


Enzymatic hydrolysis is a common method in food processing to modify the protein fraction towards an improvement of the functional and nutritional properties without loss in nutritional value (Adler-Nissen, Reference Adler-Nissen1986; Kilara & Panyam, Reference Kilara and Panyam2003). Proteolytic enzymes are specific catalysts which accelerate the cleavage of peptide bonds in proteins without generation of degradation products (as would be the case of acid or basic hydrolysis) since the reaction is carried out at moderate conditions of temperature and pH. The degree of hydrolysis (DH) is the parameter used to measure the extent of the hydrolysis reaction, i.e. ratio of number of peptide bonds cleaved to total number of peptide bonds available.
High DH improves the solubility (Abert & Kneifel, Reference Abert and Kneifel1993; Severin & Xia, Reference Severin and Xia2006) and the flavour-enhancing capacity (Nielsen, Reference Nielsen1994) of hydrolysates. However, a limited DH is preferable to improve both the foaming (Halling, Reference Halling1981) and emulsifying capacities (Tuncturk & Zorba, Reference Tuncturk and Zorba2006). With respect to the nutritional properties, the hydrolysis improves the digestibility (Potier & Tomé, Reference Potier and Tomé2008) of hydrolysates. Furthermore, a high DH is desirable in order to reduce the allergenicity and bitterness due to the presence of peptides with high molecular weight (Mahmoud et al. Reference Mahmoud, Malone and Cordle1992; Pedersen, Reference Pedersen1994).
More recently, it has been reported that some peptides released in the course of the enzymatic hydrolysis exhibit numerous bioactivities such as antihypertensive, immunomodulatory or antioxidative activities (Korhonen & Philanto, Reference Korhonen and Philanto2003). To this regard, milk proteins are one of the most studied sources of bioactive peptides (Clare & Swaisgood, Reference Clare and Swaisgood2000), including goat milk protein, which has been found to contain sequences of bioactive peptides (Park et al. Reference Park, Juárez, Ramos and Haenlein2007). In particular, the antihypertensive activity can be highlighted as a significant activity found in bioactive peptides derived from goat milk proteins. Hypertension is a very important health problem because it is one of the main risk factors in cardiovascular diseases. Indeed, the World Health Organisation estimates that by 2020, heart disease and stroke will have surpassed infectious diseases to become the leading cause of death and disability worldwide (Lopez & Murray, Reference Lopez and Murray1998).
Antihypertensive peptides have been shown to be effective in the treatment of hypertension (Jauhiainen & Korpela, Reference Jauhiainen and Korpela2007), mainly due to the inhibition of angiotensin converting enzyme (ACE). This enzyme plays a crucial role in blood pressure regulation, converting angiotensin I into angiotensin II, a highly potent vasoconstrictor molecule, as well as inactivating the vasodilator bradykinin (Johnston, Reference Johnston1992). Therefore, using these food derived inhibitors may reduce the blood pressure. Although the inhibitory capacity of food derived peptides is much lower than that of chemically produced drugs, they do not present side effects attributed to the use of synthetic compounds. For instance, captopril, which is widely used clinically to manage hypertension and congestive heart failure, can provoke dry cough and alterations in serum lipid metabolism (FitzGerald & Meisel, Reference FitzGerald and Meisel2000). Therefore, the introduction of milk derived peptides in food formulas (De Leo et al. Reference De Leo, Panarese, Gallerani and Ceci2009) for the prevention or control of hypertension is of interest.
To exert their effect, the peptides must resist the gastrointestinal digestion as well as pass through the intestine. Normally, peptides are metabolised to their constituent amino acids by brush border membrane peptidases after oral dosing. However, there is a specific peptide transport system that allows di- and tripeptides to pass through the intestine wall without any modification (Foltz et al. Reference Foltz, Cerstiaens, Van Meensel, Mols, Van der Pijl, Duchateau and Augustijns2008; Miguel et al. Reference Miguel, Dávalos, Manso, de la Peña, Lasunción and López-Fandiño2008). These peptides would enter the cardiovascular system and produce a systemic effect. Taking into account that ACE inhibitory peptides usually have between 2 and 12 residues (Li et al. 2004) and the potency of the peptides decreases as the chain-length increases (Roberts et al. Reference Roberts, Burney, Black and Zaloga1999), short-chain peptides resulting from high DH would be desirable.
In this work, goat milk proteins were hydrolysed with two different enzymes (subtilisin and trypsin) for 2·5 h. It has been reported in the scientific literature that the hydrolysis of milk protein with both subtilisin (Geerlings et al. Reference Geerlings, Villar, Zarco, Sánchez, Vera, Zafra, Boza and Duarte2006; Jiang et al. Reference Jiang, Chen, Ren, Luo and Zeng2007; Mao et al. Reference Mao, Ni, Sun, Hao and Fan2007) and trypsin (Pintado & Malcata, Reference Pintado and Malcata2000; Tauzin et al. Reference Tauzin, Miclo and Gaillard2002; Pan et al. Reference Pan, Cao, Guo and Zhao2012) yields peptides with good ACE inhibitory activity. Both hydrophobicity (caused by subtilisin) and basicity (caused by trypsin) are desirable in the C-terminal of peptides (Li et al. Reference Li, Le, Shi and Shrestha2004).
An experimental design was executed in order to obtain an empirical model which allowed optimisation of the operational conditions, temperature (T) and enzyme-substrate ratio (ES), leading to the maximum values of DH and ACE inhibitory activity (ACEI). Finally, the conflict between both objectives required a bi-objective optimisation technique able to determine a set of balanced solutions satisfying both objectives to an adequate degree.
Materials and methods
Substrate and enzymes
Goat milk with a protein concentration of 33 g/l was obtained from local supermarkets. The hydrolysis was undertaken by means of the enzymes subtilisin (EC 3.4.21.62) and trypsin (EC 3.4.21.4) supplied as Alcalase 2·4 l and PTN 6.0S by Novozymes (Bagsvaerd 2880, Denmark). Subtilisin is a serine endoprotease, which hydrolyses peptide bonds with a broad specifity. It is stable within a wide range of pH, between 5·0 and 11·5, showing a maximum activity at temperatures between 50 and 60 °C (Adler-Nissen, Reference Adler-Nissen1986). Trypsin, also a serine endoprotease, cleaves specifically peptide bonds at the C-terminal side of Lys and Arg residues, except for Arg-Pro and Lys-Pro bonds which are normally resistant to proteolysis. The pH optimum for trypsin is 7–9 (Sipos & Merkel, Reference Sipos and Merkel1970) and it is stable within the range of 30–60 °C (Guadix et al. Reference Guadix, Guadix, Páez-Dueñas, González-Tello and Camacho2000).
Hydrolysis procedure
Prior to hydrolysis, goat milk was skimmed in a Sigma 6k15 centrifuge (Sigma Laborzentrifugen, 37520 Osterode am Harz, Germany) at 4 °C and 4800 g for 20 min. The hydrolysis reaction was conducted in a 250 ml jacketed reactor connected to a thermostatic bath (Haake F4S, Thermo Scientific, Karlsruhe D-76227, Germany).
For each experiment, 200 ml skimmed milk was poured into the reactor and the pH was adjusted to 8 by adding 1 m NaOH. When the desired temperature was reached, the reaction was started by adding the enzyme. pH was kept constant in the course of the reaction by means of a automatic titrator (model 718 Stat Titrino, Metrohm, CH-9100 Herisau, Switzerland) which employed 1 m NaOH as titration agent.
According to the pH-stat method (Adler-Nissen, Reference Adler-Nissen1986), the amount of NaOH consumed in the course of the reaction can be related to the DH, Eq. (1):
 $${\rm DH} = n_B /({\rm \alpha} {\cdot} m_P {\cdot}h_{{\rm TOT}} )$$
$${\rm DH} = n_B /({\rm \alpha} {\cdot} m_P {\cdot}h_{{\rm TOT}} )$$where DH is the ratio between the number of peptide bonds cleaved and the number of peptide bonds in the substrate, n B (mol) is the amount of NaOH consumed to keep the pH constant, α is the average degree of dissociation of α-NH2 groups released during hydrolysis, m P=6·6 g is the mass of protein in the substrate and h TOT=0·0082 mol/g is the amount of peptide bonds per mass unit of protein.
After completing 2·5 h of reaction, the enzyme was deactivated by heating at 100 °C for 15 min. Subsequently, the hydrolysates were cooled and frozen at −20 °C until the ACE-inhibition analysis was performed.
ACE inhibitory assay
The spectrophotometric method described by Shalaby et al. (Reference Shalaby, Zakora and Otte2006) was used to determine the ACEI of the hydrolysates. The assay is based in the hydrolysis of the tripeptide N – [3-(2-N-[3-(2-Furyl)acryloyl]-L-phenylalanyl-glycyl-glycine (FAPGG, Sigma, St Louis MO 63178, USA) with the angiotensin converting enzyme from rabbit lung (EC 3.4.15.1, Sigma, St Louis MO 63178, USA). The assay was carried out in a 96-well microplate at 37 °C monitoring the absorbance at 340 nm in a Multiskan FC microplate photometer (Thermo Scientific, Vantaa FI-01621, Finland). This assay is based on the linear decrease of absorbance observed during the hydrolysis of FAPGG with ACE. The slope of this descent is related to the enzymatic activity of ACE and will be less pronounced in the presence of an inhibitor.
In each well 10 μl enzyme solution, 10 μl sample (control or hydrolysate) and 150 μl substrate solution were added. Each sample was analysed in triplicate. The enzyme solution was prepared at 0·25 U/ml with Milli-Q water. The substrate solution was 0·88 mm FAPGG in buffer Tris-HCl 50 mm, pH 7·5 and 0·3 m of NaCl. In order to have better absorbance values, the hydrolysate solutions used in the assay were prepared diluting the original hydrolysates ten-fold with water. Control samples were prepared with Milli-Q water instead of hydrolysates solutions.
The slope of the control sample represents 100% enzyme activity. Therefore, the numerical value of ACEI of each hydrolysate was calculated as the difference from 1 of the quotient between the slopes in the presence of inhibitor and the slope obtained in the absence of inhibitor. These slopes were calculated with the values in the interval from 10 to 25 min, since the linearity is better in this range.
Experimental design
The effect of T and ES (input variables) on the DH and the ACEI (output variables) was investigated for each enzyme using a 5×6 factorial design and response surface methodology (RSM) (Myers et al. Reference Myers, Montgomery and Anderson-Cook2009; Contreras et al. Reference Contreras, Hernandez-Ledesma, Amigo, Martin-Alvarez and Recio2011). T was varied according to the thermal stability of the enzymes assayed: subtilisin was tested at 45, 50, 55, 60, 65 and 70 °C, while trypsin was at 30, 35, 40, 45, 50 and 55 °C. The levels assayed for the ES were 1, 2, 3, 4 and 5% (grams of enzyme per 100 g of substrate).
The Statgraphics software (version 5.1) was used to generate the experimental designs, the statistical analysis and the regression model. The output variables (Y) were related to the input variables by second order polynomials, Eq. (2):
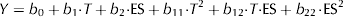 $$ Y = b_0 + b_1 {\cdot} T + b_2 {\cdot} {\rm ES} + b_{11} {\cdot} T^2 + b_{12} {\cdot} T {\cdot} {\rm ES} + b_{22}\! \cdot \!{\rm ES}^2 $$
$$ Y = b_0 + b_1 {\cdot} T + b_2 {\cdot} {\rm ES} + b_{11} {\cdot} T^2 + b_{12} {\cdot} T {\cdot} {\rm ES} + b_{22}\! \cdot \!{\rm ES}^2 $$Each output variable was correlated to the input variables by a model containing one constant, 2 linear terms (associated to T and ES), 2 quadratic terms (T 2 and ES2) and one interaction term (related to the cross-product T·ES). An analysis of variance (ANOVA) analysis was then conducted to evaluate the statistical significance of each term on both output variables. By this approach, an associated probability or P-value was computed for each term in the quadratic model. Setting a level of confidence of 95%, those terms having a P-value below 0·05 will be statistically significant on the output variables.
The regression models obtained by RSM permitted generation of contour plots, where levels of each output variable were represented as a function of both input factors (T and ES). Besides, the quadratic model was optimised in order to determine the combination of input variables (T, ES) leading to a maximum value for each output variable (DH or ACEI).
Bi-objective optimisation
The separated optimisation of the DH and the ACEI, may lead to different optimum values of T and ES. This conflicting behaviour of the experimental factors towards the simultaneous accomplishment of both objectives can be solved by employing a bi-objective optimisation approach. This problem arises when two objectives (possibly conflicting) must be satisfied: attaining a maximum DH and a maximum ACEI simultaneously. More specifically, a bi-objective optimisation technique provides a set of non-inferior solutions, referred to as Pareto Front. In our case, each non-inferior solution is formed by a couple of values of the output variables or objectives (DH, ACEI) satisfying that one single objective cannot be improved without the degradation of the other (Halsall-Whitney & Thibault, Reference Halsall-Whitney and Thibault2006). Among the different multi-objective optimisation techniques, the weighted-sum method (Kim & Weck, Reference Kim and de Weck2005) was chosen. Following this method, an objective function (OBJ) was defined as a linear combination of both output variables (DH and ACEI), Eq. (3):
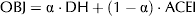 $${\rm OBJ} = {\rm \alpha} \cdot{\rm DH} + (1 - {\rm \alpha} )\cdot{\rm ACEI}$$
$${\rm OBJ} = {\rm \alpha} \cdot{\rm DH} + (1 - {\rm \alpha} )\cdot{\rm ACEI}$$where α stands for a weight factor (0⩽α ⩽1), which quantifies the relative importance attached to each individual objective (e.g. for α=1, the optimisation problem comes down to the maximisation of the DH only). Then, the new bi-objective optimisation problem consisted of estimating the combination of input variables (T, ES), laying within their experimental ranges, which maximises OBJ for a given value of α. The Pareto front was hence generated as the set of solutions obtained by varying α between 0 and 1. A similar procedure was successfully applied to the bi-objective optimisation of the enzymatic hydrolysis of porcine blood protein (Perez-Galvez et al. Reference Perez-Galvez, Almecija, Espejo, Guadix and Guadix2011). Eventually, the selection of a single optimum solution, given by a unique value of α, among the set of non-inferior solutions contained in the Pareto front should be based on economical, technical or safety requirements.
Results and discussion
Degree of hydrolysis
A total of 30 experiments were conducted for each enzyme (Table 1). For subtilisin, the final DH attained after 2·5 h, varied from 0·155 to 0·223. The DH obtained with trypsin was lower, ranging between 0·0936 and 0·159, which could be attributed to the lower hydrolytic capacity of trypsin, which only cleaves two kinds of peptidic bonds.
Table 1. Experimental design and measured values of the output variables

The DH was related to the experimental factors (T and ES) by fitting the experimental data to a second order polynomial. The analysis of variance (ANOVA) for the DH was performed for each enzyme as shown in Table 2a, which also contains the estimated coefficients of the polynomial model. Considering an interval of confidence of 95%, the DH for both subtilisin and trypsin was affected by the T and the ES, because of their respective P-values associated with linear and pure quadratic terms. On the contrary, the interaction term T·ES was not significant for the DH of either for subtilisin or trypsin. The significance of the overall model, assessed by its P-value, was less than 0·001 for both enzymes.
Table 2. (a) Regression coefficients and ANOVA for (a) degree of hydrolysis and (b) ACE inhibitory activity

† R 2=0·963; Standard error of estimate=0·003; Mean absolute error=0·002
‡ R 2=0·950; Standard error of estimate=0·004; Mean absolute error=0·003

† R 2=0·648; Standard error of estimate=0·017; Mean absolute error=0·013
‡ R 2=0·696; Standard error of estimate=0·027; Mean absolute error=0·020
Furthermore, the explained variation for the DH of the predictive models is given by the coefficient of determination (R 2) and it is shown as a footnote in Table 2a. It can be seen that these indicators were 0·9634 for subtilisin and 0·9503 for trypsin, which shows that the quadratic model obtained for DH fitted the data adequately. Table 2a also provides information about the residuals (i.e. the difference between calculated and observed DH values) for both enzymes. These errors are distributed around a mean value (noted as mean absolute error) and present a standard deviation which is indicative of the degree of fit of the model assayed. Besides this, the predicted values of DH (i.e. calculated by the second order model) were represented against the experimental measures in Fig. 1a. These points were arranged along a straight line whose equation was inserted in the figure, so the degree of coincidence between this line and the diagonal is indicative of the goodness of the polynomial fit. Also, all the experimental points are arranged between two dotted lines on both sides of the diagonal represent a deviation of ±10% regarding calculated and observed values.

Fig. 1. Calculated and experimental values of (a) the degree of hydrolysis and (b) ACE inhibitory activity for (1) subtilisin and (2) trypsin.
According to the contour plots shown in Fig. 2a, higher ES values produced hydrolysates with higher DH values at any temperature. This was an expected result since as the concentration of enzyme increases, the number of active centres available rises so that the substrate can react faster. Then, an ES of 5% (highest level) was the optimum for both enzymes. With respect to the temperature, a maximum of DH was found at 60·5 °C for subtilisin (Fig. 2a1), which indicates that higher temperatures lead to an excessive thermal deactivation of the enzyme. Differently, the optimum T for trypsin (Fig. 2a2) was not found inside the experimental range, but coincided with the highest level of T assayed (55 °C), which means that thermal deactivation was not relevant. In summary, the maximum attainable values of DH were 0·218 for subtilisin and 0·159 for trypsin.

Fig. 2. Contour plots of (a) the degree of hydrolysis and (b) ACE inhibitory activity for (1) subtilisin and (2) trypsin.
ACE-inhibitory activity
With respect to the output variable ACEI, it varied during the experiments from 0·250 to 0·347 for subtilisin and from 0·227 to 0·395 for trypsin (Table 1). As performed for the DH, the measured values of ACEI were related to the T and the ES by means of quadratic models. According to the ANOVA analysis (Table 2b), when subtilisin was used in the hydrolysis, only the linear and pure quadratic terms of T significantly affected the inhibitory activity at a confidence level of 95%. In the case of trypsin, only T and ES were statistically significant on the ACEI, where both input variables exert a negative contribution. The goodness of fit of these models was found to be lower than that of the DH. Indeed, according to the values of the determination coefficient (R 2), the ACEI model for subtilisin explained 64·8% of the variability of the experimental data while that for trypsin explained 69·4%. This fact is reflected in Fig. 1b, where the dispersion of the experimental points with respect to the diagonal is broader than that observed for the DH models. Nevertheless, most of the experimental points were inside the dotted lines which limit the 10% deviation.
The contour plot depicted in Fig. 2b1 shows the influence of T upon the ACEI of the hydrolysates for subtilisin. At low temperatures, ACEI increased with T at a decreasing rate, attaining a local maximum of 0·325 at 52·7 °C and ES of 5% (highest level). For trypsin (Fig. 2b2), it can be seen in the contour plot that a maximum ACEI of 0·399 was reached at the lowest levels of T (30 °C) and ES (1%) assayed. Similar findings have been reported by Van der Ven et al. (Reference Van der Ven, Gruppen, de Bont and Voragen2002) for Corolase PP (a mixture of porcine pancreatic enzymes containing trypsin and chymotrypsin) and cardoon extract (Tavares et al. Reference Tavares, Contreras, Amorim, Martín-Álvarez, Pintado, Recio and Malcata2011).
In order to identify a correlation between both output variables DH and ACEI, linear regression of ACEI against DH was performed. In the case of subtilisin, no correlation was found (R 2=0·0005). However, in the case of trypsin, an R 2 of 0·6132 with a slope of −2·1258 was obtained. Such negative effect was also described by Van der Ven et al. (Reference Van der Ven, Gruppen, de Bont and Voragen2002) for the hydrolysis of whey protein.
Bi-objective optimisation
As a result of the optimisation of DH and ACEI, it was observed that, for each enzyme, the maximum values of each objective were reached at different conditions of T and ES. However, the overall purpose of our optimisation approach was to obtain hydrolysates with a high DH (which is desirable in order to obtain a product mixture rich in peptidic fractions with low molecular weight) and, at the same time, exhibiting a high ACEI. This conflict was tackled by performing a bi-objective optimisation aiming to produce a set of solutions which satisfied both objectives to an adequate degree.
By following the sum weighted method, the weight factor (α) was varied from 0 to 1, representing the relative importance attached to the DH with respect to the ACEI. The closer α is to the unit, the higher is the preference towards DH, whereas low values of α favour the maximisation of the ACEI. For each given value of α, a non-inferior solution was determined, i.e. a combination of T and ES ratio which maximise the objective function.
The results generated for the bi-objective optimisation when the enzyme was subtilisin can be seen in Fig. 3a, where the optimum values for the input variables (T and ES) as a function of α are represented. As α increases from 0 to 1, the optimum T increased from 52·7 to 60·5 °C following a convex curve. On the other hand, the whole set of non-inferior solutions corresponded to the maximum ES assayed (5%).

Fig. 3. Set of optimal values of temperature (solid line) and enzyme to substrate ratio (dotted line) for (a) subtilisin and (b) trypsin as a function of the weight factor.
The values for the output variables (DH and ACEI) associated to this bio-objective optimisation are shown in Fig. 4a, which is also known as Pareto front. The DH increased continuously from 0·213 for α=0 until attaining a maximum value of 0·218 for α=1, which coincided with the optimum value when DH is optimised alone. This curve presented an inflection point at α≈0·9, which involved a change from convex to concave. For ACEI, in contrast, a decreasing curve was obtained from its maximum value of 0·325 for α=0 down to 0·310 for α=1. Such descent was more pronounced for values of α higher than 0·6. As a consequence, within the range 0<α<0·6, the response DH could be improved (by raising the T up to 55 °C) while keeping the ACEI in the vicinity of its maximum value (ACEI=0·324). On the contrary, the slight increase in DH achieved within the interval 0·6<α<1 was detrimental to the ACEI. Moreover, this improvement was accompanied by a steep increase in T, which is undesirable from an economical point of view.

Fig. 4. Set of optimal values of degree of hydrolysis (solid line) and ACE inhibitory activity (dotted line) for (a) subtilisin and (b) trypsin as a function of the weight factor.
When this bi-objective optimisation approach was applied to trypsin, the optimum values for T and ES represented in Fig. 3b were calculated. Differently from subtilisin, where a continuous behaviour was computed, it can be observed that for trypsin there were only two optimum solutions which maximise the sum-weighted objective function. For any α<0·70, the optimisation problem converged to a single non-inferior solution which corresponded to the lowest T (30 °C) and ES (1%) assayed. When α is above 0·70, which indicates a rather high preference to DH, the optimum values for T and ES were the highest assayed (55 °C and 5%, respectively). With regard to the Pareto front (Fig. 4b), the values of DH and ACEI were related to the two optimum solutions separated at α=0·70. Indeed, DH took an optimum value of 0·095 from α=0 to α=0·70, and of 0·159 for α above 0·7. In the case of ACEI, optimum solutions corresponded to 0·399 (α<0·7) and 0·254 (α>0·7).
This work was funded by the project P07-TEP-02579 from the Consejería de Innovación, Ciencia y Empresa of Junta de Andalucía, Spain.







