INTRODUCTION
A central palaeoecological paradigm has long been that there was an alternation of glacial and interglacial biota via large-scale migrations, where whole communities shifted to southern refugia during glacial periods and northward during interglacial periods (de Lattin, Reference de Lattin1967). Increasing interest in the location of such refugia for temperate species during the harshest part of the last glacial period—the last glacial maximum (LGM)—has been evident during the last two decades (e.g., Pfenninger et al., Reference Pfenninger, Posada and Magnin2003; Pinceel et al., Reference Pinceel, Jordaens, Pfenninger and Backeljau2005; Kotlík et al., Reference Kotlík, Deffontaine, Mascheretti, Zima, Michaux and Searle2006; Magri et al., Reference Magri, Vendramin, Comps, Dupanloup, Geburek, Gomory and Latalowa2006; Sommer and Nadachowski, Reference Sommer and Nadachowski2006; Feuerdean et al., Reference Feurdean, Wohlfarth, Björkman, Tantau, Bennike, Willis, Farkas and Robertson2007; Bhagwat and Willis, Reference Bhagwat and Willis2008; Provan and Bennett, Reference Provan and Bennett2008; Juřičková et al., Reference Juřičková, Horáčková and Ložek2014a; Jamrichová et al., Reference Jamrichová, Potůčková, Horsák, Hajnalová, Bárta, Tóth and Kuneš2014b; Peterson and Graves, Reference Peterson and Graves2016). The idea that species survived elsewhere than they recently occur, usually in the warmer margins of Europe (Huntley and Birks, Reference Huntley and Birks1983), has been tested using phylogeography (e.g., Kotlík et al., Reference Kotlík, Deffontaine, Mascheretti, Zima, Michaux and Searle2006) and/or direct fossil evidence (e.g., Juřičková et al., Reference Juřičková, Horáčková and Ložek2014a). Palaeontological records can give detailed and precise information about past distributions of species, but finding them in suitable areas is largely a matter of luck. Genetic data are easily available, but their interpretation is based on the analysis of modern populations at current locations and may be affected by past bottlenecks when the majority of populations became extinct, leaving only part of the genetic diversity preserved.
This paradigm has been called into question by the finding of so-called northern or cryptic refugia, mostly in the Alps and Carpathians (for a review, see Schmitt and Varga, Reference Schmitt and Varga2012). Both fossil and genetic evidence have indicated the survival of some temperate vertebrates (e.g., Horáček, Reference Horáček2006; Kotlík et al., Reference Kotlík, Deffontaine, Mascheretti, Zima, Michaux and Searle2006; Sommer and Nadachowski, Reference Sommer and Nadachowski2006; Ursenbacher et al., Reference Ursenbacher, Carlsson, Helfer, Tegelström and Fumagalli2006), snails (Pfenninger et al., Reference Pfenninger, Posada and Magnin2003; Pinceel et al., Reference Pinceel, Jordaens, Pfenninger and Backeljau2005; Juřičková et al., Reference Juřičková, Horáčková and Ložek2014a), and plants (Birks, Reference Birks2003; Magri et al., Reference Magri, Vendramin, Comps, Dupanloup, Geburek, Gomory and Latalowa2006; Jankovská and Pokorný, Reference Jankovská and Pokorný2008) during glacial periods in central Europe. But this is not only the case in Europe; for example, New Zealand temperate forest trees were widespread but scarce during the LGM (McGlone, Reference McGlone1983). Similarly, modelling by Snell and Cowling (Reference Snell and Cowling2015) and Peterson and Graves (Reference Peterson and Graves2016) shows that the rapid migration rates of temperate plant species in North America can be explained only as a combination of spread from southern refugia and the existence of northern refugia. Higher latitudes were previously regarded as being inhospitable. Cryptic refugia of temperate species situated there certainly did not cover large areas. The survival of temperate species might have been possible in microrefugia of favourable microclimates (Rull, Reference Rull2009; Dobrowski, Reference Dobrowski2011) within harsh areas, thus enabling temperate species to survive, scattered or at low densities but nevertheless remaining present.
There are two hypotheses for how temperate taxa survived glacial periods in central Europe: either in refugia in southern European mountain areas or in broader cryptic refugia of scattered populations situated farther north. These hypotheses, however, should probably be seen as endpoints of a continuum (Provan and Bennett, Reference Provan and Bennett2008).
Tzedakis et al. (Reference Tzedakis, Emerson and Hewitt2013) critically evaluated this debate, highlighting that the polarization of refugia into northern and southern is confusing, and they preferred to study the northernmost borders of species ranges during the LGM and postglacial colonization dynamic. They questioned the occurrence of temperate trees north of 45°N but accepted a west–east asymmetry in boreal tree distribution. While western Europe was treeless north of 46°N during the LGM, restricted boreal populations persisted in eastern Europe up to 49°N.
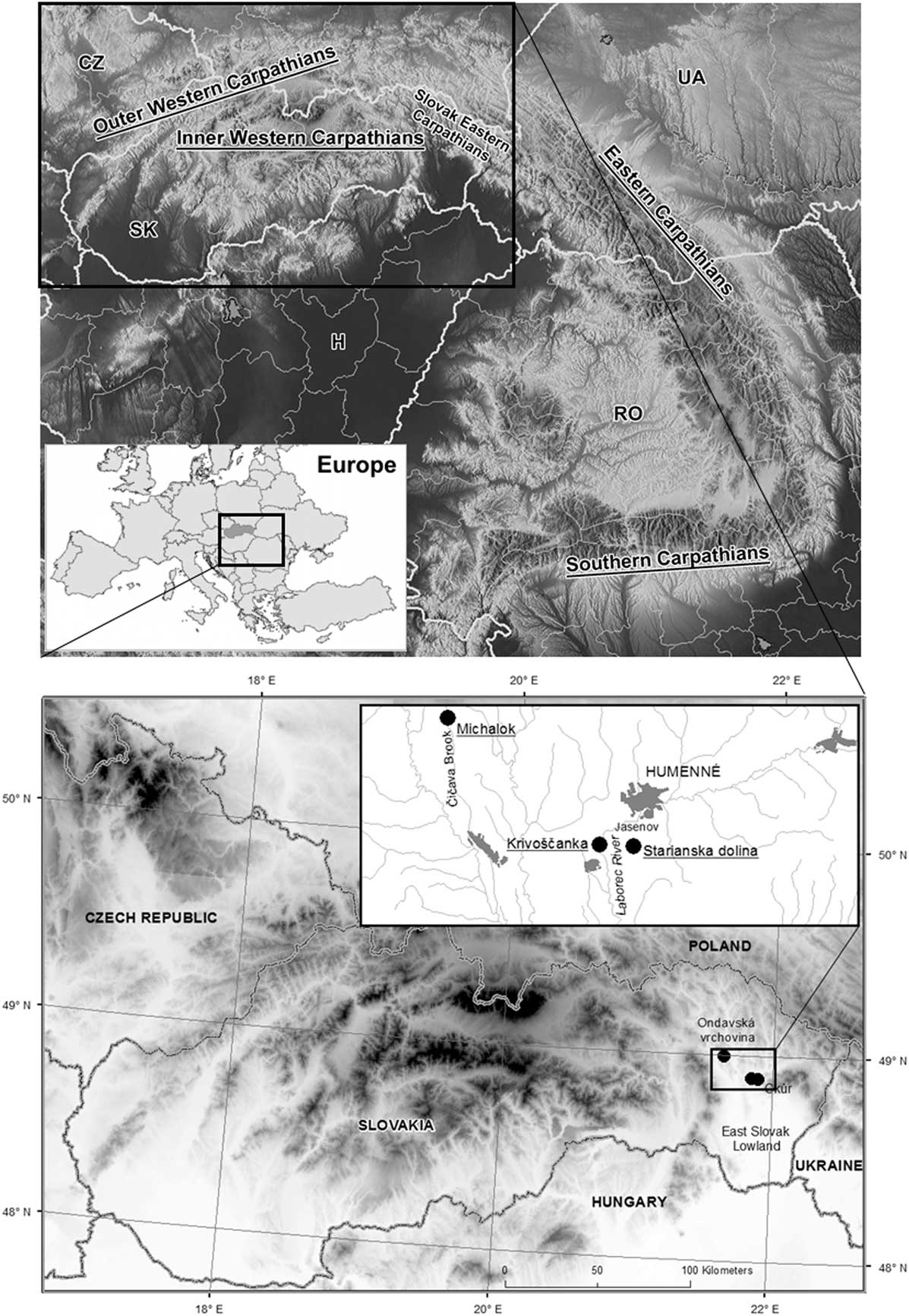
The Carpathian mountain belt (Fig. 1) can be divided into three different and isolated geological parts: the western, eastern, and southern Carpathians, each of which differs in its recent unique fauna and flora as well as natural development (Ronikier, Reference Ronikier2011). The Carpathian Mountains are considered an important biodiversity hot spot (e.g., Feurdean et al., Reference Feurdean, Wohlfarth, Björkman, Tantau, Bennike, Willis, Farkas and Robertson2007; Ronikier, Reference Ronikier2011; Mráz and Ronikier, Reference Mráz and Ronikier2016; Jamrichová et al., Reference Jamrichová, Petr, Jimenéz-Alfaro, Jankovská, Dudová, Pokorný and Kołaczek2017) with a high level of local endemism (Bálint et al., Reference Bálint, Ujvárosi, Theissenger, Lehrian, Mészáros and Pauls2011; Barkasi, Reference Barkasi2016; Kliment et al., Reference Kliment, Turis and Janišová2016), a high level of local genetic diversity in some species (Kotlík et al., Reference Kotlík, Deffontaine, Mascheretti, Zima, Michaux and Searle2006; Wielstra et al., Reference Wielstra, Babik and Arntzen2015), and the presence of fossil records of temperate species during glacial periods (Ložek, Reference Ložek2006; Jankovská and Pokorný, Reference Jankovská and Pokorný2008; Juřičková et al., Reference Juřičková, Horáčková and Ložek2014a). Knowledge of the Quaternary development of Carpathian biota can shed light on the present zoogeography and species diversity of this mountain range, regional climatic changes, and the timing and response of these faunas to regional climatic changes.
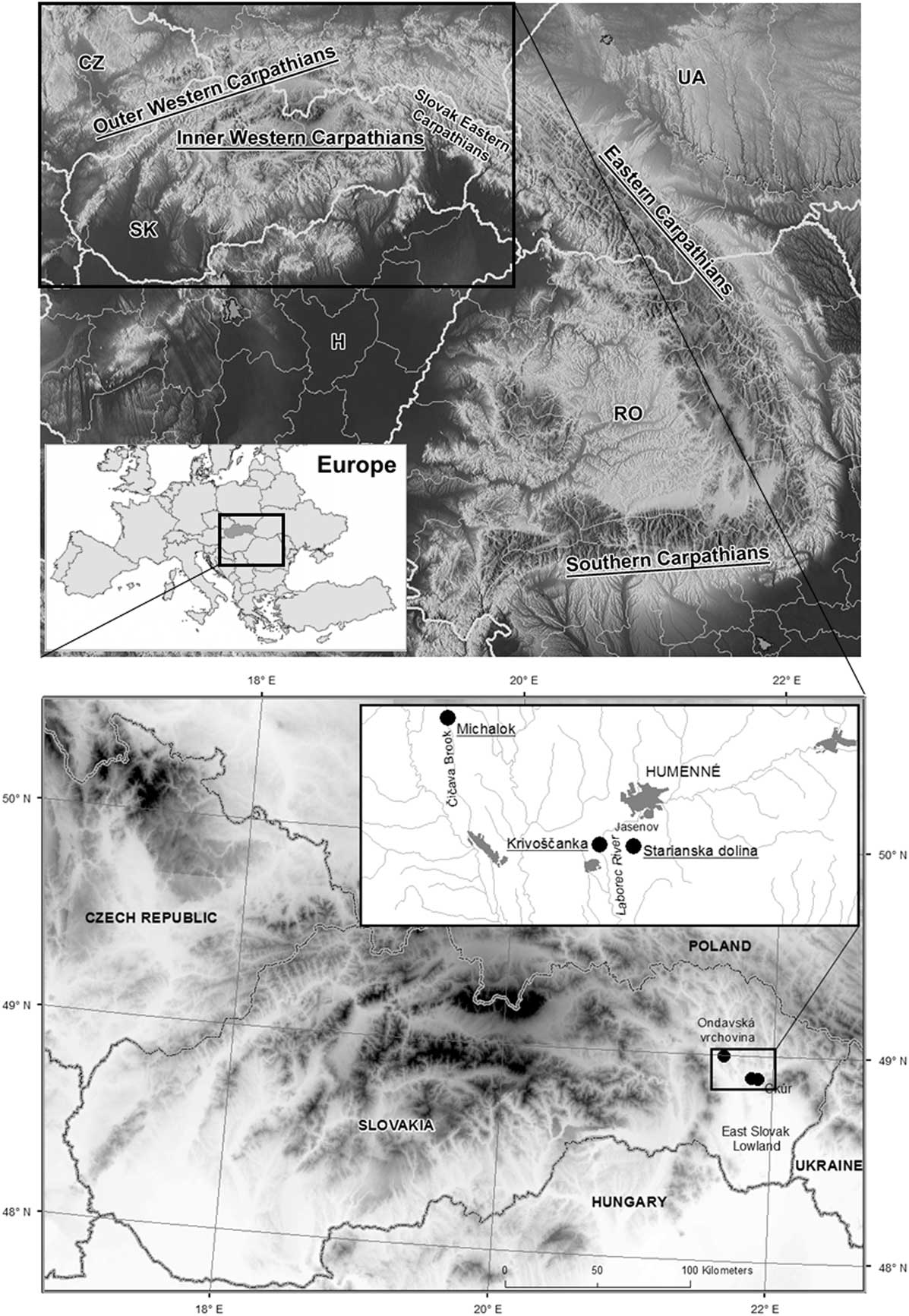
Figure 1 Upper map: location of the study area in Europe and in the Carpathian Mountains. CZ, Czech Republic; H, Hungary; RO, Romania; SK, Slovakia; UA, Ukraine. Lower map: situation of the studied successions in the Slovak eastern Carpathians. Higher altitudes are marked in black, lower in white.
Unfortunately, information on postglacial faunal and floral successions remains sparse and uneven for these particular Carpathian regions. Probably the best data exist from the western Carpathians, which have been demonstrated to be the northern glacial refugium of many plants (Jankovská and Pokorný, Reference Jankovská and Pokorný2008; Jamrichová et al., Reference Jamrichová, Potůčková, Horsák, Hajnalová, Bárta, Tóth and Kuneš2014b, Reference Jamrichová, Petr, Jimenéz-Alfaro, Jankovská, Dudová, Pokorný and Kołaczek2017) and animals (Ložek, Reference Ložek2006; Kotlík et al., Reference Kotlík, Deffontaine, Mascheretti, Zima, Michaux and Searle2006; Juřičková et al., Reference Juřičková, Horáčková and Ložek2014a). An important network of palaeobotanical records is also known from the Romanian part of the eastern Carpathians, documenting that some coniferous and broadleaf trees were present there before 14,700 cal yr BP (Feurdean et al., Reference Feurdean, Wohlfarth, Björkman, Tantau, Bennike, Willis, Farkas and Robertson2007, Reference Feurdean, Klotz, Brewer, Mosbrugger, Tămaş and Wohlfarth2008a, Reference Feurdean, Klotz, Mosbrugger and Wohlfarth2008b, Reference Feurdean, Spessa, Magyari, Willis, Veres and Hickler2012). The more northerly situated Slovak part of the eastern Carpathians, in particular, remains poorly understood with respect to both its phylogeography and palaeoecology. Except for two sites of tufaceous fens with typical wetland fauna and flora of subrecent age (Hájková et al., Reference Hájková, Horsák, Hájek, Lacina, Buchtová and Pelánková2012; Jamrichová et al., Reference Jamrichová, Hájková, Horsák, Rybníčková, Lacina and Hájek2014a) and one site of lake sediments, where temperate forest trees appeared at the very beginning of the Holocene (Hájková et al., Reference Hájková, Pařil, Petr, Chattová, Grygar and Heiri2016), the late glacial and Holocene biota of the Slovak eastern Carpathians remains virtually unknown. Nevertheless, these lower mountains in the westernmost part of the eastern Carpathians, rising above the east Slovak lowlands, have been shown to be an important biogeographic boundary (e.g., Zemánek, Reference Zemanek1991; Ronikier, Reference Ronikier2011; Mráz and Ronikier, Reference Mráz and Ronikier2016; Šrámková-Fuxová et al., Reference Šrámková-Fuxová, Záveská, Kolář, Lučanová, Španiel and Marhold2017). In particular, the Slovak eastern Carpathians seem to be an important zoogeographic boundary for modern molluscan species ranges. The easternmost ranges of many central European and western Carpathian species end just before the eastern Carpathian foothills, and many eastern Carpathian species have their westernmost boundary there (Kerney et al., Reference Kerney, Cameron and Jungbluth1983; Welter-Schultes, Reference Welter-Schultes2012). The specific character of the Slovak eastern Carpathians molluscan fauna also supports high local species endemism (Horsák et al., Reference Horsák, Juřičková and Picka2013).
We have no information so far about shifts in these species ranges during the late glacial period and Holocene. Such shifts in the species ranges may depend on local, terrain-modified mesoclimatic changes slightly differing from general trends and may more realistically reflect how the species spread. Holocene land snail assemblages are a useful proxy for studying this relatively narrow boundary. Molluscan shells are usually deposited where the animal actually lived, unlike pollen or vertebrate fossils. Moreover, shells can be identified to the species level, which is essential for studies of local endemism. The most commonly used palaeoecological proxy, pollen, is only partly identifiable to a comparably exact taxonomic level (e.g., Ložek, Reference Ložek1964, Reference Ložek2000; Evans, Reference Evans1972; Sümegi, Reference Sümegi2005; Davies, Reference Davies2008).
We present here the three late glacial and Holocene molluscan successions, two of them from the westernmost part of the Slovak eastern Carpathians, and one from the easternmost part of the western Carpathians, to elucidate the palaeoecological characteristics of this zoogeographic boundary.
MATERIALS AND METHODS
Site characteristics
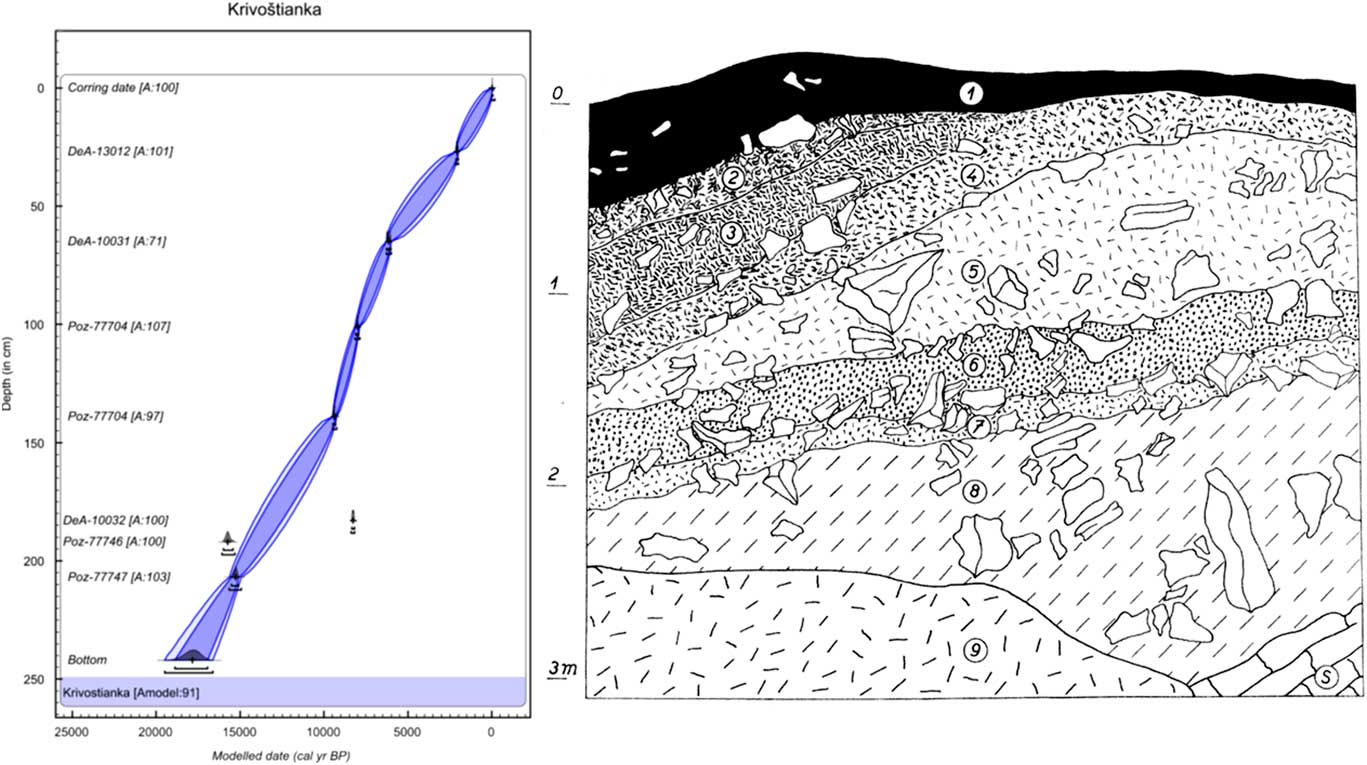
We analysed the three molluscan successions from eastern Slovakia (Fig. 1) in this study. Krivoštianka, Okúr (48°53′28.136″N, 21°50′31.384″E; 175 m asl), is a profile through 3.5-m-thick colluvium at the westernmost split of the Krivoštianka Ridge called Okúr (Humenné vrchy Mountains) projecting into the great east Slovak lowland above the Laborec River (138 m asl). The depositional sequence is located on a west-facing, steep rocky slope of that ridge built by Mesozoic limestones (for lithology, see Fig. 2). This is reflected in the recent vegetative cover, which includes xerothermophilous shrubland with patches of rocky steppe in the immediate neighbourhood of oak–hornbeam and submontane beech forests. These woodlands sharply border the deforested plain of the eastern Slovak lowland. The oldest loess layer, which is not associated with the rest of sampled sediments, originated from this border.

Figure 2 (color online) Lithology and age–depth model of the colluvial deposit at Krivoštianka Ridge, Okúr in the Humenné vrchy Mountains above the Laborec River. Model was made in OxCal v4.3.2 (Bronk Ramsey, Reference Bronk Ramsey2017) with InCal 13 atmospheric curve (Reimer et al., Reference Reimer, Bard, Bayliss, Beck, Blackwell, Bronk Ramsey, Buck, Cheng, Edwards, Friedrich, Grootes, Guilderson, Haflidason, Hajdas, Hatté, Heaton, Hoffman, Hogg, Hughen, Kaiser, Kromer, Manning, Niu, Reimer, Richards, Scott, Southon, Staff, Turney and van der Plicht2013). Two radiocarbon dates (presented in models only as outliers) were excluded from the final model to obtain a reliable Bayesian model with the maximum possible agreement value as shown at the bottom of all three models (Amodel = age-depth model agreement in %). The laboratory codes of all radiocarbon dates used in the models are in accordance with Table 1. Numbers of layers indicate the centre of the sample. 1, greyish-black humic loam with scattered scree fragments (rendzina soil); 2, brownish-grey, slightly humic loam, richer in scree; 3, fine rubble with a loose, slightly humic loamy matrix and numerous shells; 4, greyish-brown loam, rich in fine to medium-coarse scree, partly compacted; 5, pale greyish-brown compact loam, rich in medium-coarse scree with scattered blocks; 6, brownish-grey humic crumbly loam, rich in rubble with coarser fragments; 7, pale brownish-grey loam, rich in scree; 8, pale greyish-brown (some loess) loam with scree of all sizes, including coarse blocks; 9, talus; S, limestone bedrock.
Table 1 Results of accelerator mass spectrometry radiocarbon age determination.Footnote a
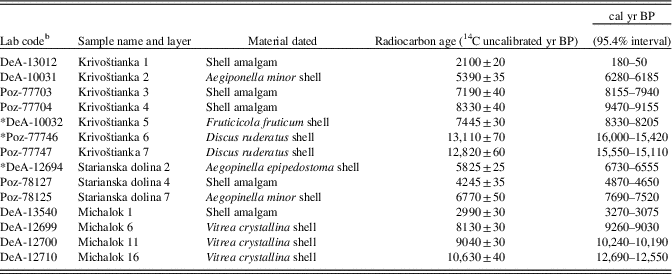
a Calibration was performed in OxCal v. 4.3 (Bronk Ramsey Reference Bronk Ramsey2009a). All ages are rounded according to Stuiver and Pollach (Reference Stuiver and Pollach1977).
b DeA, laboratory code of Izotoptech Zrt., Debrecen, Hungary; Poz, laboratory code of Poznań Radiocarbon Laboratory. Radiocarbon dates preceded by an asterisk (*) were excluded from the age–depth models.
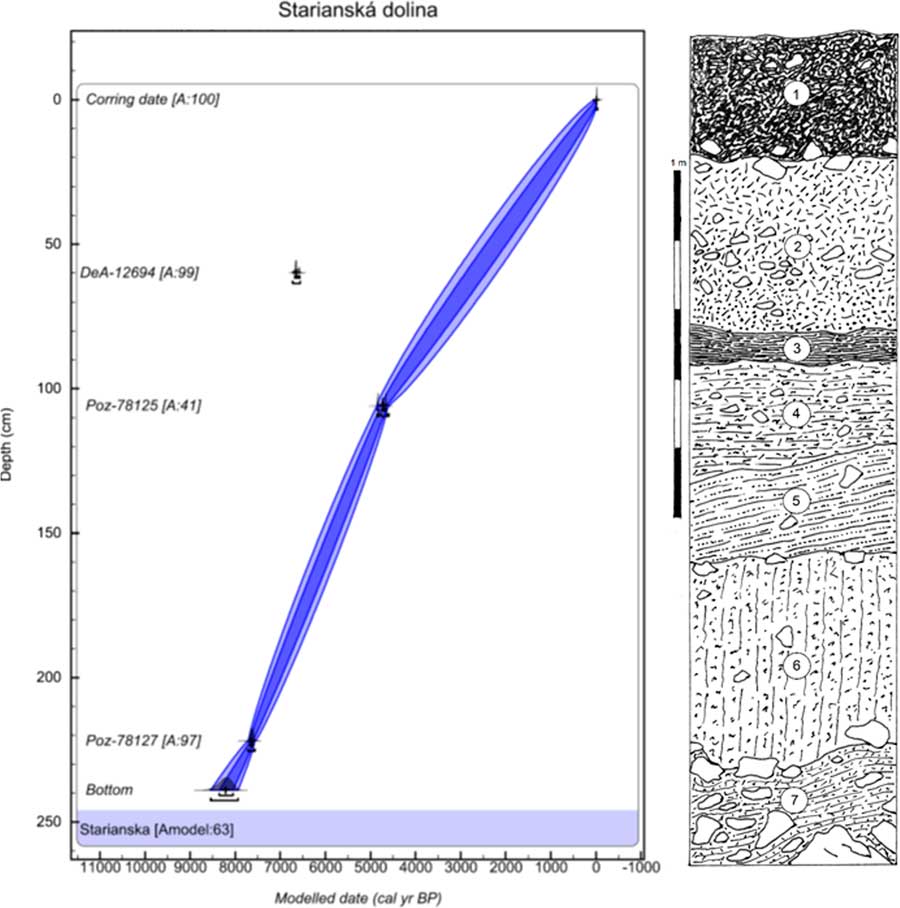
The next profile is in the Starianska dolina Valley near Jasenov (48°53′25″N, 21°53′36″E; 320 m asl), a short valley incised in the north-facing side of the Krivoštianka Ridge 1.2 km west of Jasenov Castle near Humenné in the Humenné vrchy Mountains. The site is situated in the middle of the ridge, where the valley bottom, filled by slopewash loams and limestone scree with a tufa admixture, is cut at a gully by a trench that exposes the sedimentary sequence (Fig. 3). The surrounding slopes are covered by calciphilous and submontane herb-rich beech forest with local linden–maple stands on talus screes.
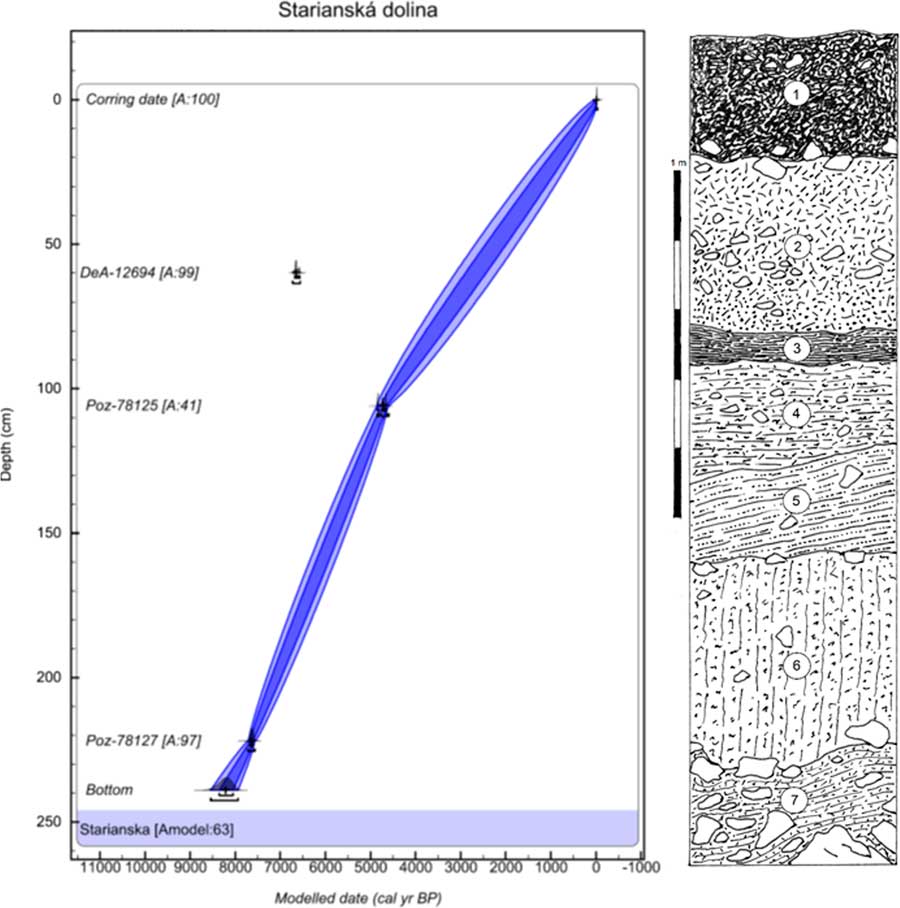
Figure 3 (color online) Lithology and age–depth model of the Starianska dolina Valley near Jasenov Castle in the Humenné vrchy Mountains. Model was made in OxCal v4.3.2 (Bronk Ramsey, Reference Bronk Ramsey2017) with InCal 13 atmospheric curve (Reimer et al., Reference Reimer, Bard, Bayliss, Beck, Blackwell, Bronk Ramsey, Buck, Cheng, Edwards, Friedrich, Grootes, Guilderson, Haflidason, Hajdas, Hatté, Heaton, Hoffman, Hogg, Hughen, Kaiser, Kromer, Manning, Niu, Reimer, Richards, Scott, Southon, Staff, Turney and van der Plicht2013). One radiocarbon date (presented in models only as an outlier) was excluded from the final model to obtain a reliable Bayesian model with the maximum possible agreement value as shown at the bottom of all three models (Amodel value in %). The laboratory codes of all radiocarbon dates used in the models are in accordance with Table 1. Numbers of layers indicate the centre of the sample. 1, dark brownish-black, humus-rich crumby topsoil, scattered limestone fragments, coarser scree at the base; 2, reddish-brown, rather small-sized (5 to 10 cm) limestone scree with a coarse-granular incoherent limestone matrix dominating at the base; 3, greyish-black, moderately clayey humic loam with scattered fine fragments and sporadic charcoals; 4, greyish-brown, slightly humic calcareous loam, scattered small fragments, and tufa particles; 5, brown, highly calcareous loam, fine limestone fragments, tufa admixture; 6, yellowish-brown calcareous loam with fine rubble and scattered tufa particles; 7, coarse limestone scree with fine angular rubble in interstices and a greyish pale-brown compacted loamy matrix.
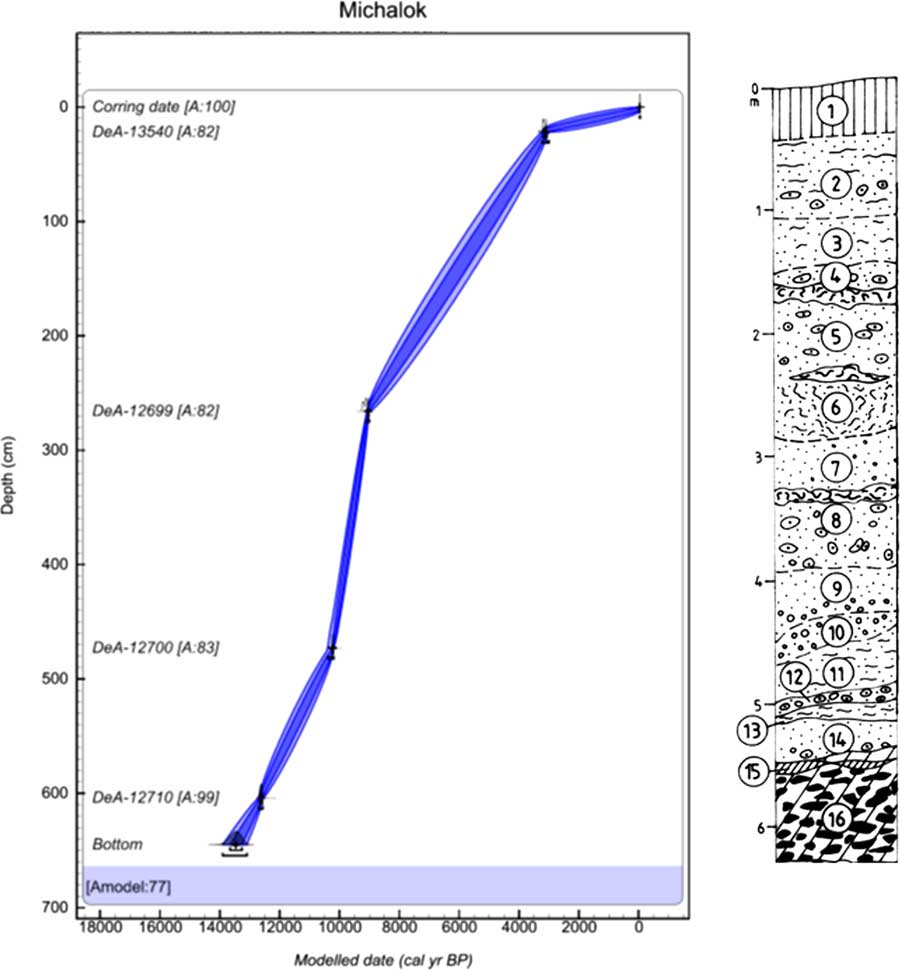
The final Michalok profile (48°59′53.823″N, 21°37′53.193″E; 315 m asl) is situated in the easternmost split of the western Carpathians in the Ondavská vrchovina highlands. The site is part of the Klippen belt that separates the inner and outer western Carpathians. At this site, the tufa was deposited in the flysch valley 0.5 km north of Michalok village, approximately 8–10 m above the right bank of the Čičava Brook (Fig. 4).
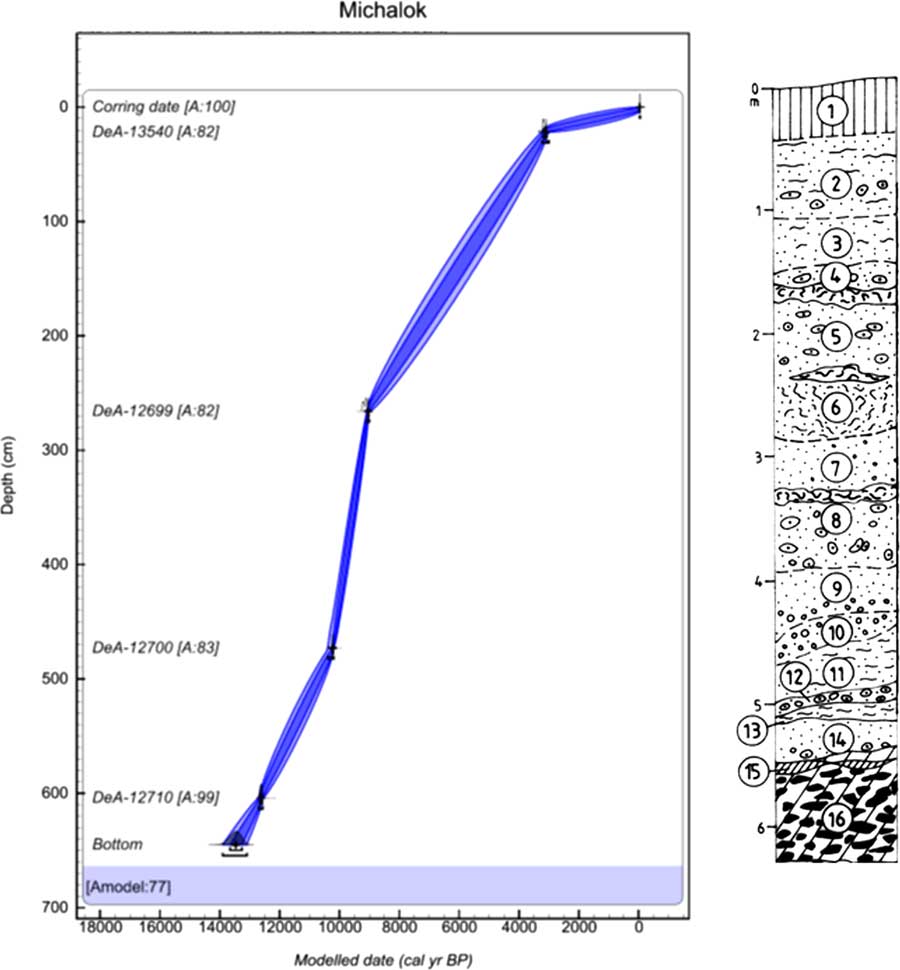
Figure 4 (color online) Lithology and age–depth model of Michalok in the Ondavská vrchovina highlands. Model was made in OxCal v4.3.2 (Bronk Ramsey, Reference Bronk Ramsey2017) with InCal 13 atmospheric curve (Reimer et al., Reference Reimer, Bard, Bayliss, Beck, Blackwell, Bronk Ramsey, Buck, Cheng, Edwards, Friedrich, Grootes, Guilderson, Haflidason, Hajdas, Hatté, Heaton, Hoffman, Hogg, Hughen, Kaiser, Kromer, Manning, Niu, Reimer, Richards, Scott, Southon, Staff, Turney and van der Plicht2013). The laboratory codes of all radiocarbon dates used in the models are in accordance with Table 1. Numbers of layers indicate the centre of the sample. 1, brown-greyish rendzina; 2, pale grey and grey loamy tufa (white efflorescences and concretions); 3, grey, ochre-grey, rusty brown, and black finely cloudy loam tufas; 4, pale and dark grey sand tufa with larger incrustations; pale and dark grey sand tufa, and rusty spotted decaying structural tufa at the base; 5, gray and rusty sand tufa with larger incrustations and tubers, based on a lens of rusty structural tufa; 6, upper part of the layer grey, stronger structural, degradable, and sandy tufa; 7, beige-grey sand tufa with larger incrustation, grey structural tufa at the base; 8, light beige-grey sand-clay incrustations and cobbles with lenses of structural tufa; 9, light ochre-grey sandy tufa with a larger incrustations at the bottom; 10, grey, rusty, spotted, slightly wrinkled coarse incrustation; 11, white-grey sandy sand tufa; 12, yellow-grey small incrustations and tufa cobbles; 13, light grey sandy and earthy tufa with small incrustations, black curled smudge at the base; 14, grey earthy-sandy tufa with a lens of rusty yellow slope sediments with incrustations; 15, lens of grey marl and scree; 16, grey and yellow spotted sandstones in clayey and sandy sediment, arrangement mostly subhorizontal and oblique
Fossil mollusc sampling
Fossil molluscs from all sites were taken from cleaned profiles resulting from artificial extraction (Krivoštianka) or from trenches in naturally eroded sediments. The Krivoštinka and Starianska sites were sampled in 1963, and the Michalok site in 1986. All molluscan successions were sampled by standard methods (Ložek, Reference Ložek1964): 8 dm3 of space-discrete samples of the sediment were taken from the central part of each layer of distinguishable lithology (the central part of the sample is marked by layer number in Figs. 2–4). Mollusc shells were extracted from the sediments by a combination of floating and sieving. After careful drying, each sample was disaggregated in water and then hydrogen peroxide was applied to remove organic material. Floating snails were repeatedly decanted into a 0.5 mm sieve and dried under laboratory conditions. Afterwards, the sediment including the shells that did not float was dried and sorted by sieving. Shells and their fragments were systematically removed from the sediment and determined under a binocular microscope.
Ecological groups were used sensu Juřičková et al. (Reference Juřičková, Horsák, Horáčková, Abrhám and Ložek2014b); the nomenclature follows Welter-Schultes (Reference Welter-Schultes2012). Mollusc diagrams express the absolute proportions of the total number of species (malacospectra of species [MSS]) in separated layers.
Radiocarbon dating
Radiocarbon data analyses were performed in the Poznań Radiocarbon Laboratory, Poland, and in the Izotoptech Zrt. in Debreceen, Hungary. Mollusc shells were measured by the accelerator mass spectrometry method and calibrated for variable initial 14C concentration using the OxCal v. 4.3.2 calibration program (Bronk Ramsey, Reference Bronk Ramsey2009a) (Table 1). Shells can contain so-called dead carbon, which can lead to an overestimation of their age (Goodfriend and Stipp, Reference Goodfriend and Stipp1983). To minimize this dating error, we used the shells of small species, predatory members of the family Zonitidae, and shells that have yielded reliable ages in our previous studies (e.g., Juřičková et al. Reference Juřičková, Horsák, Horáčková, Abrhám and Ložek2014b). Although some inaccuracy in dating cannot be ruled out, such errors are likely on the order of centuries (at most) based on 14C analyses of small terrestrial gastropods in North America (Pigati et al., Reference Pigati, Rech and Nekola2010).
The lithology was used as another proxy to describe the sedimentation characteristics of particular sites (Figs. 2–4). The ages of nondated layers were estimated based on an age–depth model constructed for each succession (Supplementary Material 1) using 13 radiocarbon dates (see Table 1). Dates were extrapolated using a script in OxCal v. 4.3.2 (Bronk Ramsey, Reference Bronk Ramsey2009b). Three radiocarbon dates (DeA-10032 and Poz-77746 from Krivoštianka and DeA-12694 from Starianska dolina) were excluded, because they caused an age reversal and decreased the quality of the age–depth models to zero. We tried to obtain the first excluded date (DeA-10032) from a large-sized snail Fruticicola fruticum, while the other excluded 14C date (Poz-77746) was interpreted as an outlier by outlier analysis in OxCal (Bronk-Ramsey, 2009b), probably because there was a very similar date in sequence layer 7 of the Krivoštianka succession. The age–depth model was calculated (model agreement = 91%, Supplementary Material 1) with 1 cm resolution based on a P_Sequence function with the k parameter equal to 0.5 cm−1 and log10(k/k0) equal to 0.3. The 14C date (DeA-12694) from layer 2 of the Starianska dolina succession was also excluded, again because it was revealed to be an outlier by outlier analysis in OxCal. The age–depth model was calculated (model agreement = 63%, Supplementary Material 1) with 1 cm resolution based on a P_Sequence function with the k parameter equal to 1 cm−1 and log10(k/k0) equal to 1. The age–depth model of Michalok was calculated (model agreement = 77%, Supplementary Material 1) with 1 cm resolution based on a P_Sequence function with the k parameter equal to 0.5 cm−1 and log10(k/k0) equal to 0.3. All age–depth models were modelled using OxCal v. 4.3.2 (Bronk Ramsey, Reference Bronk Ramsey2017).
RESULTS
In the easternmost outcrops of the western Carpathians and the westernmost eastern Carpathians, which tower above the east Slovak lowland, we describe three molluscan successions covering the late glacial period, the early and middle Holocene, and part of the late Holocene (sensu Walker et al. Reference Walker, Berkelhammer, Björck, Cwynar, Fisher, Lond, Lowe, Newham, Rasmussen and Weiss2012). These are the first molluscan successions described from the eastern Carpathians.
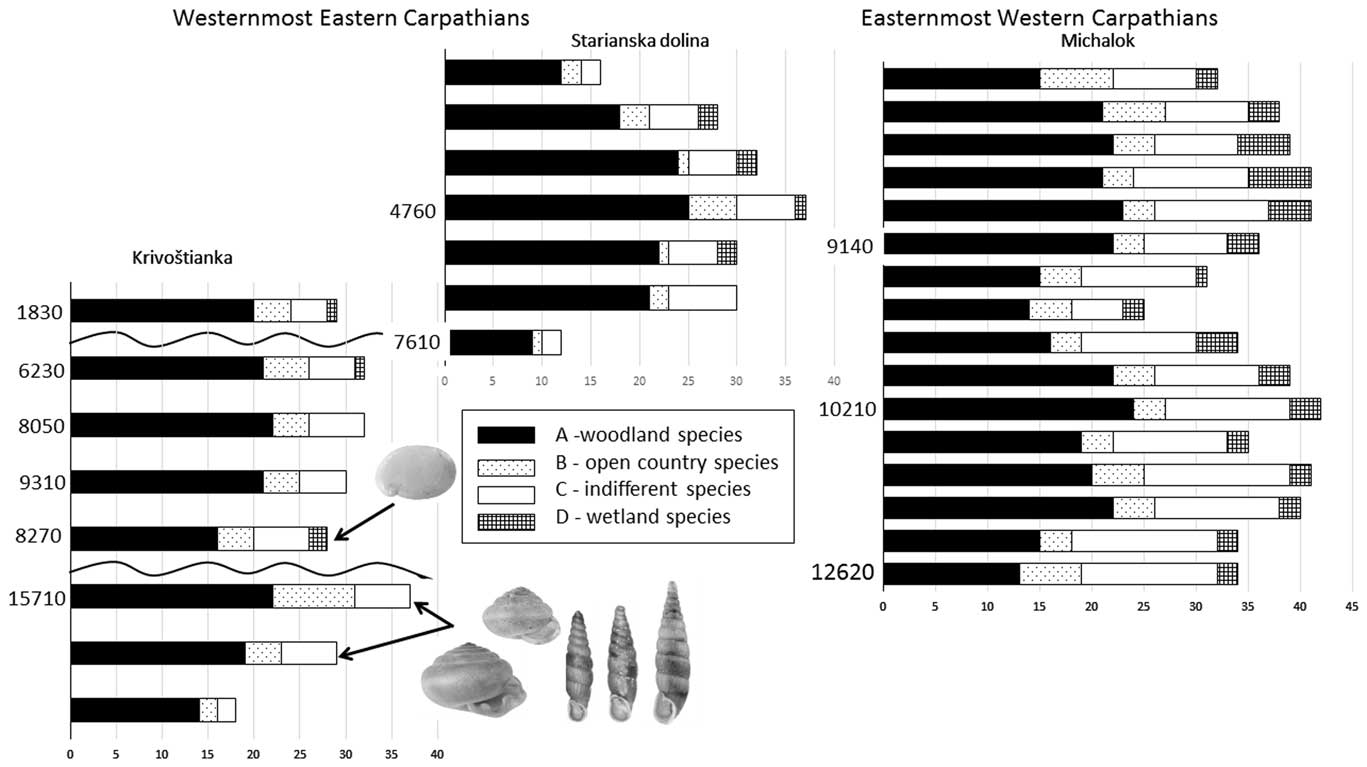
The first known fossil records of three local endemics were found: Carpathica calophana occurred only in the eastern Carpathians, while Petasina bielzi and Perforatella dibothrion both occurred in the eastern Carpathians and the easternmost western Carpathians. The succession from the western Carpathian foothills contained the easternmost occurrences of some western Carpathian species (Orcula dolium) and central European species (e.g., Discus rotundatus, Eucobresia diaphana), and showed a shift in the ranges of east Carpathian species to the west during the middle Holocene (Table 4). The two molluscan successions of the Slovak eastern Carpathians lack a considerable number of common mid-European species (see “The Slovak Eastern Carpathians—An Important Zoogeographic Boundary”), while canopy forest species occurred there very early during the late glacial period (Fig. 5).
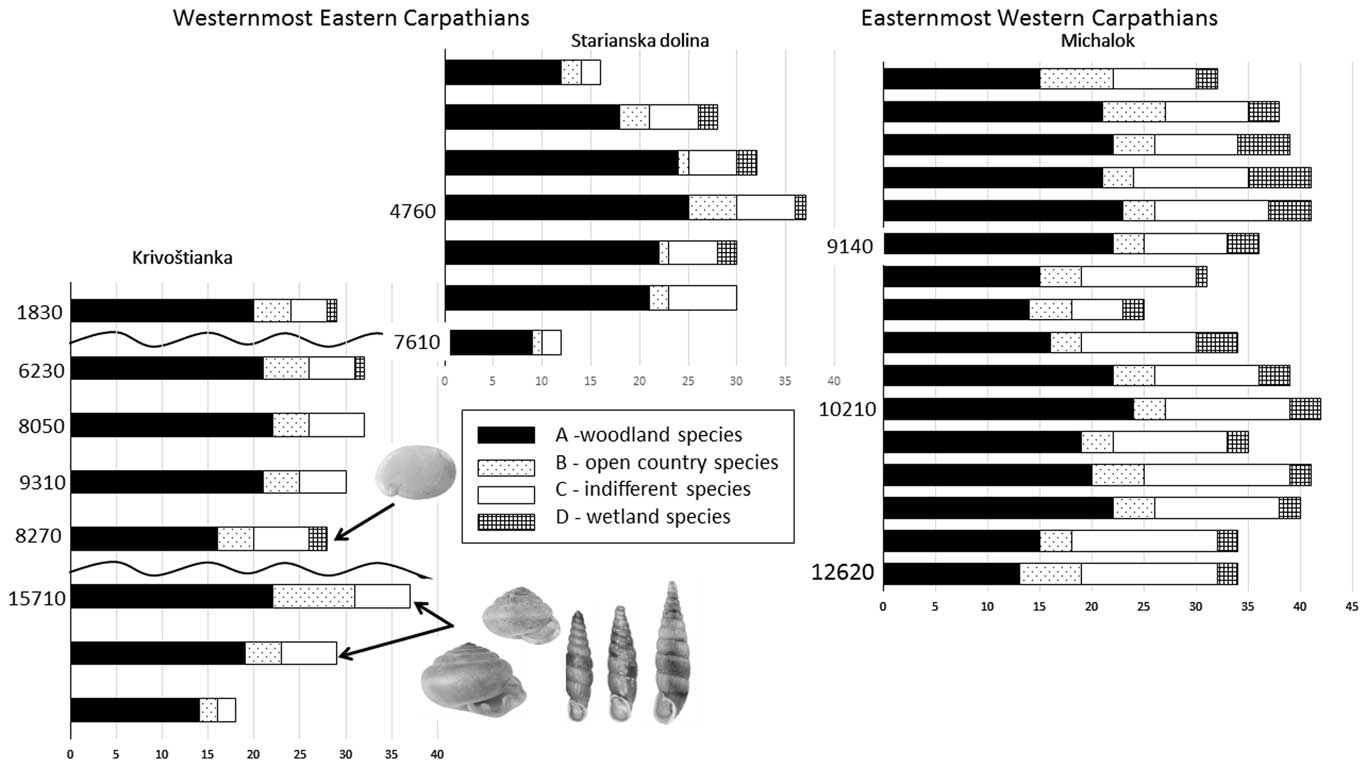
Figure 5 Comparison of total number of molluscan species (malacospectra of species [MSS]) of all three molluscan successions of the Slovak eastern Carpathians. Ecological groups sensu Juřičková et al. (Reference Juřičková, Horáčková and Ložek2014b). On the left side of every MSS malacospectra are radiocarbon data for particular layers in cal yr BP (see Table 1). The first occurrence of each local endemic is marked by an arrow. From left to right: Carpathica calophana, Perforatella dibothrion, and Petasina bielzi.
Molluscan succession from Krivoštianka
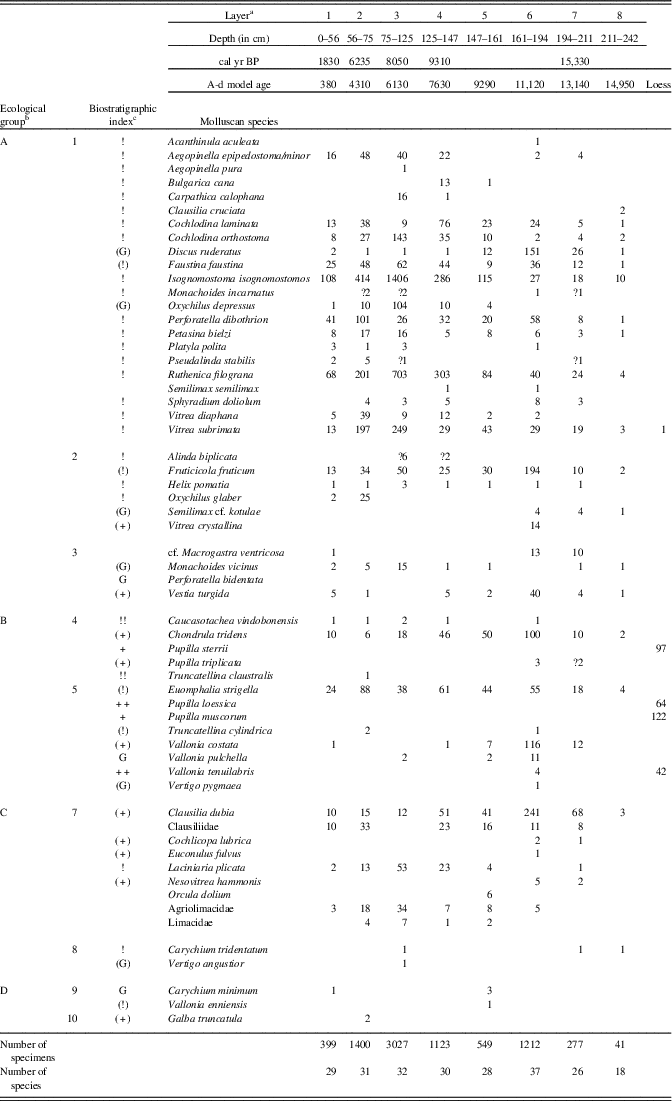
At the foot of the Okúr slope, a small loess deposit was exposed that is not associated with the main profile. It provided characteristic pleniglacial fauna (rightmost column in Table 2) consisting of Pupilla muscorum, P. sterrii, P. loessica, and Vallonia tenuilabris (tens of shells in 8 dm3 samples). One shell of the mountain woodland species Vitrea subrimata was also found.
Table 2 Number of mollusc specimens in separate layers of the Krivošťanka, Okúr, profile.
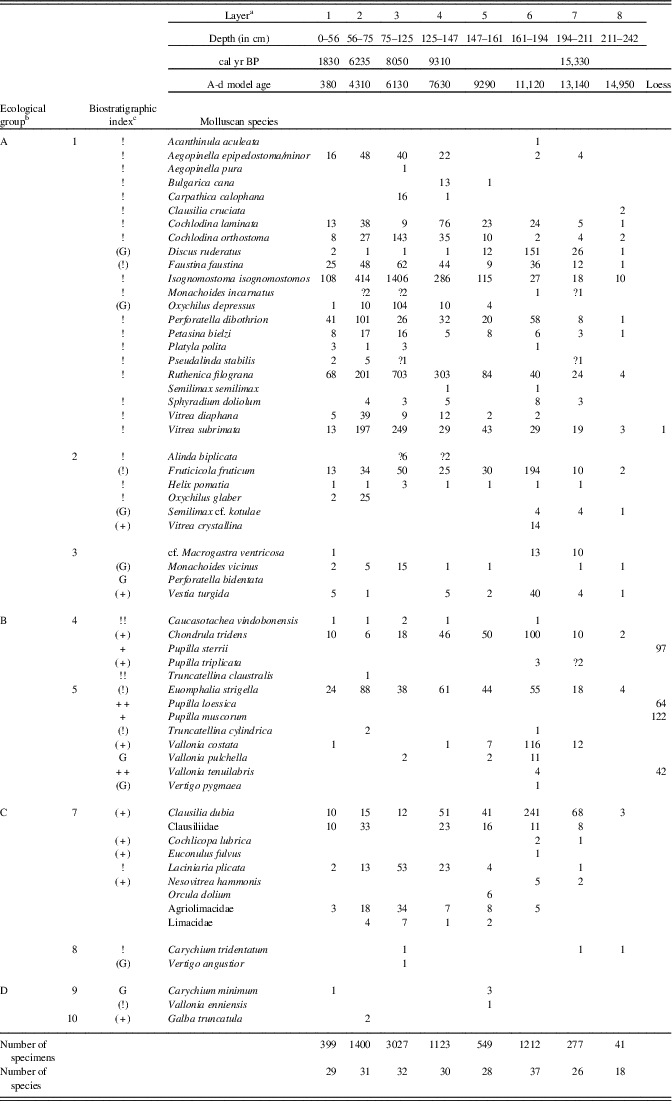
a Layer number related to Fig. 2.
b Ecological characteristics: general ecological groups: A, woodland (in general); B, open country; C, woodland/open country; D, water, wetland. Ecological groups: 1, woodland (sensu stricto); 2, woodland, partly semi-open habitats; 3, damp woodland; 4, xeric open habitat; 5, open habitats in general (moist meadows to steppes); woodland/open country: 6, predominantly dry; 7, mesic or various; 8, predominantly damp; 9, wetlands, banks; 10, aquatic habitats.
c Biostratigraphic characteristics: +, characteristic loess species; (+), local or occasional loess species; !, species of warm phases; (!), eurythermic species of warm phases; !!, index species of warm phases; G, species surviving glacial out of loess zone; (G), species surviving glacial out of loess zone as relics.
The main profile consists of colluvial strata and is situated statigraphically above the loess profile. The first two layers are humic. Triassic limestone fragments occur inside the lower nonhumic loam layers (Fig. 5). The strata of layers 8 to 6 (Fig. 5, Table 2) seem to have originated during the warm late glacial Bølling oscillation, which has been assigned an age range of 15,000–14,000 cal yr BP, and end in other humic strata (layer 6). Our radiocarbon data are somewhat older (15,330 cal yr BP) and come directly from the shells of the forest species Discus ruderatus. However, there was also a rich forest fauna, including the local woodland endemic species Perforatella dibothrion and Petasina bielzi. The sediments between this period and the onset of the Holocene were eroded. The bedrock consisted of Triassic limestone.
The part of the slope sequence between layers 5 and 3 probably contains the early Holocene sedimentation period, with a fully developed woodland fauna occurring there and again including another east Carpathian endemic, Carpathica calophana. The woodland fauna also survived the 8.2 ka cold event, when a fluctuation in the eastern border of the central European species Monachoides incarnatus (uncertain determination) appeared. The occurrence of Vertigo angustior during this cold event may indicate a temporary wetland.
The temporary appearance of Truncatellina claustralis and Oxychilus glaber during the Holocene climate optimum (layer 2: 6235 cal yr BP) may indicate additional warming. The sediments between layers 1 and 2 were eroded. The surface soil (layer 1: 1830 cal yr BP) also included a well-developed canopy forest fauna, which differs considerably from the present-day poor local communities, which are dominated by a reduced number of dryness-tolerant species. The younger sediments were completely eroded, so we have no information about the late Holocene fauna of this site.
Molluscan succession from Starianska
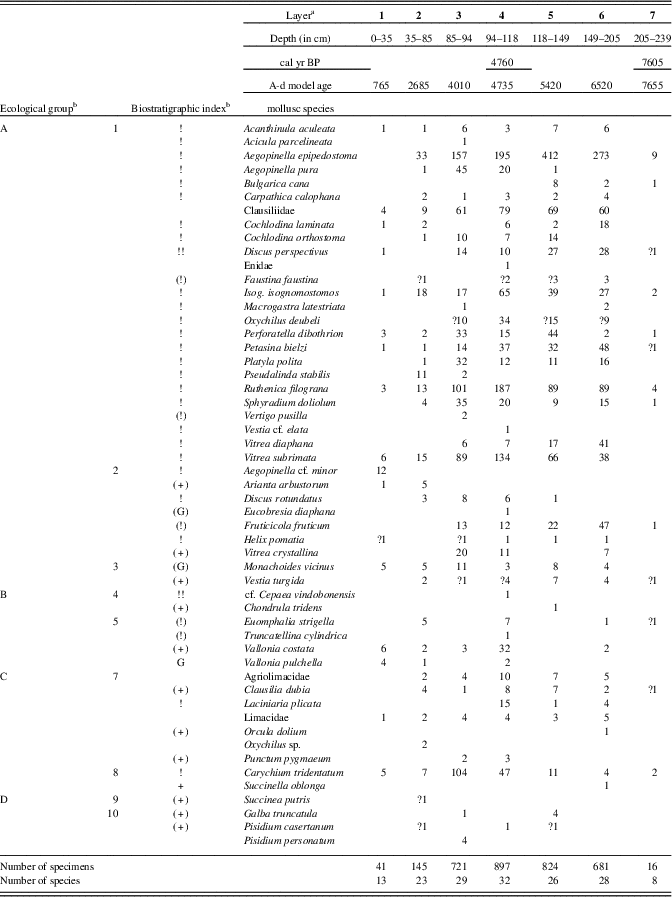
The colluvial sediments of Starianska contain slopewash loams and limestone scree with a tufa admixture (Fig. 3). The upper layer consists of humic soils with Triassic limestone. Starting with layer 4, scattered tufa particles appear. The bedrock consists of Triassic limestone scree.
As seen in Table 3 and Figure 5, the major part of the succession (layers 6–3) was dominated by fully developed Holocene climate optimum assemblages of a temperate canopy forest, including a number of species that prefer the submontane zone. Index species of the early Holocene such as Discus ruderatus, Nesovitrea petronella, and Vertigo substriata are absent, and succession begins before the Holocene climate optimum, which is in agreement with the radiocarbon dating of 7610 cal yr BP for the basal layer 7. Older sediments had previously eroded. The west Carpathian species Orcula dolium, which has been recently extirpated in the eastern Carpathians (Lisický, Reference Lisický1991), was temporarily recorded during the onset of the Holocene climate optimum (layer 6). In layer 4 (4760 cal yr BP), the highest species diversity (32 species) as well as the highest species abundances were found. This period can be considered as the maximum of forest species, whose abundance increased to 87% of the total. The temporary occurrence of the central European species Eucobresia diaphana during this optimum and the extinction of the west Carpathian endemic Acicula parcelineata in this area thereafter could indicate a moderate decrease in humidity. Also noteworthy is the late occurrence of the eastern Carpathian species Pseudalinda stabilis (first appearance since layer 3), as well as the occurrence of the central European species Discus rotundatus only during the Holocene climate optimum. The occurrence of the steppe species Caucasotachea vindobonensis and Chondrula tridens indicate deforestation, which, given natural conditions not suitable for steppe occurrence, may point to local human impacts. The decrease of the woodland faunal diversity in layer 2 may correspond with climate deterioration during the Subboreal period (sensu Jäger, Reference Jäger1969), which is in agreement with the change in sedimentation indicated by scree accumulation (Fig. 3).

Molluscan succession from Michalok
The upper layer of naturally cut tufa sediments in the Michalok site consists of rendzina soil. Layers 14–2 consist of tufa fragments or sandstone tufa sediments. Marl and scree sediments of layer 15 lie on sandstones and sandy sediment of bedrock.
The succession contained only late glacial and early Holocene malacocoenoses (Fig. 5, Supplementary Material 1). Besides the typical late glacial species (Columella columella, Nesovitrea petronella, Vertigo substriata), molluscan assemblages of layers 16–10 contained species more typical of the early Holocene of central Europe (Discus ruderatus, Ena montana) but surprisingly also for the Holocene climate optimum (e.g., Bulgarica cana, Isognomostoma isognomostomos, Ruthenica filograna), including thermophilous species (Discus perspectivus, Sphyradium doliolum, Cochlodina orthostoma). Trochulus hispidus occurred there only during the late glacial and was extirpated after that. The west Carpathian endemic Plicuteria lubomirski occurred there only during the late glacial period and then shifted westwards. During the Preboreal period (layers 9–6), a fully developed canopy forest fauna again occurred at this site, including local eastern Carpathian endemics (Perforatella dibothrion, Petasina bielzi), which occurred there in their westernmost boundary. Some central European species occurred there for the first time since the late glacial period (Isognomostoma isognomostomos, Ruthenica filograna, Sphyradium doliolum), others for the first time since the Preboreal period (Vitrea diaphana), and still others for the first time since the onset of the Holocene climate optimum (Daudebardia rufa). Some of these taxa do not occur there today (Alinda biplicata, Discus rotundatus, Vitrea contracta) and some western Carpathian endemics appeared later (Vitrea transsylvanica, 3150 cal yr BP; Macrogastra latestriata, 7815 cal yr BP).
DISCUSSION
The influence of the Carpathians in the evolution of European biota is clear (e.g., Feurdean et al., Reference Feurdean, Persoiu, Tantau, Stevens, Magyari, Onac and Markovic2014; Mráz and Ronikier, Reference Mráz and Ronikier2016), but we tend to refer the Carpathians as a single unit. More and more studies, including ours, point to the need for accurate research on particular Carpathian regions to find the precise shape of recent zoogeographic boundaries, localize glacial refugia, and assess the role of these refugia in the postglacial colonisation of Europe.
The Carpathians as a case study area for northern glacial refugia
In discussing northern glacial refugia or northern range borders of temperate species surviving the LGM, the Carpathians and the Alps, especially their foothills, are always considered first (e.g., Schmitt and Varga, Reference Schmitt and Varga2012; Feurdean et al., Reference Feurdean, Persoiu, Tantau, Stevens, Magyari, Onac and Markovic2014; Mráz and Ronikier, Reference Mráz and Ronikier2016; Tzedakis et al., Reference Tzedakis, Emerson and Hewitt2013; Jamrichová et al., Reference Jamrichová, Petr, Jimenéz-Alfaro, Jankovská, Dudová, Pokorný and Kołaczek2017). Because the Carpathians probably contain the world’s most fully explored northern refugia, this mountain belt is an excellent case study area for the research of this problem. Nevertheless, existing phylogeographic and palaeocological studies are divided on where exactly the Carpathians refugia of temperate species were situated, what their area was, and what their species compositions were.
Based on direct pollen and/or macrofossil evidence, Tzedakis et al. (Reference Tzedakis, Emerson and Hewitt2013) questioned the occurrence of broadleaf temperate trees north of 45°N in central and western Europe including the western Carpathians. However, the survival of some boreal trees (Jankovská and Pokorný, Reference Jankovská and Pokorný2008) was directly evidenced in the western Carpathians, and Tzedakis et al. (Reference Tzedakis, Emerson and Hewitt2013) accepted the restricted persistence of boreal trees in eastern Europe up to 49°N. Our molluscan successions are situated at 48°N, and are thus important for elucidating this issue.
Based on records from the southern parts of the Pannonian lowlands (Sümegi et al., Reference Sümegi, Magyari, Dániel, Molnár and Töröcsik2013), the late glacial occurrence of temperate trees was anticipated in the southwestern foothills of the western Carpathians. But Jamrichová et al. (Reference Jamrichová, Petr, Jimenéz-Alfaro, Jankovská, Dudová, Pokorný and Kołaczek2017) modelled broadleaf tree refugia in the humid northwestern mountain part of the western Carpathians, emphasizing the interactions between temperature and precipitation, rather than temperature alone.
The existence of northern glacial (micro)refugia of woodland land snails is connected only with the occurrence of broadleaf temperate trees, especially those with citrate calcium in the leaf litter (e.g., Wäreborn, Reference Wäreborn1970), because a majority of canopy forest snails, including some identified in our study, cannot survive without them.
We have no direct (fossil) evidence for the survival of such trees in either the western Carpathians or in the eastern Carpathians, but much indirect evidence supports this possibility (Jamrichová et al., Reference Jamrichová, Potůčková, Horsák, Hajnalová, Bárta, Tóth and Kuneš2014b, Reference Jamrichová, Petr, Jimenéz-Alfaro, Jankovská, Dudová, Pokorný and Kołaczek2017; Novák et al., Reference Novák, Trotsiuk, Sýkora, Svoboda and Chytrý2014; Hájek et al., Reference Hájek, Dudová, Hájková, Roleček, Moutelíková, Jamrichová and Horsák2016). The survival of lime and elm have been modelled in the western Carpathians, and both species offer suitable conditions for canopy forest land snails.
Are we on the trail of the eastern Carpathian refuge for forest species?
Unlike plant remains, we have fossil evidence of six temperate woodland snail species surviving the LGM in the western Carpathians (Ložek, Reference Ložek2006; Juřičková et al., Reference Juřičková, Horáčková and Ložek2014a), but there is as yet no such evidence for the eastern Carpathians. However, Feurdean et al. (Reference Feurdean, Wohlfarth, Björkman, Tantau, Bennike, Willis, Farkas and Robertson2007, Reference Feurdean, Klotz, Brewer, Mosbrugger, Tămaş and Wohlfarth2008a, Reference Feurdean, Klotz, Mosbrugger and Wohlfarth2008b, Reference Feurdean, Spessa, Magyari, Willis, Veres and Hickler2012) documented the occurrence of some coniferous and broadleaf trees in the Romanian eastern Carpathians before 14,700 cal yr BP. The problem of direct evidence of an LGM refugia for temperate species in the Carpathians and elsewhere north of 45° (Tzedakis et al., Reference Tzedakis, Emerson and Hewitt2013) is related to finding suitable sites, which is largely a matter of luck and suitable fossilization conditions.
Unfortunately, apart from several localities of poor loess fauna with leading species such as Vallonia tenuilabris and Pupilla loessica (Košťálik, Reference Košťálik1999; our data), glacial sediments with molluscan fauna or other fossil remains have not yet been found in the eastern part of Slovakia. The only exceptions are the lake sediments from the Vihorlat Mountains (Hájková et al., Reference Hájková, Pařil, Petr, Chattová, Grygar and Heiri2016) and our data. The open woodlands of Pinus, Betula, Larix, and Picea were reconstructed by Hájková et al. (Reference Hájková, Pařil, Petr, Chattová, Grygar and Heiri2016) from the late glacial period in the Slovak eastern Carpathians, replaced by broadleaf temperate forests dominated by Betula and later by Ulmus at the onset of the Holocene. But their site is situated relatively high and in a north-facing location.
We found late glacial colluvial deposits in layers 6, 7, and 8 in the Krivoštianka site and slightly younger sediments from the Michalok site (layer 12). Radiocarbon data come directly from the several specimens of Discus ruderatus from layers 6 and 7 of the Krivoštianka profile. It is striking that the fully developed canopy forest malacofauna, including local endemic species, dates back to the period just before the warm oscillation, usually called Bølling (GI-1e Greenland interstadial). This period, formerly called the Oldest Dryas (but see Rasmussen et al., Reference Rasmussen, Bigler, Blockley, Blunier, Buchardt, Clausen and Cvijanovic2014), was relatively cold, and therefore the presence of forest fauna is a rather unusual phenomenon. This may have been caused by the dating of shells overestimating their true age (Goodfriend and Stipp, Reference Goodfriend and Stipp1983), and this fauna may actually come from the slightly younger warm Bølling oscillation. However, six canopy forest species were also recorded from the Younger Dryas sediments (12,620 cal yr BP) in the Michalok site, although an overestimation of their age cannot be excluded. In any case, these two sites with fully developed late glacial woodland molluscan communities may indicate the presence of a glacial refugium for forest fauna in these warm foothills of the Slovak eastern Carpathians.
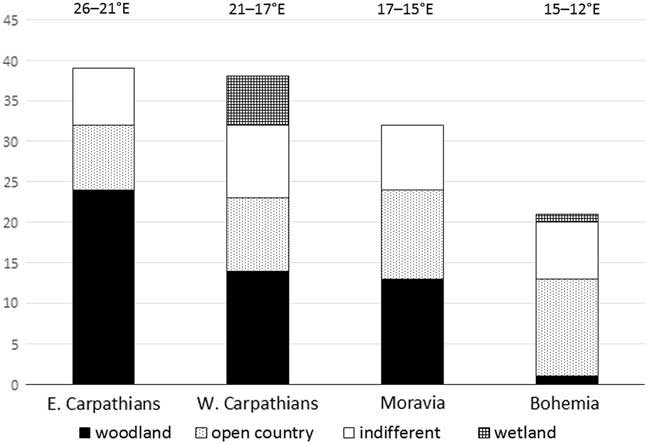
To assess the possibility of an east–west climatic gradient of the survival of demanding woodland species during the late glacial in central Europe, we compared the proportion of particular ecological groups in similarly aged radiocarbon-dated (about 15,000 cal yr BP) molluscan assemblages in four central European areas: Bohemia and Moravia as the western and eastern parts of the Bohemian Massif, and the western and eastern Carpathians. As can be seen from Table 4, the proportion of forest species declines significantly towards the west, and in contrast, the proportion of open-country and indifferent species slightly increases in the opposite direction. The proportion of particular ecological groups is practically the same in all four areas recently (e.g., Welter-Schultes, Reference Welter-Schultes2012; Horsák et al., Reference Horsák, Juřičková and Picka2013). There is no doubt that more accurate dating and additional profiles will be needed to precisely describe the faunal changes in this area.
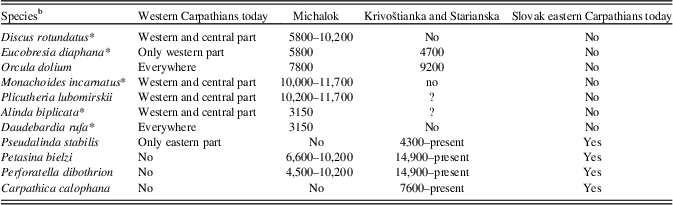
Table 4 Occurrence of the western Carpathian and eastern Carpathian species during the Holocene in the molluscan successions of Michalok, Krivoštianka, and Starianska in comparison with their modern distribution in the western and the Slovak eastern Carpathians.Footnote a

a Time span of occurrence in cal yr BP.
b An asterisk (*) indicates species also occur in central Europe.
The Slovak eastern Carpathians—an important zoogeographic boundary
The most important Carpathian zoogeographic boundary seems to be the border between the western and the eastern Carpathians. This boundary was defined based on geomorphology, recent endemic species’ distribution ranges, and the phylogeography of especially high mountain taxa (for a review, see Mráz and Ronikier, Reference Mráz and Ronikier2016). But did the same boundary exist in the Quaternary?
Loess is the most common type of sediment related to glacial activity in central Europe, but it sedimented in lowlands. Thus, loess molluscan assemblages provide information only about lowland environments (e.g., Ložek, Reference Ložek1964). The eastern Carpathian lowlands together with the marginal zone of the adjacent eastern and western Carpathian foothills differ markedly from the lowlands of southwestern Slovakia, south Moravia, and middle Bohemia. These latter are characterised by a warm-dry climate (450–550 mm annual rainfall), extensive calcareous loess covers, and typical chernozem soils, whereas the eastern Slovak lowland is dominated by CaCO3-deficient dust loams and a reduced occurrence of true loess that is mostly low in carbonate, despite its more continental warm climate (January 3–4°C, July 19–20°C, but 650–730 mm annual rainfall). Chernozem soils occur only in minor areas and are entirely decalcified and luvic. Such conditions are characteristic of the so-called humid loess landscape (sensu Fink, Reference Fink1956, Reference Fink1961). This is reflected in the composition of the local loess fauna without the index species Helicopsis striata (Košťálik, Reference Košťálik1999; our data) but with the occurrence of Vitrea subrimata, which is a species known so far from the loess context only from the Riss glacial period at Nové Mesto nad Váhom (western Slovakia) (Kukla et al., Reference Kukla, Ložek and Bárta1962). Thus, the presence of a local zoogeographic boundary is already apparent from the late glacial period.
The role of the Slovak eastern Carpathians as an important zoogeographic boundary for temperate fauna and flora has also been discussed (for a review, see Mráz and Ronikier, Reference Mráz and Ronikier2016). However, this is evident only when evaluating modern distributions of many European organisms, including molluscs (Kerney et al., Reference Kerney, Cameron and Jungbluth1983; Welter-Schultes, Reference Welter-Schultes2012). Twenty western European, central European, and western Carpathian molluscan species have their eastern borders just outside the gates of the Slovak eastern Carpathian foothills (Orcula dolium, Sphyradium doliolum, Chondrina clienta, Chondrina tatrica, Pyramidula pusilla, Cochlodina cerata, Discus rotundatus, Semilimax semilimax, Eucobresia nivalis, Daudebardia rufa, Aegopinella nitens, Macrogastra plicatula, Clausilia parvula rugosa, Alinda biplicata, Xerolenta obvia, Urticicola umbrosus, Monachoides incarnatus, Petasina unidentata, Helicodonta obvoluta, Isognomostoma isognomostomos, and Causa holosericea). Five of these were recorded in the Michalok site at the easternmost split of the western Carpathians, which is situated a little less than 20 km from the eastern Carpathian profiles. While the woodland species Daudebardia rufa, which needs abundant leaf litter, appeared only in the youngest layer, other demanding woodland species (Isognomostoma isognomostomos, Spyradium dolium) occurred there throughout the Holocene without crossing the 20-km-wide lowland. On the contrary, two east Carpathian species have their western border in the Slovak eastern Carpathians (Carpathica calophana, Petasina bielzi).
However, our data show that this zoogeographic boundary shifted slightly to the west during the Holocene. While C. calophana never occurred in the western Carpathians (Michalok and other Carpathian profiles [Horáčková et al., Reference Horáčková, Ložek and Juřičková2014]), P. bielzi had already reached this site during the late glacial period. On the contrary, the mid-European species Discus rotundatus and Eucobresia diaphana, which only recently reach central Slovakia, had already shifted their easternmost borders by hundreds of kilometres (Michalok) in the Preboreal period, reaching the eastern Carpathians (Starianska dolina) during the Holocene climate optimum. While D. rotundatus spread gradually (see below), E. diaphana probably reached the Michalok site via long-distance transport, because we have no Holocene record of this species in the western Carpathians (Ložek, Reference Ložek1964, Reference Ložek1982; unpublished own data) (Table 4).
Finally, Discus rotundatus is a common European species whose recent range centre lies in central Europe. Shifts of its range border westerly have regularly been used as an indication of the Holocene climate optimum in western Europe (e.g., Meyrick, Reference Meyrick2001; Limondine-Lozouet and Preece, Reference Limondin-Lozouet and Preece2004). Discus rotundatus reached the Rhineland area at approximately 7400 cal yr BP and Normandy around 7910 cal yr BP. The occurrence of this species in the opposite border of its range in the easternmost western Carpathians (Michalok) was 10,215 cal yr BP. Scattered occurrences of this species were recorded in the whole profile, but it is recently extinct there. On the contrary, in the westernmost eastern Carpathians (Starianska), fluctuation of its range was recorded only during the Holocene climate optimum.
It is clear that the boundary of many species shifted mostly in the south–north direction at a scale of hundreds kilometres between the glacial and interglacial periods in Europe (e.g., de Lattin Reference de Lattin1967; Feuerdan et al., Reference Feurdean, Persoiu, Tantau, Stevens, Magyari, Onac and Markovic2014), but it seems that some long-distance shifts, mostly in the west–east direction, also took place also during the Holocene, though far fewer. Our data here show that this is not just the case for a single species. Such range-border shifts for many species probably indicates climatic changes in the east–west climatic gradient between continental and oceanic European climates (e.g., Peel et al., Reference Peel, Finlayson and McMahon2007).
CONCLUSIONS
Three late glacial and Holocene molluscan successions were recorded in the easternmost outcrops of the western Carpathians and the westernmost eastern Carpathians. Krivoštianka and Starianska were the first molluscan successions described in the eastern Carpathians, and yielded the first fossil records of the east Carpathian endemic species Carpathica calophana, Petasina bielzi, and Perforatella dibothrion. The easternmost borders of western and central European species and the westernmost borders of eastern Carpathian species shifted during the Holocene climate optimum, probably as reaction to climatic changes in the east–west climatic gradient. We provide evidence of the survival of 21 canopy forest species approximately 15,000 cal yr BP ago in the Slovak eastern Carpathians. There is no evidence of surviving that much demanding species during the late glacial period in the Western Carpathians, Moravia, and Bohemia (Figure 6). Our data support the important role of the Slovak eastern Carpathians as a zoogeographic boundary and a potential glacial refugium.
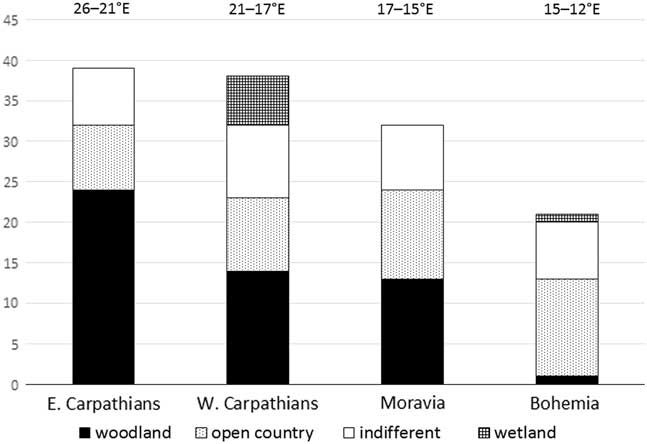
Figure 6 Proportions of molluscan species of particular ecological groups (for a description, see Table 2) in the east–west gradient of central European molluscan assemblages around 15,000 cal yr BP. Sites in Bohemia are situated approximately between 12°E to 15°E, in Moravia between 15°E to 17°E, in the western Carpathians between 17°E to 21°E, and in the eastern Carpathians at 21°E. The sites used (and dates in cal yr BP) are from the Czech Quaternary database (Horáčková et al. Reference Horáčková, Ložek and Juřičková2014). Eastern Carpathians: Krivoštianka (15,710–15,329 BP, 48°53′28.136″N, 21°50′31.384″E). Western Carpathians: Maštalná cave, Slovak Karst (15,830 cal yr BP, 48°36′05.305″N, 20°27′50.012″E); Hámorská cave, Slovak Karst (14,540 cal yr BP, 48°32′48.514″N, 20°24′37.885″E); Čierna dolina, Slovensky Raj (14,580 cal yr BP, 48°55′00″N, 20°19′45″E); Pivková dolinka, Nízké Tatry Mountains (14,380 cal yr BP, 48°58′26″N, 19°39′48″E); Laskár, Malá Fatra Mountains (14,470 cal yr BP, 48°57′42.962″N, 18°52′37.299″E). Moravia: Pekárna cave, Moravian Karst (14,830 cal yr BP, 49°14′30.927″N, 16°44′45.931″E); Zazděná cave, Moravian Karst (14,680 cal yr BP, 49°22′21.583″N, 16°43′30.044″E); Pálava, Soutěska, southern Moravia (15,840 cal yr BP, 48°51′57.384″N, 16°38′33.663″E); Pálava, Martinka, southern Moravia (15,250 cal yr BP, 48°51′55.986″N, 16°38′4.409″E). Bohemia: Dobrkovický komín, southern Bohemia (14,500 cal yr BP, 48°49′16″N, 14°17′38″E); Štětí, lower Elbe River (15,780 cal yr BP, 50°28′30″N, 14°20′48″E); Karlštejn, Moucha, Bohemian Karst (14,490 cal yr BP, 49°56′6.332″N, 14°11′49.490″E); Tetín, Bohemian Karst (15,970 cal yr BP, 49°56′28″N, 14°06′31″E).
ACKNOWLEDGMENTS
The work was supported by the Czech Science Foundation (GAČR) project 17-05696S and the HACIER project funded from the Norwegian Financial Mechanism 2009–2014 and the Ministry of Education, Youth and Sports under project contract no. MSMT-28477/2014. We are grateful to Jeff S. Pigati and Petra Hájková for their valuable comments on a previous version of the article and David Hardekopf for the English revision.
SUPPLEMENTARY MATERIAL
To view supplementary material for this article, please visit https://doi.org/10.1017/qua.2018.68