Introduction
The Late Pennsylvanian entomofauna of the Souss basin was discovered in 2001 during paleontological fieldwork in lacustrine black shales at Tanamert village in the western High Atlas mountains of Morocco (Fig. 1.1, 1.2). It is so far the only known late Paleozoic entomofauna of North Africa and, judging from the composition of the plant and insect assemblages, the southernmost Euramerican one (Hmich et al., Reference Hmich, Schneider, Saber and El Wartiti2003, Reference Hmich, Schneider, Saber, El Wartiti, Lucas and Zeigler2005, Reference Hmich, Schneider, Saber, Voigt, El Wartiti, Lucas, Cassinis and Schneider2006). In this context, the Souss insect localities are of considerable interest compared to the well-known European and North American insect sites, such as Mazon Creek (North America; Selden and Nudds, Reference Selden and Nudds2012), Nord Pas-de-Calais and Commentry (France; e.g., Brongniart, Reference Brongniart1893; Pruvost, Reference Pruvost1919; Carpenter, Reference Carpenter1943; Schneider, Reference Schneider1983b; Béthoux and Nel, Reference Béthoux and Nel2003), and Wettin (Germany; Schlechtendal, Reference Schlechtendal1912; Schneider, Reference Schneider1978b). These classical insect collections from roof shales and interbeds of coal seams were sampled on mine dumps and reflect the autochthonous to parautochthonous entomofauna of densely vegetated swamps and mires of the paleoequatorial belt (DiMichele, Reference DiMichele2014). By contrast, the Souss insect localities are situated in different paleogeographical positions within the Souss basin (Fig. 1.1), and the individual insect beds are scattered across a nearly 900 m thick part of the section (Figs. 2, 3) representing different sedimentary and biotic environments from swamps and mires to shallow and deep lakes within fluvial-dominated landscapes. In addition, the insect beds cover a time span of ~1.5 Myr, roughly estimated from sedimentation rates of fluvial sandstones and lacustrine silt- and claystones in the Souss basin. This provides the opportunity to investigate the evolution of single taxa as well as the development of faunal compositions related to changing environmental conditions through time and space.

Figure 1. Location and geological overview of the study area in Morocco. (1, 2) Position of the working area in the western High Atlas mountain region in Morocco and the main fault systems. (3) Simplified geological map of the Souss basin with the tectonically bordered Ida Ou Ziki subbasin in the North and the Ida Ou Zal subbasin in the South. Numbered stars indicate the insect fossil localities discussed in the text (after Saber, Reference Saber1998; Saber et al., Reference Saber, El Wartiti and Broutin2001, Reference Saber, El Wartiti, Hmich and Schneider2007).

Figure 2. Profile of the Ida Ou Zal subbasin (after Saber, Reference Saber1994; Saber et al., Reference Saber, El Wartiti and Broutin2001, Reference Saber, El Wartiti, Hmich and Schneider2007). Hercynian folded basement covered by the conglomeratic Ikhourba Formation, followed by the braidplain sandstone/siltstone deposits of the Stephanian A/B (Kasimovian) El Menizla Formation. Indicated are the positions of the insect localities (cf. Table 1). The Stephanian is overlain with an angular and erosive unconformity by middle to late Permian red beds of the Ikakern Formation.

Figure 3. Profile of the Ida Ou Ziki subbasin (after Saber, Reference Saber1994; Saber et al., Reference Saber, El Wartiti and Broutin2001, Reference Saber, El Wartiti, Hmich and Schneider2007). Hercynian folded basement covered by the conglomeratic Tajgaline Formation, followed by the braidplain sandstone/siltstone deposits of the Stephanian A/B (Kasimovian) Oued Issene Formation. Indicated are the positions of the insect localities (cf. Table 1). The Oued Issene Formation is erosively overlain by fluvial conglomerates of the ?late Stephanian Tirkou Formation. It is followed by an angular and erosive unconformity by middle to late Permian red beds of the Ikakern Formation. See Figure 2 for legend.
Table 1. Locations and specimens of Phyloblattidae and Compsoblattidae of the Stephanian Souss basin in Morocco.

Each of the several insect beds in the Souss basin has been documented and sampled in great detail with regard to the sedimentology (= depositional environment) and to the whole floral and faunal content (= composition of biota, paleoecology). Altogether, these data provide insights into environmental factors determining the composition and evolution of insect associations. The study, from fieldwork to taxonomy, of the individual insect taphocoenoses of the Souss basin is still in progress. The paper presented here is one in a series of forthcoming publications on the taxonomy, paleoecology, and evolution of the Souss entomofauna in a global context.
Geological setting
The Late Pennsylvanian (Kasimovian, early Stephanian) sub- to perimontaneous Souss basin is situated in the present-day southwestern High Atlas mountains of Morocco (Fig. 1.1, 1.2). It consists of the two tectonically separated subbasins of Ida Ou Zal and Ida Ou Ziki, which formed originally as a single basin and ultimately separated into two subbasins after the early Stephanian (Kasimovian) and before the late middle Permian (Capitanian?) by a sinistral movement of the west branch of the Tizi N'Test Fault (TZTF; Fig. 1.3) at the end of the Mauretanid phase of the Variscan (Hercynian) Orogeny in Morocco (Saber, Reference Saber1994; Saber and El Wartiti, Reference Saber and El Wartiti1996; Saber et al., Reference Saber, El Wartiti, Hmich and Schneider2007). The Late Pennsylvanian deposits form a maximum thickness of 1,800 m resting directly on the Variscan deformed and metamorphosed basement of the Souss basin (Saber et al., Reference Saber, El Wartiti and Broutin2001). The basin fill from the base to the top (Figs. 2, 3) comprises basal conglomerates of the Ikhourba Formation in the Ida Ou Zal subbasin and the Tajgaline Formation in the Ida Ou Ziki subbasin followed by more than 1,000 m of gray sediments comprising fluvial sandstones, lacustrine black shales, and, in places, up to decimeter thick coal seams of the El Menizla Formation in the Ida Ou Zal subbasin and the Oued Issene Formation in the Ida Ou Ziki subbasin (Figs. 4–6) (Saber, Reference Saber1994; Saber et al., Reference Saber, El Wartiti and Broutin2001, Reference Saber, El Wartiti, Hmich and Schneider2007; Lagnaoui et al., Reference Lagnaoui, Voigt, Belahmira, Saber, Klein, Hminna and Schneider2018). The Oued Issene Formation is erosively overlain by the coarse clastics of the ?late Stephanian Tirkou Formation. In both subbasins, these Carboniferous units are overlain with an angular and erosive unconformity by middle to late Permian red beds of the Ikakern Formation (T1 and T2; Voigt et al., Reference Voigt, Hminna, Saber, Schneider and Klein2010; Hminna et al., Reference Hminna, Voigt, Saber, Schneider and Hmich2012) and in places by Early to Late Triassic Timesgadiouine Formation (T3 to T5; Klein et al., Reference Klein, Voigt, Hminna, Saber, Schneider and Hmich2010, Reference Klein, Voigt, Saber, Schneider, Hminna, Fischer, Lagnaoui and Brosig2011; Voigt et al., Reference Voigt, Schneider, Saber, Hminna, Lagnaoui, Klein, Brosig and Fischer2011; Lagnaoui et al., Reference Lagnaoui, Klein, Voigt, Hminna, Saber, Schneider and Werneburg2012).

Figure 4. Outcrop situation of the El Menizla Formation (Stephanian A/B, Kasimovian) at the mountain road from El Menizla to Agadir Ou Anzizen in the Ida Ou Zal subbasin. Shown is a part of the monotonous sandstone/siltstone/black shale interbeddings as they are typically for the braidplain deposits of the El Menizla and Oued Issene formations. In the level of the road (at scale bar) laminated lacustrine black shales of the insect locality CDUE 79-AGXXVI crop out. Scale bar = 2 m.

Figure 5. Outcrop situation in the lower part of the Oued Issene Formation, Stephanian A/B (Kasimovian) of the Ida Ou Ziki subbasin in the Oued Issene canyon downstream of the village Tanamert. The insect beds of the CDUE locality 80-TaV are situated in dark gray, bivalve-rich, and plant-containing littoral lacustrine (foreground right) and in profundal lacustrine black shales with generally rare bivalves but common freshwater medusas (middle ground). In the background are dominating fluvial sandstones, in places with tetrapod tracks and conifer remains, interbedded with lacustrine pelites. Man for scale.

Figure 6. Outcrop situation of CDUE locality 82-TaI at Tanamert village in the Oued Issene canyon, Ida Ou Ziki subbasin. Shown is the inclined uppermost part of the Stephanian A/B (Kasimovian) Oued Issene Formation (Is) conformable but erosively overlain by the conglomerates of the Tirkou Formation (Tir) of assumed Stephanian age. It follows red beds of the middle to late Permian Ikakern Formation (T2) and the basal conglomerate, the Aid Driss Member, of the Triassic Timezgadiouine Formation (T3), both with an angular erosive unconformity at the base. The insect bed TaI of the CDUE locality 82 is situated in a sequence of mainly littoral dark gray siltstones and subordinately profundal dark black claystones.
The age of the insect-bearing beds was determined by insect biostratigraphy (Hmich et al., Reference Hmich, Schneider, Saber, El Wartiti, Lucas and Zeigler2005, Reference Hmich, Schneider, Saber, Voigt, El Wartiti, Lucas, Cassinis and Schneider2006) based on morphophylogenetic spiloblattinid lineages (Schneider and Werneburg, Reference Schneider, Werneburg, Lucas, Schneider and Cassinis2006, Reference Schneider and Werneburg2012) as Stephanian A/B of the West European Regional Scale. Co-occurrences of spiloblattinid zone species together with marine guide fossils such as conodonts and fusulinids in mixed continental-marine deposits of North America allow for direct correlations with the Late Pennsylvanian Kasimovian of the marine Standard Global Chronostratigraphic Scale (Lucas et al., Reference Lucas, Barrick, Krainer and Schneider2013; Schneider et al., Reference Schneider, Lucas and Barrick2013).
Materials and methods
The taxonomic analysis of the blattoids here described is based on 56 specimens from the Souss entomofauna in comparison with the well-investigated European Commentry and Wettin entomofauna and data from North America. The fossil insects analyzed herein were collected during fieldwork in 2001, 2004, and 2012–2018 in outcrops along the mountain road from the El Menizla to the Agadir Ou Anzizen villages (El Menizla Formation) and in the Oued Issene canyon south of the Abdelmoumen dam, close to Tanamert village (Oued Issene Formation) (Figs. 1, 4–6; Table 1).
Photographs were taken with a Canon EOS 500D camera equipped with macro lenses Canon EF-S 2.6–60 mm and Canon MP-E 65mm as well as with a KEYENCE VHX 5000 digital microscope. Photographs show the specimens in their natural position; nevertheless, the drawings are presented as left wings (mirrored if they are right wings) to facilitate better comparison. Drawings were prepared using a series of photographs obtained with different illumination settings and by direct observation of specimens under a stereomicroscope, either dry or wetted with ethanol. Draft drawings were scanned and edited with graphic software. Retrodeformation of tectonically deformed wings, based on the outline of undeformed wings, was performed manually and using graphic software CorelDraw X4. Consequently, the sizes in the drawings may differ from those of the corresponding photographs. Notice that in the figure captions the preserved size of the wings or wing fragments is given.
Various contributions were used as references for species delimitations. For the revision of late Paleozoic blattoid families and genera, see Schneider (Reference Schneider1983a); for the revision of the phyloblattid taxa and the relevant literature, see also Schneider (Reference Schneider1978b, Reference Schneider1983b) and Schneider et al. (Reference Schneider, Lucas, Rowland, Lucas and Zeigler2004). These contributions diverted from those by earlier workers, including Anton Handlirsch, whose principle was that each specimen should be described provisionally as a separate species. Later, with increasing knowledge, revisions would unify specimens to species (Handlirsch, Reference Handlirsch1937, p. 65). Consequently, if any new species of any author was based, besides the holotype, on additional specimens, all these specimens were named separately as new species by Handlirsch (e.g., Reference Handlirsch1906a, Reference Handlirschb, Reference Handlirsch1922). Unfortunately, researchers who did not read German misunderstood Handlirsch´s principle as a valid concept of α-taxonomy up to present times. The consequence is an oversplitting of species and of higher taxonomic categories, too. In a revision of the Late Pennsylvanian and early Permian blattoids of the Wettin entomofauna of the Saale basin as well as from some other German localities, the number of species was reduced from formerly about 150 to 35, a decrease of about 80% (Schneider, Reference Schneider1978b). An indispensable basis for this revision was the detailed investigation and documentation of the individual, intraspecific, and interspecific variability of 99 forewing pairs of males and females of the extant Periplaneta americana (Linnaeus, Reference Linnaeus1758) and 47 forewing pairs of males and females of P. australasiae (Fabricius, Reference Fabricius1775) (Schneider, Reference Schneider1977, Reference Schneider1978a). With this experience in mind, a worldwide revision of (nearly) all known late Paleozoic blattoids was carried out by Schneider (Reference Schneider1983a). As a result, the number of species was reduced by, again, roughly 80%, if the last complete documentation by Handlirsch (Reference Handlirsch1922) of all fossil insects worldwide known at this time is used as a database. Handlirsch listed in this compilation ~800 late Paleozoic blattoid species belonging to about 125 genera and 13 families. Up to the eighties of the past century, this number had increased to an estimated 900 species. Only ~190 species belonging to 21 genera and eight families remain after revision (Schneider, Reference Schneider1983a; Jarzembowski and Schneider, Reference Jarzembowski and Schneider2007).
Species formerly described from various localities and strata will be used for comparison in the systematic section. The corresponding occurrences are Havercampf farm, SE of Lawrence, Lawrence Shale, Washington Formation, lower Douglas Group, Mid-Continent Basin, Kansas, Kasimovian–Gzhelian transition, in the following shortened to Lawrence; Grande Couche, Commentry basin, French Massif Central, Stephanian B/C, early Gzhelian, shortened to Commentry; Wettin Subformation, Siebigerode Formation, Saale basin, Germany, Stephanian C, Gzhelian, shortened to Wettin. The herein studied material of the Souss entomofauna is shortened to Souss. The stratigraphic designations follow Lucas et al. (Reference Lucas, Allen, Krainer, Barrick, Vachard, Schneider, DiMichele and Bashforth2011), Schneider and Werneburg (Reference Schneider and Werneburg2012), and Schneider and Scholze (Reference Schneider, Scholze, Lucas and Shen2016).
Repositories and institutional abbreviations
The studied material is housed at the Chouaïb Doukkali University El Jadida, Morocco, abbreviated as CDUE and supplemented by the locality (e.g., TaV), bed (e.g., 13d), and specimen number (e.g., 54). Discussed type specimens are stored at Martin-Luther-University Halle-Wittenberg, Institute of Geosciences and Geography, Germany, abbreviated as MH; the Smithsonian Institution, National Museum of Natural History, Washington, DC, USA, abbreviated as USNM; and Muséum National d'Histoire Naturelle, Paris, France, abbreviated as MNHN.
Systematic paleontology
The wing venation nomenclature follows the serial insect wing venation pattern paradigm of Lameere (Reference Lameere1922). Wing venation abbreviations relevant for this contribution are as follows: Sc = subcosta; R = radius; RA = radius anterior; RP = radius posterior; M = media; MA = media anterior; MP = media posterior; CuA = cubitus anterior; CuP = cubitus posterior. The nomenclature of the anal field veins generally follows Schneider (Reference Schneider1984a), except that instead of An1 and An2, the designations AnA and AnP are used here; AnP1 = first posterior analis, and so on.
For the crossvenation (archedictyon), the following descriptive terms are used: reticulate = a regular polygonal meshwork of fine veinlets (Fig. 7.1, 7.2); anastomosing-striate = an irregular meshwork of striate erratic joined fine veinlets (Fig. 7.3); crossvein seams = densely arranged crossvein stubs along main veins and their branches (Fig. 7.4).

Figure 7. Crossvenations of phyloblattids and of Compsoblatta Schlechtendal in Handlirsch, Reference Handlirsch1906b. (1) Finely reticulate crossvenation of Phyloblatta occidentalis (Sellards, Reference Sellards1908) forewing, CDUE TaV-3-X1-4 (Fig. 8.7); (2) typical transitional anastomosing-striate to reticulate crossvenation in the anal field and the CuP area of forewings in the Anthracoblattina ensifera-gigantea group, characteristic is the clear differentiation between AnA and AnP, CDUE TaI-13e-2 (Fig. 13.1); (3) typical fine reticulate crossvenation in the middle basal part of forewings in the Anthracoblattina ensifera-gigantea group, notice sporadically occurring small veinlets (arrows) between main veins in the basal wing part, CDUE TaI-13d-48 (Fig. 13.3); (4) typical irregularly reticulated crossvenation in the upper basal part of forewings of Compsoblatta between the main veins and their branches the crossvenation is partially reduced to seams; areas framed by these seams are paler than the remaining wing surface, CDUE TaI-13d-34 (Fig. 15.1). Scale bars = 2.5 mm. Sc = subcosta; R = radius; M = media; CuA = cubitus anterior; CuP = cubitus posterior; AnA = analis anterior ; AnP = analis posterior.
In the description, distinction is made between vein forking (symmetrical, Y-form) and vein branching (asymmetrical). Despite that, in both cases the resulting veins are called branches or twigs. ‘Proximal’ or ‘basal’ is related to the wing base, ‘distal’ or ‘apical’ to the wing apex.
Because of the generally strong compaction, the natural relief of the wings is mostly not preserved. Therefore, in most cases it is not possible to decide whether it is a left or a right wing. Only when this decision is unambiguously possible do we give corresponding information in the descriptions.
Class Insecta Linnaeus, Reference Linnaeus1758
Superorder Dictyoptera Latreille, Reference Latreille1829
Order Blattodea Brunner von Wattenwyll, Reference Brunner von Wattenwyl1882
Family Phyloblattidae Schneider, Reference Schneider1983a
Genus Phyloblatta Handlirsch, Reference Handlirsch1906a
Type species
Phyloblatta schroeteri (Giebel, Reference Giebel1867); Wettin; MH 32, by original designation.
Diagnosis
Elongate ellipsoidal forewings of about 10–35 mm in length. Costal field strip-like, up to about 70% of forewing length. Sc pectinate; branches end at the anterior wing margin. R sigmoidal; R branches terminate at anterior wing margin. M sigmoidal, multiple-forked, branches covering an area extending from the wing tip to the transition between wing tip and posterior wing margin. CuA sigmoidal. First CuA twigs arise by branching from CuA stem and are rarely forked; all further branches arise by furcation. CuP regularly bended. Crossvenation (archedictyon) reticulate to anastomosing striate.
Occurrence
Pennsylvanian to Permian, Europe, North America, Siberia, rare in China, South America, and South Africa (Schneider, Reference Schneider1983a); Pennsylvanian, North Africa (herein).
Phyloblatta occidentalis (Scudder, Reference Scudder1890)
Figures 7.1, 8–12
- Reference Scudder1890
Etoblattina occidentalis Scudder, p. 410, pl. 32, fig. 4.
- Reference Handlirsch1906a
?Phyloblatta occidentalis; Handlirsch, p. 739.
- Reference Handlirsch1906b
Phyloblatta occidentalis; Handlirsch, p. 205, pl. 21, fig. 16.
- Reference Sellards1908
Etoblattina obscura Sellards, p. 509, pl. 70, fig. 5; pl. 79, figs. 1, 2; pl. 81, figs. 2, 4; pl. 83, figs. 1, 2.
- Reference Sellards1908
Etoblattina occidentalis; Sellards, p. 512, pl. 70, figs. 1, 2; pl. 78, fig. 3; pl. 79, fig. 4.
- Reference Sellards1908
Etoblattina fulva Sellards, p. 512, pl. 70, figs. 4, 6, 9; pl. 79, fig. 3; pl. 81, figs. 3, 6.
- Reference Handlirsch1920
Phyloblatta obscura; Handlirsch, p. 443.
- Reference Handlirsch1920
Phyloblatta fusca Handlirsch, p. 443.
- Reference Handlirsch1920
Phyloblatta lugubris Handlirsch, p. 443.
- Reference Handlirsch1920
Phyloblatta magna Handlirsch, p. 443.
- Reference Handlirsch1920
Phyloblatta fulva; Handlirsch, p. 442.
- Reference Handlirsch1920
Phyloblatta fulvana Handlirsch, p. 442.
- Reference Handlirsch1920
Phyloblatta fulvella Handlirsch, p. 442.
- Reference Meunier1921
Etoblattina parva Meunier, p. 93, fig. 126, pl. 18, fig. 9.
- Reference Meunier1921
Etoblattina queribunda Meunier, p. 93, fig. 127, pl. 18, fig. 10.
- Reference Meunier1921
Etoblattina flebilis Meunier, p. 94, fig.128a, b, pl. 18, fig. 11.
- Reference Meunier1921
Etoblattina subtilis Meunier, p. 97, fig. 133, pl. 19, fig. 3.
- Reference Meunier1921
Etoblattina vulpis (partim) Meunier, p. 95, fig. 130, pl. 18, fig. 13, pl. 20, fig. 9.
- Reference Handlirsch1937
Phyloblatta parva; Handlirsch, p. 77.
- Reference Handlirsch1937
Phyloblatta queribunda; Handlirsch, p. 77.
- Reference Handlirsch1937
?Phyloblatta flebilis; Handlirsch, p. 77.
- Reference Handlirsch1937
Phyloblatta subtilis; Handlirsch, p. 77.
- Reference Schneider1983b
Phyloblatta parva; Schneider, p. 78, pl. 1, figs. 1–4, pl. 5, fig. 3.

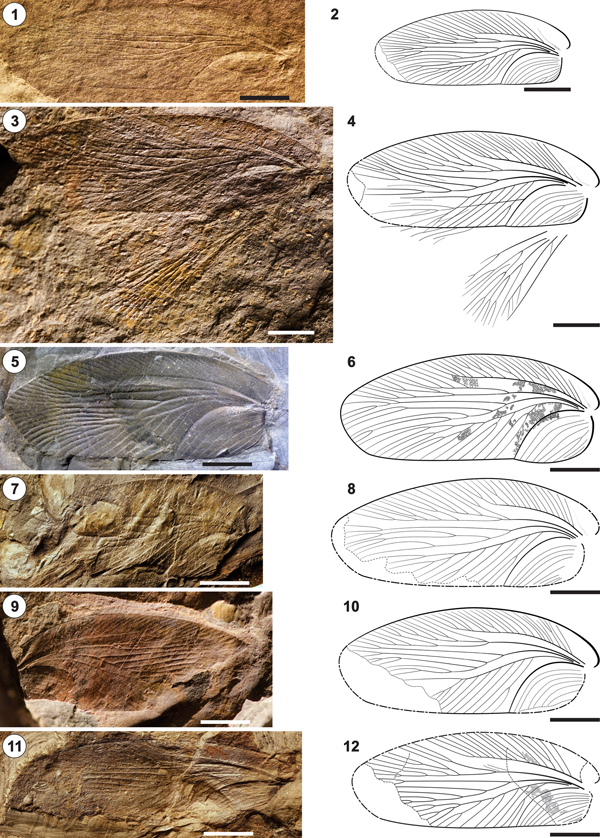
Figure 8. Phyloblatta occidentalis from CDUE localities 80-TaV and 82-TaI near Tanamert village, Oued Issene Canyon, Ida Ou Ziki subbasin, Oued Issene Formation, Stephanian A/B (Kasimovian). Drawings show specimens partially retrodeformed and mirrored to facilitate comparison. Size data refer to the preserved sizes of the specimens. (1, 2) CDUE TaV-3-4a,b (31 mm × 9 mm); (3, 4) CDUE TaV-3-X0-14 (25 mm × 12 mm); (5, 6) CDUE TaI-13d-50 (32 mm × 9 mm); (7, 8) CDUE TaV-3-X1-4 (29 mm × 9 mm); (9, 10) CDUE TaV-3-10a,b (24 mm × 8.5 mm); (11, 12) CDUE TaV-3-49 (25 mm × 9 mm); (13, 14) CDUE TaI-13d-26 (29 mm × 8.5 mm); (15, 16) CDUE TaV-3-17 (21 mm × 7 mm). Scale bars = 5 mm.

Figure 9. Phyloblatta occidentalis from CDUE localities 80-TaV and 82-TaI near Tanamert village, Oued Issene Canyon, Ida Ou Ziki subbasin, Oued Issene Formation, Stephanian A/B (Kasimovian). Drawings show specimens partially retrodeformed and mirrored to facilitate comparison. Size data refer to the preserved sizes of the specimens. (1, 2) CDUE TaV-3-X1-25 (30 mm × 8 mm); (3, 4) CDUE TaI-13d-61a (28 mm × 9 mm); (5, 6) CDUE TaV-3-19 (26 mm × 10 mm); (7, 8) CDUE TaI-13d-54 (26 mm × 11 mm); (9, 10) CDUE TaI-3-47a,b (25 mm × 11 mm); (11, 12) CDUE TaI-13e-1 (235 mm × 8 mm). Scale bars = 5 mm.

Figure 10. Phyloblatta occidentalis from CDUE locality 80-TaV near Tanamert village, Oued Issene Canyon, Ida Ou Ziki subbasin, Oued Issene Formation, and from CDUE locality 77-A5 at the mountain road from El Menizla to Agadir Ou Anzizen, Ida Ou Zal subbasin, El Menizla Formation, Stephanian A/B (Kasimovian). Drawings show specimens partially retrodeformed and mirrored to facilitate comparison. Size data refer to the preserved sizes of the specimens. (1, 2) CDUE TaV-3-X1-22 (21 mm × 10 mm); (3, 4) CDUE A5/13 (21 mm × 10 mm); (5, 6) CDUE A5/14 (20 mm × 13 mm). Scale bars = 5 mm.

Figure 11. Phyloblatta occidentalis from CDUE locality 80-TaV near Tanamert village, Oued Issene Canyon, Ida Ou Ziki subbasin, Oued Issene Formation, and from CDUE locality 79-AGXXIV at the mountain road from El Menizla to Agadir Ou Anzizen, Ida Ou Zal subbasin, El Menizla Formation, Stephanian A/B (Kasimovian). Size data refer to the preserved sizes of the specimens. (1, 2) CDUE TaV-3-X1-30 (length of wing fragment 17 mm); (3, 4) CDUE TaV-3-X1-27 (wing 22.5 mm × 10 mm); (5, 6) CDUE AGXXIV-17 (left wing 28 mm × 11 mm). an = antenna; ar = arolium; cx = coxa; fe = femur; hc = head capsule; ma = ?mandible; ps = pronotal shield; ta = tarsus; ti = tibia. Scale bars = 5 mm.

Figure 12. Reconstruction of Phyloblatta occidentalis based on specimens CDUETaV-3-X1-30, TaV-3-XI-27, and AGXXIV-17 (Fig 11.1–11.6). The forewings are from CDUE TaV-3-4a,b (Fig 8.2). The position of the head capsule in front of the pronotal shield is possibly taphonomically caused. Scale bar = 5 mm.
Holotype
Etoblattina occidentalis Scudder, Reference Scudder1890; Lawrence; USNM 38071 (part and counterpart).
Diagnosis
As for the genus with the following specifics. Elongate ellipsoid forewings of about 20–35 mm in length. Costal field about 50%–70% of forewing length. Sc pectinate, with 10–23 branches terminating at the wing margin. R weakly sigmoidal, terminating just anterior of the wing tip; first or second R branch once or multiple forked; altogether 10–15 R branches run against the anterior wing margin. M sigmoidal, divided at about one-third to half wing length in two parts; altogether with eight to 11 branches covering an area extending from just anterior of the wing tip down to the transition of the wing tip into the posterior wing margin. CuA gently sigmoidal, first two to six posteriorly pectinate, rarely forked twigs arise by branching from CuA stem, all further veins arise by multiple furcation. CuA sends altogether 10–16 branches to the posterior wing margin. CuP regularly curved. Crossvenation fine reticulate (Fig. 7.1).
Occurrence
Lawrence, Commentry, Souss.
Description
The following is based on more than 25 drawings out of 36 forewings; 21 of them are figured here (Figs. 8–11). The size of the Souss specimens varies from 22 mm in length and 8 mm in width (CDUE TaV-3-17; Fig. 8.16) to 29 mm in length and 11 mm in width (CDUE TaV-3-X0-14; Fig. 8.4), as reconstructed. Costal field about 53%–70% of forewing length. Nine to ~19 pectinate simple or forked branches arise at an angle of about 45°–50° from the Sc. The number of Sc branches terminating at the wing margin is not strongly related to Sc length. In CDUE TaI-13d-26 (Fig. 8.14), 11 Sc branches (lowest number) terminate at the wing margin; the Sc is 62% of the wing length. In CDUE TaV-3-19 (Fig. 9.6), 23 Sc branches (highest number) terminate at the wing margin, and the Sc is 68% of the wing length. R branches arise from R stem by forking mostly. The arrangement of the first two branches is highly variable (cf. Fig. 9.10, 9.12). In 12 out of 20 wings, where it is visible, the first branch arises from the R stem in a distinct distance to the next following ones. In these cases the first branch is multiple forked with altogether mostly four branches; exceptions are CDUE TaI-3-47a,b (Fig. 9.10), with six branches, and CDUE TaI-13e-1 (Fig. 9.12), with only two. In CDUE TaV-3-X1-22 (Fig. 10.2), the first R branch ends blind in the area between R and Sc. In the remaining seven specimens, the first two branches arise close together from the R stem but at a distinct distance to the next following ones; in these cases, the second branch usually has more twigs than the first branch (e.g., CDUE TaI-13d-54, Fig. 9.8). The first branch of the M runs straight to the wing tip, where it ends forked into two or four branches close before the apical wing margin. The remaining M stem is two or three times forked into simple or up to three times forked branches. Crossvenation (Fig. 7.1) reticulate, forming a fine meshwork of six to eight cells between CuA and M, of eight to nine cells at the widest distance between M and Sc, and of five to six cells between the anterior CuA branches.
Description of specimen CDUE TaV-3-X1-30: cockroach in dorsal view, tectonically slightly deformed (Fig. 11.1, 11.2); the dimensions in the description are based on the retrodeformed drawing (Fig. 12). Basal part of a right forewing with the completely preserved costal field of 17 mm length. Sc emits fourteen partially furcated twigs; altogether sixteen branches end at the anterior wing margin. Pronotal shield of 10 mm × 7.5 mm size. In front of the pronotal shield is an oval head capsule of 3.5 mm × 2.5 mm dimension; because of deformation, no details visible. An only slightly coalified crescent-shaped and vaguely dotted area on the left side of the head may represent remnants of a compound eye. Parts of both filiform antennae preserved, the left one with the scape at the base. A triangular element at the anterior border of the head capsule may belong to the left mandible. On the right site all three legs are preserved; on the left side the prothoracic leg only. Coxae are not visible, with the exception of the right prothoracic leg, where the coxa is indistinctly observable below the pronotal shield. The same concerns the femora, which could only be badly identified below the pronotal shield and the right forewing; trochanter could not be recognized. The tibiae bear two rows of robust spines. The tarsi consist of five segments, the last one, the pretarsus, consists of a pair of claws and a roundish median lobe, the arolium. The reconstructed lengths of the tibia and the tarsus are 5.1 mm and 5.5 mm, respectively, at the prothorax, 5.5 mm and 5.5 mm at the mesothorax, and 7.6 mm and 7.1 mm at the metathorax.
Description of specimen CDUE TaV-3-X1-27: counter imprint of a nearly complete right forewing and the basal part of the left forewing, part of the pronotal shield as well as the right prothoracic, and a fragment of the left mesothoracic leg (Fig. 11.3, 11.4). Preserved size of the right wing 22.5 × 10 mm, reconstructed natural size 26.6 mm × 10 mm. No differentiation between the R branches. Reconstructed width of pronotal shield ~9 mm. The tibiae of both preserved legs bear two rows of robust spines. Length of tibia at the prothoracic leg not exactly determinable, reconstructed length of tarsus 4.8 mm; the pretarsus shows the arolium framed by a pair of claws.
Description of specimen CDUE AGXXIV-17: imprint of two nearly complete forewings, fragments of the associated hindwings, and the pronotal shield with questionable remnants of the head capsule in front of it (Fig. 11.5, 11.6). Preserved size of the left forewing 28 mm × 11 mm, reconstructed natural size 28.6 mm × 11 mm, preserved size of the left hindwing 18.8 mm × 8.4 mm. The number of main vein branches terminating at the wing margins is nearly identical in both forewings; the branching pattern varies slightly as follows. The number of Sc branches is nearly identical—left wing 15, right wing 16. The same is observed at the R—left 15 branches, right 16. At the left wing the multiple-forked first R branch arises from the R stem in a certain distance to the next following ones; at the right wing the distances between all branches are the same. M in both wings with nine branches; first M branch arises in both wings at a distinct distance from the next following ones; at the left wing simple forked, at the right wing three times forked. CuA with 19 branches terminating at the wing margin at left wing and 18 branches at the right wing. Hindwings not well enough preserved to allow a detailed description. Size of the pronotal shield 8.7 × 10 mm.
Materials
Figured specimens: CDUE TaV-3-4a,b (Fig. 8.1, 8.2), CDUE TaV-3-X0-14 (Fig. 8.3, 8.4), CDUE TaI-13d-50 (Fig. 8.5, 8.6), CDUE TaV-3-X1-4 (Fig. 8.7, 8.8), CDUE TaV-3-10a,b (Fig. 8.9, 8.10), CDUE TaV-3-49 (Fig. 8.11, 8.12), CDUE TaI-13d-26 (Fig. 8.13, 8.14), CDUE TaV-3-17 (Fig. 8.15, 8.16), CDUE TaV-3-X1-25 (Fig. 9.1, 9.2), CDUE TaI-13d-61a,b (Fig. 9.3, 9.4), CDUE TaV-3-19 (Fig. 9.5, 9.6), CDUE TaI-13d-54 (Fig. 9.7, 9.8), CDUE TaI-3-47a,b (Fig. 9.9, 9.10), CDUE TaI-13e-1 (Fig. 9.11, 9.12), CDUE TaV-3-X1-22a,b (Fig. 10.1, 10.2), CDUE A5/13 (Fig. 10.3, 10.4), CDUE A5/14 (Fig. 10.5, 10.6), CDUE TaV-3-X1-30 (Fig. 11.1, 11.2), CDUE TaV-3-XI-27 (Fig. 11.3, 11.4), CDUE AGXXIV-17 (Fig. 11.5, 11.6); all from Souss (see Table 1 for details).
Unfigured specimens: CDUE M2-12, CDUE M4-15a,b, CDUE M4-18a,b, CDUE M4-48a,b, CDUE A5/75, CDUE AGV-22c-30, AG XXIV-16, CDUE TaI-13-4, CDUE TaI-13d-29, CDUE TaI-13d-30, CDUE TaV-3-18, CDUE TaV-3-XI-21, CDUE TaV-3-XI-31, CDUE TaV-3b, CDUE Ig1-6, CDUE Ig1-15, CDUE TaI-13d-46, CDUE TaI-13d-60; all from Souss (see Table 1 for details).
Remarks
The figure of the holotype (and only specimen) of Etoblattina occidentalis in Scudder (Reference Scudder1890) does not provide enough information. Precise information on the venation pattern of the holotype is gained from Handlirsch (Reference Handlirsch1906b, pl. 21, fig. 16) and the reinvestigation of the type specimen USNM 38071. An additional four specimens from Lawrence, referred to this species and figured in Sellards (Reference Sellards1908) (see synonymy list), give some information on intraspecific variability. Because of this, Etoblattina obscura and Etoblattina fulva, also from Lawrence, are synonymized here with Phyloblatta occidentalis. The Souss specimens described here are, in all diagnostic features of the venation pattern, identical with P. occidentalis. That concerns, above all, the construction of R, M, and CuA. Etoblattina parva, Etoblattina queribunda, Etoblattina flebilis, Etoblattina subtilis, and Etoblattina vulpis described by Meunier (Reference Meunier1921) from Commentry were synonymized as Phyloblatta parva by Schneider (Reference Schneider1983b). Here, the latter and the included synonymous species are synonymized with P. occidentalis because of the complete identity of the venation pattern of the forewings.
Genus Anthracoblattina Scudder, Reference Scudder1879
Type species
Anthracoblattina spectabilis (Goldenberg, Reference Goldenberg1869); Wettin; MH 1125, by original designation.
Diagnosis
Elongate ellipsoidal forewings of up to 50 mm length, up to three times as long as wide. Costal field basally up to half as wide as distally, about 70%–75% of forewing length. Sc with pectinate branches inclined apically, some branches forked. R weakly sigmoidal, multiple forked with the first fork originating anterior of the distal end of the anal field, branches terminating anterior of the wing tip on the anterior wing margin. M divided shortly behind the first fork of the R stem into multiple-forked MA and MP, branches covering an area extending from the wing tip to the transition between wing tip and posterior wing margin. CuA runs, after the basal curve, inclined and more or less straight toward the posterior wing margin; first posteriorly pectinate twigs arise mainly by branching from CuA stem; all further branches arise by (multiple) forking. Basal part of CuP gently curved, apical part straighter. AnA simple and clearly separated from basal CuP and from first AnP (Fig. 7.2). AnP branches simple or forked. Crossvenation reticulate and/or anastomosing-striate (Fig. 7.2, 7.3).
Occurrence
Pennsylvanian to Permian, Europe, North and South America, Siberia (Schneider Reference Schneider1983a); Pennsylvanian, North Africa (herein).
Remarks
Anthracoblattina seems to be the most conservative phyloblattid genus of the late Paleozoic. Well-developed RA/RP and MA/MP as in this genus (e.g., Schneider, Reference Schneider1983b, pl. 3, figs. 1–11) and especially the clear differentiation of the AnA from the CuP and the AnP are typical elements of the venation pattern of Westphalian (Bashkirian–Moscovian) blattoids in general (e.g., Wei et al., Reference Wei, Béthoux, Guo, Schneider and Ren2013, figs. 1–3; Zhang et al., Reference Zhang, Schneider and Hong2013, fig. 6E). This basic venation pattern could be traced through time from Anthracoblattina poryvaikensis (Becker-Migdisova, Reference Becker-Migdisova, Rodendorf, Becker-Migdisova, Martynova and Sharov1961), Westphalian–Stephanian (Moscovian–Kasimovian) transition, Kuznetsk basin, Russia, up to Anthracoblattina sp. from the early Permian, Sakmarian, Obora entomofauna, Czech Republic. High intraspecific and individual variability, even in one wing pair (e.g., Schneider, Reference Schneider1978b, pl. 41, fig. 3a–c, Reference Schneider1983b, pl. 3, fig. 3a, b), hampers the determination of species. So far, mainly the crossvenation structures and wing sizes are used for species determination.
Anthracoblattina ensifera-gigantea group
Figures 7.2, 7.3, 13.1–13.8
Holotype
Anthracoblattina gigantea Brongniart, Reference Brongniart1893: MNHN.F.R51446; Anthracoblattina ensifera Brongniart, Reference Brongniart1893: MNHN.F.R51420; both from Commentry. Synonyms are listed in Schneider (Reference Schneider1983b).
Diagnosis
As for the genus with the following specifications. Mean length of forewings 40–50 mm. Sc with ~10–15 partly forked branches arising at an angle of about 30°–35° from the Sc stem. R mostly forked in RA and RP, the latter with more branches than the former. CuA runs more or less straight toward the transition of the wing tip into the posterior wing margin or ends simply bended at the posterior wing margin; the first up to seven posteriorly pectinate twigs arise mainly by branching from CuA stem, all further by (multiple) forking. Crossvenation mainly anastomosing-striate in A. ensifera and mainly reticulate in A. gigantea.
Occurrence
Commentry, Souss.
Description
Specimen CDUE TaI-13e-2 (Figs. 7.2, 13.1, 13.2): left forewing of 40 mm × 16 mm natural size. Sc with 15 pectinate branches; some branches forked. Altogether, 19 branches terminate at the anterior wing margin. RA multiple forked, sending six branches against the anterior wing margin; multiple forked, sending seven branches against the wing margin and terminating shortly above the middle of the wing tip. MA multiple forked into 12 branches; MP multiple forked into six branches. CuA runs, after the basal curve, inclined and more or less straight toward the posterior wing margin; first two or three posteriorly pectinate twigs arise by branching from CuA stem, all further branches arise by forking; CuA sends altogether 12 branches to the posterior wing margin. Crossvenation (Fig. 7.2) in the anal field and in the basal part, as well as in a small area in the middle of the wing, is anastomosing-striate with transition into reticulated over the rest of the wing.

Figure 13. Anthracoblattina Scudder, Reference Scudder1879. (1–8) Forewings of the Anthracoblattina ensifera-gigantea group from CDUE locality 82-TaI near Tanamert village, Oued Issene Canyon, Ida Ou Ziki subbasin, Oued Issene Formation; (9, 10) forewing of Anthracoblattina striata (Meunier, Reference Meunier1921) from CDUE locality 74-M4 at the mountain road from El Menizla to Agadir Ou Anzizen, Ida Ou Zal subbasin, El Menizla Formation, Stephanian A/B (Kasimovian). Drawings show specimens partially retrodeformed and mirrored to facilitate comparison. Size data refer to the preserved sizes of the specimens. (1, 2) CDUE TaI-13e-2 (40 mm × 16 mm); (3, 4) CDUE TaI-13d-48a,b (33 mm × 14 mm); (5, 6) CDUE TaI-13d-25 (38 mm × 19 mm); (7, 8) CDUE TaI-13e-3 (24 mm × 20 mm); (9, 10) CDUE M4-56a,b (22 mm × 10 mm). Scale bars = 5 mm.
Specimen CDUE TaI-13d-48a,b (Figs. 7.3, 13.3, 13.4): nearly complete right forewing of 33 mm × 14 mm preserved size, natural length about 37 mm. Sc with 13 pectinate branches, some branches forked; altogether 19 branches terminate at the anterior wing margin. RA multiple forked, sending four branches against the anterior wing margin; RP multiple forked, sending six branches against the wing margin and terminating shortly below the middle of wing tip. MA and MP multiple forked; number of branches unknown because of preservation. CuA runs, after the basal curve, strongly inclined toward the posterior wing margin; all branches arise by forking; CuA sends altogether only nine branches to the posterior wing margin. Crossvenation in the whole wing polygonal reticulate (Fig. 7.3).
Specimen CDUE TaI-13d-25 (Fig. 13.5, 13.6): apical part of a forewing, 38 mm × 19 mm preserved size, natural length about 49 mm, with all diagnostic features of Anthracoblattina. Crossvenation polygonal reticulate.
Specimen CDUE TaI-13e-3 (Fig. 13.7, 13.8): basal part of a right forewing, 24 mm × 20 mm preserved size, natural length about 50 mm, with all diagnostic features of Anthracoblattina. Crossvenation, so far preserved, in the anal field and in parts of the proximal half of the wing anastomosing-striate.
Materials
Two nearly complete forewings, CDUE TaI-13e-2 (Fig. 13.1, 13.2) and CDUE TaI-13d-48a,b (Fig. 13.3, 13.4), as well as the apical half of a forewing, CDUE TaI-13d-25 (Fig. 13.5, 13.6) and the basal half of a right forewing, CDUE TaI-13e-3 (Fig. 13.7, 13.8) and two basal parts of forewings, CDUE TaI-13b-6a,b and TaI-13d-78a,b (not figured); all from Souss (see Table 1 for details).
Remarks
Anthracoblattina occurs in nearly all late Paleozoic blattid-dominated entomofaunas but is known by only single specimens in each association (e.g., Schneider, Reference Schneider1983a; Schneider et al., Reference Schneider, Lucas, Rowland, Lucas and Zeigler2004). A larger number of forewings is known only from Commentry. Altogether, nearly 30 forewings from there have been described and figured by Brongniart (Reference Brongniart1893), Meunier (Reference Meunier1921), and Bolton (Reference Bolton1925) and regarded as belonging to about 25 different species by Handlirsch (Reference Handlirsch1937). They have been synonymized by Schneider (Reference Schneider1983b) as the Anthracoblattina ensifera-gigantea group of species because the only difference among all of them could be a mainly anastomosing-striate crossvenation as seen in A. ensifera and a mainly reticulate crossvenation in A. gigantea.
All previously described specimens correspond in their venation pattern to the A. ensifera-gigantea group as it was characterized and figured by Schneider (Reference Schneider1983b, pl. 3, figs. 1–11). Specimens CDUE TaI-13e-2 and CDUE TaI-13e-3 match in their crossvenation with A. ensifera Brongniart, Reference Brongniart1893, which shows basally a larger area with anastomosing-striate crossveins changing gradually toward the apex into anastomosing-reticulate crossveins (Schneider, Reference Schneider1983b, fig. 2). Specimens CDUE TaI-13d-48 and CDUE TaI-13d-25 are in the type of the crossvenation identical with A. gigantea Brongniart, Reference Brongniart1893, which possesses a nearly uniform reticulate crossvenation in the forewings. Anthracoblattina mendesi Pinto and Sedor, Reference Pinto and Sedor2000 (early Permian, Parana basin, Brazil), recently revised on the basis off new well-preserved body remains and wings (Ricetti et al., Reference Ricetti, Schneider, Iannuzzi and Weinschütz2016), differs from the A. ensifera-gigantea species group in the nearly strip-like and basally only minimally narrowed costal field (Ricetti et al., Reference Ricetti, Schneider, Iannuzzi and Weinschütz2016, figs. 4–6); the reticulate crossvenation (Ricetti et al., Reference Ricetti, Schneider, Iannuzzi and Weinschütz2016, fig. 6D) is similar to A. gigantea.
The short CuA, with only nine branches in CDUE TaI-13-48 (Fig. 13.4), is a morphological variation; the last branch of MA is most likely an anterior branch of CuA translocated onto MA, a type of transformation recurrent in insects (Béthoux and Wieland, Reference Béthoux and Wieland2009; Béthoux, Reference Béthoux2012), including blattoids (e.g., Béthoux et al., Reference Béthoux, Schneider and Klass2011; Ross, Reference Ross2012; Guo et al., Reference Guo, Béthoux, Gu and Ren2013). A similarly shortened CuA is observed in the basal forewing fragment CDUE TaI-13e-3 (Fig. 13.8) with seven preserved of primarily about eight or nine branches only.
Anthracoblattina striata (Meunier, Reference Meunier1921)
Figure 13.9, 13.10
- Reference Meunier1921
Etoblattina striata Meunier, p. 91, fig. 123, pl. 18, fig. 6.
- Reference Handlirsch1937
Phyloblatta striata; Handlirsch, p. 77.
- Reference Schneider1983b
Anthracoblattina striata; Schneider, p. 84, pl. 5, fig. 5.
- Reference Hmich, Schneider, Saber, Voigt, El Wartiti, Lucas, Cassinis and Schneider2006
Anthracoblattina sp.; Hmich et al., fig. 8B.
Holotype
Etoblattina striata Meunier, Reference Meunier1921; Commentry; MNHN.F.R.51541.
Diagnosis
As for the genus with the following specifications. Forewings of about 30 mm × 10 mm size. Sc with ~10–15 partly forked branches. First branch of R rarely simple, mostly multiple forked; remaining R stem multiple forked. CuA runs weakly sigmoid to nearly straight toward the transition of the wing tip into the posterior wing margin; first up to about eight posteriorly pectinate twigs arise mainly by branching from CuA stem, all further by (multiple) forking. Crossvenation polygonal reticulate with transitions into anastomosing-striate.
Occurrence
Commentry, Souss.
Description
Left forewing, 22 mm × 10 mm preserved size, natural size 29.5 mm × 10 mm. Sc with 14 partially forked pectinate branches. First two R branches arise close together from the R stem, all further branches with a distinct distance from the former; first R branch multiple forked into four twigs, the second simple forked. R branches cover the area between the end of the costal field and the middle of the wing tip. M divided shortly anterior of half of the wing length by a fork into MA and MP. MA with a long stem, forked. MP multiple forked. CuA runs nearly straight toward the transition of the wing tip into the posterior wing margin; the first five pectinate simple or forked twigs arise by branching, all further by forking from the CuA stem. More than six An veins. Crossvenation mainly polygonal reticulate; in the posterior apical part of the anal field and in the middle of the wing between CuA branches as well as between CuA and M transitions to irregular reticulate and anastomosing-striate.
Remarks
CDUE M4-56a,b is in all diagnostic features identical with the type (and only) specimen of A. striata. Minimal differences relate to the CuA, which is weakly sigmoid in the holotype but nearly straight in the Souss specimen, as well as to the more RA-like construction of the first (two) R branches in the latter compared to the unspecific first R branches in both forewings of the holotype (Meunier, Reference Meunier1921, fig. 123). These differences are within the whole range of variability of Anthracoblattina, as demonstrated by Schneider (Reference Schneider1983b, pl. 3, figs. 1–11).
Phyloblatta sp.
Figure 14.1–14.8
Materials
Figured specimens: CDUE AGXXIV-16 (Fig. 14.1, 14.2), fragment of a left forewing (preserved size 22 mm × 10 mm) and a left hindwing (preserved size 21 mm × 10 mm). CDUE TaV-3-X0-16 (Fig. 14.3, 14.4), strongly deformed hindwing (preserved size 24.5 mm x 15 mm); CDUE TaI-13d-52a,b (Fig. 14.5, 14.6), hindwing fragment (preserved size 20 mm x 10 mm); CDUE TaV-2a (Fig. 14.7, 14.8), hindwing fragment (preserved size 20 mm x 10 mm); all from Souss (see Table 1 for details).

Figure 14. Hindwings of Phyloblatta Handlirsch, Reference Handlirsch1906a from CDUE locality 79-AGXXIV at the mountain road from El Menizla to Agadir Ou Anzizen, Ida Ou Zal subbasin, El Menizla Formation, and from CDUE localities 80-TaV and 82-TaI near Tanamert village, Oued Issene Canyon, Ida Ou Ziki subbasin, Oued Issene Formation, Stephanian A/B (Kasimovian). Size data refer to the preserved sizes of the specimens. (1, 2) CDUE AG XXIV-16 (fragment of a left forewing 22 mm × 10 mm, left hindwing 21 mm × 10 mm); (3, 4) CDUE TaV-3-X0-16 (strongly deformed, 24.5 x 15 mm); (5, 6) CDUE TaI-13d-52a,b (20 x 10 mm); (7, 8) CDUE TaV-2a (20 x 10 mm). Scale bars = 5 mm.
Unfigured specimens: CDUE TaI-13d-21; CDUE TaI-13d-55; CDUE TaV-X1-31; CDUE AGV-22c-28; CDUE AGV-22c-33; CDUE AGXXIV-3-18; all from Souss (see Table 1 for details).
Remarks
Identifying hindwings of Paleozoic blattoids (and of Paleozoic Neoptera in general) at the species level is challenging. Hindwings are rarely, and then often incompletely, preserved because of their delicate structure. If they are preserved articulated to the body together with forewings, on which the blattoid classification is based, the hindwings are often more or less covered by the forewings, making the hindwings incompletely discernible. A few exceptional preserved specimens allowed fore- and hindwings to be related (e.g., Béthoux et al., Reference Béthoux, Schneider and Klass2011; Guo et al., Reference Guo, Béthoux, Gu and Ren2013). To contribute to the identification of hindwings, we illustrate some of those best preserved from the Souss basin. They are all regarded as belonging to Phyloblatta sp. because of their length of roughly 20 mm and by comparison with the venation pattern of the hindwings in the Phyloblatta occidentalis specimen CDUE AGXXIV-17 (Fig. 11.6). Hindwings of Compsoblatta are of similar size to those of Phyloblatta but would have a color pattern of similar design to the corresponding forewings (JWS personal observation). Hindwings of the Anthracoblattina ensifera-gigantea group are larger, having a length of about 40–50 mm (e.g., Brongniart, Reference Brongniart1893, pl. 47, fig. 5).
Family Compsoblattidae Schneider, Reference Schneider1983a
Genus Compsoblatta Schlechtendal in Handlirsch, Reference Handlirsch1906b
Type species
Compsoblatta mangoldti Schlechtendal in Handlirsch, Reference Handlirsch1906b; MH 36/14; Halle Formation, Lower Rotliegend (Asselian, early Cisuralian); Germany.
Diagnosis
Elongate ellipsoidal forewings of about 20–35 mm in length. Costal field strip-like, up to about 60%–70% of forewing length. Sc pectinate, branches about 45° inclined to the apex and ending at the anterior wing margin. R sigmoidal, R branches terminate at anterior wing margin. M sigmoidal, multiple forked, branches covering an area extending from the wing tip to the transition between wing tip and posterior wing margin. CuA strongly sigmoidal. First CuA twigs arise by branching from CuA stem and rarely forked; all further branches arise by furcation. CuP regularly bended. The general venation pattern is similar to that of Phyloblatta, but in contrast to this genus the crossvenation in the basal three quarters of the wings consists of seams formed by crossvein bases along the main veins and their branches; the remaining wing surface shows a delicate irregularly reticulated cross venation. In contrast to the spiloblattinids, which show similar crossvein seams, the areas between the main veins are not distinctly broadened and the venation is generally denser in compsoblattids.
Occurrence
Pennsylvanian to Permian, Europe (Schneider, Reference Schneider1983a); Pennsylvanian, North Africa (herein).
Compsoblatta sp. Figures 7.4, 15.1, 15.2
Description
Elongate ellipsoidal forewing of 18 mm × 9 mm preserved size, and 20 mm × 9 mm reconstructed natural size. Costal field about 60% forewing length. Sc with eleven pectinate branches arising at an angle of about 40°–45° from the Sc stem; some branches forked. Altogether, twelve branches terminate at the anterior wing margin. R sigmoidal with five branches; the first branch arises shortly anterior of the distal end of the anal field from the R stem, some branches forked; first one RA-like multiple forked. Altogether, 10 R branches terminate on the anterior wing margin. M strongly sigmoidal, divided by forking just before half of wing length into MA and MP. MA multiple forked. MP branches arise by forking and are partly forked; the last one ends at the transition of the wing tip into the posterior wing margin. CuA sigmoidal, first five twigs arise by branching from proximal CuA stem, all further branches in the apical part of the CuA arise by multiple forking; first fork of CuA stem at about half of the wing length; CuA sends altogether 13 branches to the posterior wing margin. CuP simple and curved. Eight An veins. Crossvenation (Fig. 7.4) consists of seams formed by crossvein bases along the main veins and their branches in parts of the wing; the wing surface encircled by the seams shows a delicate irregularly reticulated crossvenation.

Figure 15. Compsoblatta sp., counterpart of a left forewing from CDUE locality TaI-82 near Tanamert village, Oued Issene Canyon, Ida Ou Ziki subbasin, Oued Issene Formation, Stephanian A/B (Kasimovian). Preserved size of the specimen CDUE TaI-13d-34 = 18 mm x 9 mm. Scale bar = 5 mm.
Remarks
The family Compsoblattidae includes blattoids displaying an enigmatic mixture of a Phyloblatta-like venation pattern combined with a wing coloration comparable to the Spiloblattinidae (Schneider, Reference Schneider1983a, pl. 3, figs. 7, 10). The main veins and their branches in the basal two-thirds of the forewings are framed by seams of short crossvein stubs. These seams and the remaining wing surface are black colored if coalified organic material is preserved (Schneider, Reference Schneider1984a, pl. 7, figs. 2, 4). The areas enclosed by the seams are pale and show the color of the embedding rock (for details of color patterns in Compsoblattidae and Spiloblattinidae, see Schneider, Reference Schneider1984a). This wing coloration is somewhat more indistinct than in spiloblattinids, and till Bolton (Reference Bolton1925, p. 42) it was (and still is) overlooked by researchers. Consequently, some forms formerly described as phyloblattids may be in reality compsoblattids.
The specimen CDUE TaI-13d-34 lacks coalified remains of the primary organic wing material; nevertheless, the diagnostic color spots and the crossvenation of the vein seams are partially visible (Figs. 7.4, 15.1). From Commentry, only two Compsoblatta species, Compsoblatta ovata (Meunier, Reference Meunier1921) with four specimens (Meunier, Reference Meunier1921, fig. 113, pl. 17, fig. 8), and Compsoblatta couloni (Meunier, Reference Meunier1921) with two specimens (Meunier, Reference Meunier1921, pl. 15, fig. 10), were identified by Schneider (Reference Schneider1983b, p. 84–85, pl. 1, figs. 6–10, pl. 5, fig. 6) based on Meunier (Reference Meunier1921). The venation pattern of CDUE TaI-13d-34 is nearly identical with C. ovata and has only one branching twig more from the CuA stem before the apical fan-like forking (i.e., five instead of four). C. couloni has only three twigs from the CuA stem before the apical forking. Phyloblatta monubilis spathulata Bolton, Reference Bolton1925 (Bolton, Reference Bolton1925, p. 40–42, fig. 14, pl. 2, fig. 19) from Commentry is synonymized here with C. ovata (Meunier, Reference Meunier1921) because of identical venation patterns and the presence of seams of crossvein bases (Bolton, Reference Bolton1925, p. 42). In this specimen, both forewings and parts of the pronotum are preserved (Bolton, Reference Bolton1925, fig. 14). One of the forewings has three branches, the other one four branches at the CuA before its apical forking. A similar individual variation in the branching and furcation pattern of the CuA is visible in two forewing pairs of Compsoblatta anaglyptica (Schlechtendal in Handlirsch, Reference Handlirsch1906b) from Wettin (Schneider, Reference Schneider1978b, pl. 62, figs. 2, 3). Substantially higher than this individual is the intraspecific variability of forms with a phyloblattid wing venation pattern. This becomes evident when larger numbers of specimens are available for investigation (Ross, Reference Ross2012), such as demonstrated here for P. occidentalis (Figs. 8–11). In this light, the differences in the venation pattern between C. ovata and C. couloni, as discussed by Schneider (Reference Schneider1983b, p. 84–85), are meaningless.
As far as preserved in CDUE TaI-13d-34 (Fig. 15.2), the size of the color spots is somewhat smaller than in C. ovata from Commentry according to our examination of a photograph of the specimen MNHN.F.R51084, holotype of Etoblattina ovata Meunier, Reference Meunier1921, available at the MNHN Paris collection website (http://coldb.mnhn.fr/catalognumber/mnhn/f/r51084), and C. anaglyptica from Wettin (cf. Schneider, Reference Schneider1983a, pl. 3, fig. 7). Whether this is caused by sexual dimorphism only, as in spiloblattinids, or these differences could be used for the definition of a new species must be clarified with further finds from Souss.
Discussion
Although the late Carboniferous Souss basin is situated in the present North Africa at the former southern fringe of the Variscan Central Pangean Orogen and therefore close to Gondwana, the biotas of this basin are definitely of Euramerican character. This is evidenced by the composition of the flora (Broutin et al., Reference Broutin, Ferrandini and Saber1989; Hmich et al., Reference Hmich, Schneider, Saber, Voigt, El Wartiti, Lucas, Cassinis and Schneider2006), the tetrapod tracks (Lagnaoui et al., Reference Lagnaoui, Voigt, Belahmira, Saber, Klein, Hminna and Schneider2018), and the entomofauna (Hmich et al., Reference Hmich, Schneider, Saber and El Wartiti2003, Reference Hmich, Schneider, Saber, El Wartiti, Lucas and Zeigler2005). In Euramerica, the Souss is the oldest diverse post-Westphalian entomofauna, followed in age by the Commentry and Wettin entomofaunas (Schneider and Werneburg, Reference Schneider and Werneburg2012). The Souss entomofauna is dominated, both in the number of specimens and in diversity, by the family Phyloblattidae. Phyloblattids first appear with typical forms in the late Westphalian of the Euramerican and Angaran provinces, for example, Phyloblatta mazona (Scudder, Reference Scudder1882) (Mazon Creek, Illinois) and Phyloblatta curvinervosa (Becker-Migdisova, Reference Becker-Migdisova, Rodendorf, Becker-Migdisova, Martynova and Sharov1961) (Kuznetsk basin, Siberia). They became dominant during the Stephanian A (Kasimovian) climatic wet phase after the disappearance of archimylacrids and a decrease in the diversity of mylacrids during the late Westphalian (late Moscovian) dry phase (Roscher and Schneider, Reference Roscher, Schneider, Lucas, Cassinis and Schneider2006; Tabor and Poulsen, Reference Tabor and Poulsen2008; DiMichele, Reference DiMichele2014).
The distribution of phyloblattids in the various insect beds of the Souss basin supports some older observations regarding habitat preferences of single genera (Schneider, Reference Schneider1983a). Phyloblatta is one of the most common forms in the late Paleozoic Euramerican entomofauna of ever-wet to seasonal wet climates. It occurs in the thickly vegetated habitats of swamps and mires as well as in more sparsely vegetated lakeshore areas and in the riparian vegetation of river courses of intra- and perimontaneous basins and foreland basins and in the coastal plains of seashores (Schneider, Reference Schneider1983a; Rasnitsyn et al., Reference Rasnitsyn, Aristov, Gorochov, Rowland and Sinitshenkova2004; Schneider et al., Reference Schneider, Lucas, Rowland, Lucas and Zeigler2004, Reference Schneider, Lucas, Trümper, Stanulla and Krainer2016). With increasing aridization in the early Permian, small forms such as Phyloblatta praecompacta Schneider, Reference Schneider1984b and Phyloblatta compacta (Sellards, Reference Sellards1908) appear, for example, at Obora (late Artinskian), Czech Republic, and Elmo (Kungurian), Kansas (Schneider, Reference Schneider1984b). They were living in a semiarid climate marked by strong seasonality.
Anthracoblattina occurs from the Late Pennsylvanian (Westphalian–Stephanian transition) to the middle Cisuralian (Artinskian–Kungurian) with single finds at all European and North American insect sites, but has a seemingly limited habitat preference (Schneider, Reference Schneider1983a). In the insect associations of thickly vegetated mires, such as the Wettin insect sites, Anthracoblattina is rare. By contrast, lake deposits such as Commentry and marine lagoonal deposits such as those of the Parana basin can be rich in Anthracoblattina (Schneider, Reference Schneider1983b; Ricetti et al., Reference Ricetti, Schneider, Iannuzzi and Weinschütz2016). This and the finds in the mixed marine and continental coastal pond deposits of Carrizo Arroyo, New Mexico (Schneider et al., Reference Schneider, Lucas, Rowland, Lucas and Zeigler2004, Reference Schneider, Lucas, Trümper, Stanulla and Krainer2016; Rasnitsyn et al., Reference Rasnitsyn, Aristov, Gorochov, Rowland and Sinitshenkova2004), as well as in the plant fragment– and bivalve-rich lacustrine littoral sediments of the CDUE 82-TaI site of the Souss basin, support the conclusion that sparsely vegetated shores of lakes and seas were the preferred habitat of Anthracoblattina.
The database for Compsoblatta is currently too small to allow solid paleoecological conclusions. However, because of the common co-occurrence with Phyloblatta, a similar habitat preference is assumed.
Conclusion
In summary, the Souss entomofauna, comprising phyloblattids, compsoblattids, spiloblattinids, poroblattinids, and mylacrids (Hmich et al., Reference Hmich, Schneider, Saber and El Wartiti2003, Reference Hmich, Schneider, Saber, El Wartiti, Lucas and Zeigler2005, Reference Hmich, Schneider, Saber, Voigt, El Wartiti, Lucas, Cassinis and Schneider2006), is similar to Euramerican entomofauna of the Late Pennsylvanian and earliest Permian but displays some differences. Interestingly, the Ida Ou Zal subbasin is characterized by the dominance of xeromorphic opsiomylacrids (Hmich et al., Reference Hmich, Schneider, Saber and El Wartiti2003) and the rarity of phyloblattids, whereas in the Ida Ou Ziki subbasin, opsiomylacrids are comparatively rare and phyloblattids are common. Further study of the insect faunas of the subbasins will contribute to the understanding of late Paleozoic insect evolution in the context of paleoecology and paleoenvironment in general.
Acknowledgments
This publication is part of a Ph.D. project at the Chouaïb Doukkali University and the Technical University Bergakademie Freiberg, generously supported by the grant A/10/9816 of the German Academic Exchange Survey (DAAD) to the first author. JWS acknowledges the support of the German Research Foundation (DFG), grant DFG Schn 408/21, which enables the fieldwork in Morocco and further studies as well as the support by a subsidy from the Russian Government to support the Programme of Competitive Growth of Kazan Federal University among the World's Leading Academic Centres. The first author thanks M. Magnus for support and training in photographic techniques. Our colleagues D. Hmich, A. Lagnaoui, S. Voigt, and A. Brosig are thanked for common fieldwork and stimulating discussions. J. Utrup and S.H. Butts, Yale Peabody Museum of Natural History, New Haven, are thanked for a series of excellent photographs of the topotype of E. mazona, which help us to understand what kind of blattoid it is. A. Rasnitsyn and E. Jarzembowski helped with literature. S.G. Lucas, L. Rinehardt, and I. Kogan are thanked for the improvement of the English and taxonomic discussions. We thank the e-recolnat project, the MNHN Paris, and G. Doitteau for providing useful data. The reviewers O. Béthoux, A.J. Ross, and A. Nel are thanked for careful review and valuable hints, which have greatly improved the manuscript. The editors, B. Hunda and J. Haug, are thanked for their helpful comments. Our always good-humored driver H. Manssouri brought us with safety to the most remote places in Morocco—choukran! Last but not least, the friendly villagers of Tanamert and El Menizla are thanked for their support of our fieldwork. This publication aims to contribute to the tasks of the ‘Late Pennsylvanian – Permian – Early Triassic Non-Marine – Marine Correlation Working Group’ of the International Subcommissions on Carboniferous, Permian, and Triassic stratigraphy.