Introduction
Intelligent life is one of the most controversial issues in astrobiology. Of particular interest are the related questions of intelligent life's likelihood and time required for it's evolution. The fi term of the Drake equation represents the fraction of planets in the galaxy with life that have developed intelligent life, and therefore embodies these questions. In Rare Earth, Ward & Brownlee (Reference Ward and Brownlee2000) provide multiple lines of argument that intelligent life in the universe is rare even though microbial life is common. Counterarguments often cite the importance of convergence in evolution (e.g. Conway-Morris Reference Conway-Morris1998, Reference Conway-Morris2003, Reference Conway-Morris2010), implying that intelligence is an attribute upon which lineages, or at least some lineages, will converge, and will therefore be more common in the universe.
Research on the evolution of intelligence can be classified into two types: those focusing on the evolution of human intelligence (human intelligence phenomenon), and those with broader perspectives focusing on the evolution of intelligence in general (broad-sense intelligence phenomenon). A large body of literature addresses questions regarding the evolution of intelligence in the hominin lineage 5–7 mya to present, our lineage on the tree of life. From an astrobiological perspective, while the human intelligence phenomenon is of interest, the fi term of the Drake equation actually hinges on answers to the broad-sense intelligence phenomenon. The following questions highlight this distinction. What caused the human lineage to undergo substantial evolutionary change in brain size and cognition? Instead we might ask how and why did a species emerge with the potential to undergo this evolution in the first place? This second question changes the focus of inquiry and the time scale of interest.
If parallels exist between human and animal intelligence evolution, an effective strategy for generating questions in an astrobiological context could be to review the human intelligence literature for hypotheses that might generalize beyond the human lineage. Each reviewed hypothesis can be assessed to determine if the processes proffered would operate beyond the human lineage and its time frame. Here I discuss intelligence in the context of the Phanerozoic (542 mya to present). However, the original Drake equation fi term focuses on the evolution of intelligence after life's initiation (∼3.5 Ga).
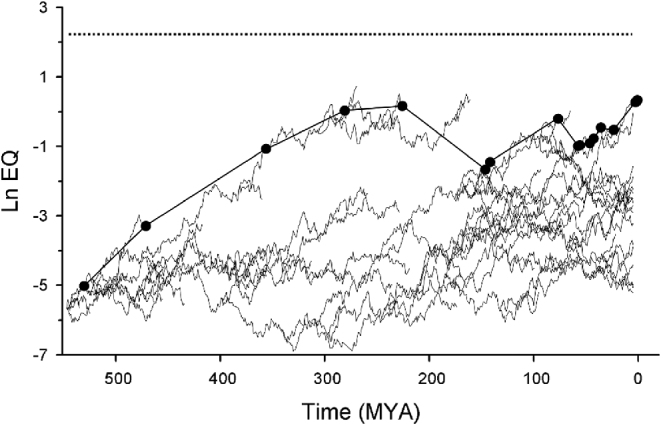
The above strategy could be useful. A paper entitled ‘Exponential evolution: Implications for intelligent extraterrestrial life’ (Russell Reference Russell1983) demonstrated a semi-log linear relationship between maximum EQ (encephalization quotient – a body mass transformed measure of brain size) and time from 542 million years ago to present. EQ is not synonymous with intelligence, but is a reasonable proxy for behaviour associated with intelligence (see hypothesis vi; Dunbar Reference Dunbar1998; Reader & Laland Reference Reader and Laland2002; Jerison Reference Jerison1955; Changizi Reference Changizi2003; Deaner et al. Reference Deaner, van Schaik and Johnson2006; Lee Reference Lee2007; Sol et al. Reference Sol, Bacher, Reader and Lefebvre2008). Importantly, EQ is fossilizable. Plotted in Russell's graph are the most encephalized organisms on Earth at each time interval (Fig. 1). This and other patterns presented in that paper suggest that the evolution of human intelligence is part of a more general process of greater encephalization over time. This paper is not well cited, suggesting that it has been largely overlookedFootnote 1. Additionally, recent reviews suggest that the difference between human and animal intelligence is of quantity not of quality (Roth & Dicke Reference Roth and Dicke2005), further supporting the notion that human intelligence can be looked at as part of a broader evolutionary process.

Fig. 1. Scatter plot of the most encephalized lineage on Earth (natural log of EQ), at each time plotted, over geological time (mya). Data are from Russell (Reference Russell1983).
The definition of intelligence has been the subject of much discussion. Sternberg (Reference Sternberg, Sternberg and Kaufman2002) provides a quick overview on the various approaches adopted for defining intelligence. Most definitions include the ability to respond flexibly and successfully to one's environment and to learn from experience. Given the wide temporal scale of interest here, we should be interested not just in abstract problem-solving intelligence but also in proxies, components and antecedents of that kind of intelligence. A large class of cognitive capabilities or behaviours might be classified as ‘intelligent’ in that they satisfy the demands inherent in the definition above (memory, domain general learning, behavioural flexibility). Determining whether or not these behavioural/cognitive capabilities tend to be correlated across species or correlated with metrics of encephalization is an empirical problem of interest to astrobiologists.
Outlined below are ten hypotheses related to both the evolution of human intelligence and intelligence in the broad sense. All can be evaluated, reviewed and compared for their relevance to each other and to the broad-sense intelligence phenomenon. Hypothesis i explores implications of random macroevolutionary processes on the evolution of intelligence. Hypotheses ii–vi are frequently expressed in the human (or primate) context while the remaining are frequently expressed in the broader context.
Some of the hypotheses I outline below have been largely overlooked by the astrobiological community. For example, the paper that introduced the expensive tissue hypothesis (hypothesis iv; Aiello & Wheeler Reference Aiello and Wheeler1995) has been cited 573 times in evolutionary and anthropological journals and elsewhere (Google Scholar search 29-12-2009) but not once in the International Journal of Astrobiology or Astrobiology. Transparently, opportunities exist in linking astrobiological perspectives with evolutionary and ecological research.
Hypotheses
i. Diffusion hypothesis or the random walk: The evolution of intelligence or encephalization is a random walk for each lineage, with a bounded lower limit of complexity and an essentially unbounded upper limit of complexity. The idea that encephalization might evolve randomly (increase or decrease) is popular among biologists because larger brains are not viewed as necessarily better than smaller brains in all ecological contexts; large brains have advantages, but come with energetic trade-offs (treated in hypotheses iii and iv). However, provided enough lineages are randomly moving along an encephalization spectrum with a lower boundary or limit, both the mean and maximal values of encephalization should increase over time. Long-term trends in evolution can exist due to passive mechanisms or driven mechanisms (McShea Reference McShea1994; Marcot & McShea Reference Marcot and McShea2007); the random walks of lineages in the diffusion hypothesis are a passive mechanism. An over-riding long-term selective pressure for more intelligent organisms is not necessary to make production of an intelligent species virtually inevitable; trends can exist without evolutionary directionality. More precisely, a trend for increasing maximal intelligence of a clade (or biota) can exist even if its individual lineages do not possess a tendency to increase or decrease in intelligence or encephalization. Humans are just the latest and highest-EQ species to walk in the ‘right’ direction. The data exist to address this hypothesis (Drake Reference Drake2003Footnote 2; Chyba & Hand Reference Chyba and Hand2005), and some work has been done (e.g. Marino et al. Reference Marino, McShea and Uhen2004; Finarelli & Flynn Reference Finarelli and Flynn2007). Importantly, depending on the random walk's parameters, it could produce either a predictable or unpredictable time frame for the evolution of intelligent life, much like the rate of diffusion of a gas can be predictable despite the fact that the motions of its particles are random. Such a pattern would have important implications for astrobiology.
Testing the diffusion hypothesis with the Russell data set
The diffusion hypothesis can be statistically tested. The Russell data (Fig. 1) represent observed patterns and can be compared with a null model simulation, based on diffusion model principles. The Russell data set may require update or alteration, but the concept will remain unchanged. Biotas could be simulated with species undergoing random encephalization changes, diversifications and extinctions, and iterated (e.g. Fig. 2). Parameters could be modified and explored and the resulting null biota simulations could be statistically compared with the observed curve. Simulations such as these have been developed for evolutionary change processes in general (McShea Reference McShea1994); the same logic applies whether the y-axis is EQ or any other character. The Russell data set represents, at each time sampled, the most encephalized organism on Earth rather than the mean EQ of all species in the biota. Tracking the most encephalized organism seems appropriate given that the object of interest is the evolution of intelligent life: an intelligent species likely being one of, or the most encephalized organism from the biota. Maxima tend to be more susceptible than means to stochastic variation (McShea personal communication). This suggests that the regularity of increasing EQ observed in Russell's data set may be difficult to account for with a diffusion model. On the other hand, some temporal stochastic variation in EQ maxima is expected in a diffusion model and the amount observed in the Russell data (Fig. 1) may not be an unlikely result of a diffusion process. Determining if observed patterns are consistent with a diffusion model will require a detailed statistical examination of the behaviour of maxima under diffusion model simulations with reasonable parameters.
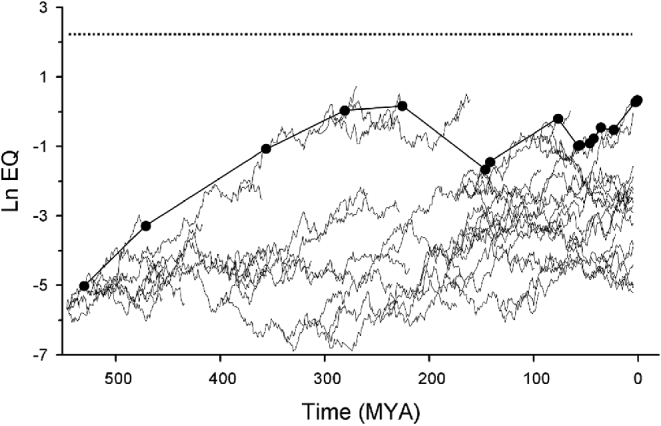
Fig. 2. Hypothetical iteration from a test of the diffusion hypothesis (produced by the author as an example). This graph is meant to be comparable to Fig. 1. The axes are the same as in Fig. 1. Plotted is a simulated biota, with diversification, extinction and EQ change parameters per time step (542 one million year intervals). The maximal EQ line is drawn in the same manner as in Russell's data set, with the same temporal sampling pattern. Interestingly, this biota does not generate a lineage with human-level EQ (dotted line) in a comparable time frame. As in Fig. 1, which lineage is the most encephalized can (and does) change from one time step to the next. The most encephalized lineage in the fifth sampled time is extinct by the sixth – in this case without a compensating EQ increase in any other lineage.
Figure 2 is an example random diffusion simulation generated by the author for the purposes of illustration. Maximal EQ has a tendency to rise in diffusion models, although in Fig. 2 this rise is less regular than Russell's observed pattern and does not attain human-level EQ in a comparable time frame. However, this is just one possible result. Models with slightly different parameters can produce results more comparable to observed patterns. If the extinction likelihood was lower, EQ drops would presumably be less likely in any particular model iteration. For a more complete description of diffusion models and a list of parameters, see McShea (Reference McShea1994).
Three main patterns are visible in Russell's data set: a general semi-log linear increase in EQ of the maximal EQ species in the biota (R 2=0.91); modifying this pattern is a change in the rate of EQ increase at the K/T boundary extinction (65 mya); and an apparent absence of instances of drops in maximal EQ between any time steps (maximal EQ always increased or remained nearly at the same level). The third pattern is surprising, given the five major mass extinctions that occurred in the last 542 million years. These three apparent regularities can be tested using diffusion simulations. The following questions can be asked. Given a set of diffusion model parameters, will the degree of regularity in observed patterns occur frequently over many model iterations or are observed patterns highly unlikely? Does the observed pattern of increase in highest EQ conform to a model of random EQ changes over time? Possible alternative models would include those hypotheses discussed in this review or a diffusion model with different parameters. Here exists an opportunity to quantitatively test ideas concerning the evolution of intelligent life with real biological data.
ii. Auto-catalysis or positive feedback: During four million years of hominid evolution, encephalization has increased exponentially (De Miguel & Henneberg Reference De Miguel and Henneberg2001). Auto-catalytic evolutionary processes have been provided as explanations for this pattern. This is a large class of hypotheses and many processes have been implicated as mechanisms underlying auto-catalytic human evolution. Wilson (Reference Wilson1975) suggests that within-species selection pressures (sociality, sexual selection) could drive auto-catalytic evolution where increasing social complexity leads to greater selection pressures for social cognitive skills, leading to even more social complexity – a positive feedback loop. Pinker (Reference Pinker2010) suggests that external selection factors can be important, where humans use intelligence (and cooperative behaviour) to extract resources from environments, in what Pinker terms our cognitive niche. Selection for a cognitive niche, perhaps interacting with social evolution, could yield self-reinforcing evolution for higher intelligence, sociality and language.
The role of social behaviour in the evolution of primate cognition has been explored in detail through the social brain hypothesis; among primates, strong correlations exist between social group size and brain size metrics, suggesting that social behaviour drives encephalization (Dunbar Reference Dunbar1998). Appropriately, astrobiologists have recently directed their attention to the role of social behaviour, particularly social structure, in intelligence evolution (António & Schulze-Makuch Reference António and Schulze-Makuch2010). If sociality were more common later in the Phanerozoic, even if the reason is simply that there are more species later in this eon, then intra-specific (within species)-driven auto-catalytic evolutionary events would plausibly be more common as well.
Auto-catalysis or evolutionary positive feedback loops need not be limited to human evolution (or the associated short time scale), and could be driven through inter-specific (between species) rather than intra-specific evolutionary mechanisms. At broader temporal and phylogenetic scales, auto-catalysis for greater intelligence and encephalization could plausibly be mediated through competitive biotic interactions (Dawkins & Krebs Reference Dawkins and Krebs1979; Russell Reference Russell and Seth Shostak1995, Reference Russell2009). Broad-scale EQ patterns appear consistent with this view (Jerison Reference Jerison1955; Jerison Reference Jerison1973). Early Paleozoic animals had small nervous systems relative to body size, and Mesozoic reptiles and dinosaurs had lower EQs than Cenozoic mammals. Indeed, early mammals had lower EQs than late Cenozoic mammals; all early mammals were either in the bottom third of extant mammalian inter-species EQ variation, or entirely below extant variation (Jerison Reference Jerison1973). The maximum, mean and mode of mammal EQs increased in the Cenozoic.
Not all mammalian faunas experienced the same EQ increases in the Cenozoic. The South American mammalian fauna was separated from the Holarctic mammalian fauna for roughly 40 million years until 2–3 mya, effectively running an evolutionary experiment. The South American herbivore fauna did not experience EQ increases comparable to the EQ increases observed among Holarctic herbivores over the same interval. To explain this discrepancy, Jerison (Reference Jerison1973) suggests that the relevant difference between the two faunas was in the presence or absence of large eutherian carnivores. In South America these carnivores were absent; in the Holarctic they were present. Jerison argues that in the Holarctic fauna an open-ended evolutionary race ensued, where predators were continually getting better at catching prey and prey were continually getting better at avoiding predators. EQ increased in both predator and prey guilds in association with the behavioural changes to avoid and catch prey (see Radinsky Reference Radinsky1978 for a critical re-analysis of carnivore and ungulate EQ patterns). Dawkins & Krebs (Reference Dawkins and Krebs1979) refer to these evolutionary feedback phenomena as ‘arms races’, and argue that they represent one theoretical rationale for an open-ended escalating positive feedback mechanism in evolution. This evolutionary process represents one theoretical reason to expect increases in EQ over long evolutionary time.
The South American ‘experiment’ seems to demonstrate that EQ increases were not inevitable at least among mammals over that 40 million year interval. Those persuaded by the view that high EQ and intelligence are inevitable should be given pause by the 40 million years where EQ remained unchanged among South American herbivores. However, if Jerison and Dawkins are correct about the existence of arms races, this has implications for the interpretation of EQ increases where and when they do occur. Trends of increasing EQ are often not simply random chances of history but the result of an evolutionary process that, though not inevitable, plausibly has a high probability. The existence of multiple lineages and continents increases the likelihood that such positive feedback systems will initiate by providing greater numbers of opportunities. Jerison's South American analysis could be regarded as preliminary, given the small sample size of South American mammals in the study. However, the intent of the research is entirely appropriate. Summarized, the goal is to look for evolutionary experiments in the various biogeographically isolated regions of the past. Patterns of morphological evolution can be compared between isolated regions. More attention should be paid to these patterns because the results could be of interest to astrobiologists.
iii. Life-history theory: In biology, the term life-history refers to the pattern and timing of events (such as age of first reproduction or age of death) in an individual's lifetime. The life-history of a species refers to the typical pattern of events in the lifetime of an individual organism of that species. Thus in biology life-history does not have the same meaning as history of life.
Humans can be understood through a life-history trait trade-off theory (Wilson Reference Wilson1975; Kaplan et al. Reference Kaplan, Hill, Lancaster and Hurtado2000). The r–K theory (Wilson Reference Wilson1975) was an early attempt to describe a spectrum of species traits that correlate in species due to ecological trade-offs. One finds a correlated set of traits among species along an r–K spectrum. K strategists focus on reproductive assurance and have low fecundity, high investment in individual offspring and long life spans, whereas r strategists invest less per offspring in large litters and have short lifespans. High EQ is a K strategy, low EQ an r strategy (Charnov & Berrigan Reference Charnov and Berrigan1993; Kaplan et al. Reference Kaplan, Hill, Lancaster and Hurtado2000; Barrickman et al. Reference Barrickman, Bastian, Isler and van Schaik2008). Trade-offs prevent one species from simultaneously holding both r and K strategies. K and r strategies are both successful, but different; humans represent an extreme K strategy. The r–K spectrum is often referred to as a fast–slow life-history spectrum.
For astrobiologists evaluating a potential trend in EQ over geological time, the lesson from r–K theory (or related theories invoking life-history trait trade-offs) is that associated trends may be expected (given an EQ trend) and could be considered germane. For example, is there a long-term trend in parental care? If so why? r–K and life-history theory establishes the link between parental care and EQ. Evaluating the broader ecological context in which EQ patterns emerge conceivably will deepen our mechanistic understanding of any such pattern, and provide a path to multiple lines of evidence for any EQ trend.
Some studies already provide information. Parental care has evolved readily in ray-finned fishes (Mank et al. Reference Mank, Promislow and Avise2005) in multiple lineages. Once a fish lineage evolves parental care, loss of parental care is rare. Yopak et al. (Reference Yopak, Lisney, Collin and Montgomery2007) point out that among the chondrichthyans (cartilaginous fishes), those with the relatively largest brains, the carcharhiniformes, are also viviparous (have live birth) and have evolved placental vivaparity. A phylogenetically broad analysis of parental care at the class or order level will be very useful for astrobiologists interested in intelligence.
iv. Expensive tissue hypothesis or brain/gut trade-off hypothesis (Aiello & Wheeler Reference Aiello and Wheeler1995): Evolution of human intelligence involved a trade-off between the metabolically expensive nervous tissues and gut tissues, thereby altering diet significantly in favour of high-quality food. The idea is that if an organism allocates to expensive CNS tissue, that organism may need to scale back allocation to other metabolically expensive tissue, such as the gut. But if you are going to have a smaller gut, higher quality foods will be required to meet organismal energetic demands. These ideas logically lead to a dependence of such intelligent organisms on properties of the biota (presence and abundance of high-quality foods). Organisms require a viable ecological strategy to support energetically expensive encephalization. High-quality foods may present a more complex foraging problem to organisms, an issue explored in the following section. Consistent with the expensive tissue hypothesis, the squirrel monkey (genus Saimiri), which has a very large ratio of brain size to body mass (and very large social groups), also eats high-quality food (fruit) and has a smaller gut than comparably sized primates. Can we expect similar patterns in other lineages that evolve human-level intelligence? Fruit needs to exist for Saimiri to employ that ecological strategy to support its encephalization. Again, this scenario suggests some kind of dependence of animal intelligence on attributes of the primary trophic level. Plants predominate in biomass on the primary trophic level on terrestrial Earth and so attention should be paid to the role of plants in the evolution of intelligence.
Recent work points to a more general brain/energy trade-off than that outlined by Aiello & Wheeler (Reference Aiello and Wheeler1995). Organisms that evolve larger brains incur the energetic cost of this tissue, but offset this cost in a variety of ways not necessarily limited to gut tissue (expensive brain hypothesis; Isler & van Schaik Reference Isler and van Schaik2009). For example, encephalized organisms, rather than just possessing less gut tissue than less encephalized organisms, may instead (or as well) support less muscle tissue, or may possess lower annual fertility rates, longer birth intervals or slower growth. This broader set of offsets places the expensive brain concept within the realm of life-history theory.
v. Trophic facilitation: Human civilization is completely dependent on angiosperms (particularly the Poaceae, Solonaceae and Fabaceae; Diamond Reference Diamond1997). This means our civilization is dependent on a group of plants that did not exist 130 million years ago (much less 542 mya!). Does intelligence have similar biotic dependencies? If so, was intelligence less likely to evolve without a group such as angiosperms and the high-quality foods associated with them? The expensive tissue hypothesis could provide a mechanism for EQs dependence on developments in the primary trophic level of the biota.
Primates rely on edible plant resources. Fruiting tropical plants, because of their diversity, have complex temporal (seasonal) and spatial distributions. So plants present primates with a cognitive problem and an energetic supply (Milton Reference Milton1981). Much attention has been devoted to the dietary ecology of primates for clues about primate evolution and how a complex ecology can promote intelligence. Cenozoic terrestrial ecologies are the most diverse and complex that have ever existed on Earth, more diverse than Paleozoic and early Mesozoic ecosystems, in large part due to the angiosperm radiation (Knoll & Niklas Reference Knoll and Niklas1987; Russell Reference Russell and Seth Shostak1995; but see Peters & Foote Reference Peters and Foote2001). If ecosystem complexity and diversity has an important role in the evolution of encephalization (e.g. Milton Reference Milton1981), this would suggest that the likelihood of the evolution of intelligent life was much greater later in the Phanerozoic. Therefore, astrobiologists should be concerned with the evolution of ecosystem diversity and complexity (see hypothesis x). Attention should be directed to both derived encephalized lineages such as primates and also to basal organisms and prehistoric ecosystems in comparison with contemporary ecosystems.
Potential connections between trophic facilitation, the expensive tissue hypothesis and life-history theory are apparent. If intelligence is viewed as a property of an organism, then humans are simply one very rare organism of many, illustrating intelligence's rarity (Simpson Reference Simpson1964). However, the above hypotheses suggest an important role (in the evolution of encephalization) for the ecology of the organism in the context of its biota. Thus, evolution of intelligence might be thought of more as a property of a biota than a property of a species; this changes the probabilistic scenario. This view is consistent with the theme that humanity is part of the natural ecosystem and not an entity that dominates it (Haqq-Misra Reference Haqq-Misra2007).
To explore potential associations between EQ and trophic ecology, many questions can be asked. For instance, are high (or low) EQ organisms homogeneously distributed across global geographical environmental gradients? Or are these organisms in higher relative frequency in some kinds of environments? How do high EQ organisms fit into food webs? Some of these patterns are already apparent. More formal quantitative study could clarify the ecology of high EQ and help us better understand its evolution.
vi. General intelligence hypothesis: Intelligence is a controversial concept. The empirical rationale for the so-called unitary nature of intelligence derives from correlated performances by human and animal subjects on a wide variety of cognitive tasks. For example, an individual or individual species that performs well on one cognitive task will also tend to perform well on other kinds of cognitive tasks (Deaner et al. Reference Deaner, van Schaik and Johnson2006). Factor analysis reveals a single dimension of variation in performance on these cognitive tasks, termed the ‘g factor’ for general intelligence. ‘g’ is high or moderate and has been observed both within humans and between animal species (Deaner et al. Reference Deaner, van Schaik and Johnson2006; Lee Reference Lee2007). Brain size (EQ is one kind of brain size metric) moderately correlates with g within humans (Andreasen et al. Reference Andreasen, Flaum, Swayze, O'Leary, Alliger, Cohen, Ehrhardt and Yuh1993); importantly, correlations appear stronger when comparisons are made between species (Lee Reference Lee2007). In human inter-population comparisons, the g factor, intelligence and even their heritability may largely be explainable in terms of environmental causes, particularly through environmental feedbacks that amplify initially weak heritable differences (Dickens Reference Dickens2005 and references therein). However, inter-species intelligence and g variation are more likely to have a basis in inter-species genetic differences.
Importantly, the inter-specific (between species) g phenomenon may result from either correlated selection pressures or structural constraints. For example, it may be possible to generate a brain with very high long-term memory and very low spatial intelligence, but one does not tend to find organisms that possess such brains because selection does not tend to favour that combination of cognitive performances; instead one finds coincident positive (or coincident negative) selection for both cognitive processes. Alternatively, it may be difficult to produce a brain that bestows high performance on one kind of cognitive task without having spillover effects where performances on other cognitive tasks are improved. Or, once a species evolves high performance on one cognitive process, improving other cognitive processes may be sufficiently eased such that the benefits of generalized improvement of cognitive capacity outweigh the now lowered costs, and the species evolves.
Within this review there is an emphasis on EQ and intelligence. Intelligence, general intelligence and encephalization are three distinct concepts or quantities; whether relationships between g and EQ or intelligence and EQ generalize beyond vertebrates is an empirical question. The relationship between encephalization and intelligence does appear to generalize beyond vertebrates. Large meta-analyses that can determine the slope and strength of the relationship between encephalization and intelligence may not be currently possible with invertebrates, given the relatively low number of studies on invertebrates. However, the invertebrate literature does allow an assessment of the broader generality (existence beyond vertebrates) of the EQ/intelligence relationship, if not the generality of the strength or slope of this relationship. Within the insects, beetles with generalist feeding ecologies possess larger and more complex mushroom bodies (a structure of the insect nervous system) than other beetle species (Farris & Roberts Reference Farris and Roberts2005). Cephalopods are the most encephalized invertebrates and also display the most complex behaviour and learning among invertebrates (particularly Octopus vulgaris). When estimated, some cephalopod species have EQs comparable to some reptiles and mammals (Packard Reference Packard1972). Mather (Reference Mather2008), in considering the possibility of cephalopod consciousness, provides a nice overview of cephalopod behavioural research. These organisms display observational learning (Fiorito & Scotto Reference Fiorito and Scotto1992), spatial landscape memory (Mather Reference Mather1991) and problem-solving ability (Fiorito et al. Reference Fiorito, von Planta and Scotto1990). The brains of cephalopods are analogous (not homologous) to vertebrate brains and share some striking convergent features, but also display a distinct anatomy (Nixon & Young Reference Nixon and Young2003). Thus, the generality of an EQ/intelligence link is sustained even under the context of these gross anatomical differences.
The existence of g may have high significance for astrobiology; g suggests either that selection for one kind of cognitive processing has positive cognitive pleiotropies (positive side effects in other cognitive tasks), or that selection pressures for a variety of cognitive processes tend to be correlated in Nature. If the former is correct, the implication would be that selection for a wide variety of cognitive tasks would have positive spillover effects in other cognitive domains, presumably rendering high general intelligence a more likely evolutionary outcome. Astrobiologists should be aware of research on g, particularly as attempts are made to generalize g beyond the human species.
Alternative views exist, which challenge the idea that intelligence can be meaningfully described as a singular or even real phenomenon (see Gould Reference Gould1981). Astrobiological extrapolation may be more straightforward with a unitary concept of intelligence or a high g factor, but a non-unitary view of intelligence would not preclude astrobiological extrapolations; instead the latter view simply implies a broader definition of intelligence and an expectation for more variety in the manifestations of intelligence.
vii. Brain evolution hypothesis: Many hypotheses listed here treat the nervous system as a black box influenced by ecological factors and evolutionary pressures that drive the allocation to nervous tissue. Instead, the rate of evolution of intelligence may not be limited by selection pressures, but by the rate that structures and functions of the brain are generated by mutation. The evolution of any number of internal factors ranging from structural elements to underlying genetic systems could influence the long-term rate of brain evolution. For example, the accumulation of the major morphological structures in the brain may both provide some kind of essential function for intelligence (Sagan Reference Sagan1977) and may require time. Comparative anatomy, behaviour and genetics are the means to evaluate the brain evolution hypothesis. Some researchers are using simulation, simulated environments and neural networks to evaluate brain evolution (e.g. Yaeger Reference Yaeger2009). Additionally, selection experiments on allocation to central nervous system tissue can reveal responsiveness to and behavioural consequences of such selection. Very few lineages have been evaluated in this way, due to both practical and ethical considerations. Selection experiments on learning ability have been performed on Drosophila, showing some responsiveness to selection for learning ability (e.g. Mery & Kawecki Reference Mery and Kawecki2002). Extrapolation of these results to other animal phyla, longer time scales and larger phenotypic changes is tentative. Longer term studies would be necessary to get a good sense of the longer term evolvabilitiesFootnote 3 of lineages.
Questions regarding intelligence evolution fit within larger fundamental evolutionary questions. To what extent is the pace of phenotypic evolution set by limits to evolvability versus a lack of selection pressures? Articulated in the context of intelligence evolution the above question may read like the following. With respect to intelligence and encephalization, a biota's failure to produce a highly encephalized organism at time t might be attributed to genetic or phenotypic limitations in existing organisms or a lack of appropriate selection pressures. The evolution of intelligence may require an accumulation of structural or genetic diversity associated with the nervous system. If true, derived organisms possessing such features would be expected to respond to selection for general intelligence, while organisms without such features would not. How long has the latent potential for intelligence existed on Earth?
viii. Contingency hypothesis: Many evolutionary outcomes within lineages are contingent on earlier outcomes or events. If these contingent circumstances are unpredictable, idiosyncratic, occur chaotically or are rare, then evolutionary outcomes that require these preconditions will be equivalently unpredictable. The path to intelligence is argued to be best described by this pattern of evolution (Gould Reference Gould1989; Via Reference Via2001). Simpson (Reference Simpson1964) makes a similar argument for the evolution of humans. Lineweaver (Reference Lineweaver, Seckbach and Walsh2008) argues that intelligence is not a convergent feature of evolution. Contingent events can be endogenous to an organism (e.g. whether or not an organism evolves a feature) or exogenous to an organism, such as a mass extinction event.
The contingency hypothesis predicts that intelligence may have evolved much earlier in the Phanerozoic, much later, or it may not have evolved at all (Gould Reference Gould1989; Via Reference Via2001). Advocates of a contingency view often further argue that because of underlying chaotic processes, the nature of the outcome of evolution, what intelligence ‘looks’ like and the kinds of organisms that possess it, is inherently unpredictable. Organisms such as cetaceans appear to demonstrate that the kinds of organisms that can evolve intelligence are diverse. At the same time, the existence of multiple lineages in multiple ecosystems with high intelligence and high EQ suggests that these features are both likely and likely to arise in many contexts. Superficial differences between such diverse lineages may belie commonalities in their ecology and life-history.
Lineweaver (Reference Lineweaver, Seckbach and Walsh2008) rightly argues that convergence, which is often thought of as opposing contingency, must always be scaled by the evolutionary distance between lineages. Even cetaceans, cephalopods and primates share a common ancestor that already possessed some key adaptations related to the nervous system. Thus their independent evolution of large brains cannot be regarded as entirely independent. However, if a key adaptation only evolves once, this is evidence only that the adaption didn't evolve twice, not that it couldn't have evolved twice. Rapid adaptive radiations are thought to frequently follow (and be initiated by) the acquisition of a key adaptation. Through ecological incumbency, such an adaptive radiation could prevent additional cases of evolution of that key adaptation in other lineages. An implicit idea in the contingency hypothesis is that lineages (and by extrapolation an entire biota) are strongly constrained by the key adaptations or features which did or did not evolve in their past. The strength and importance of contingency arguments depend powerfully on the evolvability of lineages. If at least some lineages have high evolvability, such constraints are likely to be overcome or a suitable key adaptation is likely to arise, allowing diverse phenotypic potentials including intelligence. There are plenty of examples of evolutionary convergence reflecting this evolvability (see Conway-Morris references herein). For example, myelin appears to have evolved independently in vertebrates and multiple invertebrate lineages (Hartline & Colman Reference Hartline and Colman2007). Alternatively, if most lineages have low evolvability in the sense discussed above, Lineweaver's (2008) arguments may be quite consequential for astrobiology.
Evidence for the contingency hypothesis tends to be anecdotal. This is justified because anecdotal or peculiar events are viewed as integral to observed evolutionary outcomes because these events constrain future potential. Peculiar events have occurred in our ancestry. If the human line were to have gone extinct some time in the last 100 000 years the pattern of intelligence evolution would have been altered. Alternatively, if multiple pathways exist for the evolution of intelligence then historical events might dictate which lineage evolves intelligence, but might not prevent the eventual evolution of some intelligent lineage regardless of its position on the tree of life. Contingency may be important for individual species, but this important role for contingency may not necessarily extrapolate to the biota as a whole. The diffusion hypothesis makes this clear; if one lineage due to highly idiosyncratic reasons fails to evolve high EQ or goes extinct, other lineages are potentially available to do so. The evolutionary motion of multiple lineages can be more predictable than single lineages.
ix. Step hypothesis: The step hypothesis treats the evolution of intelligent life as requiring several sequential steps (Carter Reference Carter1983; Watson Reference Watson2008). The likelihoods of the steps are usually modelled as low and not changing over time. Watson's (Reference Watson2008) model contains seven steps such as ‘prokaryotes to eukaryotes’, among others. The last two steps in Watson's model are the evolution of multicellular life and the evolution of intelligence. Thus the model pertains to a wider time scale than the Phanerozic, the time scale of interest herein, though the last steps of step model do overlap with this eon. In principle, additional steps could be included in a similar model. For example, McKay (Reference McKay and Doyle1996) using a similar ‘step perspective’ included the development of animals, land ecosystems and animal intelligence as additional steps along with the evolution of multicellularity and the evolution of human intelligence. The step hypothesis is similar to the contingency hypothesis (hypothesis viii) but has thematic differences. The path to intelligence in both is treated as a process of passing through a series of (usually unlikely) events. In the step hypothesis, emphasis is placed on roughly known events that are considered necessary for further evolution towards intelligent species. On the other hand the contingency hypothesis stresses unknown or idiosyncratic events and stresses the unpredictability of the nature of the organisms resulting from the process.
The step hypothesis has received attention from astrobiologists (Carter Reference Carter1983; McKay Reference McKay and Doyle1996; Watson Reference Watson2008). Proponents have identified its weaknesses. Watson's (Reference Watson2008) model simplifies the evolution of intelligent life by modelling the probabilities of each step in the chain to intelligent life as remaining low and constant over time, making the problem easier to solve analytically. The alternative to the step hypothesis, described as the ‘long-fuse’ hypothesis, suggests intelligence requires time and increases in probability through processes that are incremental and cumulative. Several of the hypotheses reviewed here strongly point to the latter scenario rather than the step hypothesis (e.g. hypotheses i–vi and x). For example auto-catalytic hypotheses propose that the probability of evolving a species with a certain EQ in the next time-step is dependent on the EQ of contemporary organisms and the selective pressures they engender in the ecosystems of the biota. In principle, the model developed by Watson (Reference Watson2008) could be applied to any kind of event (biological or not) dependent on a sequence of rare events. The only input from biological fields would be the identification of relevant rare events and their probability. The ‘long fuse’, on the other hand, would lead to a deeper involvement of evolutionary, genetic, behavioural, neurological and ecological fields into the resolution of the question of intelligent life in the universe. Many hypotheses reviewed herein pertain only to the last step of Watson's model; the step model is valuable whether or not a long-fuse model is appropriate for the last critical step (the evolution of intelligence).
x. Internal variance principle or zero-force evolutionary law: New ideas concerning the evolution of complexity are emerging. The internal variance principle (IVP) challenges our notions of null models of the evolution of complexity (McShea Reference McShea2005). The IVP proposes that statistical constraints force increases in organismal complexity through the differentiation of an organism's internal repeated parts, prior to the action of natural selection. In other words, the IVP suggests that increases in internal organismal differentiation, a form of complexity, should be the null expectation. The IVP works in the following manner. An organism's internal parts, whether they are genetic or morphological, tissues, cell types or segments are thought to be constrained to differentiate simply because there are more possible states in which the various parts are different from each other than states in which they are all the same. Thus in the absence of counteracting selection for similarity of parts, differentiation is bound to increase. This concept predicts functionally neutral increases in diversity of organismal components (complexity), disentangling complexity from function. However, complexity that evolves neutrally through this principle can later become functionally relevant through evolutionary exaptation: the re-purposing of traits or attributes from one function (or no function) to a new functionFootnote 4. An explication of the abstract IVP concept can be found in McShea (Reference McShea2005) and McShea & Brandon (Reference McShea and Brandon2010).
The evolution of complexity is not synonymous with the evolution of encephalization, although there are reasons to expect links between encephalization, nervous system complexity, general organismal complexity and the IVP. The IVP provides a neutral (or function independent) mechanism for constrained differentiation of organismal parts. Despite the neutral nature of this principle on a proximate/immediate time scale, long-term functional evolutionary consequences could emerge. Differentiated neural parts or genetic control mechanisms would plausibly be more responsive to subsequent natural selection than undifferentiated neural parts, and thus selection for greater encephalization would be more effective in terms of behavioural and cognitive consequences (a testable hypothesis). On the other hand, morphologically differentiated organisms may present an opportunity for neuronal specialization and growth to control these differentiated organismal features.
McShea & Brandon (Reference McShea and Brandon2010) have reformulated the IVP as a process that is ubiquitous in biological systems operating throughout the biological hierarchy, at molecular and ecosystem scales as well as at organismal scales. They have termed this process the zero-force evolutionary law (ZFEL; I suggest ‘differentiation principle’ might be an appropriate term as well). The zero-force term refers to the expectation of a tendency for increasing complexity whether or not other factors, such as natural selection or constraints, are present. Ecosystems such as organisms have component parts that can differentiate. Hence ecosystems are susceptible to the ZFEL/IVP's ‘zero-force’ drive for higher complexity. The ZFEL could operate in concert at organismal and ecosystem levels. Angiosperms are highly diversified components of terrestrial ecosystems, and their fruits and flowers are simply differentiated leaves. In other words, angiosperms diversified within ecosystems, and they did so in part by diversifying their own internal component structure. A complex ecosystem as detailed earlier can promote the evolution of encephalization (hypothesis v, Milton Reference Milton1981). This represents another potential link between IVP/ZFEL-driven complexity and encephalization.
The IVP/ZFEL concept in part represents just a synthesis of existing biological notions regarding variation; however, this synthesis brings clarity to the implications of how biological variation is generated, shedding light on the omnipresent tendency of biological systems to spontaneously diverge and diversify, thereby increasing complexity.
Discussion
Understanding microevolutionary mechanisms such as natural selection does not translate straightforwardly to a knowledge of life's long-term trajectories, pacing and patterns. Estimates for timing and likelihood of intelligent life vary widely. McKay (Reference McKay and Doyle1996) estimates that human-level intelligence could evolve, at the earliest, only ∼20 million years after the evolution of multicellularity. Ward & Brownlee (Reference Ward and Brownlee2000) argue that intelligence is highly unlikely. Do these wide estimates reflect genuinely wide potential evolutionary outcomes or uncertainty in our knowledge? Given the multiple currently active realms of ecology and evolution, whose unanswered questions could directly impact our understanding of intelligent life, I think we should recognize that we do not know the likelihood of intelligent life or how chaotic or deterministic its evolution may be. However, we should not be discouraged. Like chemistry or physics, biology can have universal principles (Chela-Flores Reference Chela-Flores2007). Universal principles are expected to operate elsewhere in the universe beyond Earth and can be usefully contrasted with historical contingency (unless historical contingency itself were to be thought of as a universal principle). Many astrobiologists are already confident enough to enshrine evolution by natural selection as one of these universal principles. The phenomenon of natural selection and its expectation of universality were derived from the study of Earth's life. Therefore, the study of Earth's life using rigorous scientific methods (empirical, theoretical, simulation) could lead to the inference of universal biological principles relevant for the evolution of intelligence. Universal biological principles might not be limited to microevolutionary processes such as natural selection. For example, diffusion dynamics (hypothesis i) may represent a macroevolutionary universal principle. Whether or not convergence, diffusion dynamics, contingency, IVP/ZFEL or other principles have the same expectation of universality as natural selection will be enlightened by a deeper understanding of these principles and their action on Earth's life. Thus, intelligent life's likelihood, time frame and pattern of evolution represent regular scientific research problems.
Several potential universal principles are not neutral on the question of intelligence's likelihood or inevitability. The diffusion hypothesis (hypothesis i) suggests that an intelligent species will eventually evolve whether the path to intelligence is choppy or smooth (Figs 1 and 2). Regularity of pattern of EQ or intelligence increase and predictability in timing are not required for inevitability in outcome. Diffusion dynamics therefore represents a theoretical reason to expect an intelligent organism to eventually evolve within a biota, provided the allowable time scale is long enough for diffusion processes to explore higher intelligence phenotypes.
Conclusion
The literature on the evolution of intelligence particularly as it pertains to humans does not suffer from a lack of ideas. Some hypotheses (contingency hypothesis, hypothesis viii) would suggest that the evolution of intelligent organisms may depend on idiosyncratic events. Thus the evolution of such organisms may be unpredictable, or, worse, their ultimate causes may lie hidden in an unknowable morass of factors and events. However, other hypotheses and largely overlooked data suggest otherwise. There are several theoretical reasons to support increasing maximum intelligence in a biota (hypotheses i, ii, v and x). Therefore, research into the evolution of intelligence should be addressed with vigour, both through continued examination of empirical data and through simulation and theory. The relevant questions range broadly into diverse disciplines. I suggest a critical evaluation of the Russell data set as a starting point. A distinction should be made between elucidating rules and patterns on our planetary system versus extrapolation of these discoveries to other systems. Progress in the former should aid in the latter. However, it will be difficult to know beforehand whether what we learn about the evolution of intelligent life on Earth can be extrapolated to other systems, because the ability to extrapolate will depend on the nature of the answers we get to the above questions.
Acknowledgements
Many individuals assisted me with this paper either through conversation or by reading sections of the manuscript: Mary Damm, Maxine Watson, Daniel McShea, Elizabeth Koziol, Aditya Chopra, Charles Lineweaver, Larry Yaeger, Kevin Hunt, Michael Muehlenbein, Andrew Watson, Harry Jerison, Jim Goodson, and two anonymous reviewers. I got feedback from Abgradcon2009 attendees and from the Indiana University History and Philosophy of Biology BSRG reading group.
Author disclosure statement
No competing financial interests exist.