


1. Introduction
Jiangxichelys ganzhouensis Tong et Mo, Reference Tong and Mo2010 is a moderate-sized nanhsiungchelyid turtle from the latest Cretaceous Nanxiong Group in Jiangxi Province, southern China (Tong & Mo, Reference Tong and Mo2010). The species was based on a single shell that, although complete, lacks many details of sutures and sulci. Here we report on additional material of this species which includes four shells, one of them associated with a partial skull. Our study provides additional information on the morphology of the taxon and also phylogenetic and palaeogeographical information.
The new specimens were collected during extensive urban construction at Longling Town, Nankang city, about 20km SW of Ganzhou City, Jiangxi Province (Fig. 1). In recent years, the red beds of the latest Cretaceous Nanxiong Group of the Longling area have yielded abundant and diverse vertebrate fossils, including sauropod and theropod dinosaurs (Lü et al. Reference Lü, Yi, Zhong and Wei2013 a, b, Reference Lü, Yi, Brusatte, Yang, Li and Chen2014; Wei et al. Reference Wei, Pu, Xu, Liu and Lü2013) as well as lizards, turtles and dinosaur eggs. The turtle specimens studied here are housed in the Mineral Resources Administration of Ganzhou. In this paper, we follow Zangerl (Reference Zangerl, Gans, Bellairs and Parsons1969) for the nomenclature of the shell and Gaffney (Reference Gaffney1979) for that of the skull.

Figure 1. Map showing the location of the type locality (Ganxian) and new locality (Longling, Nankang) of Jiangxichelys ganzhouensis Tong & Mo, Reference Tong and Mo2010.
2. Systematic palaeontology
Testudines Linnaeus, Reference Linnaeus1758
Cryptodira Cope, Reference Cope1868
Nanhsiungchelyidae Ye, Reference Ye1966
Jiangxichelys Tong & Mo, Reference Tong and Mo2010
Type species: Jiangxichelys ganzhouensis Tong & Mo, Reference Tong and Mo2010
Emended diagnosis: A genus of Nanhsiungchelyidae of moderate size, without midline keel; cervical notch well developed, triangular and wide, and first vertebral vase-shaped. Different from all other nanhsiungchelyids in having the first marginal scute short and wide with a long free edge and long contact with the second marginal; the second marginal is boot-shaped with a medial process reaching the first vertebral, preventing contact between the first marginal and the first pleural scute. In addition, it differs: from Hanbogdemys (including Bulganemys) in having a larger and triangular first suprapygal, a large and triangular fifth vertebral extending onto the tenth peripheral and narrower lateral marginal scutes; from Nanhsiungchelys and Anomalochelys in the smaller cervical notch, wider than long cervical scute, narrower neurals and narrow vertebrals; from Kharakhutulia in having a longer anterior plastral lobe which extends beyond the anterior edge of the carapace and pentagonal entoplastron; from Basilemys in the larger cervical notch, wider than long cervical scute and narrower peripherals; from ‘Zangerlia’ dzamynchondi in the shallower cervical notch, longer epiplastron and intergular midline length; and from ‘Zangerlia’ ukhaachelys in having a pentagonal entoplastron, a straight pectoroabdominal sulcus and the abdominal scute longer than the pectoral scute at the midline. The skull differs from Nanhsiungchelys in having deep temporal and cheek emarginations, with a narrow bar between these two emarginations and between the orbit and the temporal emargination.
Included species: Jiangxichelys neimongolensis (Brinkman & Peng, Reference Brinkman and Peng1996).
Distribution: Late Cretaceous of China.
Jiangxichelys ganzhouensis Tong & Mo, Reference Tong and Mo2010
(Figs 2–6)
Holotype: NHMG 010415, a shell, collection of the Natural History Museum of Guangxi Zhuang Autonomous Region, Nanning, Guangxi Province, China (Tong & Mo, Reference Tong and Mo2010).
Type locality and horizon: Ganxian, NE of Ganzhou city, Jiangxi Province, China; Nanxiong Formation, latest Cretaceous (Tong & Mo, Reference Tong and Mo2010).
Diagnosis: A species of Jiangxichelys. Differs from J. neimongolensis in having a pentagonal entoplastron; a straight intergularohumeral sulcus which is located far anterior to the entoplastron; and the gulars greatly reduced to absent (when present, they are situated at the lateral end of the intergularohumeral sulcus).
Referred material: JXGZ(2012)-178: a shell with very damaged carapace and complete plastron; JXGZ(2012)-179: a partial skeleton including a damaged carapace articulated with a nearly complete plastron, associated with a partial skull articulated with the lower jaw, and part of the left forelimb; JXGZ(2012)-180: a damaged partly restored carapace without plastron; JXGZ(2012)-182: a shell with articulated carapace and plastron, the right portion of both carapace and plastron is damaged; all from Longling town, Nankang city, Ganzhou, Jiangxi Province, China.
Measurements: see Table 1.
Table 1. Measurements of Jiangxichelys ganzhouensis Tong & Mo, Reference Tong and Mo2010 (in centimetres).

2.a. Description
Skull: The skull, preserved only in JXGZ(2012)-179 (Fig. 2), is incomplete, the snout being missing. The skull extends partially out of the carapace and is in a raised position. The skull is tilted towards the left, so that the right side is better exposed. The morphology of the skull closely resembles that of Jiangxichelys neimongolensis (Brinkman & Peng, Reference Brinkman and Peng1996; Brinkman et al. Reference Brinkman, Tong, Li, Sun, Zhang, Godefroit and Zhemin2015). The skull roof is broad and the orbits are facing laterally. The cheek emargination is deep with its upper margin higher than the lower margin of the orbit. Its shape matches that of J. neimongolensis. The temporal emargination is very deep, extending anterior to the cheek emargination and separated from the orbit by a narrow bar. The cavum tympani is round and deep. The incisura columellae auris is closed. The skull roof scales are not visible, contrary to J. neimongolensis in which a network of scutes of moderate size covers the skull roof.

Figure 2. Jiangxichelys ganzhouensis, JXGZ(2012)-179, skull in (a, b) right lateral, (c, d) inclined dorsal and (e, f) front views. Scale bar = 5 cm. Abbreviations: ang – angular; art pro – articular process of quadrate; den – dentary; fr – frontal; ju – jugal; lj – lower jaw; mx – maxilla; pa – parietal; po – postorbital; qu – quadrate; sq – squamosal; sur – surangular.
The maxilla is deep below the orbit and meets the jugal posterodorsally. Posteriorly it forms the anterior margin of the cheek emargination. The postorbital forms the narrow bar between the temporal emargination and the orbit. It contacts the frontal medially and the jugal laterally. There is also a short contact with the parietal posterior to the frontal. The frontal-parietal suture is transverse and posterior to the orbit as in J. neimongolensis. The jugal has a narrow exposure on the skull roof; it forms part of the lower margin of the orbit and the upper margin of the cheek emargination. The squamosal is a conical bone that contacts the postorbital anteriorly and the quadrate ventrally. The suture of the quadratojugal is unclear so the relationships with the surrounding bones are uncertain. The broken snout region exposes the nasal passages, connecting the fossa nasalis and the apertura narium interna that is bordered by the maxilla laterally and the prefrontal dorsally. The fissura ethmoidalis is rounded in front view.
Lower jaw: The lower jaw is attached to the skull and only the outer surface is exposed. The lower jaw is low, with an unfused symphysis as in J. neimongolensis. The outer surface of the lower jaw is mostly formed by the dentary, the surangular forms the posterodorsal part and the angular has a small exposure on the posteroventral end of the lower jaw. The coronoid process appears to be lower than that of J. neimongolensis (Brinkman & Peng, Reference Brinkman and Peng1996). The retroarticular process is not developed.
Two slender hyoid bones are preserved, exposed ventrally.
Carapace: Four additional shells are available (Figs 3–5), although none of them has a complete carapace. The carapace is low domed with a flat top. It has an almost circular outline, with the length only slightly greater than the width. The cervical notch is wide and relatively deep. It is bordered by the nuchal and the first peripherals, not by the nuchal only as in Anomalochely angulata (Hirayama et al. Reference Hirayama, Sakurai, Chitoku, Kawakami and Kito2001).

Figure 3. Jiangxichelys ganzhouensis, JXGZ(2012)-179, shell in (a, b) dorsal, (c, d) ventral, (e, f) left lateral and (g, h) posterior views. Scale bar = 10 cm.
The nuchal is trapezoidal and wide, with forward convergent lateral margins. Its free margin contributes to the middle portion of the cervical notch. The complete neural series is preserved in JXGZ(2012)-180 and JXGZ(2012)-182, with a neural formula of 6<4>6>6>6>6>6>6 that reaches the suprapygal 1. The neurals are narrow, except for neurals 7 and 8 as seen in JXGZ(2012)-180 which are wider than long. There are two suprapygals; the second is much larger than the first. The pygal preserved in JXGZ(2012)-178 is wider than long as in the holotype, while in JXGZ(2012)-182 it is slightly longer than wide. The first costal is longer than the second costal. The lateral and posterior peripherals are high and the posterior peripherals are subvertical. The posterior margin of the carapace turns inward.
The cervical scute, as preserved in JXGZ(2012)-179, is short and wider than long. The first vertebral scute is vase-shaped as in the holotype, with the front part restricted to the nuchal. The second and the third vertebrals are narrow, with length nearly twice their width. The fourth vertebral is also longer than wide, but shorter than the third. The fifth vertebral is large and triangular as in the holotype, extending onto the tenth peripheral. The pleural scutes extend greatly onto the peripherals. The first and second marginal scutes are well preserved in JXGZ(2012)-179, JXGZ(2012)-180 and JXGZ(2012)-182. The morphology of these scutes matches that of the holotype in that the first marginal is short and wide along the anterior boarder of the carapace; the second marginal has a medial process extending onto the nuchal, reaching (JXGZ(2012)-182) or nearly reaching (JXGZ(2012)-180) the first vertebral. The marginals are all much lower than the corresponding peripherals except the 11th and 12th marginals, which are greatly expanded and cover the posterior half of the second suprapygal.
Plastron: The plastron is complete in JXGZ(2012)-178 and JXGZ(2012)-179 and nearly so in JXGZ(2012)-182. The anterior margin of the plastron extends beyond that of the carapace as in the holotype. The anterior end of the plastron develops a large lip on the dorsal side with a sculptured surface, indicating that the area was covered by scutes. The bridge is long and narrow. The anterior lobe matches that of the holotype. It is triangular and long, with a narrow anterior end. The front border is rounded. The lateral margins are pinched at the lateral ends of the gular/humeral sulcus. The posterior lobe is short and wide, being shorter and wider than that of the holotype. The posterior border is almost straight, and an anal notch is absent. The length proportions are bridge>anterior lobe>posterior lobe.
The epiplastron is large and has a long midline contact with its counterpart. The midline length of the epiplastron is almost equal to that of the entoplastron. The entoplastron is a large pentagonal plate that is much wider than long, with a convex anterior margin and an almost straight posterior rim. It is located posteriorly with the posterior end at the level of the line connecting the bottoms of the axillary notches. The hyoplastron is longer than the hypoplastron at the midline. The hypoplastron-xiphiplastron suture is close to the base of the posterior lobe, so that the contribution from the hypoplastron to the posterior lobe is small. The xiphiplastron is triangular and its length is roughly equal to its width.
The intergular is unusually large with a nearly transverse posterior sulcus that is located well anterior to the entoplastron. In JXGZ(2012)-179 and JXGZ(2012)-182, this scute appears to be single and no midline sulcus is discernible; in JXGZ(2012)-178, the intergular is at least divided anteriorly (Fig. 4). The gulars are greatly reduced in size; they are very small and are widely separated from one another by the large intergular in JXGZ(2012)-179 (Fig. 3) and absent in JXGZ(2012)-180 (Fig. 4). The gulars, when present, are located on the lateral border of the anterior lobe instead of on the anterolateral corner of the anterior lobe as in other turtles. The humeral scute is much longer laterally than medially. The humeropectoral scute is strongly convex forwards, covering the posterior half of the entoplastron as in other nanhsiungchelyids. The pectoral scute is triangular, with its midline length more than four times its length at the lateral end. It is excluded from the axillary notch as in other nanhsiungchelyids. The abdominal is the largest scute of the plastron, with straight anterior and posterior margins. The femoral and anal scutes have a similar midline length. The femoroanal sulcus is convex forwards and located far posterior to the hypoplastron/xiphiplastron suture. There is a complete row of five inframarginal scutes preserved on the left side of JXGZ(2012)-179; the first to the fourth are very narrow and the fifth is much wider. The almost complete inframarginals are preserved on both sides of JXGZ(2012)-178 and their shape matches that of JXGZ(2012)-179, while the exact number is not determinable.

Figure 4. Jiangxichelys ganzhouensis. Upper: JXGZ(2012)-180, carapace in dorsal view. Lower: JXGZ(2012)-178, plastron in ventral view. Scale bar = 10 cm.
Limb bones: A partial left forelimb is preserved in JXGZ(2012)-179. It is mostly covered by matrix; no detail is visible. A partial left hind limb is preserved in JXGZ(2012)-182. The femur is incomplete, with both ends damaged and exposed in ventral view. The shaft is slightly curved and two trochanters are separated by a large intertrochanteric fossa.
3. Discussion
3.a. Systematic assignment
The specimens described above are referred to Jiangxichelys ganzhouensis Tong & Mo, Reference Tong and Mo2010 on the basis of the similarities in the shell, including the almost circular carapace outline, the relatively large and deep cervical notch, the wider than long cervical scute, the morphology of the first and second marginal scutes and the pleural scutes extending well onto the peripherals. The plastron of the new specimens also resembles that of the holotype of J. ganzhouensis, particularly in the long triangular anterior lobe that extends beyond the anterior margin of the carapace, with a rounded anterior border and a long midline suture of the epiplastra. The shape of the posterior lobe of the plastron varies, however: it is narrow and relatively long in the holotype and shorter and wider in JXGZ(2012)-179 and JXGZ(2012)-180. This may be related to sexual dimorphism.
Although the holotype of J. ganzhouensis is an almost complete shell, the carapace underwent a distortion and the plastron lacks many sutures and sulci (Tong & Mo, Reference Tong and Mo2010). The new specimens add to our understanding of the shell morphology of J. ganzhouensis in the following details: the carapace is almost circular, with its length only slightly greater than its width and widest at the midlength of the shell; the complete series of eight neurals reaches the suprapygal 1 with a neural formula of 6<4>6>6>6>6>6>6; most neurals are narrow except for neurals 7 and 8 which are wider than long; vertebrals 2 and 3 are narrow with their length nearly twice their width; vertebral 4 is also narrow, but is shorter or wider than vertebral 3; the entoplastron is pentagonal; the large intergular has a straight posterior sulcus that is located far anterior to the entoplastron; the gular scutes are greatly reduced to absent; there is a complete row of five inframarginal scutes, most of them narrow except the last one which is as wide as long; the medially expanded marginal 6 on the plastron; a plastral lip is well developed on the dorsal side of the anterior lobe; and scutes cover the dorsal surface of the lip. A new reconstruction of both carapace and plastron is shown in Figure 6.
3.b. Phylogenetic relationships
The data matrix as updated in Brinkman et al. (Reference Brinkman, Tong, Li, Sun, Zhang, Godefroit and Zhemin2015) is used here for the phylogenetic analysis. The dataset is completed for Jiangxichelys ganzhouensis based on the new material. Two characters are added as follows. Character 46: neurals (0) = narrow (J. neimongolensis), (1) = wide (Anomalochelys angulata). Character 47: cervical scute (0) = wider than long (Jiangxichelys), (1) = as wide as long (Anomalochelys), (2) = longer than wide (Basilemys). A total of 34 characters among 47 are coded in J. ganzhouensis, which represents 72% of the total number of characters instead of 40% in the previous analysis (Brinkman et al. Reference Brinkman, Tong, Li, Sun, Zhang, Godefroit and Zhemin2015). The updated dataset, including 13 taxa of Nanhsiungchelyidae with Adocus as outgroup, and 47 characters are analysed using Paup 4.0b10. All characters are unordered and of equal weight. The analysis resulted in a single tree of 68 steps, with a consistency index (CI) of 0.74, homoplasy index (HI) of 0.27, retention index (RI) of 0.71 and a rescaled consistency index (RC) of 0.52. The topology of the tree remains unchanged relative to that of Brinkman et al. (Reference Brinkman, Tong, Li, Sun, Zhang, Godefroit and Zhemin2015, fig. 5).
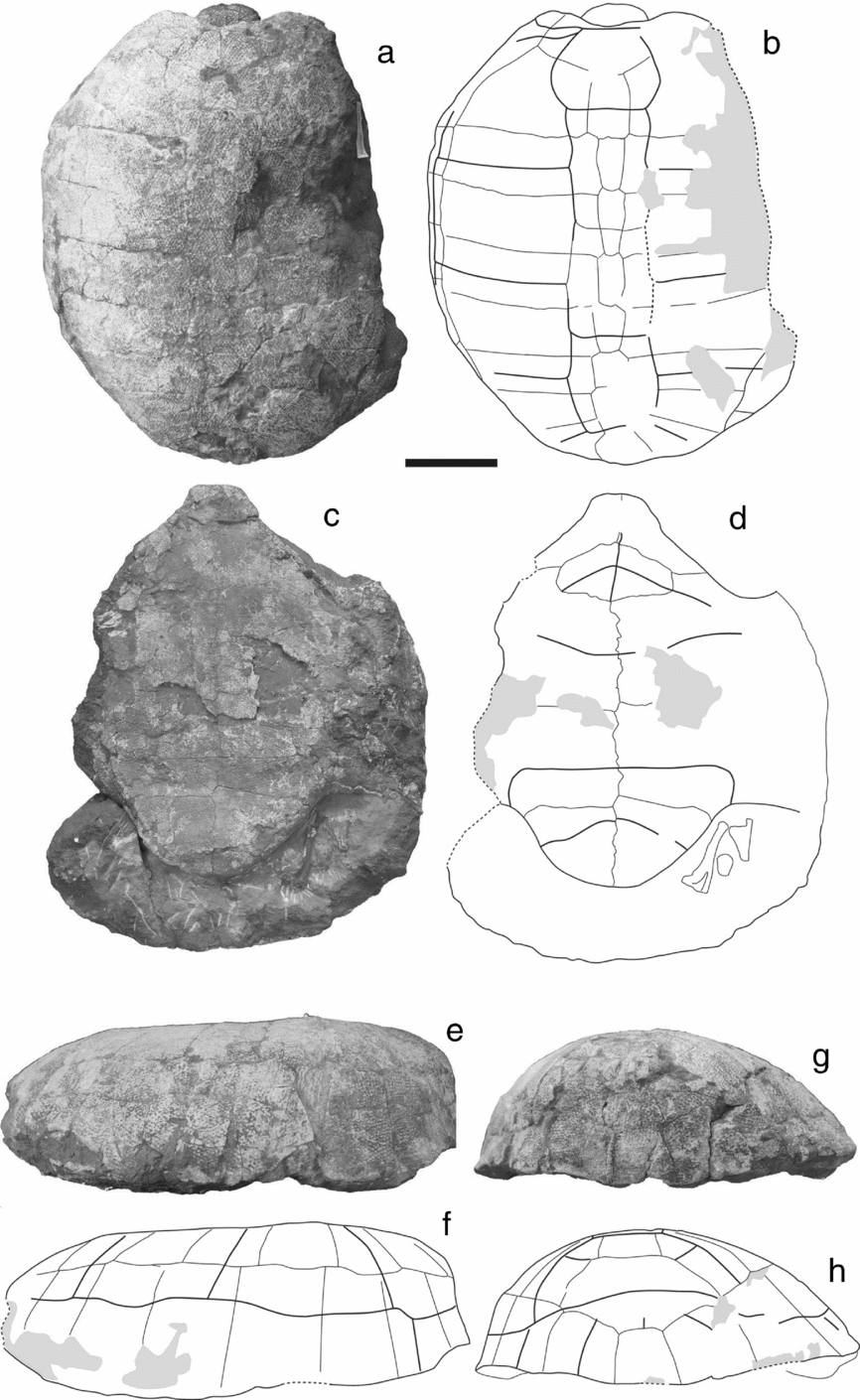
Figure 5. Jiangxichelys ganzhouensis, JXGZ(2012)-182, shell with articulated carapace and plastron in (a, b) dorsal, (c, d) ventral, (e, f) left lateral and (g, h) posterior views. Scale bar = 10 cm.

Figure 6. Reconstruction of the shell of Jiangxichelys ganzhouensis Tong & Mo, Reference Tong and Mo2010.
Our analyses continue to support the monophyly of the J. ganzhouensis – J. neimongolensis – ‘Z.’ dzamynchondi – ‘Z.’ ukhaachelys clade, and its separation from the type species of the genus Zangerlia, Z. testudinimorpha Młynarski, Reference Młynarski1972. As noted by Brinkman et al. (Reference Brinkman, Tong, Li, Sun, Zhang, Godefroit and Zhemin2015), this result is in agreement with Hirayama et al. (Reference Hirayama, Sakurai, Chitoku, Kawakami and Kito2001) and Danilov, Sukhanov & Syromyatnikova (Reference Danilov, Sukhanov, Syromyatnikova, Brinkman, Holroyd and Gardner2013), but contrary to that of Joyce & Norrell (Reference Joyce and Norell2005) and Sukhanov, Danilov & Syromyatnikova (Reference Sukhanov, Danilov and Syromyatnikova2008) which support the monophyly of the genus Zangerlia. With the new material, the similarities between J. ganzhouensis and J. neimongolensis Brinkman & Peng, Reference Brinkman and Peng1996 from Upper Cretaceous deposits of Inner Mongolia, China are even more apparent. The carapace of J. ganzhouensis is similar to that of J. neimongolensis in the following features: (1) the almost circular carapace outline; (2) the nearly vertical lateral and posterior peripherals; (3) the posterior margin of the carapace turned inwards; (4) the knob on suprapygal 1; (5) the wide and triangular cervical notch; (6) narrow neurals; (7) wider than long cervical scute; (8) vase-shaped vertebral 1; (9) narrow vertebrals 2–3; (10) vertebral 5 enlarged posteriorly, overlapping the peripherals 10; (11) pleural scutes greatly extending onto the peripherals; (12) wide and short marginal 1; and (13) the medial process of marginal 2 (Brinkman & Peng, Reference Brinkman and Peng1996; Brinkman et al. Reference Brinkman, Tong, Li, Sun, Zhang, Godefroit and Zhemin2015). The characters 1, 4, 5, 7 and 11 are shared also with ‘Z.’ dzamynchondi Sukhanov & Narmandakh, Reference Sukhanov and Narmandakh2006 (Sukhanov, Reference Sukhanov, Benton, Shishkin, Unwin and Kurochkin2000; Sukhanov & Narmandakh, Reference Sukhanov and Narmandakh2006; Danilov, Sukhanov & Syromyatnikova, Reference Danilov, Sukhanov, Syromyatnikova, Brinkman, Holroyd and Gardner2013).
Reported for the first time, the skull of J. ganzhouensis closely resembles that of J. neimongolensis in its general morphology including: the deep temporal emargination that extends anterior to the cheek emargination, separated from the orbit by a narrow bar; the deep cheek emargination with its upper margin higher than the lower margin of the orbit; and the bar between the cheek emargination and the temporal emargination is greater than that between the temporal emargination and the orbit. The lower jaws of these two taxa are similar to one another in the unfused symphysis and low rami. The narrow postorbital bar and the deep temporal emargination on the skull and the unfused symphysis of the lower jaw are also shared with ‘Z.’ ukhaachelys (Joyce & Norell, Reference Joyce and Norell2005).
Based on the abovementioned similarities, we include ‘Zangerlia’ neimongolensis Brinkman & Peng, Reference Brinkman and Peng1996 in the genus Jiangxichelys. Although ‘Z.’ dzamynchondi shares several shell characters and ‘Z.’ ukhaachelys shares some skull features with J. ganzhouensis and J. neimongolensis, both taxa are poorly documented for the carapace. We refrain from including these taxa to the genus Jiangxichelys, pending the discovery more complete material.
J. ganzhouensis differs from J. neimongolensis mainly in features of the plastron, especially on the anterior lobe of the plastron. The anterior lobe of the plastron in J. ganzhouensis is more elongated, extending well beyond the anterior border of the carapace with a narrower front end; the midline contact of the epiplastra is longer and the entoplastron is pentagonal in shape. In J. neimongolensis the anterior lobe is shorter with a more rounded anterior border; the midline contact of the epiplastra is shorter and the entoplastron is diamond-shaped. The most notable difference is the morphology of the gular and intergular. In J. ganzhouensis the posterior sulcus of the intergular is straight and located far anterior to the entoplastron and the gulars are greatly reduced, being very small or absent; in J. neimongolensis the intergular has an angled posterior sulcus that extends to the entoplastron and the gular is a long strip.
3.c. Palaeobiogeographical implications
The close relationship between Jiangxichelys and ‘Zangerlia’ spp. is of palaeobiogeographical significance (Fig. 7). As noted by Danilov, Sukhanov & Syromyatnikova (Reference Danilov, Sukhanov, Syromyatnikova, Brinkman, Holroyd and Gardner2013), the geographical and temporal distributions of ‘Zangerlia’ spp. (including ‘Z.’ neimongolensis, ‘Z.’ dzamynchondi and ‘Z.’ ukhaachelys) were restricted to the Campanian and probably Maastrichtian deposits of southern Mongolia and adjacent areas of China in Inner Mongolia. As currently documented, Jiangxichelys ganzhouensis specimens have only been collected in the uppermost Cretaceous Nanxiong Formation of Ganzhou area, Jiangxi Province in southern China. This moderate-sized nanhsiungchelyid group flourished during latest Cretaceous (Campanian–Maastrichtian) time in both southern and northern China and Mongolia. The similarities between the southern China dinosaur assemblages and those of Mongolia and Inner Mongolia in northern China during Late Cretaceous time have been pointed out recently by Lü et al. (2015); these regions share several clades of oviraptorids. This appears to be different from the palaeobiogeographical distribution pattern during Early Cretaceous time. A distinct southern province including Thailand, Laos and part of southern China has been recognized on the basis of similar Lower Cretaceous vertebrate assemblages (Aptian–Albian) which are distinct from the coeval vertebrate faunas in the more northern parts of Asia, such as the Jehol biota. This southern province includes the mid-Cretaceous (Aptian) fauna of the Xinlong Formation in Fusui, Guangxi Province, but where its northern boundary was located remains uncertain (Mo et al. Reference Mo, Buffetaut, Tong, Amiot, Cavin, Cuny, Suteethorn, Suteethorn and Jiang2015). A difference in climate during that period was suggested to explain the geographical isolation of the southern province, on the basis of geochemical evidence (Amiot et al. Reference Amiot, Wang, Zhou, Wang, Lécuyer, Buffetaut, Fluteau, Ding, Kusuhashi, Mo, Phillippe, Suteethorn, Wang and Xu2015; Mo et al. Reference Mo, Buffetaut, Tong, Amiot, Cavin, Cuny, Suteethorn, Suteethorn and Jiang2015). Based on the fish faunas, Zhang & Miao (Reference Zhang, Miao, Arratia and Tintori2004) recognized three distinct zones in the Early Cretaceous of Asia, a western zone, a northeastern zone and a southeastern zone. The southeastern zone to which southern China belonged also included southern Korea and northern Kyushu, Japan, and is characterized by the presence of ichthyodectiforms and clupeomorphs. These SW–NE-oriented areas are parallel to the tectonic structures that control the drainage, explaining the freshwater fish distribution. Even if land animals were submitted to different constraints as compared to freshwater fauna, the Jiangxichelys–‘Zangerlia’ spp. clade provides additional evidence for only weak physical barriers affecting the dispersal of vertebrates between southern China and the northern parts of Asia in terrestrial ecosystems during Late Cretaceous time.

Figure 7. Geographical distribution of Jiangxichelys – ‘Zangerlia’ spp. clade. 1, Jiangxichelys ganzhouensis; 2, J. neimongolensis; 3, ‘Zangerlia’ dzamynchondi; 4, ‘Z.’ ukhaachelys.
Acknowledgements
We would like to thank Yanyin Wang and Yufeng Liu for their help during the examination of the specimens and Eric Buffetaut for improving the manuscript. This work is supported by the National Natural Science Foundation of China Grant 41172017, as well as the National Natural Science Foundation of China (Grant 41272022) and the China Geological Survey (Grant 12120114026801) to JC Lü.








