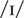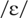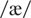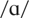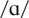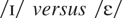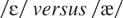INTRODUCTION
Pure word deafness (PWD) is a rare neurological syndrome characterized
by severe difficulties in understanding and reproducing spoken language,
with otherwise intact speech production and written language comprehension
(Kussmaul, 1877; Lichtheim,
1885). The deficiencies in decoding language occur at a stage of
analysis that markedly impairs the processing of speech sounds, words,
phrases, and sentences, but leaves environmental sound recognition and
identification relatively preserved. Because deficits are specific to the
auditory modality, this pattern of impairment has also been referred to as
“word sound deafness” (Kohn & Friedman,
1986; Franklin, 1989) or “verbal
auditory agnosia” (Ulrich, 1978; Wang et al., 2000). In the majority of published
reports, it is the sequelae of cerebrovascular accidents, but it has also
been described in association with tumor (Goldstein,
1948), seizures (Stefanatos, 1993; Fung et al., 2000), head injury (Seliger et al., 1991), degenerative aphasia (Mesulam, 1982; Croisile et al.,
1991; Otsuki et al., 1998), encephalitis
(Goldstein et al., 1975), and drug toxicity
(Donaldson et al., 1981).
The syndrome is of significant theoretical importance because it
provides clinical support for the modularity of speech perception as
separable from nonverbal auditory recognition systems and more central
language computational networks (Allport & Funnell,
1981; Polster & Rose, 1998; Poeppel, 2001; Pinard et al.,
2002). However, longstanding disagreement exists regarding the
nature of the pathognomonic auditory comprehension deficits and their
neurological basis (Goldstein, 1974; Buchman et al., 1986; Praamstra et
al., 1991). Phenomenological descriptions, in some cases, have
implicated a substantial dissolution of auditory perceptual processing,
sufficient in severity to impede recognition of the acoustic
characteristics of the human voice (Miceli,
1982). Speech is described in such terms as “a noise”
(Coslett et al., 1984; Buchman et al., 1986), “a hurr or buzzing”
(Mendez & Geehan, 1988), like “wind in
the trees” (Ziegler, 1952), or “the
rustling of leaves” (Luria, 1966).
However, other subjective descriptions allude to subtler disturbances,
suggesting a continuum of severity. Spoken communications are recognized
as speech, but a breakdown appears to occur early in the process of
mapping the acoustic input to lexical representations of words. Discourse
sounds like “jabbering” or “a foreign language”
(Denes & Semenza, 1975; Auerbach et al., 1982; Buchman et
al., 1986; Mendez & Geehan, 1988), or
simply does not “register” (Saffran et al.,
1976). There are also frequent suggestions that perceptual or
cognitive resources are incapable of keeping up with the rate at which
speech is produced: “words just run together” (Klein & Harper, 1956) or “come too
quickly” (Albert & Bear, 1974).
Experimental investigations of word deafness have yielded
substantively different conceptions of the functional locus of the
underlying processing disturbances. Several reports have noted fundamental
problems in basic aspects of auditory temporal processing, including
deficient intensity-duration functions (Kanshepolsky et
al., 1973) and poor resolution of temporally distinct auditory
events (Albert & Bear, 1974; Auerbach et al., 1982; Tanaka et
al., 1987; Buchtel & Stewart, 1989;
Best & Howard, 1994; Godefroy et al., 1995). Based on such findings, it has
been argued that PWD results from general limitations in fine-grained
auditory temporal analysis (Albert & Bear,
1974; Auerbach et al., 1982; Phillips & Farmer, 1990) that are particularly
detrimental to language comprehension because they impede the ability to
perceive brief spectrotemporal cues in speech that are important to the
derivation of linguistic meaning. In contrast, others have noted
systematic patterns of error on speech discrimination and identification
tasks that implicate problems at phonetic levels of analysis, a
higher-order stage of auditory processing specific to speech (Saffran et al., 1976; Caramazza et
al., 1983; Metz-Lutz & Dahl, 1984;
Praamstra et al., 1991).
There is also disagreement regarding the neuroanatomical substrate of
the syndrome. The majority of cases reported in the literature demonstrate
bilateral temporal lobe lesions, particularly involving the middle and
posterior portions of the superior temporal gyrus or underlying
geniculotemporal pathways (Bauer & Zawacki,
2000; Poeppel, 2001). However, PWD was
first described in patients considered to have unilateral lesions of the
left posterior temporal lobe (Kussmaul, 1877;
Lichtheim, 1885), and a number of subsequent
case reports have affirmed that subcortical-cortical lesions in this
localization can result in the symptom complex (Saffran
et al., 1976; Kamei et al., 1981; Metz-Lutz & Dahl, 1984; Shindo
et al., 1991; Takahashi et al., 1992;
Wang et al., 2000).
To reconcile these contrasting views, it has been suggested that there
may be at least two types of PWD (Auerbach et al.,
1982; McCarthy et al., 1990; Phillips & Farmer, 1990). One proposed form is the
sequelae of bitemporal lesions and is associated with
“prephonemic” disturbances of auditory temporal resolution. By
contrast, a second form stems from lesions of the left superior temporal
lobe and underlying white matter and is linked to impairment at phonetic
levels of processing.
In this article, we describe the results of a detailed analysis of
auditory processing in a prototypical case of PWD resulting from a
well-defined, discrete, unilateral left temporal lesion. Our findings
revealed substantial impairment in the ability to process rapid
spectrotemporal variations in speech and nonspeech sounds. However, other
aspects of auditory processing were relatively spared, even though they
required a higher degree of temporal resolution. These observations have
important implications for our conceptualization of PWD and its
subtypes.
CASE REPORT
NH (PR00001-NH)a
Human subjects policy at
the sponsoring institution now precludes identifying participants by their
actual initials. The identifier used here is a code. Future reports issued
from our institution that involve this subject will identify her by this
same code (PR00001-NH).
is a 43-year-old right-handed Caucasian
female who sustained a left temporal lesion secondary to a ruptured middle
cerebral artery aneurysm. Several months previously, she began to
experience transient visual disturbances, lasting 1 to 2 min, during which
areas of her visual field darkened. This was sometimes followed by
scintillating scotoma (seeing stars). At first, these events were isolated
and she did not seek medical attention. However, after a few months, they
were regularly followed by headache, distributed predominantly in the
basal posterior area of her head. The frequency of headache increased
until they were almost daily occurrences. On the day of her admission, she
experienced an “unbearable” headache and had to be escorted to
her family physician, where she subsequently lost consciousness in the
waiting room. She was then rushed to the emergency room of a local
university hospital. Neuroimaging studies revealed a left perisylvian
subarachnoid hemorrhage. She immediately underwent endovascular treatment
with implantation of Guglielmi Detachable Coils to seal the aneurysm. The
procedure was completed without complication, and she made a full recovery
from the operation.
As awareness returned postoperatively, NH noted that she could not
understand the dialogue in programs aired on her hospital room television,
although she could pick out words periodically. She readily oriented to
and could hear spoken communications by hospital staff but had profound
difficulty comprehending the meaning of simple verbal questions or
statements. She was transferred to an inpatient rehabilitation hospital at
3 weeks post-onset.
Assessment of speech and language during the fourth week revealed
average oromotor function. There was no facial asymmetry, and labial,
lingual, and velar movements were within normal limits. Expressive
language was fluent with good intelligibility, articulatory precision,
rate, and prosody. She produced infrequent phonemic paraphasias. Object
naming was adequate but repetition was impaired at the word level.
Understanding of basic one-step commands generally required frequent
repetition along with a decreased rate of speech. While she had briefly
demonstrated reading problems postoperatively, these had resolved at the
time of this evaluation. Narrative writing skills were also adequate.
Following a week of inpatient rehabilitation, she was discharged to a
Day Program, which she attended for 4 weeks. She continued to receive
outpatient speech-language therapy twice weekly. While she learned
compensatory strategies for her receptive problems, the auditory
processing deficits remained unchanged. The investigations described below
were completed during this time.
Premorbid History and Medical Examination
NH's history prior to the events surrounding her aneurysm bleed
was noncontributory. She had no history of seizures, significant head
injury, or neurologic disease. She had attended public schools and
generally received B's and C's in her mainstream classes. On
completing the 11th grade, she obtained her General Education Diploma and
worked as a salesperson until she started having children.
On physical examination a few months post-onset, she was found to have
nonnodular thyroid swelling but thyroid function tests were normal. Her
balance was slightly impaired. She could stand on her right foot for 12 s
and on her left for 15 s. She did not demonstrate evidence of lateralized
motor deficits, although she reported that her right hand had diminished
sensation and fatigued easily. She demonstrated a full range of motion in
her upper and lower extremities and, consistent with her subjective report
of somatosensory changes, she had mildly decreased sensation to sharps on
the right side of her body.
Audiological Evaluation
An audiological screening was completed using a standard calibrated
clinical audiometer varying in 5 dB steps. Pure-tone thresholds were
within normal limits for the left ear. Her threshold was at 10 dB at 250
Hz, 500 Hz, 750 Hz, 1 kHz, 2 kHz, 3 kHz, and 4 kHz while at 8 kHz her
threshold was 15 dB. In the right ear, sensitivity remained between 10 and
20 dB at frequencies from 250 Hz to 3 kHz. However, slightly raised
thresholds (30 dB) were evident at 4 and 8 kHz.
Structural Neuroimaging
Proton density, T1-, and T2-weighted magnetic resonance images were
obtained using a 1.5 Tesla scanner (see Fig. 1).
These scans revealed encephalomalacia and gliosis involving the dorsal
surface of the left temporal lobe extending into subadjacent white matter.
This included the planum polare, the transverse temporal (Heschl's)
gyrus, and the planum temporale. In the lateromedial plane, the
abnormalities extended from the dorsal convexity of the superior temporal
gyrus to the insula. At points, there was also minimal involvement of
adjoining intrasylvian cortex in the frontoparietal operculum. Homologous
areas of the right temporal and perisylvian region appeared intact. A
small amount of artifact was present from the implanted coil.
Magnetic resonance images showing encephalomalacia and gliosis in the
left temporal region involving the planum polare, the transverse temporal
(Heschl's) gyrus, and the planum temporale and subadjacent white
matter. Abnormalities extended in the lateromedial axis from the dorsal
surface of the superior temporal gyrus to the insula. This study was
obtained 4 months post-onset.
Neuropsychological Evaluation
Nonverbal cognitive ability, assessed with the General Assessment of
Mental Abilities (Naglieri & Bardos, 1997),
revealed a nonverbal IQ of 87, corresponding to the upper end of the low
average range. Academic abilities were commensurate with expectations
given her general level of cognitive function and educational background.
On the Wide Range Achievement Test–Third Edition (Jastak & Wilkinson, 1992), her reading/word
recognition skills fell in the low average range in comparison with other
individuals her age. On the Gates-MacGinitie Reading Tests (MacGinitie & MacGinitie, 1978), her score on the
reading comprehension subtest corresponded to a grade level equivalent of
10.9.
Assessment of speech and language utilizing the Western Aphasia
Battery (WAB) (Kertesz, 1982) revealed marked
problems with auditory language comprehension and repetition. She had
substantial difficulty understanding sequential commands (e.g.,
“point to the window and then the door”) (20/80) and in
correctly answering biographical/nonbiographical questions requiring a
basic “yes/no” response (27/60). She made only a few
mistakes in her auditory recognition of color names, real objects, body
parts, numbers, and letters (56/60), although this appeared to be
facilitated by the fact that items are identified from a small closed set
of alternatives (6) and one repetition is allowed. Consistent with other
reports of PWD (Shindo et al., 1991; Jacobs & Schneider, 2003), she also appeared to
utilize lip reading to facilitate her auditory comprehension. Her ability
to correctly repeat single words, phrases and sentences was significantly
impaired (39/100).
By contrast, expressive language was relatively spared. Speech was
fluent and grammatically correct with normal phrase length. Rare phonemic
paraphasias were noted and appeared related to periodic lapses in
self-monitoring. She obtained a perfect score (60/60) on the naming
subtest of the WAB, and word fluency was adequate (14/20). Performance
on the sentence completion (2/10) and responsive speech (2/10)
subtests was poor secondary to her comprehension deficits. Reading
comprehension (34/40) on the WAB was within normal limits. Spontaneous
writing was adequate, but she had significant difficulties writing to
dictation. Overall, she obtained an Aphasia Quotient of 69.7.
Several ancillary measures of language were also administered. On the
Token Test (DeRenzi, 1978), a measure of
auditory comprehension using nonredundant commands, she scored at the 1st
percentile in comparison with individuals her age. Her performance on
measures of speech discrimination was also impaired. On Benton's
Phoneme Discrimination Test (Benton et al.,
1983), she was required to make perceptual judgments
(same-different) on pairs of nonsense syllables/words. Half of the
pairs differed in one major phonemic feature. She correctly discriminated
only 18 of 30 items. A score less than 22 is below the lowest performance
of the normative controls and is considered “defective.”
Similarly, she performed poorly on a word discrimination task from the
Psycholinguistic Assessments of Language in Aphasia (Kay et al., 1992) requiring her to match single spoken
words to a corresponding picture in a three-alternative forced-choice
format. Whereas the mean score for control subjects is 39/40
(SD = 1.7), NH obtained a score of 33/40.
NH was administered a dichotic listening test using words (Damasio & Damasio, 1980) presented at
approximately 60 dB above threshold. During practice trials with monaural
presentation, she demonstrated significant difficulty, correctly repeating
only 30% of items. There did not appear to be a remarkable difference in
performance comparing left and right ears, although she subjectively noted
that speech presented to the right ear was more difficult to identify. On
dichotic presentation, she demonstrated comparable levels of performance.
However, right ear extinction was apparent in a complete inability to
accurately recall words presented to the right ear.
To further examine the extinction effect, NH was administered the
consonant-vowel dichotic task described by Hugdahl et al. (1991). Stimuli consisted of a set of 36 dichotic
consonant-vowel pairs produced by combining the stop
consonants—/ba/, /da/, /ga/, /ka/,
/pa/, /ta/—in all possible combinations (including
six identical pairs, e.g., /ba/-/ba/). Three different
attentional conditions included: (1) a nonforced (NF) attention condition,
in which she was asked to indicate what she heard on each trial; (2) a
forced left (FL) condition, where she was asked to recall only stimuli
presented to the left ear; and (3) a forced right (FR) condition, on which
she was instructed to focus and identify only right ear stimuli. Following
Hugdahl and Asbjornsen (Manual for Dichotic Listening
with CV-Syllables), only the first response was scored from each
trial. In the NF condition, NH demonstrated better recall from the left
ear (26.67%) compared to the right ear (16.67%). This ear asymmetry was
accentuated in the FL condition (L ear = 40%, R ear = 3.32%), but there
was little change in her overall level of performance. In the FR
condition, left ear recall was 36.67% while right ear recall was 16.67%.
When monaural consonant-vowels were presented to the right ear, her
accuracy (23.3%) was not substantially different from her best right ear
performance on the dichotic pairs. Similarly, when monaural stimuli were
presented to only the left ear, her performance (40%) remained close to
her best left-ear performance with the dichotic pairs. When compared to
age norms, she demonstrated depressed recall of information presented to
her right ear in all dichotic conditions.
NH was administered a 40-item environmental sounds recognition test
(Stefanatos & Madigan, 2000). Each item was
a 2 s segment of an environmental sound corresponding to one of four sound
categories: (1) human nonverbal (e.g., laughing, coughing); (2) man-made,
inanimate (e.g., car crash, toilet flushing); (3) nonhuman, animate (e.g.,
dog barking, cow mooing); and (4) natural, inanimate (e.g., wind, fire
crackling). After hearing each sound, she was asked to point to the
corresponding picture in a four-alternative forced-choice paradigm. Each
response card included a pictorial representation corresponding to the
target, an acoustic foil, a semantic foil, and an object that was neither
acoustically nor semantically related to the target. Acoustic distractors
produced sounds similar to the target but were from a different semantic
category. Semantic distractors were from the same semantic category as the
target but were acoustically disparate. A third distractor type was
neither semantically or acoustically confusable with the target. She
produced only four errors on this entire task, which is within normal
limits based on comparison with a small normative sample. She produced two
acoustic errors (volcano erupting → pistol firing, heavy footsteps on
wooden stairway → hammering a nail) and two semantic errors
(avalanche → tree falling, baby cooing → baby sleeping, mouth
closed). These findings suggested that her auditory processing problems
did not extend to environmental sounds. Moreover, she claimed no change in
her perception and appreciation of music, except that she could no longer
understand the lyrics.
Experimental Investigations
Speech and nonspeech perception
Three parallel tasks were devised to examine NH's perception of
speech (vowels and consonant-vowels) and nonspeech sounds (complex tones).
All stimuli were digitally synthesized (16-bit resolution at a sampling
rate of 44.1 kHz) using PRAAT 4.1 software to permit precise control over
acoustic parameters such as onset/offset, envelope shape, and
frequency characteristics. The digital waveforms were converted to analog
signals by a high-performance external USB soundcard/amplifier (Edirol
UA-5) and passed through custom-built attenuators prior to transduction by
Sennheisser HD-580 headphones. Unless otherwise indicated, tasks required
a perceptual discrimination between pairs of stimuli presented with an 800
ms interstimulus interval (ISI). She was instructed to listen to each pair
and press one button on a response pad to indicate that the two sounds
presented were the “same” or an adjacent button to indicate
they were “different.” Stimulus presentation and response
collection were controlled by using E-prime software (Psychology Software Tools, 2001). Each task included
six tokens that were contrasted in all possible combinations.
On a vowel discrimination task, NH was asked to discriminate between
pairs of the following vowels: /i/ as in beet,
as in bit,
as in bet,
as in bat,
as in cot, and /u/ as in boot. These were generated utilizing the
parameters for vowel formant frequencies outlined by Peterson and Barney
(1952). The second task, a nonspeech analogue of
the vowel discrimination task, required NH to discriminate pairs of
complex tones. Each token was comprised of four pure-tone frequency
components corresponding to the center formant frequencies of the
equivalent vowel described above. A third task assessed consonant-vowel
(CV) discrimination. The CV tokens were synthesized by pairing six stop
consonants (/b/, /d/, /g/, /p/,
/t/ and /k/) with one of the synthesized vowels,
.
The consonantal burst appropriate to each CV was sampled from natural
productions by a male speaker and inserted at the onset of each CV. All
stimuli were 250 ms in duration. Spectrograms of a representative complex
tone, vowel, and CV are provided in Figures 2A, 2B, and
2C, respectively. As can be appreciated in these illustrations,
both complex tone and vowel stimuli were characterized by steady state
spectra, whereas CV syllables contained rapidly changing spectrotemporal
cues at onset. Normal controls perform at or near ceiling on all three of
these tasks (92–100%).
These spectrograms plot the frequency spectra of the four types of
speech and nonspeech stimuli. A represents a four component
complex tone, B depicts the corresponding vowel
,
C shows the CV syllable
,
and D provides examples of the single formant stimuli. The
extent of these formant transitions was modeled after the second formant
in
.
Each contrast was presented 12 times in random order in the course of
two sessions. NH demonstrated significant difficulties in discriminating
the consonant-vowel syllables. She correctly identified 69% of the items,
which was not significantly better than chance (Binomial Test). There were
no systematic error patterns related to distinctive feature contrasts. She
demonstrated as much difficulty with contrasts of voicing as she did with
place of articulation. However, she performed better when speech sounds
differed by more than one distinctive feature (both voicing and place of
articulation).
By contrast, she was able to correctly discriminate complex tone
contrasts on 90% of trials, which is not substantially different from
performance seen in age and sex matched control subjects. An intermediate
level of performance was evident on the vowel discrimination task. She
correctly identified 82% of the contrasts correctly. She made errors on
closely related contrasts such as
and
.
This is slightly lower than performance we have seen in controls.
Decomposition of speech transitions: of a single
formant
Because formant frequency transitions are fundamental cues to
distinguishing consonants, we devised a task to assess her ability to
discriminate transitions in a single formant. By parametrically varying
the time course of transitions, we aimed to determine which durations
posed difficulty for her. The extent of the frequency transition was
modeled after the second formant transitions in the CV's
.
The up-going formant ramp started at 900 Hz and transitioned to a
steady-state frequency of 1240 Hz. The comparison down-going formant ramp
started at 1580 Hz and transitioned to the same steady-state frequency
(1240 Hz). These stimuli were paired in all possible combinations: up-up,
down-down, up-down, down-up. The duration of the formant transition was
the same for each pair. Five different durations were examined (40, 80,
120, 200, 500) with 20 trials per stimulus pair. Examples of the single
formant stimuli are depicted in Figure 2D.
NH's performance on the single formant ramp perceptual
discrimination task is depicted graphically in Figure
3. She responded with 45 to 70% accuracy on ramp durations from 40
to 200 ms. Binomial tests suggested that her discrimination accuracy was
no better than chance for all but the 500 ms ramp duration. Normal
listeners score between 93 and 100% on these discriminations, so her
performance remained somewhat poor even at the longest ramp duration.
Results from the single formant perceptual discrimination task. The
abscissa is the duration of the formant transition and the ordinate is the
percent correct.
Click Fusion
Auditory temporal resolution was assessed with a click fusion task in
which very brief, spectrally broadband stimuli (clicks) are presented in
rapid succession separated by an intervening period of silence. When this
ISI is extremely short, the temporal boundary between each click is
insufficient to result in separate percepts, so listeners
“fuse” two clicks and report hearing one. Extending the
duration of the ISI beyond a threshold value results in the perception of
two distinct clicks.
In the current implementation of this task, the duration of a silent
interval inserted between two 0.5 ms binaurally presented square wave
clicks was varied from 0 to 10 ms in 1 ms steps. Ten stimuli were
presented at each ISI. NH was asked to press one button on a response box
if one click was heard, and another button if two clicks were perceived.
Figure 4 plots her percent correct
identifications as a function of ISI.
Results from the click fusion task. The abscissa denotes the duration
of the silent ISI between 0.5 ms square wave clicks. The ordinate is the
percent correct.
NH was able to discriminate one from two “clicks” with
100% accuracy when separated by an ISI of 3 ms or more. She correctly
identified clicks with no intervening silence (0 ms) as a single click on
80% of trials. Overall, her performance compares favorably with results
from normal listeners (Patterson & Green,
1970). The findings suggest that NH was able to adequately process
brief temporal cues in sound defined by onset/offset
characteristics.
DISCUSSION
Historical Considerations
Following Wernicke's (1874) seminal
description of “sensory aphasia,” Kussmaul (1877) noted similar deficits in auditory comprehension
in a patient who did not demonstrate the coexisting problems with reading
or the copious “paraphasias” (a term he coined) that Wernicke
had noted. Kussmaul postulated that this symptom complex constituted a
distinguishable syndrome “word deafness” that resulted from
destruction of the first left temporal gyrus. He contrasted this with an
analogous disturbance in processing language in the visual modality,
“word blindness,” which he believed was secondary to left
angular gyrus and supramarginal gyrus lesions. When the two syndromes
occurred together, he suggested, they represented Wernicke's sensory
aphasia.
Lichtheim (1885) also regarded
“isolated word deafness” as a distinct clinical entity,
characterizing it in terms of selective deficits in auditory language
comprehension, repetition, and writing to dictation. He supported this
with a description of a case who demonstrated early symptoms of
Wernicke's aphasia that improved rapidly, leaving a more selective
pattern of deficits that spared volitional speech, reading, copying
written material, and spontaneous writing (p. 460). While Kussmaul had
implicated destruction of auditory cortex, Lichtheim speculated that word
deafness resulted from a deep left temporal lesion that essentially
isolated Wernicke's area from the “auditory reception
center” (primary auditory cortex). His conceptualization implied
that Wernicke's area and auditory cortex might independently be
capable of normal or near normal functioning. Subsequently, several case
studies confirmed an association of PWD with deep unilateral superior
temporal lesions (Liepmann & Storch, 1902;
Schuster & Taterka, 1926).
A number of reports also emerged suggesting that PWD could result from
bilateral temporal lesions (Pick, 1892; Déjerine & Serieux, 1898; Ballet, 1903; Barrett, 1910;
Henschen, 1919, 1920).
In contrast to the unilateral cases, these patients frequently
demonstrated persisting aphasic disturbances or collateral symptomatology
suggestive of a resolving general auditory agnosia or cortical deafness
(Buchman et al., 1986). The rarity of cases
fully meeting criteria for PWD prompted some to question the existence of
the syndrome. Pierre Marie (1906) remarked in no
uncertain terms that PWD was “a simple myth” stating “I
must declare, first of all, that to my knowledge pure word deafness does
not exist from either the clinical or the anatomical pathological point of
view … it is impossible to find an authentic case of this pretended
clinical form” (translated by Cole & Cole, 1971, pp. 77–78). Head (1926) concurred, noting that in every instance where
case reports provided sufficient detail, nonverbal perceptual impairment
existed or there was clear evidence of an aphasic disturbance apparent in
problems executing both written and oral commands. Despite persisting
concerns over definitional issues, many cases labeled as PWD were
subsequently reported with unilateral left temporal or bitemporal lesions,
which did not entirely comply with the original criteria.
The broadening concept of PWD arguably contributed to confusion
regarding its validity, its basis, and its boundaries that continues to
the present day. Both the selectivity for verbal material and the
preservation of other language functions remain focal points in
contemporary controversies (Goldstein, 1974;
Buchman et al., 1986). A current review of the
literature reveals a predominance of cases (∼72%) secondary to
bitemporal infarcts, typically involving fairly symmetric
cortico-subcortical lesions compromising middle and posterior regions of
the superior temporal gyrus (Poeppel, 2001),
some with sparing of Heschl's gyrus (Bauer &
Zawacki, 2000). Word deafness resulting from unilateral lesions is
more rarely reported, and in only a few cases has the location of the
lesion been adequately delineated (Hamanaka et al.,
1980; Kamei et al., 1981; Metz-Lutz & Dahl, 1984; Takahashi et al., 1992; Wang et
al., 2000). In all but one unilateral case, PWD has been reported
in association with left hemisphere pathology.
As a consequence of both the higher proportion of bitemporal lesions
and the general rarity of cases that adhere to strict diagnostic criteria,
prevalent views regarding the functional basis of PWD are
disproportionately influenced by studies in which word deafness was
identified as a symptom mixed with other aphasic components (Miceli, 1982; Buchman et al.,
1986; Buchtel & Stewart, 1989; Praamstra et al., 1991) or with features of a broader
auditory recognition disorder (Auerbach et al.,
1982; Vignolo, 1982). As a case in point,
the only report of PWD resulting from a unilateral right temporal lesion
(Roberts et al., 1987) is a patient who lost his
ability to recognize both spoken words and musical tunes and
arguably should not be regarded as a “pure” case of word
deafness. In reviewing the literature, Buchman et al. (1986) suggested that the modifier “pure”
be dropped but supported the concept that “word deafness” was
a distinct clinical entity.
A Prototypical Presentation of PWD
NH represents one of the rare instances where the clinical picture
conforms well to the original descriptions of PWD. Consistent with
Lichtheim's description, she exhibited early signs of a broader
aphasic disturbance resembling a Wernicke's aphasia that rapidly
resolved in the course of a couple of weeks, leaving her with a remarkably
circumscribed disturbance of auditory language comprehension. She
demonstrated moderate to severe difficulties in understanding and
repeating spoken language with otherwise intact speech production and
nonauditory language comprehension. Both reading and spontaneous writing
were preserved. In addition, there was no evidence of a broader auditory
agnosia. She demonstrated intact environmental sound recognition and
reported no change or difficulties in her appreciation of music, although
she complained that she could no longer understand the lyrics. Her
peripheral hearing sensitivity in the frequency range most critical to
speech was broadly within normal limits, although her audiogram showed
mildly raised thresholds at 4 and 8 kHz in the ear contralateral to her
lesion.
Similar audiological findings have been described in other unilateral
cases of PWD (e.g., Saffran et al., 1976)
although interaural disparities in sensitivity have also been evident at
somewhat lower (250–1000 Hz) frequencies (Takahashi et al., 1992; Wang et
al., 2000). By comparison, patients with word deafness secondary to
bitemporal lesions commonly demonstrate bilateral increases in pure-tone
thresholds (Lhermitte et al., 1971; Kanshepolsky et al., 1973; Phillips
& Farmer, 1990) that are also frequently associated with a
gradient whereby higher frequencies (≥4 kHz) are more affected (Jerger et al., 1972; Auerbach et
al., 1982; Motomura et al., 1986; Tanaka et al., 1987; Yaqub et al.,
1988; Praamstra et al., 1991; Griffiths, Rees & Green, 1999).
Analysis of NH's ability to perceive speech revealed a
substantial impairment in her ability to distinguish between stop
consonants while her perception of vowels was significantly better. This
pattern of phonemic imperception has been previously described in PWD
(Saffran et al., 1976; Auerbach et al., 1982; Miceli,
1982; Yaqub et al., 1988) but is not
specific to it. Inordinate difficulty with consonant processing can emerge
in aphasia (Miceli et al., 1978; Baker et al., 1981; Gow &
Caplan, 1996; Caramazza et al., 2000) and
problems with specific stop consonants (for example, alveolars t, d) can
be secondary to moderate hearing loss (Walden &
Montgomery, 1975). However, NH's difficulties with phonemic
perception are both more severe and pervasive than those typically
associated with aphasia or mild to moderate hearing loss. In the context
of her overall presentation, they suggest that a fundamental breakdown
exists in processes that mediate the mapping of acoustic features of
consonants onto discrete phonological representations or in the
representations themselves. Viewed from the perspective of current models
of speech perception (McClelland & Elman,
1986; Franklin, 1989; Marslen-Wilson & Warren, 1994; Frauenfelder & Floccia, 1998; Norris et al., 2000), this would impede the matching
of acoustic input with lexical entries that in turn could result in
failures to activate the retrieval of meaning attributes of words in the
semantic system.
The prelexical impairment in auditory analysis associated with
“word sound deafness” can be contrasted with “word
meaning deafness” (Bramwell, 1927; Kohn & Friedman, 1986) in which patients have
severe difficulties understanding auditory language but are able to repeat
speech and write to dictation. Preservation of the ability to repeat
speech in these cases suggests that their analysis of phonological
information is relatively intact (but see Tyler &
Moss, 1997). Rather, the disruption of language comprehension in
word meaning deafness appears to occur at the level of lexical access,
possibly secondary to a failure to match the intact phonological code with
corresponding lexical representations or to a “post access”
failure of lexical items to activate corresponding representations in the
semantic system (Ellis, 1984; Kohn & Friedman, 1986; Franklin
et al., 1996).
NH also exhibited significant perturbations in dichotic listening
performance that are consistent with previous descriptions of unilateral
cases of PWD (Albert & Bear, 1974; Saffran et al., 1976). Specifically, she demonstrated
extinction of words presented to the right ear during dichotic stimulation
and relative suppression of right ear recall when listening to dichotic CV
syllables. A strong left ear advantage on dichotic listening tasks has
also been observed in patients with aphasia and interpreted as evidence of
functional neuroplasticity and a greater role of right hemisphere
mechanisms in mediating speech processing (Moore & Papanicolaou, 1988, 1992). It is somewhat
unusual to obtain complete or near complete extinction of the right ear in
aphasia (Niccum et al., 1986), although it can
occur with lesions to Heschl's gyrus or geniculotemporal pathways
with or without aphasia.
Several factors may account for this pattern in NH. First, the extent
and distribution of her lesion likely prevents right ear input from
reaching Wernicke's area via crossed geniculocortical auditory
pathways. Right ear input may conceivably reach Wernicke's area via
transcallosal pathways, although it must compete against left ear input
arriving at right auditory cortex via stronger contralateral pathways.
Given that contralateral pathways are functionally predominant over
ipsilateral channels, this competition is biased against the right ear
input. Degradation of the signal may result from the circuitous path to
the language dominant hemisphere, and this may be further affected by
alterations in hearing competence seen contralateral to hemispheric
lesions (Linebaugh, 1978; Niccum & Speaks, 1991). Finally, it is also
possible that transcallosal pathways have been compromised by the
subcortical extension of her lesion (Gazzaniga et al.,
1973). Selective attention appeared to have little effect on this
asymmetry.
It is noteworthy that NH showed generally lower overall performance on
dichotic word tasks than has been noted in other unilateral cases of PWD.
We used an open set of words while both Saffran et al. (1976) and Albert and Bear (1974) used small (≤12) closed sets of dichotic
stimuli (monosyllabic names and digits). Because patients with PWD appear
to make use of contextual cues (Saffran et al.,
1976; Best & Howard, 1994), repeated
presentation of distinct stimuli from a closed set may have contributed to
the relatively better overall accuracy in those studies.
Word Deafness, Speech Processing, and Temporal
Imperception
The fundamental nature of the processing disturbance that
characterizes PWD has not been firmly established, nor is it clear that
the same mechanism is operative in all cases. Recent attempts to identify
the functional locus of their verbal auditory recognition disorder have
focused on detailed analyses of their difficulties with speech perception
(Denes & Semenza, 1975; Saffran et al., 1976; Auerbach et
al., 1982; Yaqub et al., 1988; Praamstra et al., 1991). A key issue concerns the
basis for the dissociation between consonant and vowel perception and
whether a lawful breakdown occurs in their processing of particular
phonetic features during consonant perception.
Vowels are characterized acoustically by a formant frequency structure
that maintains a relatively steady state for 100–150 ms. Although
not invariably the case (Tanaka et al., 1987;
Praamstra et al., 1991; Tanji
et al., 2003), the reasonably static cues for vowel perception,
such as the frequency of the first formant (F1) and the relative spacing
between F1 and F2 (Delattre et al., 1952; Ladefoged, 2001), are generally processed without
substantial difficulty in PWD, even in patients with bitemporal lesions
(Auerbach et al., 1982; Miceli, 1982; Yaqub et al.,
1988).
By contrast, the processing of consonants is impaired in both
unilateral and bitemporal cases of PWD (Denes &
Semenza, 1975; Auerbach et al., 1982;
Yaqub et al., 1988; Kazui et
al., 1990; Godefroy et al., 1995; Jacobs & Schneider, 2003). This has been observed
when tested by means of natural CV syllables (Denes
& Semenza, 1975; Saffran et al.,
1976) as well as computer-synthesized speech sounds (Saffran et al., 1976; Miceli,
1982). The perception of consonants necessitates successful online
perceptual elaboration of short-term acoustic features in the speech
signal. Cues important for the perception of stop consonants of English
(p, t, k, b, d, g) include the spectrum of brief (5 and 15 ms) noise
(consonantal) bursts (Fant, 1973), and the rate,
duration, and direction of rapid (20–50 ms) formant frequency
transitions (Delattre et al., 1952; Liberman et al., 1967; Keating
& Blumstein, 1978; Diehl, 1981). The
frequency of the burst and the trajectory (rising vs. falling) of
formant transitions give rise to the perception of different places of
articulation (labial, alveolar, or velar). There are also multiple cues to
distinguish voiced (b, d, g) from voiceless (p, t, k) consonants. A
primary cue related to voiced-voiceless distinctions is the
“voice-onset time,” which corresponds to the time lag between
the consonantal burst and the onset of voicing (the first glottal
pulse).
Some studies have suggested that patients with PWD demonstrate a
systematic breakdown involving the coding of particular phonetic features.
This is of considerable interest because it may specify the level of
phonemic processing where problems arise (Saffran et
al., 1976; Caramazza et al., 1983; Metz-Lutz & Dahl, 1984). Saffran et al. (1976) noted greater difficulty in identifying
consonants differentiated by voiced-voiceless distinctions than place of
articulation in a patient with word deafness secondary to left hemisphere
pathology. More frequently, however, the converse pattern has been
observed (Miceli, 1982; Yaqub
et al., 1988), or patients demonstrate roughly equal problems with
consonant perception whether the critical feature contrast is dependent on
place or voicing cues (Tanaka et al., 1987;
Praamstra et al., 1991). Given the relatively
small number of cases, variations in underlying pathology, and differences
in methodology, there is insufficient information at present to discern
whether systematic dissimilarities in consonant perception may exist
between patients with unilateral and bilateral lesions. However, the
observed patterns of error do not appear to be predictable on the basis of
problems at the level of phonological representation. Consequently, it has
been suggested that their difficulties with speech perception may be
related to prephonemic disturbances (Saffran et al.,
1976; Auerbach et al., 1982).
Prephonemic Disturbance in Word Deafness
The precise nature and extent of prephonemic disturbances associated
with PWD remain to be elaborated. A number of studies have suggested that
deficits in temporal resolution may be key to their difficulties in
processing speech. While normal individuals demonstrate click fusion
thresholds on the order of 1 to 3 ms (Miller &
Taylor, 1948; Patterson & Green,
1970; Hirsh, 1975), it has been reported
that patients with PWD require 15–300 ms between two clicks before
they are able to reliably make this discrimination (Albert & Bear, 1974; Auerbach
et al., 1982; Tanaka et al., 1987; Best & Howard, 1994; Godefroy
et al., 1995). Auerbach et al. (1982)
proposed that this elevated threshold reflected impaired auditory temporal
acuity, which in turn could cause special difficulties with consonant
perception by impeding the analysis of rapid formant frequency
transitions.
The link between impairment of auditory temporal resolution and
problems with specific aspects of speech perception or language
comprehension in PWD has remained speculative. Indirect evidence
supporting an association may be garnered from a study by Godefroy et al.
(1995) who followed a patient with
“auditory agnosia” in the course of his recovery. They found
that click fusion thresholds assessed in the acute stage were
significantly elevated. However, as the word deafness resolved, there was
a corresponding normalization of click fusion thresholds. The parallel
recovery of word deafness and click fusion strengthens the argument for a
common underlying disturbance, although clearly it is not possible to
suggest causal relations from such correlations.
The precise nature of the temporal processing deficits indexed by
faulty click fusion also remains at issue. Kanshepolsky et al. (1973) found that as the duration of auditory stimuli
became very short, sound intensity had to be raised appreciably (20 to 25
dB) in order for their patient with word deafness to hear the stimulus. A
few studies have shown that these problems tend to co-occur in patients
with word deafness secondary to bitemporal infarction, particularly with
lesions involving or extending into subcortical white matter (Motomura et al., 1986; Tanaka et
al., 1987). However, Buchtel and Stewart (1989) suggest that poor temporal resolution is evident
in PWD in the absence of difficulties perceiving brief stimuli in
isolation. Noting that their patient with PWD was completely unable to
perceive clicks due to their brevity, they utilized a modified fusion
task, replacing clicks with 30 ms tone bursts that their patient had no
difficulty hearing. Manipulating the duration of the silent ISI between
these tone bursts, they found that their patient demonstrated profound
difficulties in discriminating one from two tones presented in rapid
succession. Whereas normal subjects reliably differentiated two tones when
separated by silent intervals of approximately 15 ms, their patient
required a silent gap of approximately 250 ms.
Based on observations of a frequent co-occurrence of abnormal
intensity-duration functions and elevated fusion thresholds, Tanaka et al.
(1987) speculated on a common neurophysiological
basis for these diverse disturbances of timing. They suggested that such
problems might be explicable in terms of abnormal persistence of neural
activity, prolonged refractory periods, or slow initial recruitment. Some
studies have observed that timing anomalies are also evident in PWD in the
perception of rapid visual events (Best & Howard,
1994; Tanaka et al., 1987) raising the
possibility of a supramodal timing deficit. Extending a conceptualization
by Auerbach et al. (1982), Best and Howard
(1994) suggested that a slow or inconsistent
neural clock may the most parsimonious explanation for the diverse
temporal processing deficits associated with PWD. They conceived of this
biological timing mechanism in terms of a reciprocal loop from the cortex
to the cerebellum and suggested that it may serve as a central timing
mechanism for several neural systems.
Temporal Processing in Unilateral Cases
A problem with the slow clock hypothesis and other accounts positing
that PWD is based in fundamental disturbances of temporal resolution is
that the supporting data are derived almost entirely from patients with
bilateral lesions. Only one study has examined click fusion in a
unilateral case of PWD, and while the findings were abnormal (Albert & Bear, 1974), the observed threshold (15
ms) was substantially lower than that described in several cases of word
deafness secondary to bitemporal lesions (30 to 300 ms) (Auerbach et al., 1982; Motomura et
al., 1986; Tanaka et al., 1987).
Evidence from NH suggests that temporal processing deficits can exist
in PWD in the absence of basic deficiencies in resolving distinct auditory
events. NH demonstrated a normal click fusion threshold, yet had profound
difficulties with the spectrotemporal analysis of transient modulations in
formant frequency occurring within a temporal window of up to 200 ms.
Because short-term dynamic patterns such as frequency modulations (FM) are
ubiquitous to speech and serve as fundamental cues to the phonemic
identification of consonants, the observed deficits would pose significant
obstacles in understanding spoken language. By contrast, she appeared able
to process slower modulations. Although not specifically examined, this
may support satisfactory processing of suprasegmental aspects of speech
analysis such as prosody and intonation contours, consistent with a case
described by Coslett et al. (1984).
Deficiencies in FM analysis of tonal stimuli have also been described
by Wang et al. (2000) in a PWD patient with a
unilateral left hemisphere lesion involving cortical and subcortical white
matter of left temporal lobe extending superiorly into frontoparietal
regions. Their patient had difficulty in discriminating the directional
trajectory (up-going vs. down-going) of pure-tone glides 302 ms
in duration as well as 50 ms linear frequency ramps followed by a
steady-state tone. These observations are broadly consistent with our
findings with formant frequency modulations. Wang et al. (2000) did not utilize a measure of temporal acuity, so
it cannot be discerned whether the deficits observed in their case were
also independent of more basic problems with temporal resolution.
Neurophysiological and Neuroanatomical
Considerations
Overall, our findings provide compelling evidence that prephonemic
auditory temporal processing disturbances can occur in PWD secondary to
unilateral left temporal lobe lesions and result in substantial problems
with auditory comprehension. The dissociation between click fusion and
frequency modulation analysis observed in NH cannot readily be
accommodated by the concept of a slow clock as suggested by Best and
Howard (1994). Rather, the results appear to
implicate specialized mechanisms that mediate rapid FM analysis. Evidence
from animal neurophysiological studies and human psychophysical
experiments suggest that temporally varying sounds undergo specialized
analysis involving modulation sensitive neural mechanisms that are
distinct from processes underlying the analysis of steady-state sounds
(Kay, 1974). These mechanisms or
“channels” are not merely concerned with the detection of a
change from one frequency to another but are sensitive to the
instantaneous temporal properties of frequency change such as the rate,
shape, direction, and periodicity of modulation (Kay
& Matthews, 1972; Green & Kay,
1973; Collins & Cullen, 1978; Gardner & Wilson, 1979; Regan
& Tansley, 1979). They are physiologically distinguishable from
analogous mechanisms that process amplitude modulations in sound (Regan & Tansley, 1979; Kay,
1982). The temporal tuning that characterizes the response
properties of FM sensitive neurons, their connectivity with other auditory
neurons, and their hierarchical organization in the auditory system
suggests an intrinsic capacity to track modulations in time.
Frequency modulation sensitive mechanisms have functional
characteristics critical to speech reception (Kay,
1982). Studies of the cortical organization of these mechanisms
suggest that they are elaborated in the superior temporal cortex in areas
confluent with primary auditory cortex and classical language reception
areas (Arlinger et al., 1982; Hari & Makela, 1986; Makela et
al., 1987). Recent functional magnetic resonance imaging studies
have localized FM sensitive mechanisms to so-called belt and parabelt
association cortex anterolateral and lateral to Heschl's gyrus (Johnsrude et al., 1997; Binder et
al., 2000; Zatorre et al., 2002) in areas
that appear to be specialized for processing rapid time varying aspects of
sound (Hart et al., 2003). Joanisse and Gati
(2003) observed that speech and dynamically
varying nonspeech stimuli produce remarkable overlap in neural activation
on functional magnetic resonance imaging. Indeed, they suggest that
differences appeared to be related to the degree of activation rather than
differences in spatial localization.
Interestingly, these areas have been implicated in cortical mapping
studies as critical to consonant perception. Examining patients undergoing
cortical mapping in preparation for surgical treatment of intractable
epilepsy, Boatman et al. (1995) demonstrated
that electrical stimulation to lateral mid and posterior superior temporal
cortex disrupted the perception of consonant-vowel syllables but not
vowels. These regions may form a functional subunit of a posterior stream
of auditory processing distributed along the supratemporal cortical plane
(Wise et al., 2001).
While there is evidence of bilateral mediation of the speech code,
there is growing support from dichotic listening, electrophysiological,
neuromagnetic, and functional neuroimaging studies to suggest that there
are hemispheric differences in the computational networks that specialize
in the analysis of rapid acoustic modulations in speech. Poeppel (2003) has proposed that the neural architecture that
analyzes speech signals is asymmetrically analyzed in the time domain,
with left-hemisphere mechanisms preferentially extracting information over
shorter (25–50 ms) temporal integration windows while right
hemisphere mechanisms integrate temporal variations over longer
(150–250 ms) windows. Our findings are broadly consistent with this
viewpoint. However, the dissociation between click fusion and frequency
modulation analysis suggests that the temporal window concept is in need
of a higher degree of specification. Specifically, our results suggest
that window size may depend on the type of information that is being
analyzed in the time domain. Differentiation of auditory objects based on
information present in amplitude modulations (on and off) was not affected
in our patient with PWD. We would therefore suggest that hemispheric
asymmetries are more related to the analysis of rapid spectrotemporal
variations that require the tracking of relational structure or temporal
patterns in sound.