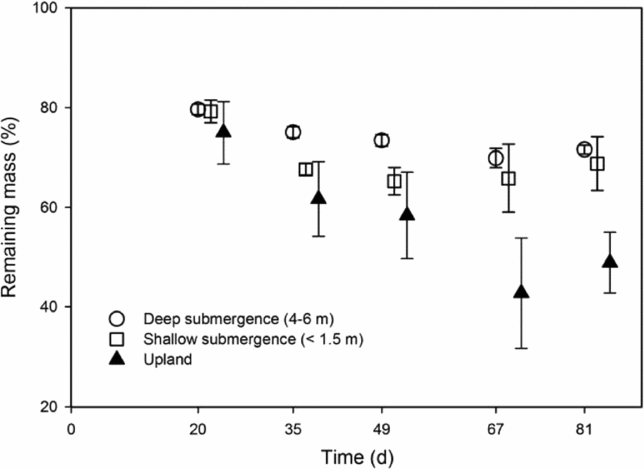
As predicted by H1 (Figure 1), leaf decomposition was faster on the upland than in shallow or deep water (P < 0.05). But decay was not slower in deep oxygen-depleted water compared with shallow water. Leaf decomposition constants were 0.0092 ± 0.0049 (95% CI) in the upland forest, 0.0028 ± 0.0013 in shallow water and 0.0018 ± 0.0002 in deep oxygen-depleted water. Explained variance was low, with R2 = 0.18, 0.23 and 0.50 for the upland, shallow and deep-water regressions, respectively.

Figure 1. Decomposition of Clitoria fairchildiana leaves submerged in black-water floodplain and on adjacent upland forest floor of the lower Rio Negro in the Central Brazilian Amazon. Error bars are 95% CI of the mean.
Both the root mat and the overlying litter layer held more fine fuel in the floodplain forest than in the upland forest (Table 1), as predicted by H2. All differences between the two habitats were highly significant (t-tests, P < 0.001). Contrary to the prediction of H3a, we found no relationship between height of the water mark in the flood zone and the combined root mat plus litter layer fine-fuel load (P = 0.67). H3b was also not supported: within the floodplain sites, soil sand content ranged from 0.1% to 92%, but had no effect on total fine-fuel load (P = 0.65). Within the upland sites, soil sand content also had no significant effect on fine-fuel load (P = 0.07). The flooded-forest soils had less sand (17% ± 28%) than the upland forest soils (34% ± 18%). Yet in the floodplain a root mat was present at 63% of the sample points when we measured fuel thickness, but only at 11% of the points in the upland forest.
Table 1. Fine-fuel loads compared between black-water floodplain forest and upland forest of the middle Rio Negro in the West Central Brazilian Amazon. Confidence interval is 1 SD.

We infer that slower leaf decomposition leads indirectly to higher tree mortality after fire in floodplain forests by allowing more fine fuel to accumulate and by leading to the formation of a carpet-like root mat above the soil where tree roots are vulnerable to fire. A mat of apogeotropic nutrient-seeking roots forms in Amazonian forests whenever slow decomposition leads to an accumulation of fine litter and humus (Cuevas & Medina Reference CUEVAS and MEDINA1986, Sanford Reference SANFORD1987).
Why do submerged leaves decompose more slowly? Amazonian black-water rivers and streams lack the abundant invertebrate leaf shredders which facilitate decomposition in temperate streams (Wantzen et al. Reference WANTZEN, YULE, MATHOOKO, PRINGLE and Dudgeon2008). The litter layer also lacks termites during its submerged phase. These have been found to remove more than 40% of decomposing leaf litter in Amazonian upland forest (Luizão & Schubart Reference LUIZÃO and SCHUBART1987). Leaves in our upland-forest litter bags had many small circular excisions typical of termites. Large residuals found when fitting the upland data to equation 2 suggest a patchy harvesting behaviour, which is typical of termites.
Litter decomposition rates in black-water forests will also be lowered by the sclerophyllous leaves with high C:N ratios found in this nutrient-poor environment. But studies are lacking comparing C:N ratios and decomposition rates for a representative mix of upland leaves against a mix of leaves from black-water forest.
Two other studies in Amazonia have compared leaf decomposition for the same species across flooded and non-flooded environments. On a riparian forest floor in the Colombian Amazon, Rueda-Delgado et al. (Reference RUEDA-DELGADO, WANTZEN and TOLEDO2006) found slower decomposition of leaves of Cecropia latiloba, Tessaria integrifolia and Symmeria paniculata during the submerged season compared with the mostly non-submerged low-water season. For two tree species in the Ecuadorean Amazon, Capps et al. (Reference CAPPS, GRAÇA, ENCALADA and FLECKER2011) also found slower leaf decomposition for permanently submerged and intermittently submerged leaves in the Tiputini River, compared with non-submerged upland forest.
Our study found a two–three-fold difference in leaf decomposition rates between upland and submerged litterbags, consistent with a similar ratio in the total standing stock of fine fuel between upland and floodplain (Table 1). The root mat with humus was responsible for most of this difference in fine-fuel load between the two forest types. Total fine-fuel load and root-mat thickness did not increase with the depth of submergence in the floodplain forests, contrary to the observations of Klinge (Reference KLINGE1973) on the lower Rio Negro. However, the annual flood pulse on the lower Rio Negro has almost twice the amplitude as at our study site on the middle Rio Negro, so any change in fuel load with depth will be less evident at our site.
ACKNOWLEDGEMENTS
The Coordination for the Improvement of Higher Level Personnel (CAPES) provided a fellowship to ARS. The GEOMA Environmental Modeling Network and the National Institute for Amazon Research provided field support. Bruce Forsberg, Manuel Graça, Paulo Graça, Flávio Luizão, Regina Luizão, John Melack, Euler Nogueira, Maria Piedade, Jochen Schöngart and two anonymous reviewers provided valuable suggestions in the planning and revision stages. João dos Santos and Amir Luiz Alves da Silva provided field assistance.