1. Introduction
Evidence from cryptospores and cuticule-like fragments indicates that embryophytes had evolved by Darriwilian times (Middle Ordovician; Strother, Al-Hajri & Traverse, Reference Strother, Jansonius and Mc Gregor1996). According to Steemans et al. (Reference Steemans, le Hérissé, Melvin, Miller, Paris, Verniers and Wellman2009), the origin of vascular land plants, indicated by the occurrence of hilate/trilete spores, dates back to the Katian, more than 20 million years after the colonization of the land by bryophyte-like plants. However, vascular plant megafossils appear later in the fossil record: the earliest fertile axial land plant fossil is reported from the Homerian (Middle Silurian; Edwards & Feehan, Reference Edwards and Feehan1980). The Lower Devonian plant-bearing deposits record some of the most striking changes occurring in the evolution of plants; by Emsian times, most major plant lineages were probably already established (Steemans et al. Reference Steemans, Pétus, Breuer, Mauller-Mendlowicz, Gerrienne and Talent2010). The occurrence of deposits of this age is therefore of great importance for our knowledge of early land plants.
A few plant-bearing deposits have been reported from Devonian sequences of the Armorican Massif, but they have not yet been studied in detail. The Armorican Massif is a small part of the Variscan belt that extends throughout western and central Europe (Ballèvre et al. Reference Ballèvre, Bosse, Ducassou and Pitra2009). It includes four domains, namely the Léon domain and the North, Central and South Armorican domains (Ballèvre et al. Reference Ballèvre, Bosse, Ducassou and Pitra2009). The South Armorican domain comprises the sedimentary series of the Chalonnes area. This series shows plant-bearing deposits within two distinct stratigraphic levels that are located below and above marine limestones (Le Maître, Reference Le Maître1934). Plants occurring in these deposits are evidence of the proximity of land and, in addition to their palaeobotanical interest, they provide valuable information for palaeogeographic reconstruction. The upper plant level, the Sainte-Anne Formation, was first reported by Bureau et al. (Reference Bureau, Bureau, Davy and Dumas1908). It has been recently reinvestigated and has been given an Emsian age (Ducassou et al. Reference Ducassou, Strullu-Derrien, Ballèvre, Dabard, Gerrienne, Lardeux and Robin2009; Ballèvre et al. Reference Ballèvre, Ducassou, Lardeux and Régnault2010). The lower plant level was observed for the first time in 1969 by one of us (H.L.). It is mainly characterized by a rich and promising plant fragment content and by spherical quartz grains attributed to an aeolian sedimentation (Cavet et al. Reference Cavet, Arnaud, Barbaroux, Blaise, Brosse, Chauris, Gruet, Lardeux and Rivière1970; Cavet, Lardeux & Phillipot, Reference Cavet, Lardeux and Phillipot1971; M. Dubreuil, unpub. thesis, Univ. Nantes, 1986). The age of this plant-bearing level is still a matter of debate. On the basis of the stratigraphic succession, Cavet et al. (Reference Cavet, Arnaud, Barbaroux, Blaise, Brosse, Chauris, Gruet, Lardeux and Rivière1970) and Cavet, Lardeux & Phillipot (Reference Cavet, Lardeux and Phillipot1971) suggested an age ranging from Silurian to Early Devonian. On miospore evidence, McGregor (unpub. report no. F1-15-1980-DCM, 1980) tentatively derived a late Pragian (Lower Devonian)–Eifelian (Middle Devonian) age.
Here we provide a sedimentological and palaeontological analysis of the lower plant-bearing level. The main aims of the study are: (1) to confirm the age of the deposits, (2) to characterize the depositional environment, and (3) to report and briefly describe the flora still under investigation. The plant megafossils from these deposits are placed in a chronological overview of the Devonian flora of the Armorican Massif.
2. Geological setting
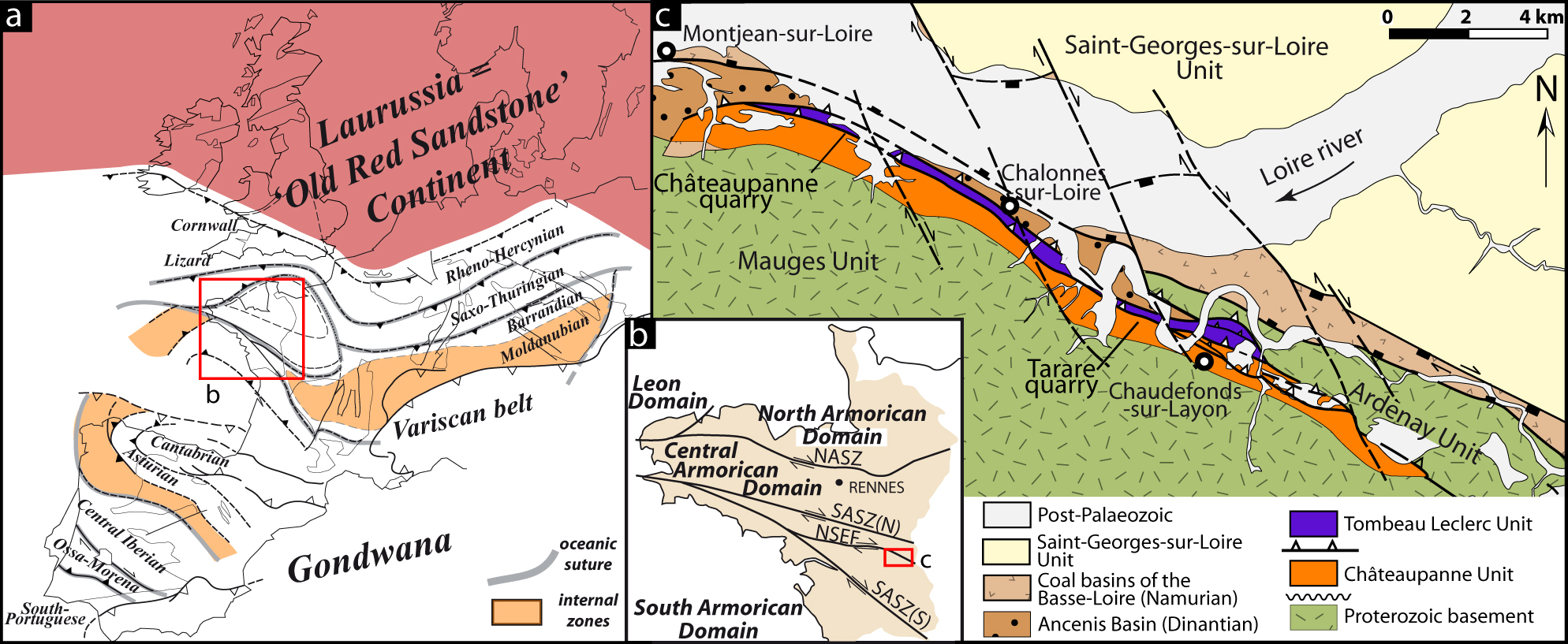
The Armorican Massif is a small part of a Late Palaeozoic mountain belt, the Variscan belt, that resulted from the convergence of Laurussia and Gondwana (Fig. 1a). It is divided by late Carboniferous transcurrent shear zones into four domains: the Léon domain, and the North, Central and South Armorican domains (Fig. 1b). The South Armorican Domain is located south of the South Armorican Shear Zone. This major fault is divided in two branches (SASZ(N) and SASZ(S) in Fig. 1b). Between these two branches, the Nort-sur-Erdre Fault (NSEF in Fig. 1b) separates the northern Lanvaux Domain from the southern Mauges Domain (Ballèvre et al. Reference Ballèvre, Bosse, Ducassou and Pitra2009). In the latter, the Mauges Unit comprises deformed Proterozoic sediments unconformably covered by Cambrian sediments on its southern part (Cavet, Gruet & Pillet, Reference Cavet, Gruet and Pillet1966; Thiéblemont et al. Reference Thiéblemont, Guerrot, Le Métour and Jézéquel2001) and by Ordovician to Devonian sediments on its northern part. These northern Palaeozoic sediments are structured in two main units separated by a southward thrust (Figs 1b, 2) (Ducassou et al. Reference Ducassou, Strullu-Derrien, Ballèvre, Dabard, Gerrienne, Lardeux and Robin2009):

Figure 1. Location (rectangle) of the study area within the European Variscan belt (a) and the Armorican Massif (b). NASZ – North Armorican Shear Zone; SASZ(N) and SASZ(S) – South Armorican Shear Zone, northern branch and southern branch, respectively; NSEF – Nort-sur-Erdre Fault. (c) Structural map of the studied area. A colour version of this figure is available at http://journals.cambridge.org/geo.

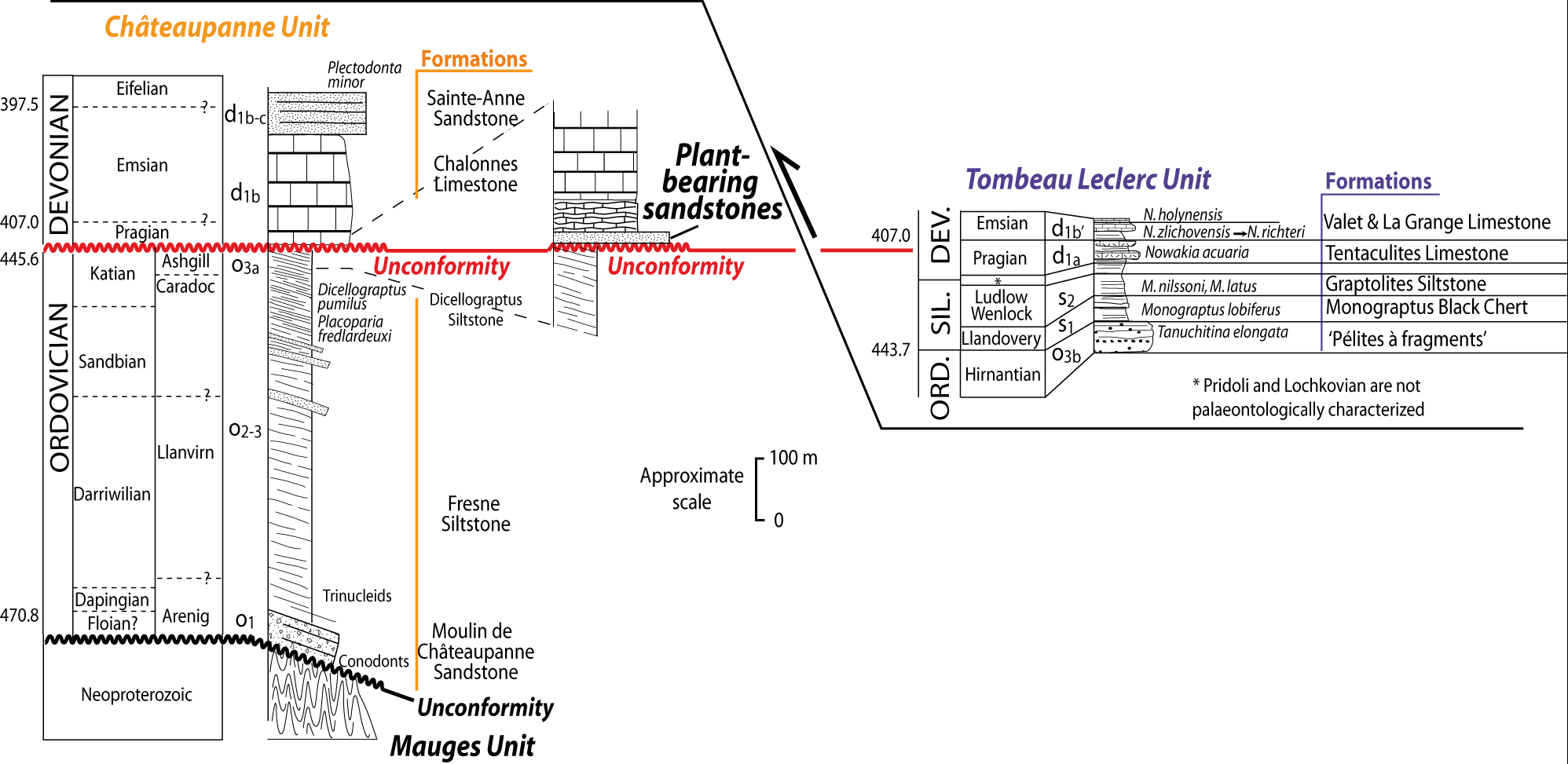
Figure 2. Lithostratigraphic columns of the Châteaupanne and Tombeau Leclerc units. A colour version of this figure is available at http://journals.cambridge.org/geo.
(1) The southern unit, the Châteaupanne Unit (Fig. 2) (Cavet, Lardeux & Phillipot, Reference Cavet, Lardeux and Phillipot1971), consists of Ordovician marine sediments (Moulin de Châteaupanne Sandstone and Fresne Siltstone) unconformably overlain by the Emsian Chalonnes Formation. This Formation, predominantly composed of massive limestone (Le Maître, Reference Le Maître1934), shows at its base terrigeneous layers with occurrences of plant fragments (= Basal Member of the Chalonnes Formation). Conformably overlying the limestone is the Sainte-Anne Formation of Emsian age.
(2) The northern unit, the Tombeau Leclerc Unit (Fig. 2), consists of an inverted Hirnantian (Upper Ordovician) to Emsian condensed sequence (Ducassou et al. Reference Ducassou, Strullu-Derrien, Ballèvre, Dabard, Gerrienne, Lardeux and Robin2009).
3. Materials and Methods
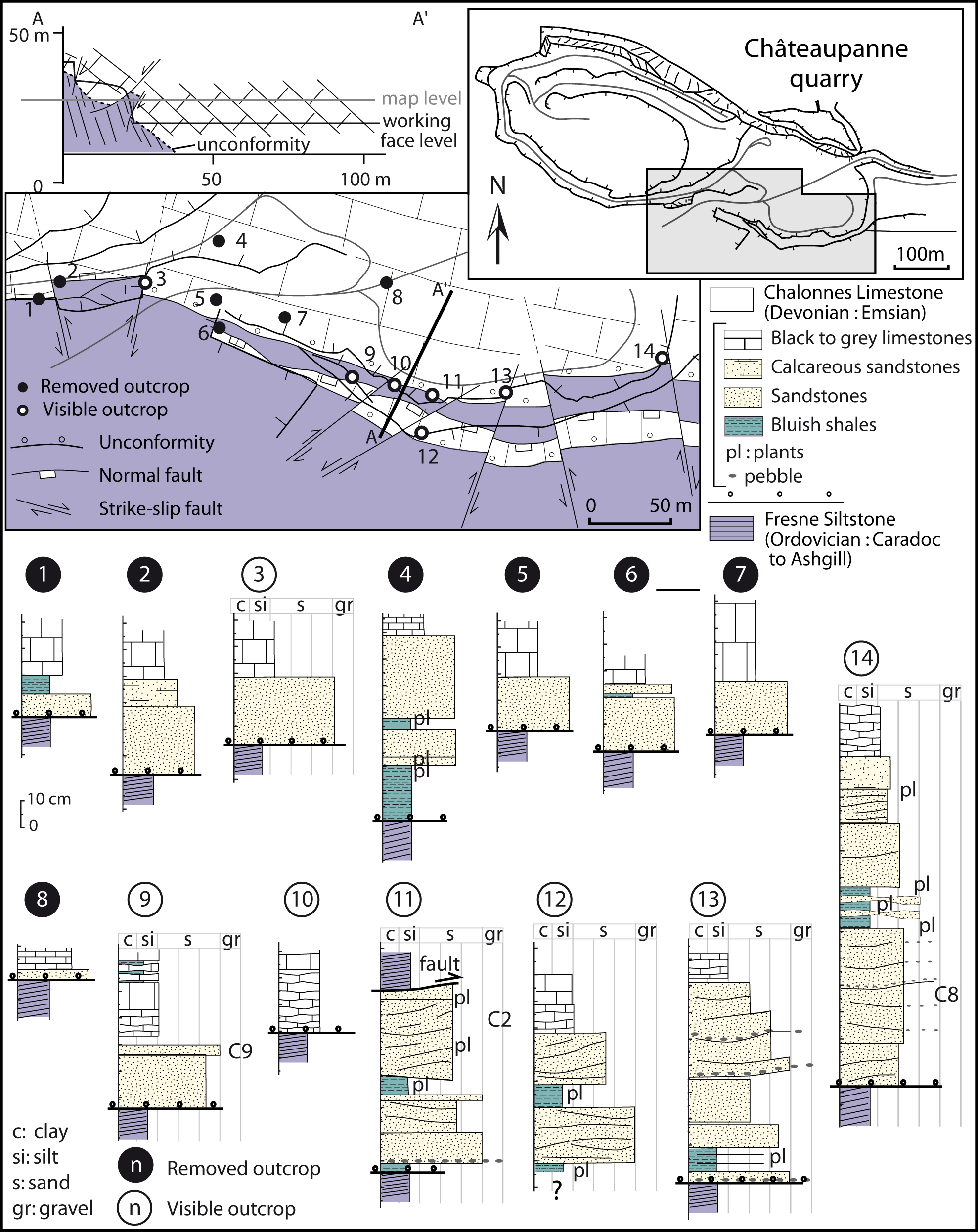
The rock samples were collected from the plant-bearing deposits that crop out at seven places (to date) in the Châteaupanne quarry (Montjean/Loire, Maine et Loire, France); their locations are given in Figure 3. Plant macrofossils described here were collected from localities 11, 12 and 13; samples for palynological study were collected from localities 11, 13 and 14.

Figure 3. Schematic map of the southern working face of the Châteaupanne quarry (location shown on Fig. 1b), established at a nearly 30 m level as schematized on the cross-section A–A′. The position of the logs detailed below is reported on the map. Positions of samples C2, C8, C9 are shown on the logs. For the removed outcrops, the granulometry reported on the logs is only schematic. A colour version of this figure is available at http://journals.cambridge.org/geo.
The plant specimens and the sections were observed using a Zeiss photomacroscope with a polarized light source. Approximately 1.5 mm thick sections were prepared through each pyritized axis embedded in resin. These sections were mounted on glass slides with epoxy resin and ground to a thickness sufficient for examination under the photomacroscope. The method described by Kenrick (Reference Kenrick, Jones and Rowe1999) was used for etching the sections before examination.
Five samples were processed for palynological study in the Paléobotanique, Palynologie et Micropaléontologie Unit at the University of Liège. They were prepared using standard palynological acid maceration techniques. The organic matter required oxidation in Schultz's solution during two hours. Slides of the samples were examined using standard light microscope analysis. England-Finder coordinates were used to locate the specimens on the palynological slides.
Specimens and palynological slides are housed respectively in the Laboratoire Mycorhizes of the University of Angers (France) and in the Paléobotanique, Palynologie et Micropaléontologie unit of the University of Liège (Belgium).
4. Results
4.a. Sedimentology and petrography of the sandstones
The basal unit of the Chalonnes Formation is rarely visible because of the poor outcrop, but it is well displayed along the southern working-face of the Châteaupanne quarry (Montjean-sur-Loire) (Fig. 1c). It is also present in the Tarare quarry (Blaise et al. Reference Blaise, Arnaud, Brossé, Cavet, Depagne, Gruet, Lardeux and Limasset1986) (Fig. 1c) at the bottom of the Chalonnes Formation, but the contact with the underlying Fresne Formation has not been observed.
4.a.1. Facies description
The Basal Member of the Chalonnes Limestone is located just above the unconformity. This level is generally made up of a single sandy bed, 10 to 20 cm thick (Fig. 3), but it can be absent. Locally (e.g. 14 in Fig. 3), it reaches more than 1 m and is then composed of fine- to coarse-grained sandy layers interbedded with bluish shale. In the sandy beds hummocky cross-stratification (HCS) and swaley cross-stratification (SCS) are visible (Fig. 4a). These bedforms are characterized by gently curved and often erosional bases, and by laminae that are nearly parallel to the lower bounding surface; the latter thicken or get thinner laterally. The HCS shows a high rate of amalgamation (Fig. 4a) that records a low rate of preservation. Very coarse-grained sandy beds or lenses with pebbles and shaly clasts are present just above the unconformity and sometimes interbedded in the sandstone. The plant fragments are contained in the shale intercalations. The thickness of these micaceous shaly layers is generally less than 1 cm but can locally reach 20 cm. Bioturbation is rarely present.

Figure 4. (a) Outcrop of the Basal Member of the Chalonnes Formation that unconformably overlies the Fresne Siltstone showing hummocky cross-stratifcations (log 14 on Fig. 3), scale bar = 20 cm. (b) General view of the sandstone petrography showing well-rounded quartz grains and laminae, PPL (sample C2, Fig. 3); scale bar = 2 mm. (c) Photography of the sand grains under stereomicroscope (fraction > 160 μm, < 500 μm on sample C9, Fig. 3) showing mainly round and matte grains (RM). Some angular grains (NU), aggregates (Ag.) of quartz grains with matrix and argillite or siltite fragments (Si) are also present. Scale bar = 500 μm. A colour version of this figure is available at http://journals.cambridge.org/geo.
4.a.2. Mineralogical analysis
Petrological and morphoscopic studies have been performed on samples from Châteaupanne quarry (samples C2, C8 and C9; Fig. 3). The sandstone consists of well-rounded detrital quartz (>95%), mainly monocrystallline, with some rare argillite fragments (Fig. 4b). Dark and light laminae are defined by the relative abundance of argillite fragments (Fig. 4b). The grain size ranges between 100 and 200 μm in fine-grained laminae, and between 300 and 500 μm in coarse-grained laminae. Some coarser grains (up to 2 mm) are found dispersed in the sandstone (Fig. 4b). Grains are predominantly well-rounded to sub-rounded with a matte surface (more than 75%) (Fig. 4c). They are sometimes broken (less than 10%), and angular or subangular grains occur (less than 25%).
4.b Palynological analyses
The palynomorph assemblages contain much phytodebris (e.g. well-preserved tracheids), abundant terrestrial plant spores and relatively few marine elements. The latter include acritarchs and prasinophycean phycomata, but no chitinozoans or scolecodonts. In all samples, palynomorphs are poorly preserved, with colour ranging from brown to black. Many specimens are not suitable for detailed description. Nevertheless, the information provided by identifiable spores and marine palynomorphs is essential to document the age and the stratigraphic relationships of the local geological units.
4.b.1. Spores
The spore assemblage is of low diversity and exclusively represented by trilete spores (Fig. 5). Spore size varies from 18 to 45 μm. The assemblage is dominated by Dibolisporites wetteldorfensis Lanninger, Reference Lanninger1968 and Apiculiretusispora plicata (Allen) Streel, Reference Streel1967. The second component of the assemblage comprises Amicosporites jonkeri (Riegel) Steemans, Reference Steemans1989, which is relatively abundant, cf. Amicosporites discus Wellman & Richardson, Reference Wellman and Richardson1996 and Ambitisporites avitus Hoffmeister, Reference Hoffmeister1959. Other species such as Emphanisporites rotatus McGregor, Reference McGregor1961; E. multicostatus Rodriguez, Reference Rodriguez1978; Synorisporites sp. and Archaeozonotriletes chulus (Cramer) Richardson & Lister, Reference Richardson and Lister1969 are represented in the assemblage, together with rare specimens of Acinosporites sp. and Raistrickia sp. A few specimens are referred with doubt to the genus Camarozonotriletes Naumova ex Ishchenko, Reference Ishchenko1952. One specimen resembles C. filatoffii described by Breuer et al. (Reference Breuer, Al-Ghazi, Al-Ruwaili, Higgs, Steemans and Wellman2007), but is too poorly preserved to confirm the determination. Rare specimens of Apiculiretusispora brandtii Streel, Reference Streel1964 and Dictyotriletes cf. kerpii Wellman, Reference Wellman2006 are also present.

Figure 5. Spores. 1 – Dibolisporites wetteldorfensis Lanninger, Reference Lanninger1968 (slide no. 64343); 2 – Apiculiretusispora plicata (Allen) Streel, Reference Streel1967 (slide no. 64349); 3 – A. brandtii Streel, Reference Streel1964 (slide no. 64064); 4 – Ambitisporites avitus Hoffmeister, Reference Hoffmeister1959 (slide no. 64343); 5 – Emphanisporites rotatus Mc Gregor, Reference McGregor1961 (slide no. 64349); 6 – E. multicostatus Rodriguez, Reference Rodriguez1978 (slide no. 64064); 7 – Archaeozonotriletes chulus (Cramer) Richardson & Lister, Reference Richardson and Lister1969 (slide no. 64065); 8 – Acinosporites sp. (slide no. 64065); 9 – Amicosporites jonkeri (Riegel) Steemans, Reference Steemans1989 (slide no. 64064); 10 – Dictyotriletes cf. kerpii Wellman, Reference Wellman2006 (slide no. 64065); 11 – Raistrickia sp. (slide no. 64064); 12 – Synorisporites sp. (slide no. 64064); 13 – cf. Amicosporites discus Wellman & Richardson, Reference Wellman and Richardson1996 (slide no. 64064). Scale bar = 12 μm. A colour version of this figure is available at http://journals.cambridge.org/geo.
4.b.2. Marine palynomorphs
Marine palynomorphs are dispersed in the material. They are mainly represented by acritarchs and prasinophytes (Fig. 6). Taking account of their known distribution, the assemblage of acritarchs and prasinophycean phycomata appears to be mixed, containing Devonian species and reworked ones. The Devonian assemblage of acritarchs includes the following species: Winwaloeusia distracta (Deunff, Reference Deunff1966) Deunff, Reference Deunff1977; Multiplicisphaeridium raspa (Cramer) Eisenack, Cramer & Diez, Reference Eisenack, Cramer and Diez1973; Multiplicisphaeridium cf. paraguaferum (Cramer, Reference Cramer1964) Lister, Reference Lister1970; Micrhystridium sp.; ?Induoglobus sp.; Ammonidium sp.; Veryhachium cf. rosendae Cramer, Reference Cramer1964; Evittia sp. prasinophycean phycomata consist of simple leiospheres and Cymatiosphaera spp. Acritarch species such as Eupoikilofusa platynetrella Loeblich & Tappan, Reference Loeblich and Tappan1978; Eupoikilofusa stiatifera (Cramer, Reference Cramer1964) Cramer, Reference Cramer1970; Veryhachium subglobosum Jardiné et al. Reference Jardiné, Combaz, Magloire, Peniguel and Vachey1974 and Villosacapsula setosapellicula (Loeblich, Reference Loeblich1970) Loeblich & Tappan, Reference Loeblich and Tappan1976 are reworked from Upper Ordovician sediments, although some species are known to range into the Silurian.

Figure 6. Marine palynomorphs. Reworked acritarchs: 1 – Eupoikilofusa platynetrella Loeblich & Tappan, Reference Loeblich and Tappan1978 (slide no. 64064); 2 – Eupoikilofusa striatifera (Cramer, Reference Cramer1964) Cramer, Reference Cramer1970 (slide no. 64343); 3 – Villosacapsula setosapellicula (Loeblich, Reference Loeblich1970) Loeblich & Tappan, Reference Loeblich and Tappan1976 (slide no. 64064); 4 – Veryhachium subglobosum Jardiné et al. Reference Jardiné, Combaz, Magloire, Peniguel and Vachey1974 (slide no. 64064). prasinophycean phycomata: 5, 6 – Leiospheres (slide nos 64348 and 64064); 7 – Cymatiosphaera sp. (slide no. 64065). In situ (Devonian) acritarchs: 8 – Winwaloeusia distracta (Deunff, Reference Deunff1966) Deunff, Reference Deunff1977 (slide no. 64064); 9 – Multiplicisphaeridium raspa (Cramer) Eisenack et al. Reference Eisenack, Cramer and Diez1973 (slide no. 64065); 10 – Multiplicisphaeridium cf. paraguaferum (Cramer, Reference Cramer1964) Lister, Reference Lister1970 (slide no. 64348); 11 – Micrhystridium sp. (slide no. 64349); 12 – ?Induoglobus sp. (slide no. 64348); 13 – Ammonidium sp. (slide no. 64343); 14 – Veryhachium cf. rosendae Cramer, Reference Cramer1964 (slide no. 64065); 15 – Evittia sp. (slide no. 64065). For specimens 6, 7, 8, 9, 10, 11, 12, 13, 14, the scale bar = 10 μm: for specimen 3, the scale bar = 15 μm: for specimens 1, 2, 4, 5, 6, 15, the scale bar = 20 μm. A colour version of this figure is available at http://journals.cambridge.org/geo.
4.c. Plant megafossils
The plant megafossils occurring in these deposits are represented by compression/impression and permineralized specimens (Fig. 7). The compressions have been found at locality 11 (see log 11, Fig. 3). Axes are up to 15 cm long. They divide anisotomously and produce helically arranged lateral branches that divide again up to three times. They bear longitudinal ribbing and punctiform scars. Only vegetative lateral branches have been found in attachment. Some of those plants are close in vegetative morphology to the trimerophyte genera Pertica, Trimerophyton and Psilophyton. Their gross morphology also resembles the plant described by Gensel (Reference Gensel1984) from the Battery Point Formation (Gaspé Bay, Canada).

Figure 7. Plant megafossils occurring in the Basal Member of the Chalonnes Formation. (a) Compressions found at locality 11 (see log 11, Fig. 3); scale bar = 5 mm (sample no. CSD-07M-01). (b) Short length axes, permineralized by pyrite, occurring isolated in the sediment at locality 13 (see log 13, Fig. 3); scale bar = 0.5 mm (sample no. CSD-06C-501A). (c) Pair of fusiform sporangia twisted around each other, characteristic of Psilophyton; scale bar = 0.5 mm (sample no. CSD-06M-02B). (d) Transverse section of an axis of the first plant type; scale bar = 2 mm (sample no. CSD-06C-C08). (e) Transverse section of an axis of the second plant type; scale bar = 0.4 mm (sample no. CSD-06C-004). A colour version of this figure is available at http://journals.cambridge.org/geo.
Short length axes are permineralized by pyrite and occur isolated in the sediment, at locality 13 (see log 13, Fig. 3). Their anatomical structure is relatively well preserved. Two main types of structures have been observed. Fertile organs of two types also occur.
First plant type. Numerous axes that are 1 to 3 mm in diameter have been found. Pairs of fusiform sporangia twisted around each other, characteristic of Psilophyton (Gerrienne, Reference Gerrienne1997), also occur dispersed in the sediment. In transverse section, axes show a massive circular to elliptical xylem strand with an elongate centrarch to mesarch protoxylem area. The central part of the strand with randomly arranged tracheids is surrounded by a zone of tracheids placed in radial rows. Xylem comprises P-type tracheids with well-preserved scalariform pitting. Those anatomical features are characteristic of the genus Psilophyton. To date, it is unknown if these pyritized axes and the axes preserved in compression (see above) belong to the same plant. On the basis of their anatomy and the presence of dispersed paired fusiform sporangia, our permineralized specimens are attributed to Psilophyton sp. Psilophyton is known from a number of Lower Devonian localities distributed worldwide. The stratigraphic range of the genus is Pragian–lowermost Middle Devonian (Gerrienne, Reference Gerrienne1997). The genus is of great importance in the evolution of vascular plants, as it is considered the earliest Euphyllophyte, a clade that comprises both the Monilophytes (ferns sensu lato) and the Lignophytes (seed plants and their free-sporing ancestors).
Second plant type. Rather badly preserved tiny axes display a dissected stele of primary xylem comparable in transverse section either to a Gothanophyton-type or to a small compressed cladoxylopsid axis. Reticulate elements with slightly bordered pits similar to those of Foozia decribed by Gerrienne (Reference Gerrienne1992) from the Early Devonian of Belgium have been observed in the palynological slides. Additionally, pairs of sporangia similar in size and morphology to those of Foozia have been found dispersed in the sediment. Dibolisporites echinaceus has been identified in the sporangia of Foozia while Dibolisporites wetteldorfensis occurs, dispersed in the sediment from the Basal Member of the Chalonnes Formation. This suggests that the plant from Châteaupanne, as Foozia, possibly represents an early cladoxylopsid.
5. Discussion
5.a. Age of the deposits
The in situ (Devonian) acritarch assemblage is of low diversity, represented by simple long-ranging forms, and is devoid of stratigraphically significant taxa. There are relatively few publications on Early Devonian acritarchs (Le Hérissé, Servais & Wicander, Reference Le Hérissé, Servais and Wicander2000). However, low-diversity phytoplankton assemblages associated with nearshore environment conditions are common in the Pragian–Emsian interval in other localities of northern Gondwana (e.g. North Africa, Spain) and peri-Gondwanan domain (e.g. Armorican Massif, Normandy) (Cramer, Reference Cramer1964; Rauscher, Doubinger & Manche-Bain, Reference Rauscher, Doubinger and Manche-Bain1965; Magloire, Reference Magloire1967; Jardiné, Reference Jardiné1972; Moreau-Benoit, Reference Moreau-Benoît1974; Deunff, Reference Deunff and Lardeux1976; Le Hérissé, Reference Le Hérissé1983).
Reliable age determinations for the samples studied are provided by spore assemblages. Dibolisporites wetteldorfensis represents a major component. Based on the presence of this species, the general characteristics of the assemblage (general form and size of the spores) and the absence of spores characteristic of younger intervals, it is considered that the assemblage belongs to the PoW Oppel Zone of Streel et al. (Reference Streel, Higgs, Loboziak, Riegel and Steemans1987). This zone ranges from early Pragian to earliest Emsian. Few assemblages belonging to the PoW Oppel Zone are described in the literature (e.g. Steemans, Reference Steemans1989; Wellman et al. Reference Wellman, Thomas, Edwards and Kenrick1998; Turnau, Milaczewski & Wood, Reference Turnau, Milaczewski and Wood2005; Wellman, Reference Wellman2006; Hassan Kermandji, Kowalski & Touhami, Reference Hassan Kermandji, Kowalski and Touhami2008). A comparison of most of the assemblages reported belonging to this zone has been made by Wellman (Reference Wellman2006). The majority of them are from Euramerica and northern Gondwana. Correlations with the Armorican Massif are provided by the work of Le Hérissé (Reference Le Hérissé1983), who described spore assemblages from two marine sections, Sablé-sur-Sarthe (Upper part of the Saint-Cénéré Formation) and Saint-Pierre-sur-Erve (Montguyon Formation) in the Central Armorican domain. Three spore assemblages have been recognized and dated from Pragian to Lower Emsian. It appears that the spore assemblage described herein from the Basal Member of the Chalonnes Formation has much in common with the assemblages from Saint Pierre-sur-Erve. Apiculiretusispora plicata, A. brandtii, Retusotriletes sp., Dibolisporites wetteldorfensis Raistrickia sp., Emphanisporites rotatus and Archaeozonotriletes chulus are common in the two assemblages. The Montguyon Formation has also been studied for palaeontological content and is considered to be Upper Pragian/Lower Emsian (Botquelen, Reference Botquelen2003).
5.b. Depositional environment
The basal sandy beds are mainly made up of rounded and smoothed quartz grains, shaped by the wind (Cailleux, Reference Cailleux1962). These quartz grains could come from sub-contemporaneous deposits such as beach sands, or from older eroded sedimentary series. Our study partly supports this latter hypothesis. Indeed, the ‘Pélites à fragments’ Formation, of Hirnantian age (Upper Ordovician) (Robardet & Doré, Reference Robardet and Doré1988) and belonging to the Armorican Massif, contains rounded-quartz sandstones. This glacio-marine formation is unknown in the Châteaupanne Unit, but is present in the northern Tombeau Leclerc Unit (A. Bourarhouh, unpub. Ph.D. thesis, Univ. Rennes, 2002; Piçarra et al. Reference Piçarra, Robardet, Bourarhouh, Paris, Pereira, Le Menn, Gourvennec, Oliveira and Lardeux2002, Reference Piçarra, Robardet, Oliveira, Paris and Lardeux2009) (Fig. 2). It is possible that this series has been deposited on the top of the Fresne Formation and then eroded. This erosion occurred before the sedimentation of the Basal Member of the Chalonnes Formation and consequenty enriched this level in aeolian quartz grains. The reworked acritarchs found in our assemblage include some classic Upper Ordovician species, such as E. platynetrella, V. subglosum or V. setosapellicula, that are recorded in pre-glacial series of the upper Ordovician or are restricted to the Hirnantian deglaciation time (Vecoli & Le Hérissé, Reference Vecoli and le Hérissé2004). The discovery of these acritarchs in the studied sandstones supports the reworking of the rounded quartz grains from the ‘Pélites à fragments’ Formation.
On the other hand, two points may be added: (1) E. stiatifera has been previously found in the Katian (Upper Ordovician) ‘Dicellograptus Siltstone’ (upper part of the Fresne Siltstone), located just below the Chalonnes Limestone (Moreau-Benoit, Reference Moreau-Benoît1974). (2) A large amount of dark shale fragments is present in these sandstones and could come either from sub-contemporaneous mud layers or from the underlying Fresne Formation (Fig. 2). The basal surface of the Basal Member of the Chalonnes Formation represents an unconformity within a stratigraphic gap including uppermost Ordovician, Silurian and Lochkovian (early Lower Devonian) deposits. As a result, this unconformity could be related to a major aerial erosional surface overlain by terrigeneous inputs. These inputs could come both from reworked underlying sediments (from the Fresne and the ‘Pelite à fragments’ Formations) and from contemporaneous aeolian sand and clay chips.
The sediments forming the Basal Member of the Chalonnes Formation are characterized by in situ (Devonian) acritarchs and an important input of continental palynomorphs. This palynofacies suggests a nearshore marine environment. The low diversity of the in situ (Devonian) acritarch assemblage recovered from these deposits can be related to the local nearshore situation of the deposits, which is in general an unfavourable environmental condition to produce abundant and complex phytoplankton assemblages (see e.g. Strother, Reference Strother, Al-Hajri and Traverse1996). The absence of chitinozoans and scolecodonts could be due to possible mechanical phenomena during sedimentation of organic material, separating forms with good buoyancy, like spores and phytodebris, from the heaviest forms. Despite the important continental supply, the diversity in spore species is very low for such sedimentary facies. This could reflect that sediments contain only local dispersed spores without remote input from rivers. However, these conclusions need to be tempered because of the poor palynomorph preservation which could modify the assemblage content.
Hummocky cross-stratification (HCS) and swaley cross-stratification (SCS) occur in the sandy beds of the Basal Member of the Chalonnes Formation; these structures are related to storm wave action (Leckie & Walker, Reference Leckie and Walker1982; Aigner, Reference Aigner1985; Einsele, Reference Einsele1992). According to the depositional models proposed for storm wave-dominated shelves (Dott & Bourgeois, Reference Dott and Bourgeois1982; Aigner, Reference Aigner1985; Guillocheau & Hoffert, Reference Guillocheau and Hoffert1988), they may be located from the shoreface (SCS–HCS) to the proximal and median parts of the inner shelf (HCS). The scarce bioturbation would imply that storm events were close in time. For the coarse-grained levels, they could be considered as lag deposits related to by-pass episodes or, for the lowermost one, as basal lag. The plant-rich shaly layers could have been deposited during quiet phases, perhaps in a relatively protected area. This suggestion supports the palynological conclusion expressed above.
The palynological and sedimentological data are in agreement with a nearshore depositional environment for the Basal Member of the Chalonnes Formation. This level is interpreted as a transgressive sandbody whose material has been continuously reworked by storm waves during landward migration of the shoreface.
6. The Devonian flora from the Armorican Massif
A survey of the plant-bearing deposits reported from the Devonian formations of the Armorican Massif is presented in Table 1; their stratigraphic position and location are given in Figure 8. Almost all of these remains need to be reinvestigated; some might not even be plants. In the Ancenis Basin, a flora from the Culm (Frasno-Dinantian period sensu Bureau) has been described (Bureau, Reference Bureau1911, Reference Bureau1913; Carpentier, Reference Carpentier1920; C. Beaupère, unpub. Thèse Doctorat, Univ. Paris, 1973; Ballèvre & Lardeux, Reference Ballèvre and Lardeux2005). The strictly Upper Devonian (Famennian) plant-bearing deposits are not well known in this area. To date, most of the plants reported in these articles are known as Carboniferous plants. A reinvestigation of the sedimentology and palaeontology of these deposits is necessary.
Table 1. Survey of the plant-bearing deposits reported from the Devonian formations of the Armorican Massif


Figure 8. Stratigraphic position and location of the Devonian plant-bearing deposits from the Armorican Massif. Only megafossil plant remains are taken into account. A colour version of this figure is available at http://journals.cambridge.org/geo.
The Emsian Sainte-Anne Formation is to date the only unequivocal Devonian flora described from the Armorican Massif (for the description of this flora, see Ducassou et al. Reference Ducassou, Strullu-Derrien, Ballèvre, Dabard, Gerrienne, Lardeux and Robin2009). On the other hand, palynological studies have shown spores and phytodebris in the Lower Devonian (Lochkovian to Emsian) and the oldest (Silurian) deposits from the Armorican Massif (e.g. Moreau Benoît, Reference Moreau-Benoît1974; Le Hérissé, Reference Le Hérissé1983; V. Baudu, unpub. Ph.D. thesis, Univ. Rennes, 1994; Baudu & Paris, Reference Baudu and Paris1995), but these scarce plant elements have not been reported associated with macroflora. The palynomorph assemblages studied here provide a reliable Pragian–earliest Emsian age for the Basal Member of the Chalonnes Formation. The flora from these deposits is thus older than that described from the Sainte-Anne Formation (Ducassou et al. Reference Ducassou, Strullu-Derrien, Ballèvre, Dabard, Gerrienne, Lardeux and Robin2009) and represents the earliest Devonian macroflora recorded in the Armorican Massif.
7. Conclusions
The Armorican Massif is known to provide an excellent record of the Palaeozoic history of the Variscan belt. The study presented here adds new information to this history. The Basal Member of the Chalonnes Formation consists of transgressive nearshore sediments deposited over the previously emergent land surface. The emergence preceding the transgression has never been observed before, and has still to be investigated in terms of the geodynamic evolution of the Variscan belt. The deposits described here are attributed to the Pragian–earliest Emsian interval, and record the earliest indication of terrestrial colonization in the Armorican Massif. The report of plant megafossils also shows the promising potential of the Basal Member of the Chalonnes Formation for further palaeobotanical studies.
Acknowledgements
The authors thank Mr E. Souchet and Mr J. Binet for sampling in the Châteaupanne quarry and Mr Boulétreau for sampling in Tarare Quarry. C. S. D. extends her gratitude to Dr C. M. Berry (University of Cardiff, Wales), Dr F. Paris (Université de Rennes) and Dr F. Redois (Université d'Angers). Mrs M. Giraldo and Mr J. Laval (Université de Liège) are acknowledged for the preparation of the palynological and palaeobotanical slides. The authors also thank the two reviewers for their constructive suggestions. This work was realized as part of the Ph.D. theses of the first two authors. It was partly funded by a MRT grant (attributed to C. D.).