


Introduction
In most tropical and sub-tropical countries, stalk-boring insects are of considerable economic importance as they reduce the yield of important food crops, including rice, sorghum, millet, maize and sugarcane (Polaszek, Reference Polaszek1998). Several species of Lepidoptera are responsible for the damage, mainly in the genera Chilo, Sesamia, Diatraea and Eldana. In South Africa, Eldana saccharina (Pyralidae) is considered the most important insect pest of sugarcane (Leslie, Reference Leslie and James2004). Besides being responsible for ongoing yield losses throughout most of that sugar industry, it has the potential to cause serious widespread damage, even the loss of entire fields of sugarcane (Atkinson, Reference Atkinson1981). As a result, extensive research has been conducted to identify efficient integrated control methods (Anonymous, 2005). Damage from E. saccharina increases with age of sugarcane, particularly after 12 to 15 months growth (Leslie, Reference Leslie1994). To avoid this, growers have been encouraged to reduce their crop cycle from 18 months (harvest to harvest) to around 12 months. This has resulted in reduced productivity, particularly in the rain-fed parts of the industry where a longer cropping cycle is favoured (Bezuidenhout et al., Reference Bezuidenhout, Singels and Hellmann2002); however, damage caused by E. saccharina is greatly reduced, generally to sub-economic levels. Insecticides have recently been adopted for use in sugarcane that will be ‘carried over’ the summer period, when the mills are closed, and that will be more than 12 months old when harvested (Anonymous, 2005). However, some growers are reluctant to use insecticides because of their secondary effect on natural enemies. According to Betbeder-Matibet (Reference Betbeder-Matibet1983), ants and spiders destroy 95% of each generation of E. saccharina. Despite this, and the long list of biological control agents that have been tested (Conlong, Reference Conlong1994), natural control is not sufficient to reduce numbers of the borer to an economically acceptable level.
Although the biology of the insect is well understood (Atkinson, Reference Atkinson1981; Atkinson & Carnegie, Reference Atkinson and Carnegie1989; Atkinson & Nuss, Reference Atkinson and Nuss1989; Keeping, Reference Keeping2006), less is known about its behaviour in the field over time; little is known about the way in which a newly planted area is colonized by the moth or about its subsequent colonization and distribution within the field in the following ratoon crops.
In this paper, we describe the distribution and extent of the damage caused by E. saccharina in sugarcane. The data were collected from field trials designed to measure crop loss from nematodes. This provided the opportunity to compare the damage caused by E. saccharina with that caused by plant-parasitic nematodes.
Material and methods
Sites and treatments
Data of stalk damage caused by E. saccharina were collected at harvest in eight plant cane crops and 22 ratoon crops at 12 nematicide trial sites in sugarcane fields in South Africa (table 1). These data were collected between 1995 and 2006 and came from 2180 plots. These measurements provided a summary of borer activity within the stalk for the duration of the crop. The trials were established in newly planted sugarcane and in ratoon sugarcane, with the various treatments at each site arranged in a randomized block design with six replicates.
Table 1. Characteristics of the nematicide trial sites monitored for E. saccharina damage.

PC, plant cane; R, ratoon.
Most of the plant crops were planted with setts (stalk cuttings) of certified (i.e. disease and borer free) seedcane that had no prior treatment. However, at site ZB, planting was done with setts that had been treated in a hot-water tank at 50°C for 30 min, a procedure that kills E. saccharina larvae within the stalk (Anonymous, 2005). Here, the treatments comprised soaking the hot water treated setts in suspensions of different bacterial strains, treatment with 3 kg ha−1 of the nematicide aldicarb, and an untreated control.
The AAPC and AAR trials both comprised treatments with various rates of anhydrous ammonia, urea, 3 kg aldicarb ha−1 and an untreated control. The LM22 and Z22 trials were designed to test various harvesting procedures and included a treatment with 3 kg aldicarb ha−1. The ZDO trial consisted of different rates of different nematicides, as well as 3 kg aldicarb ha−1 and an untreated control. Whereas only single cultivars were included in each of the six preceding trials, the other trials each compared the performance of 4–7 sugarcane cultivars planted and ratooned with and without 3 kg aldicarb ha−1.
Except for the PO trials, which were irrigated, all the trials were rain fed. The size of the trials varied between 3000 and 6500 m2. They were not isolated but situated within commercial fields, i.e. adjacent to normal commercial sugarcane, and, with a few exceptions, were harvested in synchrony with the surrounding sugarcane. Each trial comprised 54 to 96 plots each five or six rows wide, 8–10 m long, with a row spacing of 1 to 1.5 m; individual plot size ranged from 48 to 75 m2.
Data collection and analyses
At harvest, the centre three or four rows of each plot were cut and weighed. In addition, 10–20 millable stalks were selected at random from each plot and split lengthwise. Internodes damaged by E. saccharina were counted, together with the total number of internodes per stalk. To better analyze the damage caused by E. saccharina, a number of variables were calculated based on the damage that this pest causes to sugarcane stalks (Metcalf, Reference Metcalf, Williams, Metcalf, Mungomery and Mathes1969). From these data, we calculated the following parameters for each plot:
1. average number of internodes per stalk;
2. average percentage damaged internodes (=number of damaged internodes×100 divided by the total number of internodes);
3. average percentage damaged internodes per damaged stalk (=number of damaged internodes×100 divided by the total number of internodes of the damaged stalks only);
4. percentage damaged and undamaged stalks; and
5. average number of total internodes per stalk relative to the number of damaged internodes.
In the PO survey, levels of damage caused by E. saccharina were recorded in 20 commercial fields of 9–12-month-old sugarcane in the vicinity of the nematicide trial sites, PO1 and PO2. The survey was of 15 stalks per field, selected at random and conducted at the same time as that of the third ratoon of the two trial sites.
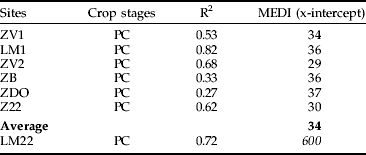
A ‘maximum Eldana damage index’ (MEDI) was determined for each site by fitting a regression for percentage internodes damaged and percentage undamaged stalks. The MEDI corresponds to the point at which there is 0% undamaged stalks in the trial (abscissa at origin). This index was calculated, separately, for the sugarcane in the untreated control plots and for sugarcane in plots treated with nematicide.
We assumed that the amount of damage caused by nematodes was equivalent to the difference between the cane yield of the untreated control and that achieved with a single application of the registered (economic) rate of aldicarb 3 kg ha−1, i.e. ((treated–control)×100/treated). Where more than one crop stage was studied at the same site, the nematicide treatment was repeated in each crop. At site ZV1, damage was also assessed by comparison with the yield achieved with ten monthly applications of aldicarb at 3 kg ha−1 in the plant crop and the following ratoons. The single application of 3 kg ha−1 is the commercial rate and, on sandy soils, normally provides an economic response. However, it gives only limited control of the nematodes. The repeated monthly treatment achieved almost complete nematode control (Berry et al., Reference Berry, Cadet and Spaull2004). Application of the nematicide was made in the furrow at planting and, for the repeated treatment, over the row thereafter. In ratoon sugarcane, the nematicide was applied over the row within 4–6 weeks after harvest of the previous crop, and, for the repeated treatment, at monthly intervals thereafter. It should be pointed out that the efficacy of the nematicide declines after application to a point that, three months after application, no active nematicide is detectable in treated soils. Thus, by the time of harvest (12 months after application of treatments) at a stage when borer pressure begins to increase, no nematicide is present (except where repeat applications were made). Details of the plant and ratoon trials discussed below are summarized in table 1.
Plant cane study
The percentage of undamaged stalks measured at harvest of the plant crop was projected on the map of the trial as normed values ((x−average x)/standard deviation), to highlight places on the grid with above and below average levels of infestation by E. saccharina. In the plots, the circles represent values of percentage undamaged stalks above average and squares, below average. The size of the symbol is proportional to the distance from the average.
Ratoon cane study
In some of the trials, E. saccharina infestation was monitored both in the plant crop and in the following ratoons or only in ratoon crops. Data collected from successive crops at the same trial site enabled the study of the distribution of the infestation from year to year.
Results
E. saccharina infestation of the plant crop
Distribution of E. saccharina infestation within trials
At harvest, when the sugarcane was 14–18 months old, of the 528 plots sampled, only one plot was free of E. saccharina. Using normed data of the percentage of undamaged stalks per plot projected on the trial maps, it was possible to locate below average infestations as represented by bigger squares and above average infestation as represented by circles (fig. 1). Within all the trial sites, infested plots occurred throughout the trial and not only on the periphery. This pattern occurred not only in trials situated in different fields but also in adjacent trials within the same field, e.g. Z22 and ZB (fig. 1). Plots with high levels of infested stalks at the bottom left corner of trial ZB were adjacent to plots with low levels of infested stalks in the top right of the neighbouring Z22 trial site.

Fig. 1. Distribution of the percentage of undamaged stalks (normed values) in the different sites in plant cane. ▪, plots with a low infestation level; ○, plots with a high infestation level; the size of the symbol is proportional to the distance from the origin. The arrow indicates that sites ZB and Z22 are located in this position on the same field, a few metres apart.
No difference in infestation distribution was apparent between trial ZB (planted with hot-water treated setts) and trial Z22 planted with setts, some of which showed symptoms of borer damage.
Level of E. saccharina infestation in trials
Aldicarb applied at 3 kg ha−1 at planting did not significantly influence E. saccharina infestation levels in plant crops (P>0.05) (table 2). Average percentage of internodes damaged for all the sites combined was 15.2%. On average, when stalks were attacked by the borer, 21.2% of the internodes were damaged. More than two-thirds (68.7%) of the stalks of plant crops were damaged. In the plots of the ZV1 trial that were treated with monthly applications of aldicarb (ZV1(*)), there was a 40% reduction in the proportion of internodes damaged and an increase in the proportion of undamaged stalks, from 19 to 57%.
Table 2. E. saccharina damage indices and (Avg) average internode numbers of the sampled stalks in plant cane, in (C) control and (T) aldicarb-treated plots.

* Repeated aldicarb treatment.
Maximum E. saccharina damage in trials
The percentage of damaged internodes in plant crops was always inversely proportional to the percentage of undamaged stalks and followed a log regression (fig. 2). The regressions for all sites were highly significant (table 3), except for the LM22 site. With the log regression model, where there are no internodes damaged, the percentage of undamaged stalks was close to 100%. The ZB site, which was the most infested (table 2), had a similar MEDI as several other sites (table 3). For all sites in plant cane, the average MEDI was 34%.

Fig. 2. Logarithmic model describing the relationship between percentage damaged internodes and percentage undamaged stalks for plant cane.
Table 3. Determination of the MEDI (maximum Eldana damage index) in plant cane with the log regression between percentage damaged internodes (x-axis) and percentage undamaged stalks (y-axis).
E. saccharina infestation of ratoon crops
Distribution of E. saccharina infestation within trials
Ratoon crops were harvested at 12 months and, as in plant crops, very few plots, 2% in rain-fed cane and 14% in irrigated cane, were free of the borer. Mapping of the distribution of the percentage of undamaged stalks per plot showed a marked change between the plant crop and the first ratoon (fig. 3). This was followed by a more stable distribution of the areas with low numbers of infested stalks (squares) and areas with high numbers of infested stalks (circles) from ratoon to ratoon (fig. 3). The distribution of percentage undamaged stalks in plant and ratoon crops was significantly correlated in only two of ten occasions; whereas within ratoon crops, it was significantly correlated in ten of 14 occasions (table 4).

Fig. 3. Distribution of the percentage of undamaged stalks (normed values) in plant cane and in successive ratoons. ▪, plots with a low infestation level; ○, plots with a high infestation level; the size of the symbol is proportional to the distance from the origin.
Table 4. Relationship between percentage of undamaged stalks in plant cane and ratoon and between ratoon crops using linear regressions.

Level of E. saccharina infestation in trials
As in plant crops, the single application of aldicarb in ratoons did not influence the level of E. saccharina damage. Similarly, the level of damage and the number of internodes bored was not influenced by the repeated application of aldicarb (ZV1(*)).
For both the treated and control plots, the percentage internodes damaged ranged between 2 and 10% with an average of 6% (table 5), much less than in plant cane (which ranged from 2 to 30% with an average of 15%, table 2). The percentage of internodes damaged per damaged stalk was 14% (approximately two internodes per stalk). In addition, the percentage of undamaged stalks in ratoon cane was twice that in plant cane. In the irrigated trials, all the indices were similar in the control and treated plots, although the percentage of internodes damaged was half that of rain-fed sugarcane.
Table 5. E. saccharina damage indices and (Avg) average internode numbers in ratoon cane, in (C) control and (T) aldicarb-treated plots.

* Repeated aldicarb treatment.
Maximum E. saccharina damage in trials
Unlike the situation in plant cane, linear regression was the best model to link the percentage of internodes damaged and the percentage of undamaged stalks in ratoon cane (fig. 4). The regressions were always highly significant (table 6). For theoretically zero E. saccharina damage, the ordinate at the origin was, as expected, close to 100% (77–96% undamaged stalks; table 6). The MEDI varied between 14 and 25%, with an average of 20% (table 6). It was lower in the irrigated trials (average 16%), which was similar to the 14% MEDI calculated from the survey of the 20 commercial fields on the same farm (all irrigated; data not shown). On average, the MEDI of the ratoon crops was half that of the plant crops.

Fig. 4. Linear model describing the relationship between percentage damaged internodes and percentage undamaged stalks for ratoon cane.
Table 6. Determination of the MEDI (maximum Eldana damage index) in ratoon cane with the linear regression between percentage damaged internodes (x-axis) and percentage undamaged stalks (y-axis).

Change in E. saccharina infestation with crop stage
To analyse changes with ratooning, the different parameters were averaged across sites and treatments. The distribution of the points for percentage internodes damaged per damaged stalk and average number of nodes per stalk were very similar to percentage internodes damaged and the MEDI – data not shown. Similarly, the percentage undamaged stalks per crop was also similar except that the points were inverted, as expected. For percentage internodes damaged and MEDI, there was a sharp drop between plant cane and ratoons (fig. 5). Plotted values in ratoons increased slowly up to the 4th ratoon. But, when all ratoon stages were used, including R4, R9 and R10, which were from single observations, the indices appeared to remain stable as judged by the fact that the linear regression model was almost parallel to the x-axis (the slope of the regression was not significantly different from 0).
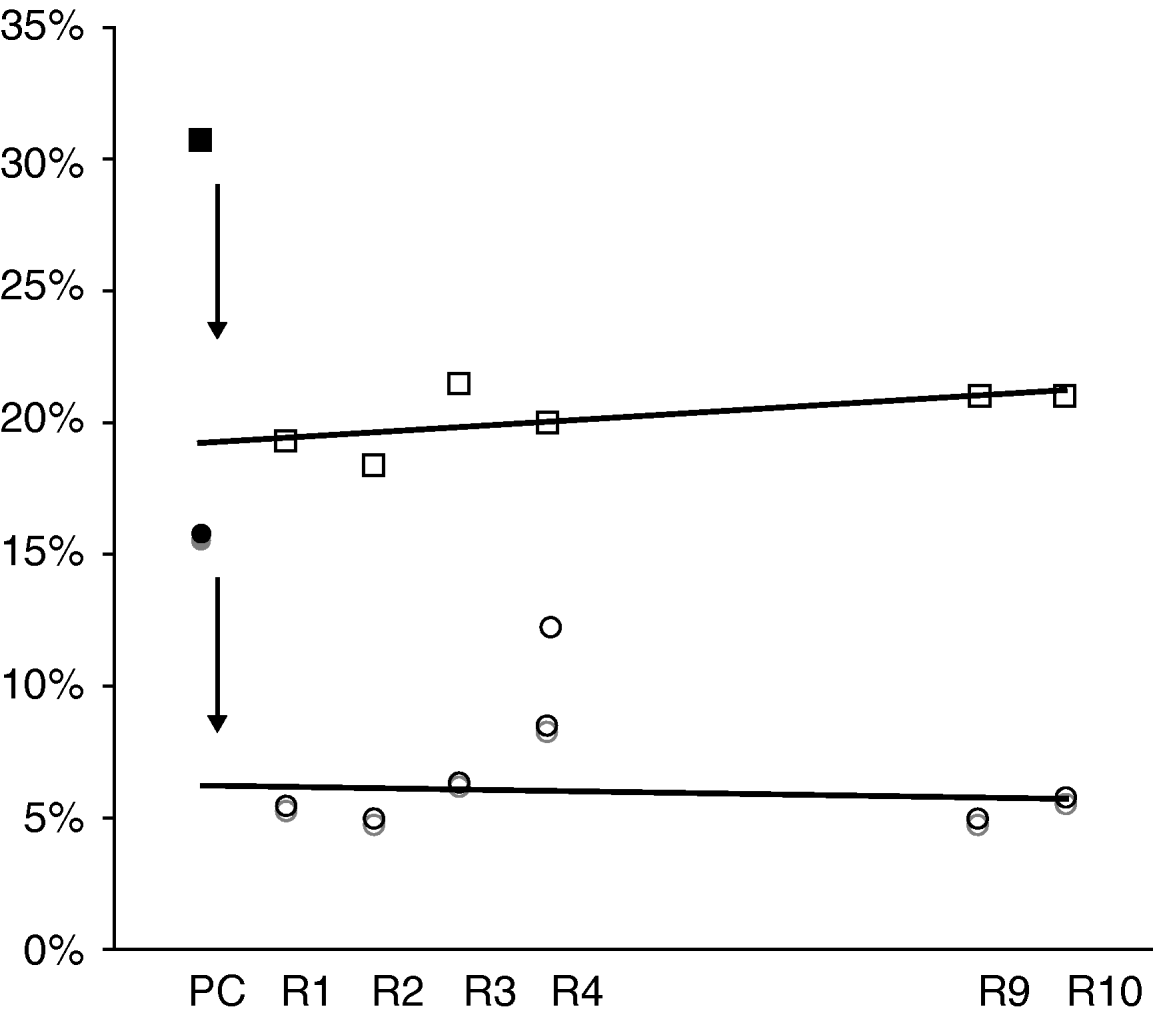
Fig. 5. Change, with crop stage, of levels of E. saccharina damage, expressed as the percentage of stalks bored and ‘maximum Eldana damage index’ (MEDI). The regression lines were drawn based on data for ratoons only. PC, plant cane; R, ratoon (○, % damage; □, MEDI (%)).
Relationships between internodes damaged and number of internodes per stalk
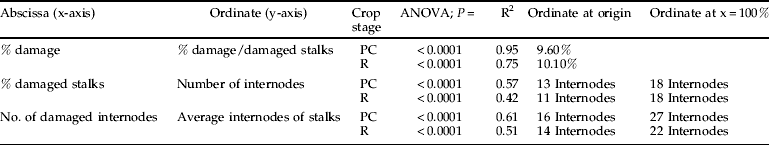
When the percentage of internodes damaged is plotted against the percentage of damage per damaged stalk and not the percentage of damaged stalks, which also includes undamaged stalks, the linear regression is highly significant (table 7). The model predicts there would be 9.6 and 10.1% internodes damaged per damaged stalk in plant cane and ratoon cane, respectively, if no stalks were damaged (ordinate at origin).
Table 7. Relationship (linear regression) between E. saccharina damage indices and between the number of internodes damaged and the average total number of internodes of the corresponding stalks.
PC, plant cane; R, ratoon.
The percentage of damaged stalks was directly related to the number of internodes per stalk. Where stalks were undamaged, the linear regression predicted that there would be 13 internodes per stalk in plant cane and 11 internodes per stalk in ratoon cane. The linear regression also predicted that, if stalks had the potential to grow 18 internodes long, they would all be infested in plant and ratoon crops. However, based on the significant linear relationship between the number of internodes damaged and the number of internodes per stalk, these values were slightly higher: namely, 16 and 14 internodes for no damage in plant and ratoon cane, respectively, and 27 and 22 internodes for 100% damaged internodes (table 7).
Comparison with nematode damage
At most of the sites, 80–90% of the nematode community comprised a mixture of Helicotylenchus dihystera, Meloidogyne javanica, Xiphinema elongatum, Pratylenchus zeae, Scutellonema spp. and Paratrichodorus spp. The combined damage caused by these nematodes, as measured by the difference between the cane yields recorded from the untreated control plots and from plots treated with 3 kg aldicarb ha−1, ranged from 8–89% in the plant crops and −1 to 114% in the ratoons (table 8). The overall average of 35% was four times greater than the actual Eldana damage of 8% on these sandy soils. The maximum damage caused by nematodes, as measured by the yield response to the repeated application of aldicarb at the ZV1(*) site, averaged 89% for plant and ratoons crops. Assuming that E. saccharina reduces sugarcane yield by 1% per 1% internodes bored (Goebel & Way, Reference Goebel and Way2003), then the maximum yield loss averaged 22% for the same soils (table 8).
Table 8. Actual and maximum potential damage per ha by E. saccharina and nematodes at the different sites.

* Maximum nematode damage in parenthesis.
Discussion
Distribution of E. saccharina
After emerging from their pupal case, female E. saccharina moths fly just above the sugarcane canopy where they are attracted by displaying males. The males often occur in groups on a plant, which is usually one of the taller in the stand (Atkinson, Reference Atkinson1981). The mated females lay about 400 eggs in batches of approximately 20 (Way, Reference Way1995). The oviposition sites include cryptic places in amongst dead leaf material at the base and lower third of the sugarcane stalk (Leslie, Reference Leslie1990). After approximately one week, the first-stage larvae hatch from the eggs and initially scavenge on the outside of the stalk before boring into the stalk (Leslie, Reference Leslie1993). Most of the borings by the larvae occur in the lower half of the stalk. E. saccharina is less frequently found in the upper, less mature part of the stalk.
Our study shows clearly that during the 14–18 months after planting, E. saccharina was able to colonize most of the field, with about two-thirds of the stalks being infested. The borders of the trial sites were no more infested than the inside, as might be expected from a progressive lateral colonization by a relatively poor flyer such as E. saccharina. Various hypotheses could explain the widespread infestation of the plant crop. One is that the borer was carried to the field with infested seedcane. This would explain the erratic distribution of heavily infested patches in the trials, but does not explain the higher level of infestation in the ZB trial where the seedcane was hot-water treated before planting; such treatment is sufficient to kill larvae inside the stalks (Anonymous, 2005). Moreover, certified and non-certified seedcane ended up with a similar infestation distribution.
An alternative explanation is that direct infestation of the new stalks took place with E. saccharina emerging from borings in the stubble of the old stools left in the ground from the previous crop. This is a possibility, as attempting to kill the old crop by ploughing out the stools or by applying glyphosate to the regrowth is, in practice, rarely completely effective.
However, a more likely explanation is that there were both invasions from outside the field plus internal spread of the borer within the field. Since sugarcane was also growing in the surrounding area, moths present in neighbouring fields could, helped by the wind, fly into the field where the trial was situated. The duration of the life cycle of E. saccharina is extremely variable (Atkinson & Carnegie, Reference Atkinson and Carnegie1989) with a generation time of approximately eight weeks (Leslie, Reference Leslie and James2004). During this time, the initial colonizers could reproduce over six generations and spread throughout the field during the plant crop. This scenario, with or without incursions from outside the field, would explain what we observed in the trials.
Mechanisms of invasion
Measuring the number of bored internodes at the time that the sugarcane was harvested provided a summary of borer activity that occurred over the duration of the crop. However, the linear regression model that we derived showed clearly that damage to the internodes in the plant crop occurred, on average, in stalks with 16 or more internodes at harvest. The highest percentages of damaged internodes were found in stalks with a high number of total internodes, that is stalks with a high growth potential. The formation of nodes in the sugarcane stalk, which are the points at which leaves are attached, is regulated by a number of factors, including temperature, time, light intensity, soil moisture and soil nutrients (Inman-Bamber et al., Reference Inman-Bamber, Bonnett, Singels and Thorburn2005). Where local environmental conditions favour growth, the sugarcane stalks will have more nodes, and thus internodes, and will be taller than stalks growing under less favourable conditions (Inman-Bamber & Smith, Reference Inman-Bamber and Smith2005). We infer from this that E. saccharina bored into stalks that were, or were destined to be, taller than other stalks. This may occur by chance or may be because the female may lay eggs at random on and around the sugarcane stalks or she, and/or her offspring, may select taller or higher potential stalks. The second scenario would suggest that the female and/or her progeny are able to identify stalks with potential to be taller. However, in either situation, the survival of the emerging neonate larvae, and of subsequent larval stages, will tend to be greater on healthier, more nutritious stalks, for example in stalks with higher levels of nitrogen, as was observed by Atkinson & Nuss (Reference Atkinson and Nuss1989) .
During the tillering phase in the development of the sugarcane crop, stalk density increases to a maximum around 3–8 months, depending on time of planting and temperature, before intra-row competition eliminates a number of the stalks and the density stabilizes at a level characteristic of the cultivar (van Dillewijn, Reference van Dillewijn1952). The death of shorter, less competitive stalks would have little effect on the E. saccharina population if most larvae occur in the better-grown stalks. However, even if E. saccharina did occur in these shorter, less competitive stalks, evidence of their presence would mostly be lost before harvest.
Since it appears that E. saccharina bores into stalks that have more internodes, it could be inferred that the insect itself, in some way, promotes the formation of nodes and perhaps increases stalk length. Growth stimulation can occur in response to low levels of insect damage (Harris, Reference Harris1974), but such an interaction appears unlikely based on the work of King (Reference King1989), who found an apparent inverse relationship between stalk length and the number of internodes damaged per stalk.
The relationship between the percentage of stalks damaged and percentage of internodes damaged showed that, initially, in plant cane, stalks were damaged at a greater rate than internodes, but as damage to the former increased beyond 70 or 80%, so there was an exponential increase in the latter. This may, in part, merely reflect the increased mathematical chance that an individual will bore into an already bored stalk, or that there is a synergistic effect, where one individual attracts others. It is also conceivable that the accumulation of larvae in the stalks could also be in response to volatiles emanating from isolates of Fusarium spp., within the borings, that are attractive to the larvae on the outside of the stalk (McFarlane & Rutherford, Reference McFarlane and Rutherford2006). Such isolates commonly occur in the borings of E. saccharina in sugarcane, and some have a marked beneficial effect on the development of the larvae (McFarlane & Rutherford, Reference McFarlane and Rutherford2005). In addition, ovipositing female E. saccharina may be attracted to Fusarium infected stalks, as was found with maize in West Africa (Ako et al., Reference Ako, Schulthess, Gumedzoe and Cardwell2003).
The physical and chemical nature of the sugarcane soils in South Africa vary within fields, over short distances (Cadet et al., Reference Cadet, Berry and Spaull2004), with the result that areas more and less conducive to sugarcane growth commonly occur within a field. Thus, the preferred sites favoured by E. saccharina may be places where the levels of N were higher in the sugarcane. It seems paradoxical to state that E. saccharina infestations appear to be an indicator of strong plant growth potential.
Regulation of E. saccharina in ratoon cane
The distribution of E. saccharina could be predicted with greater confidence from one ratoon crop to the next, than from the plant crop to the first ratoon. This meant that areas favourable and unfavourable for E. saccharina within a sugarcane field, changed after the plant crop but then remained much the same from one ratoon crop to the next. It suggests that factors affecting the spread of the borer in the plant crop may not apply to ratoons. These observations, together with the lower infestation levels and the linear model of damage distribution, suggest that there are regulatory processes in the ratoons that are not present in the plant crop. The presence of favourable sites within a variable habitat was thought to explain the distribution of the European corn borer, Ostrinia nubilalis, along the borders of maize fields in the south of France (Bailey et al., Reference Bailey, Bourguet, Le Pallec and Ponsard2007). Evidence for the presence of such sites in sugarcane fields could be explained by: (i) the reasonable assumption that a specific microenvironment favours the abundance of E. saccharina; (ii) that these occur in certain areas of the field; and/or (iii) the presence of E. saccharina males within these favoured areas that call to female moths flying nearby (Atkinson, Reference Atkinson1981).
A notable difference between a plant and a ratoon cane field is that with the former the soil is disturbed when the furrows are drawn. Thus, the much lower infestation levels, as measured by the proportion of undamaged stalks in the ratoon crops, could result from the existence of a soil environment that, for each successive ratoon, remained undisturbed for longer and longer periods. Such conditions would favour many predators of E. saccharina, in particular soil-dwelling ants (Leslie & Boreham, Reference Leslie and Boreham1981). The cane in all the trials was harvested after burning off the dead leaves and leaf sheaths. Planting the trials was done at least two months after the previous crop was burnt and harvested; thus, while burning the dead leaves at harvest would affect predators more in ratoons than plant crops, subterranean colonies of ants would soon recover from the loss of foraging individuals. All predators would be affected by the soil tillage conducted before planting. The occurrence of a regulatory process (biotic or abiotic) was also suggested by the linear regression between percentage internodes damaged and percentage undamaged stalks. This process seemingly prevented the exponential increase in damaged stalks and further damage to internodes, despite ongoing invasion and internal re-infestation, similar to that assumed for the plant crop.
Nematicide effect and regulation process
Although aldicarb is an efficient insecticide/nematicide, when applied at the registered rate, it had no significant depressive effect against E. saccharina. This is not surprising as the chemical is more effective against sap suckers than stalk borers, judging by how few stalk borers are included in the lists of pests for which aldicarb is registered (Anonymous, 2008). In addition, aldicarb is soluble in water and is effective for about six weeks (Hague & Gowen, Reference Hague, Gowen, Brown and Kerry1987); so, E. saccharina that invade the shoots when they are several weeks old would do so after the chemical had broken down. As expected, the monthly repeated treatment, applied over the row after the first month in plant cane, reduced, significantly, the level of damage caused by E. saccharina; but the same repeated treatment was inefficient in ratoon cane. The efficiency of aldicarb cannot be questioned, as almost all the nematodes were killed in these plots in both the plant cane and the ratoon (Berry et al., Reference Berry, Cadet and Spaull2004). We infer that the killing of E. saccharina in plant cane did not result from a systemic effect, as the same should have occurred in ratoon cane.
With these results, we have deduced a possible invasion process by E. saccharina in plant and ratoon cane. In plant cane, disturbance of the soil prior to planting disrupts the environment to the disadvantage of many probable predators of E. saccharina (Cherry, Reference Cherry2003). Soon after planting, invading E. saccharina moths land throughout the habitat and lay eggs on dead leaves on many of the young shoots. When in contact with the soil, the moths could be killed by the aldicarb that had been applied to the soil surface as part of the repeated monthly treatment. This would explain why the repeated treatment was so efficient in the plant crop. The absence of a noteworthy effect on the borer from the repeated treatment with aldicarb in ratoon cane may be due to: (i) the large amount of plant debris covering the soil after harvest; and/or (ii) the impact of the chemical on predators, which in ratoon cane, were presumed to have played an important role in reducing the number of E. saccharina. When the aldicarb was applied over the row, and where the plant debris was dry, the granules mostly fell through the litter layer onto the soil and would not come into contact with moths alighting on the debris. In contrast, aldicarb on the soil surface, beneath the litter, could have a considerable effect on foraging predators. Moreover, in ratoon cane a substantial residual infestation of the borer can occur within the stubble of the previous crop (Anonymous, 2005), which would be protected from the aldicarb effect. This could also explain why the repeated treatment was ineffective in ratoon cane. The limited expansion of the infestation observed in ratoon cane may possibly be explained by the attractiveness of the environment of the stools that harboured previous and overlapping generations of E. saccharina. The reduction in the percentage of stalks that are damaged in the ratoon crop can be explained by the failure of the borer to colonize certain less favourable areas, as a result of predator activity, or the existence of areas with a repulsive abiotic environment or simply because adjacent areas were more attractive. These assumptions require investigating, as they may help identify the basis of the unfavourable areas, which might then be used to promote this phenomenon in plant cane.
Change in borer infestation with crop stages
Data from several old ratoon crops from the same site are lacking, which makes prediction of the change in infestation with increasing number of ratoons less reliable. However, the available data, from 1st, 2nd and 3rd ratoons, each from five or more sites, to 9th and 10th ratoons from one site only, indicate that after the sharp decline in the level of damage from the plant crop to the 1st ratoon, average damage levels remained remarkably similar, with about 7% internodes damaged. The reduction in crop age at harvest might explain the difference in the infestation levels between the plant crops and the ratoons; whereas the former were cut at an average age of 16 months (14–18 months), the latter were cut at 12 months. Even though the shorter crop duration effectively reduced stalk damage, it probably did not influence incursion by the moths, other than reducing the opportunity for this by about four months.
Damage and threshold detection level
Even if 100% of the stalks were attacked, the proportion of internodes damaged would not reach 100% because E. saccharina would not be able to bore all the internodes before harvest and because some internodes are not palatable. In fact, the linear model describing the relation between percent damage per damaged stalk and the average percent internodes damaged, including healthy stalks, should cross the origin. When E. saccharina is not present, no stalks should be damaged. But the regression line did not cross the origin. The projected 10% stalks damaged represents the experimental error, or threshold detection level, with the sampling procedure. Collecting 20 stalks from 30 m of row represents only about 7% of the millable stalks harvested from each plot.
Interaction between nematodes and E. saccharina
Damage caused by E. saccharina is greater in drought-stressed sugarcane than unstressed sugarcane (Atkinson & Nuss, Reference Atkinson and Nuss1989). With this understanding of the relationship between the borer and sugarcane, it appears that plant parasitic nematodes could interfere with both the selection of the egg laying sites and the feeding in the stalk. During the selection phase, nematodes would be expected to reduce the prevalence of E. saccharina in the field because sett root damage to newly planted sugarcane, caused by nematodes, slows down the development of the bud and primary shoot emergence (Cadet & Spaull, Reference Cadet and Spaull1985). This would affect the subsequent appearance of the first dead leaves, the axils of which are a favoured oviposition site for E. saccharina (Leslie, Reference Leslie1990). Nematodes increase inter-tiller competition and reduce the number of stalks per ha. However, calculated as a percentage of stalks attacked, the proportions should remain the same as in the nematicide treated plots, except for the repeated treatment; but, in this case, it was suspected that the chemical was directly controlling E. saccharina. The consequence of this delay in the germination process is a reduction in the duration of the contact between stalk and insect and, thus, less insect damage.
Impact of both pests on sugarcane in South Africa
The field trials were all on poor sandy soils where nematodes are known to be a growth constraint to sugarcane. The estimated yield losses, averaging 35%, were very much greater than those due to E. saccharina. Estimates of maximum yield losses emphasize this observation. While nematodes also affect yield of cane on finer textured soils (Cadet & Spaull, Reference Cadet, Spaull, Luc, Sikora and Bridge2005; Blair & Stirling, Reference Blair and Stirling2007), the actual loss is usually smaller and less widespread, but nevertheless, significant. This contrasts with the losses due to E. saccharina which are not confined to soil texture. Thus, while nematodes present a problem to sugarcane locally, in certain fields, E. saccharina moves easily between fields and between farms and has the potential to increase to devastating levels if not controlled (Atkinson & Carnegie, Reference Atkinson and Carnegie1989). While industry-wide losses from nematodes and from E. saccharina may be similar, the perception by sugarcane growers, and other decision makers, of nematodes and E. saccharina as pests of sugarcane is very different. This misperception of the importance of nematodes has long been known (Bridge, Reference Bridge1996). Several reasons can explain it; the most important of which are: (i) the absence of diagnostic symptoms; (ii) the confusion between a plant suffering from nematodes as opposed to nutrient or water deficiency; (iii) the absence of places naturally free of nematodes; and (iv) the lack of a chemical treatment, efficient in all soils, to highlight their overall importance (Sharma et al., Reference Sharma, Price and Bridge1997). Conversely, symptoms of damage by E. saccharina are immediately visible above ground, at eye level and easily quantifiable and can be compared to a stalk free of damage collected in the immediate vicinity. The two facets of E. saccharina as a pest of sugarcane, that separate it from plant parasitic nematodes, are that it moves easily between fields and between farms and that it has the potential to increase to devastating levels if not controlled. It, therefore, requires area-wide management rather than control at a field scale.
The detection of a biotic or abiotic factor that influences the distribution of E. saccharina in ratoon sugarcane is of considerable importance. Identification of the factor should lead to the development of improved control options, not only in ratoon crops but also in plant cane, where it is presumably destroyed by the replanting practice. A strategy that focused on reducing the invasion of egg laying adults would enable an increase in crop duration (and, thus, crop yield) more so than one that concentrated on reducing the period that the larvae feed in the stalk.













