



Los índices de utilidad económica han sido ampliamente utilizados en trabajos zooarquelógicos desde la publicación de Nunamiut Etnoarchaeology (Binford Reference Binford1978). El presupuesto que subyace a la propuesta de Binford es que el valor económico —entendido en términos de cantidad de carne, grasa y médula, de las distintas regiones anatómicas de un animal— puede usarse como marco de referencia para indagar sobre las decisiones que los grupos cazadores-recolectores toman respecto de las estrategias de aprovechamiento de una presa (transporte, procesamiento, consumo y descarte) e inferir funcionalidades específicas para los espacios en los que las carcasas se encuentran depositadas. En la actualidad, tales índices son de uso común en los trabajos que analizan patrones de movilidad, estrategias de subsistencia, pautas de procesamiento y/o consumo de distintos tipos de presas, destacándose especialmente los cálculos realizados para mamíferos terrestres (e.g., Belardi y Gómez Otero Reference Belardi and Otero1998; Blumenschine y Caro Reference Blumenschine and Caro1986; Borrero Reference Borrero, Davies and Reeves1990; Outram y Rowley-Conwy Reference Outram and Rowley-Conwy1998). Asimismo, en los últimos años se han desarrollado varios índices exclusivos para distintas especies de cetáceos, fócidos y otáridos (Lyman et al. Reference Lyman, Savelle and Whitridge1992; Martinoli Reference Martinoli2018; San Román Reference San Román, López, Cartajena, García and Mena2009, Reference San Román2016; Savelle y Friesen Reference Savelle and Friesen1996; Savelle et al. Reference Savelle, Friesen and Lyman1996). En términos generales, los mamíferos marinos poseen una anatomía distinta a los terrestres; tal es el caso, por ejemplo, de la proyección de los miembros anteriores y posteriores (Lyman Reference Lyman1992).
En el caso de Sudamérica, la única especie de otárido para la cual se ha calculado un índice de utilidad económica es Otaria flavescens (San Román Reference San Román, López, Cartajena, García and Mena2009). Sin embargo, en registros zooarqueológicos provenientes de distintos sectores de ese amplio subcontinente se ha observado la presencia de otra especie de otárido: Arctocephalus australis. Esta especie ha sido explotada en distintos contextos culturales y ambientes costeros y a lo largo de diferentes momentos de la secuencia de ocupación de dicho subcontinente (Borella Reference Borella2006; Borella et al. Reference Borella, Frontini and Bayón2017; deFrance et al. Reference deFrance, Keefer, Richardson and Álvarez2001; Inda et al. Reference Inda, del Puerto, Bracco, Castiñeira, Capdepont, Gascue, Baeza and García-Rodríguez2011; López et al. Reference López, Piñeiro, Castiñeira and Gascue1997; Muñoz et al. Reference Muñoz, Götz, Roca, Muñoz, Götz and Roca2014; Orquera y Piana Reference Orquera and Piana1999; Rudolph Reference Rudolph2006; Salazar et al. Reference Salazar, Figueroa, Andrade, Salinas, Olguín, Power, Rebolledo, Parra, Orellana and Urrea2015; San Román Reference San Román2016; Schiavini Reference Schiavini1990; Volkmer de Castilho y Simões-Lopes Reference Volkmer de Castilho and Simões-Lopes2008).
El objetivo del presente trabajo es presentar el cálculo de un índice de utilidad económica específico para la especie A. australis. Esto permitirá mejorar las interpretaciones respecto de los factores que pueden haber generado los perfiles anatómicos observados en los conjuntos zooarqueológicos sudamericanos, específicamente el rol que han jugado los criterios económicos en las decisiones referidas al transporte, procesamiento y consumo de carcasas de dicha especie entre grupos cazadores-recolectores. Asimismo, se evalúa la aplicación de este índice (en función de posibles estrategias de selección y descarte) en dos conjuntos de la costa sur de Tierra del Fuego (región del canal Beagle): el Segundo Componente de Imiwaia I y la Capa C de Ajej I, con cronologías correspondientes al Holoceno medio y tardío, respectivamente.
Los Pinnípedos Sudamericanos
Los pinnípedos más habituales y abundantes en Sudamérica pertenecen a dos especies, ambas de la familia Otariidae: el lobo marino de dos pelos o lobo fino sudamericano (A. australis) y el lobo marino de un pelo o lobo marino común (O. flavescens). Además, ocasionalmente, se pueden observar ejemplares aislados o colonias pequeñas de especies antárticas y subantárticas como Arctocephalus gazella, Arctocephalus tropicalis, Mirounga leonina, Leptonychotes weddellii, Lobodon carcinophaga, Hydrurga leptonyx y Ommatophoca rossi (Bastida y Rodríguez Reference Bastida and Rodríguez2003).
La especie O. flavescens se distribuye a lo largo de las costas atlántica y pacífica de Sudamérica, incluyendo las Islas Malvinas. En la vertiente atlántica el límite norte se ubica en el Archipiélago de la Coronilla (Uruguay); en la pacífica, en la Isla de Lobos de Tierra (Perú; Bastida y Rodríguez Reference Bastida and Rodríguez2003; Schiavini Reference Schiavini1990; Vaz Ferreira Reference Vaz Ferreira, Ridgway and Harrison1981). Los lobos marinos de un pelo eligen playas con declives suaves y más reparadas, tal es así que su distribución geográfica es continua a lo largo de toda la costa, además de la ocupación de islas e islotes (Crespo, García, Dans, Pedraza, Lewis y Campagna Reference Crespo, García, Dans, Pedraza, Lewis and Campagna2008:9; Schiavini Reference Schiavini1990; Schiavini y Raya Rey Reference Schiavini and Rey2001; Sielfeld Reference Sielfeld1983, Reference Sielfeld1999). Los tamaños del lobo marino de un pelo son los siguientes: las crías pesan entre 10,2 y 14,8 kg; los machos adultos alcanzan los 2,56 m de largo y 300–340 kg de peso; las hembras adultas alcanzan los 2 m de largo y 144 kg de peso (datos procedentes de las costas uruguayas y de las Islas Malvinas; Schiavini Reference Schiavini1990; Vaz Ferreira Reference Vaz Ferreira1979).
La distribución de A. australis se extiende desde la Isla del Marco en Uruguay hasta la Isla Mayorca en Perú, incluyendo las islas de la costa sudamericana y las Islas Malvinas (Bastida y Rodríguez Reference Bastida and Rodríguez2003). Se ha informado también su presencia en sectores del sur de Brasil (Rio Grande do Sul; Figura 1; Crespo et al. Reference Crespo, Schiavini, García, Trecu, Goodall, Rodríguez, Morgante and Oliveira2015; Riedman Reference Riedman1990). Los lobos marinos de dos pelos se ubican preferentemente sobre playas rocosas y con cierta exposición al oleaje (Crespo, García, Dans y Pedraza Reference Crespo, García, Dans and Pedraza2008; Schiavini Reference Schiavini1990; Sielfeld Reference Sielfeld1983), motivo por el cual la distribución de las loberías reproductivas no es uniforme en el espacio costero y éstas se concentran principalmente en islotes escarpados (Bastida y Rodríguez Reference Bastida and Rodríguez2003; Bonner Reference Bonner1982; Schiavini Reference Schiavini1990). La mayor agrupación se encuentra en seis islas uruguayas, pero se destaca también la presencia de A. australis en el sur de Chile, en las Islas Malvinas, en Isla de los Estados y en el norte de Chile y sur de Perú (Bastida y Rodríguez Reference Bastida and Rodríguez2003; Crespo et al. Reference Crespo, Schiavini, García, Trecu, Goodall, Rodríguez, Morgante and Oliveira2015).

Figura 1. Mapa de América del Sur con la distribución de A. australis y detalle de la costa sur de Tierra del Fuego con indicación del lugar de hallazgo del individuo utilizado para el cálculo del índice de anatomía económica y de los sitios arqueológicos analizados.
Se han hallado diferencias en el nivel de unidades evolutivas entre las poblaciones de A. australis en la vertiente atlántica y pacífica (Oliveira et al. Reference Oliveira, Hingst-Zaher and Morgante2005; Túnez et al. Reference Túnez, Centron, Cappozzo and Cassini2007, Reference Túnez, Cappozzo, Paves, Albareda and Cassini2013), específicamente entre las de Perú y norte de Chile, el Atlántico sur-occidental (Brasil, Uruguay, Argentina e Islas Malvinas) y el este del Pacífico sur en Chile (Oliveira y Brownell Reference Oliveira and Brownell2014). Uno de los aspectos que nos interesa resaltar a este respecto es la variabilidad en los tamaños craneales que reportan Oliveira y coautores (Reference Oliveira, Hoffman, Hingst-Zaher, Majluf, Muelbert, Morgante and Amos2008): las muestras provenientes de Perú indican que los individuos de este espacio presentan dimensiones mayores que aquellos de Uruguay y Brasil, a la vez que previamente fueron informadas diferencias entre las poblaciones del centro-norte y del sur del subcontinente (Schiavini Reference Schiavini1990). Las dimensiones promedio informadas por Vaz Ferreira (Reference Vaz Ferreira1979) a partir de pinnípedos de Islas Malvinas, son las siguientes: las crías pesan entre 3,5 y 5,5 kg; los machos adultos alcanzan 1,9 m de largo y 159 kg de peso; las hembras adultas alzanzan 1,4 m de largo y 48,5 kg de peso. Los cálculos presentados por Schiavini (Reference Schiavini1990) postulan dimensiones promedio menores en las poblaciones uruguayas: pesos entre 68,2 y 80,9 kg para los machos adultos y 40,6 kg para las hembras adultas. En el caso de las poblaciones peruanas (Punta San Juan) se ha informado un peso promedio para las hembras adultas de alrededor de 58 kg (Majluf Reference Majluf1992:377, tabla 3). A pesar de las diferencias en las dimensiones intra-especie informadas en distintos espacios costeros sudamericanos, es importante destacar la gran diferencia de tamaño que presentan respecto de O. flavescens: estos últimos duplican en peso a los individuos de A. australis.
¿Por qué un Índice de Utilidad Económica para Arctocephalus australis?
En términos generales, la presencia de restos de A. australis se ha relevado en contextos relativamente heterogéneos a lo largo de todo el litoral sudamericano. En la vertiente pacífica, en el actual territorio de Chile ha sido documentado el consumo de esta especie en distintos yacimientos que incluyen desde la costa del desierto de Atacama (Olguín et al. Reference Olguín, Castro, Castro, Peña-Villalobos, Ruz and Santander2015; Salazar et al. Reference Salazar, Figueroa, Andrade, Salinas, Olguín, Power, Rebolledo, Parra, Orellana and Urrea2015) hasta el archipiélago fueguino y el Cabo de Hornos (Legoupil Reference Legoupil1989, Reference Legoupil1994, Reference Legoupil1997; San Román Reference San Román2010, Reference San Román2016), abarcando todo el Holoceno. Por su parte, en la costa sur de Perú se han identificado conjuntos en los cuales los otáridos (O. flavescens y A. australis) han sido presas de variable importancia en la dieta de los grupos que allí habitaron, tanto en momentos tempranos de la secuencia de ocupación (deFrance et al. Reference deFrance, Keefer, Richardson and Álvarez2001; Lavallèe et al. Reference Lavallée, Julien, Béarez, Bolaños, Carré, Chevalier, Delabarde, Fontugne, Rodríguez-Loredo, Klaric, Usselmann and Vanhaeren2011; Reitz Reference Reitz1988) como en el período correspondiente a sociedades agropastoriles (Roselló et al. Reference Roselló, Vásquez, Morales and Rosales2001). En lo que respecta a la vertiente atlántica, en el sur de Brasil (Rio Grande do Sul) se han relevado restos de A. australis en sitios especialmente del centro-sur de Santa Catarina con cronologías que abarcan desde 5000 a 900 aP; es decir, en contextos tanto cerámicos como pre-cerámicos (Volkmer de Castilho y Simões-Lopes Reference Volkmer de Castilho and Simões-Lopes2008). En el litoral atlántico uruguayo se ha informado la presencia de restos óseos de Arctocephalus sp. en conjuntos con cronologías posteriores a los 4400 años no calibrados aP (Castiñeira et al. Reference Castiñeira, Panario, Bracco, Gutiérrez, Oliva, de Grandis and Rodríguez2010; Inda et al. Reference Inda, del Puerto, Bracco, Castiñeira, Capdepont, Gascue, Baeza and García-Rodríguez2011; López et al. Reference López, Piñeiro, Castiñeira and Gascue1997; López y Villamarzo Reference López, Villamarzo, López and Gascue2009; Rudolph Reference Rudolph2006). En Argentina se han identificado restos óseos correspondientes a la especie A. australis a lo largo de toda la costa atlántica (Belardi et al. Reference Belardi, Marina and L'Heureux2011; Borella Reference Borella2006; Frontini y Bayón Reference Frontini, Bayón, Mondini, Muñoz and Fernández2017; Moreno Reference Moreno2008; Muñoz Reference Muñoz and Goñalons2004, Reference Muñoz2005; Muñoz et al. Reference Muñoz, Cruz, Lemaire, Pretto, Zangrando, Barberena, Gil, Neme, Giardina, Luna, Otaola, Paulides, Salgán and Tivoli2013). En el caso específico de los sitios arqueológicos correspondientes a adaptaciones marítimas del canal Beagle, Tierra del Fuego (Figura 1), la mayoría de los conjuntos en los que se avanzó sobre la determinación en el nivel de especie muestran el predominio en el consumo de A. australis (Martinoli Reference Martinoli2015, Reference Martinoli2017, Reference Martinoli2018; Orquera y Piana Reference Orquera and Piana1999; Piana et al. Reference Piana, Vázquez and Álvarez2008; Schiavini Reference Schiavini1990, Reference Schiavini1993; Zangrando Reference Zangrando2009).
A partir de la ubicuidad de A. australis en los registros arqueológicos sudamericanos, es posible sostener que fue un recurso de cierta relevancia. Sin embargo, la importancia económica en la dieta y las modalidades de explotación seleccionadas por los cazadores-recolectores-pescadores muestra amplia variabilidad (Martinoli Reference Martinoli2018; Muñoz et al. Reference Muñoz, Götz, Roca, Muñoz, Götz and Roca2014). Esto podría relacionarse (entre otros factores) con el hecho de que la distribución de esa especie no es homogénea a lo largo del litoral, ocupando en muchos casos islas e islotes a diferentes distancias de costas continentales o de grandes masas de tierra (Bastida y Rodríguez Reference Bastida and Rodríguez2003), lo que implicaría la disponibilidad de tecnología de navegación y captura específica para su explotación (Orquera y Piana Reference Orquera and Piana1999).
En términos teórico-metodológicos, los modelos de optimización han explorado desde múltiples perspectivas la relación entre la abundancia y la distribución de los recursos (marinos y terrestres) y las decisiones que se toman respecto de su explotación en función de la aptitud (fitness) de comportamientos alternativos (Cannon y Broughton Reference Cannon, Broughton, Broughton and Cannon2010; Kelly Reference Kelly1995; Smith y Winterhalder Reference Smith, Winterhalder, Smith and Winterhalder1992). En dicho marco, los índices de anatomía económica han sido una herramienta de gran utilidad en tanto han permitido poner a prueba, de manera sencilla, hipótesis relacionadas con el aprovechamiento diferencial de presas en contextos socioecológicos particulares (Gifford-González y Sunseri Reference Gifford-Gonzalez and Sunseri2009; Lyman Reference Lyman1992). Tal sería el caso de la evaluación de decisiones económicas relacionadas con el transporte diferencial o estrategias de procesamiento de determinadas porciones de la carcasa en función del tamaño y disponibilidad (densidad) de las presas, los lugares de captura y de los medios de movilidad disponibles (Ames Reference Ames, Duer and Turner2005; Burger et al. Reference Burger, Hamilton and Walker2005; Cannon Reference Cannon2003; Gifford-González y Sunseri Reference Gifford-Gonzalez and Sunseri2009; Lupo Reference Lupo2006; Martinoli Reference Martinoli2018; Metcalfe y Barlow Reference Metcalfe and Barlow1992), especialmente en grupos cazadores-recolectores-pescadores volcados al consumo de recursos marinos, ya sea de manera especializada u oportunista (Lyman Reference Lyman1989; Martinoli Reference Martinoli2018). Específicamente, análisis zooarqueológicos realizados en la vertiente pacífica de Norteamérica hallaron diferencias en el transporte en función del tamaño (peso) de una especie de pinnípedo (Callorhinus ursinus), con una clara tendencia a la selección de porciones de elevado valor económico en el caso de las presas de mayores dimensiones, los machos adultos (Gifford-González y Sunseri Reference Gifford-Gonzalez and Sunseri2009), los cuales tienen un rango de tamaño comparable al de A. australis (hembra adulta 40 kg; macho adulto 180 kg).
En lo que respecta a la aplicación de índices de utilidad económica específicos de una especie a otra del mismo orden, trabajos previos han indicado, a partir de análisis estadísticos, que en el caso de ser utilizados como una escala ordinal es justificado su uso inter-especie (Friesen et al. Reference Friesen, Savelle, Diab, Gerlach and Murray2001). Sin embargo, esos mismos autores observan suficientes diferencias entre distintas especies de pinnípedos como para considerar relevante la derivación de índices específicos para cada taxón. En el caso de la comparación entre otáridos de Norteamérica y Sudamérica, si bien se ha hallado una correlación muy elevada y estadísticamente significativa entre O. flavescens y Z. californianus (r s = 0,87; t = 6,06; p = 0,001; San Román Reference San Román, López, Cartajena, García and Mena2009), lo que permitió al autor sostener que las características anatómicas de ambas, particularmente las referidas a la musculatura, son similares, también fueron halladas algunas diferencias al interior de dicha familia. Entre las principales se destacan, por un lado, el peso de la piel y el tejido adiposo, mucho mayor en la especie proveniente del hemisferio norte, Z. californianus. Por otro lado, los pesos de las vísceras, escápula, tibia-peroné y aleta posterior (ver discusión al respecto del procesamiento de las aletas en San Román [Reference San Román, López, Cartajena, García and Mena2009:174]) muestra mayor valor para la especie natural del hemisferio sur, O. flavescens. En el caso de la escápula, incluso las distinciones observadas en el peso de la porción generan una inversión en la clasificación de utilidad.
Ahora bien, en las dos especies disponibles en el litoral sudamericano (A. australis y O. flavescens) se ha corroborado, sobre la base de un estudio estadístico comparativo de diferencias morfométricas, que existen notables distinciones en el desarrollo físico y morfológico, principalmente en las extremidades anteriores del cuerpo (Cadegán Sepúlveda Reference Cadegán Sepúlveda2013). Los huesos pertenecientes a la cintura escapular del lobo marino de un pelo presentaron características que evidencian un mayor rendimiento muscular, basado en el mayor tamaño proporcional de las estructuras óseas, destacando las zonas de inserción muscular y articulares. Esta diferencia en la morfometría ósea podría estar asociada a un aumento de la fuerza generada por los individuos de O. flavescens para el movimiento en tierra y la importante actividad acuática que incluye viajes a grandes profundidades, característicos de esta especie (Sielfeld Reference Sielfeld1999). Por lo tanto, a priori se esperarían diferencias en la distribución de los valores de utilidad económica, principalmente en los miembros.
Muestra
En este trabajo se presenta la información correspondiente a un individuo de la especie A. australis recolectado en las cercanías de la ciudad de Ushuaia, Tierra del Fuego, Argentina. El ejemplar fue hallado muerto en mayo de 2014 en aguas del canal Beagle (Paso Les Éclaireurs; Figura 1). El lobo marino de dos pelos fue procesado entero en el Centro Austral de Investigaciones Científicas (CADIC-CONICET). El individuo fue identificado como un macho subadulto, con una edad aproximada de cinco años, determinada por la Dra. Natalia Dellabianca a partir del conteo de los anillos de crecimiento de un canino superior. Dicho resultado mostró relativa coherencia con los estadios de fusión de los huesos largos (Borella et al. Reference Borella, Grandi, Vales, Goodall, Crespo, Zangrando, Barberena, Gil, Neme, Giardina, Luna, Otaola, Paulides, Salgán and Tivoli2013). El peso total del espécimen no pudo ser tomado por falta de una balanza adecuada; por lo tanto, el valor presentado (58 kg) fue reconstruido en función de los pesos parciales de las distintas porciones anatómicas, de las vísceras y del cuero (Tabla 1). Su longitud total fue de 162 cm (Figura 2). El individuo no tenía contenido estomacal y confirmamos la presencia de una infección pulmonar crónica como causa de la muerte (Carla Fiorito, comunicación personal 2014).
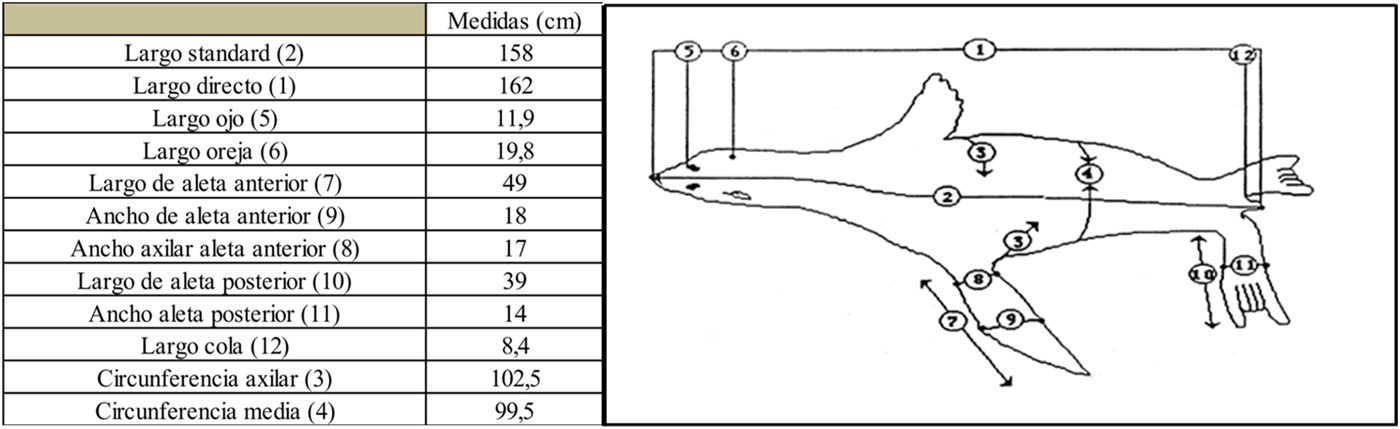
Figura 2. Medidas del espécimen de A. australis completo (modificado de Dierauf Reference Dierauf1994).
Tabla 1. Pesos de las Porciones Derechas e Izquierdas del Individuo de A. australis Analizado y Estado al Momento de ser Pesados.

Métodos
El proceso de cuereado, eviscerado y disección fue llevado a cabo con cuchillos de hoja metálica, bisturíes y dos lascas (riolita y obsidiana verde) con filo natural (experimentación realizada por la Lic. Vanesa Parmigianni y la Dra. Celina Álvarez Soncini). Se realizó el cuereado completo del animal, con excepción de las aletas. Cabe destacar que la grasa subcutánea no superó el centímetro de espesor.Footnote 1
Las vísceras fueron pesadas por separado (Tabla 1) pero, al no estar asociadas con ningún hueso en particular, no se utilizaron para el cálculo del índice de utilidad. Luego se procedió a la separación en porciones anatómicas más pequeñas según proponen Lyman y coautores (Reference Lyman, Savelle and Whitridge1992:533). En primer lugar, se hicieron cortes de desarticulación para separar la cabeza; en segundo término, se retiraron los miembros superiores e inferiores del tronco y, a su vez, los miembros fueron divididos en inferior y superior. Por último, fueron separadas las costillas, las esternebras y la columna vertebral, la cual fue seccionada en tres partes (vértebras cervicales, torácicas y lumbares-sacro-caudales).
El proceso completo de descarne y pesado del individuo tomó cinco días, por lo cual algunas unidades fueron congeladas (Tabla 1) y pesadas una vez descongeladas. Este hecho pudo afectar levemente el peso de las porciones por la pérdida de agua, aunque desconocemos en que magnitud. El cerebro no pudo ser pesado por separado dada la dificultad de su extracción; por lo tanto, el valor presentado surge del peso total de la cabeza (carne, órganos y hueso) menos el peso de los huesos del cráneo luego de hervidos. Por su parte, las aletas se separaron enteras (cuero, carne y hueso). El cuereado se realizó con bisturí por la gran cantidad de tendones que tiene esta porción; asimismo, se observó que en esta zona el panículo adiposo adquiere un espesor levemente mayor al centímetro.
El cálculo del índice de utilidad económica %MUI (meat utility index) y su versión modificada %MMUI (modified meat utility index) se efectuó siguiendo los lineamientos generales propuestos por Lyman y colaboradores (Reference Lyman, Savelle and Whitridge1992), Metcalfe y Jones (Reference Metcalfe and Jones1988) y Savelle y colaboradores (Reference Savelle, Friesen and Lyman1996). En la Tabla 1 se detallan los pesos obtenidos. El peso total corresponde a cada unidad anatómica completa (tejidos y hueso); el peso de la carne se obtuvo restando el peso del hueso hervido, por un lapso menor a dos horas, al peso total. Para llevar adelante el procedimiento se utilizó una balanza digital con precisión de 0,1 g.
Con respecto a la derivación del %MMUI, que permite dar cuenta de la presencia de elementos de bajo valor alimenticio que se introducen al estar en conexión con elementos de alto valor (riders [Binford Reference Binford1978:64]), empleamos los mismos criterios de agregación de porciones anatómicas propuestos para Phocidae (Lyman et al. Reference Lyman, Savelle and Whitridge1992), que son levemente distintos a los que utilizan Savelle y coautores (Reference Savelle, Friesen and Lyman1996) y San Román (Reference San Román, López, Cartajena, García and Mena2009) para Otariidae. No obstante, elegimos el primer criterio considerando que los autores utilizan información etnográfica para definir las unidades de trozamiento (Lyman et al. Reference Lyman, Savelle and Whitridge1992:539). Para los análisis de interdependencia (Spearman r s) realizados entre este índice modificado y las unidades anatómicas representadas en cada sitio, se consideró el valor de %MAU más alto.
Los valores generales calculados para la especie A. australis fueron sometidos a análisis de covariación con las dos especies de otáridos para las que disponemos de tales valores de referencia: O. flavescens (San Román Reference San Román, López, Cartajena, García and Mena2009) y Z. californianus (Savelle et al. Reference Savelle, Friesen and Lyman1996). De este modo, se busca dar cuenta de las posibles diferencias y similitudes entre especies cercanas. Por último, realizamos un análisis de correlación entre el índice de densidad mineral ósea para A. australis (Borella et al. Reference Borella, Gutiérrez, Foderé, Merlo, Morello, Prieto, Martinic and Bahamonde2007) y los valores de utilidad económica aquí calculados con el objetivo de reducir al mínimo problemas de equifinalidad entre ambos marcos de referencia (León Reference León2012; Lyman Reference Lyman1994). No obstante, para asegurar la aplicabilidad de este índice, de modo que potencialmente dé cuenta de comportamientos humanos relacionados con el transporte y el procesamiento de porciones esqueletarias, es necesario realizar un análisis lo más exhaustivo posible sobre los patrones de preservación diferencial de restos óseos (Marean y Cleghorn Reference Marean and Cleghorn2003).
Resultados
Los pesos totales registrados para el individuo de A. australis analizado en este trabajo son presentados en la Tabla 1. Para corroborar la consistencia en el procedimiento de desmembramiento del individuo, realizamos un cálculo de correlación entre los pesos totales de las porciones anatómicas del lado derecho y las del lado izquierdo. A partir de los resultados obtenidos es posible observar una correlación muy alta entre ambos lados y estadísticamente significativa (r s = 0,99; p < 0,01), es decir, no se detectaron inversiones de valores por diferencias en el procesamiento de cada uno de los lados.
A partir de los datos presentados por Savelle y colaboradores (Reference Savelle, Friesen and Lyman1996) para Z. californianus y por San Román (Reference San Román, López, Cartajena, García and Mena2009) para O. flavescens se realizaron análisis de correlación entre los pesos totales de esas especies y A. australis. Dichas pruebas ofrecieron resultados que indican una relación fuerte y estadísticamente significativa (O. flavescens/A. australis: r s = 0,9; p < 0,05; Z. californianus/A. australis: r s = 0,86; p < 0,05). Sumado a esto, como se puede observar en la Figura 3, al comparar los pesos de los tres individuos, no se perciben diferencias notorias entre la importancia relativa de las porciones anatómicas respecto del peso total, aunque es posible resaltar algunos aspectos. En primer lugar, los porcentajes de peso de la piel y tejido adiposo y de las vísceras de A. australis muestran un valor porcentual intermedio entre las dos especies analizadas previamente, justamente dos de los puntos en los que Z. californianus y O. flavescens indicaron mayor diferencia (San Román Reference San Román, López, Cartajena, García and Mena2009). En segundo lugar, mientras el sector de las costillas mostró un menor porcentaje de peso en A. australis respecto de Z. californianus y O. flavescens, la primera especie supera a las otras dos en el peso correspondiente a las vértebras cervicales. Por último, en el caso de las escápulas, el individuo de A. australis indicó un porcentaje de peso muy similar al de O. flavescens, pero menor al de Z. californianus.

Figura 3. Comparación de frecuencias de pesos totales sobre porciones anatómicas entre Z. californianus, O. flavescens y A. australis.
Para el cálculo del índice de utilidad económica se emplearon los pesos de los elementos anatómicos asociados a huesos y se siguió la propuesta metodológica de Lyman y coautores (Reference Lyman, Savelle and Whitridge1992) para generar los valores de %MUI para el individuo de la especie A. australis recuperado en las aguas del canal Beagle (Tabla 2). A partir de dicho resultado, se calculó su versión modificada (%MMUI; Tabla 2).
Tabla 2. Derivación del %MUI y %MMUI para la Especie A. australis.

Pues bien, si introducimos una comparación con los índices de utilidad generados para las otras dos especies de otáridos, es posible observar que el resultado de la correlación entre el %MUI de Z. californianus y A. australis (r s = 0,95; p < 0,01), al igual que la correlación entre el %MUI de O. flavescens y A. australis (r s = 0,94; p < 0,01), también muestra una relación fuerte y estadísticamente significativa entre las especies. No obstante, si observamos los gráficos de dispersión (Figura 4), algunas unidades anatómicas se alejan levemente del valor esperado.

Figura 4. Gráficos de dispersión entre valores de %MUI: (a) comparación entre A. australis y O. flavescens; (b) comparación entre A. australis y Z. californianus. CER = vértebras cervicales; COS = costillas; TOR = vértebras torácicas; CAB = cabeza; ESC = escápulas; HUM = húmeros; PEL = pelvis; LUM = vértebras lumbares; RAD = radios; EST = esternebras; AA = aletas anteriores; AP = aletas posteriores; TIB = tibias; FEM = fémures.
Si comparamos cada uno de los valores de %MUI entre las tres especies (Figura 5), una de las diferencias que podemos observar es que Z. californianus presenta un contenido cárnico menor, respecto a las otras dos especies, en las porciones correspondientes a los miembros y a la escápula (ver discusión en San Román Reference San Román, López, Cartajena, García and Mena2009). Asimismo, la porción con mayor utilidad económica en Z. californianus y O. flavescens son las costillas, mientras que en el caso de A. australis son las vértebras cervicales. Cabe destacar también que esta última especie muestra comparativamente valores más altos en el sector de las vértebras torácicas, vértebras lumbares, pelvis-sacro-caudales y de la cabeza. En resumen y a los fines del presente trabajo, las principales diferencias observadas entre O. flavescens y A. australis en términos ordinales son las siguientes posiciones: (a) las costillas y las vértebras cervicales se encuentran invertidas (en O. flavescens las costillas ocupan el primer lugar y las vértebras cervicales el segundo, y viceversa para A. australis); (b) la escápula ocupa el tercer lugar en el caso de la primera especie y el quinto en la segunda; (c) las vértebras torácicas se encuentran en el quinto lugar en O. flavescens y en el tercero en el caso de A. australis; (d) el húmero es sexto en el lobo marino de un pelo y octavo en el de dos pelos; (e) las vértebras lumbares ocupan el octavo lugar en O. flavescens y el sexto en A. australis.
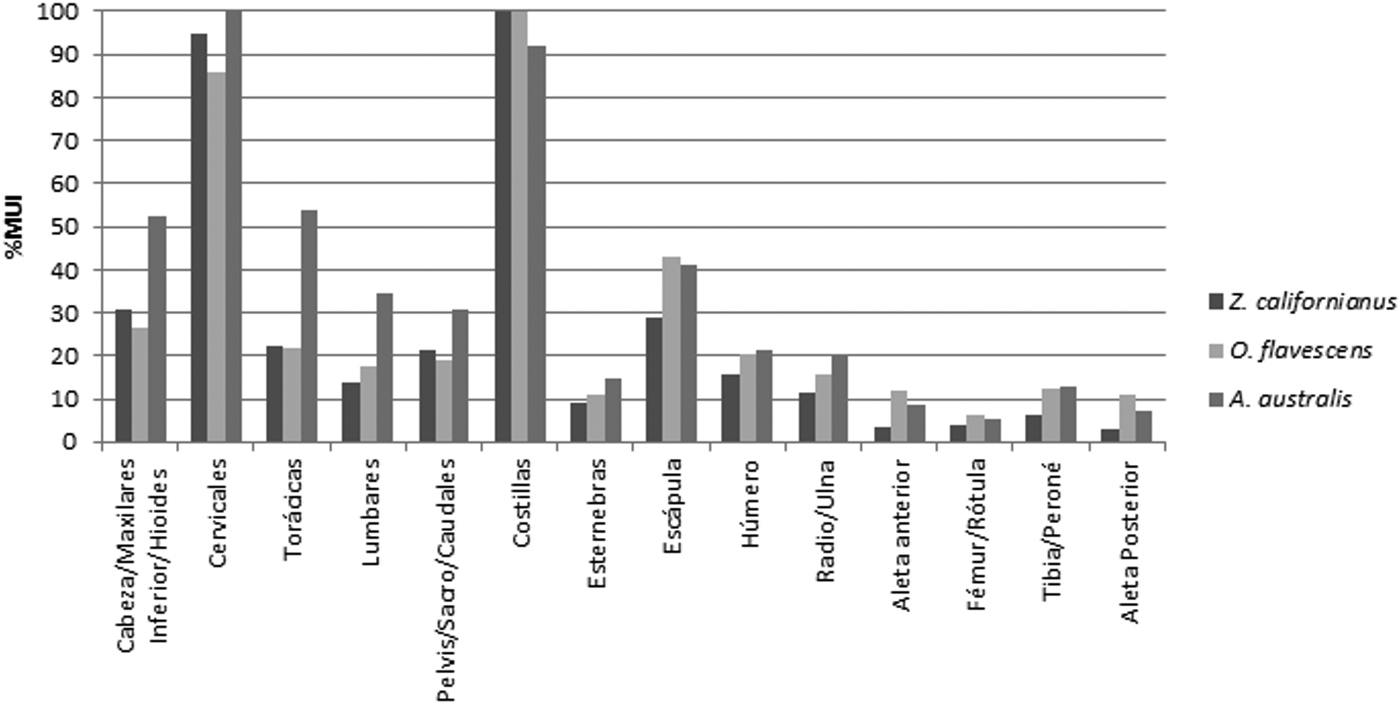
Figura 5. Histograma comparativo sobre los valores de %MUI para Z. californianus, O. flavescens y A. australis.
Por último, realizamos un análisis de correlación entre los valores de %MUI calculados para A. australis y los valores de densidad mineral ósea conocidos para dicha especie (Borella et al. Reference Borella, Gutiérrez, Foderé, Merlo, Morello, Prieto, Martinic and Bahamonde2007), con el objetivo de evaluar posibles sesgos relacionados con el uso de estos índices (Lyman Reference Lyman1994). El resultado obtenido no mostró codependencia entre dichas variables, en tanto tampoco fue estadísticamente significativo (r s = −0,1; p > 0,05).
Aplicación a dos Casos de Estudio en la Región del canal Beagle (Tierra del Fuego)
Como surge del análisis realizado en el apartado previo, las dos especies más comunes en el registro arqueológico fueguino en particular y sudamericano en general presentan algunas diferencias relativas en la importancia económica de las distintas porciones anatómicas. Mientras que en O. flavescens se observan valores más elevados en el sector del pecho (costillas), en A. australis la porción correspondiente al cuello (vértebras cervicales) es la que tiene mayor rinde cárneo. Además, se observan otras inversiones en los valores correspondientes al sector de la espalda y de los miembros. Cabe destacar que, si bien partimos del presupuesto de que debido a las diferencias en la locomoción entre ambas especies (Cadegán Sepúlveda Reference Cadegán Sepúlveda2013) era posible esperar variaciones más significativas en los valores de utilidad económica en el esqueleto apendicular (especialmente en los miembros anteriores), varias de las distinciones identificadas fueron halladas en el esqueleto axial. La diferencia observada adquiere cierta relevancia a la hora de comprender la posible variabilidad en las estrategias de transporte y procesamiento practicadas por los grupos sudamericanos, haciendo foco en las especies de pinnípedos explotadas.
A continuación, presentamos dos casos de estudio de la región del canal Beagle, cuyos perfiles anatómicos fueron evaluados previamente utilizando el índice de utilidad económica desarrollado para la especie O. flavescens (Martinoli Reference Martinoli2015, Reference Martinoli2017; Martinoli y Vázquez Reference Martinoli, Vázquez, Mondini, Muñoz and Fernández2017). Cabe destacar que en trabajos anteriores se ha testeado si la representación anatómica en tales conjuntos es producto de la preservación diferencial de restos considerando a la densidad mineral ósea (Borella et al. Reference Borella, Gutiérrez, Foderé, Merlo, Morello, Prieto, Martinic and Bahamonde2007) como factor estructurante del conjunto, pero en ninguno de los casos se encontró una relación de dependencia entre esas variables (%MAU Segundo Componente de Imiwaia I/densidad ósea: r s = 0,3; p > 0,05 [Martinoli Reference Martinoli2015]; %MAU Ajej I/densidad ósea: r s = 0,2; p > 0,05 [Martinoli Reference Martinoli2018]).
El archipiélago fuego-patagónico tiene una historia de ocupaciones humanas centradas en la explotación de recursos marinos que se remonta hasta los 6500 años aP no calibrados (Legoupil Reference Legoupil1997; Orquera y Piana Reference Orquera and Piana1999; San Román Reference San Román2010, Reference San Román2016; Schiavini Reference Schiavini1990, Reference Schiavini1993). Específicamente, los sitios arqueológicos de la región del canal Beagle muestran un claro predominio de restos óseos de pinnípedos (A. australis), es decir, particularmente estas presas eran las que aportaban el mayor ingreso energético a la dieta de los grupos del área (Martinoli Reference Martinoli2015; Orquera y Piana Reference Orquera and Piana1999, Reference Orquera and Piana2009; Schiavini Reference Schiavini1990, Reference Schiavini1993).
El sitio de Imiwaia I es un conchero multicomponente, emplazado en la bahía Cambaceres Interior (Figura 1). El mismo fue interpretado como un área residencial en la cual se realizaban múltiples actividades (Orquera y Piana Reference Orquera and Piana1999). El Segundo Componente del sitio (capas K, L y M), con una cronología de entre 6000 y 5700 años no calibrados aP (Zangrando Reference Zangrando2009:84), se encuentra dominado por restos de peces; no obstante, los pinnípedos representaron recursos importantes en la dieta de los grupos que desarrollaron allí sus actividades (10,2% del NISP% total; Martinoli Reference Martinoli2015, Reference Martinoli2018; Zangrando Reference Zangrando2009). El análisis del perfil anatómico de dichas presas mostró que la mayoría de los elementos se encontraban representados, pero con valores de %MAU variables. El examen de tales porcentajes a la luz del índice de utilidad económica generado para la especie O. flavescens arrojó como resultado una correlación con una tendencia moderada hacia la representación mayoritaria de porciones con bajo valor económico, aunque esta relación no es estadísticamente significativa (%MUI de O. flavescens y %MAU de Imiwaia I: r s = −0,49; p > 0,05; Martinoli Reference Martinoli2015, Reference Martinoli2018). Sin embargo, los perfiles taxonómicos se encuentran dominados por A. australis (8:1; Martinoli Reference Martinoli2015).
Si correlacionamos el %MUI de A. australis y el %MAU del Segundo Componente de Imiwaia I el resultado es el siguiente: r s = −0,54; p = 0,06. Evidentemente el %MUI de A. australis se ajusta mejor, aunque el cálculo de correlación sigue sin ser estadísticamente significativo. No obstante, al utilizar el %MUI de A. australis observamos en el gráfico de dispersión (Figura 6) que la tendencia que ya se advertía entre %MAU y %MUI de O. flavescens se acentúa, es decir, que los huesos que se encuentran representados en un porcentaje mayor son aquellos que tienen una utilidad económica baja (reverse bulk strategy [Binford Reference Binford1978; Thomas y Mayer Reference Thomas, Mayer and Thomas1983]). En este punto es necesario decir que el grado de significación de las correlaciones aumentará en la medida que las dos series de variables tiendan a ajustarse de manera lineal, situación característica de la curva que ilustra el modelo unbiased o su inversa (Binford Reference Binford1978; Thomas y Mayer Reference Thomas, Mayer and Thomas1983). Sin embargo, los casos que se asemejen a las curvas que modelizan las estrategias denominadas bulk y gourmet o sus manifestaciones inversas posiblemente no se ajusten a correlaciones estadísticas con altos niveles de significación; en estos casos, la visualización de las distribuciones de puntos podría ser más informativa que el resultado de las correlaciones (Vázquez Reference Vázquez2015).

Figura 6. Gráfico de dispersión entre los valores de %MAU del Segundo Componente de Imiwaia I y los valores de %MUI de A. australis y de O. flavescens.
Ajej I es un sitio monocomponente —un conchero aislado cuyo aspecto superficial denotaba un domo poco potente— ubicado en la costa de bahía Golondrina, en el margen norte del canal Beagle (Figura 1), excavado en el marco de una campaña de rescate (Piana et al. Reference Piana, Vázquez and Álvarez2008). Este yacimiento fue interpretado como un espacio de actividades específicas y acotadas (Piana et al. Reference Piana, Vázquez and Álvarez2008). Las muestras de carbón fechadas indican que la cronología del conjunto se ubica entre 1400 ± 90 y 1270 ± 180 años no calibrados aP (Piana et al. Reference Piana, Vázquez and Álvarez2008:107). En este sitio los especímenes de pinnípedos han sido relativamente los más abundantes (57,8% del NISP total; Martinoli y Vázquez Reference Martinoli, Vázquez, Mondini, Muñoz and Fernández2017; Piana et al. Reference Piana, Vázquez and Álvarez2008). Al igual que en el Segundo Componente de Imiwaia I, la mayoría de los elementos anatómicos de pinnípedos se hallaron representados, aunque en porcentajes variables, e incluso se observó la ausencia de la porción sacro-lumbar (Martinoli y Vázquez Reference Martinoli, Vázquez, Mondini, Muñoz and Fernández2017; Piana et al. Reference Piana, Vázquez and Álvarez2008). Respecto de la correlación entre los valores de %MAU de Ajej I y los valores de %MUI correspondientes a O. flavescens, los resultados obtenidos previamente arrojaron un resultado débil y estadísticamente no significativo (r s = −0,19; p > 0,05; Martinoli y Vázquez Reference Martinoli, Vázquez, Mondini, Muñoz and Fernández2017)Footnote 2. Ahora bien, en este conjunto todos los especímenes identificados corresponden a la especie A. australis (Martinoli Reference Martinoli2017; Piana et al. Reference Piana, Vázquez and Álvarez2008).
Si correlacionamos %MUI de A. australis y %MAU de Ajej I el resultado es el siguiente: r s = −0,4; p > 0,05. Esto denota una mayor relación entre variables, aunque sigue sin ser estadísticamente significativa. No obstante, en términos generales, observamos que a partir del uso del índice generado para A. australis surge una posible tendencia que no se observaba a partir de la utilización del %MUI para O. flavescens (Figura 7): mayor abundancia de porciones de moderado y bajo valor económico, lo que se describe como reverse bulk strategy (Binford Reference Binford1978; Thomas y Mayer Reference Thomas, Mayer and Thomas1983).

Figura 7. Gráfico de dispersión entre los valores de %MAU de la Capa C de Ajej I y los valores de %MUI de A. australis y de O. flavescens.
Tanto en la Capa C de Ajej I como en el Segundo Componente de Imiwaia I las correlaciones entre el %MUI para A. australis y el %MAU y la densidad mineral ósea para A. australis y el %MAU no arrojaron resultados estadísticamente significativos (ver más arriba), por lo cual si evaluamos estos conjuntos en función del modelo propuesto por Lyman (Reference Lyman1994) para la identificación de patrones de selección y descarte, ambos conjuntos formarían parte de la Clase 5, es decir, no es posible atribuir inequívocamente a ninguna de dichas variables la estructura del conjunto (Lyman Reference Lyman1994).
Ahora bien, dados tales resultados y como fue especificado en el apartado de métodos, también se calculó el %MMUI (modified meat utility index). Este índice permite considerar aquellas porciones anatómicas de menor valor económico que pueden ingresar a los sitios adosadas a porciones con un porcentaje de carne mayor (Binford Reference Binford1978). La derivación del %MMUI aquí presentada tiene implícita la elección de combinaciones de elementos anatómicos a partir de las referencias etnográficas sobre estrategias de reducción y transporte de carcasas de focas entre grupos Inuit de Canadá (Lyman et al. Reference Lyman, Savelle and Whitridge1992), a diferencia de trabajos como el de Savelle y coautores (Reference Savelle, Friesen and Lyman1996) y San Román (Reference San Román, López, Cartajena, García and Mena2009). Preliminarmente, no podemos afirmar que dichas combinaciones sean directamente aplicables en el caso de Otariidae, ya que existen algunas diferencias anatómicas, tales como el tamaño y peso de las aletas, entre estos y los fócidos (San Román Reference San Román, López, Cartajena, García and Mena2009, Reference San Román2016). Sin embargo, consideramos su aplicación en los dos conjuntos aquí presentados, ya que es la única combinación para la que se explicita la fuente a partir de la cual ha sido confeccionada. En el caso del Segundo Componente de Imiwaia I el resultado de la correlación entre el %MAU y el %MMUI para la especie A. australis arrojó un resultado negativo moderadamente fuerte y estadísticamente significativo (r s = −0,58; p < 0,05). Volviendo al modelo propuesto por Lyman (Reference Lyman1994) y a la luz de dicho resultado, el conjunto correspondiente al Segundo Componente de Imiwaia I correspondería entonces a la Clase 2, es decir, la selección o descarte en el yacimiento de porciones de bajo rendimiento económico puede ser la causa del perfil anatómico observado. Con respecto al sitio Ajej I el resultado de la correlación entre dichas variables sigue mostrando una relación negativa; sin embargo, no es estadísticamente significativo (r s = −0,3; p > 0,05), lo cual sigue siendo compatible con la Clase 5 propuesta por Lyman (Reference Lyman1994).
Tanto en el Segundo Componente de Imiwaia I como en la Capa C de Ajej I, a través de la aplicación del %MMUI calculado para A. australis (Figura 8), es posible observar la puesta en práctica de estrategias de transporte de porciones esqueletarias relativamente similares, aunque más claramente marcada en el conjunto antiguo: se encuentran más representados los conjuntos anatómicos con menor rinde económico. No obstante, mientras que Imiwaia I fue considerado un área residencial (Orquera y Piana Reference Orquera and Piana1999), Ajej I fue interpretado como un sector de actividades específicas (Piana et al. Reference Piana, Vázquez and Álvarez2008). Los resultados aquí presentados podrían indicar que los sitios tenían funcionalidades comparables, ya que el perfil anatómico en ambos es el esperado en sectores de matanza y procesamiento primario (Binford Reference Binford1978); sin embargo, en Ajej I los resultados no son inequívocos, es decir, no es posible descartar un fenómeno de equifinalidad. De todos modos, como hemos indicado en el presente trabajo, no hemos hallado correlaciones positivas entre el %MUI para A. australis y los valores de densidad mineral ósea para dicha especie. Por lo tanto, es necesario considerar estos resultados realizando una evaluación integral de todas las evidencias presentes en los yacimientos. Por un lado, los perfiles anatómicos observados podrían ser indicadores de las estrategias de procesamiento seleccionadas, como por ejemplo la decisión de transportar al campamento residencial las porciones más dificultosas de procesar por su bajo contenido de carne (e.g., vértebras lumbares-pelvis-sacro-caudales o fémur-tibia-peroné-aletas posteriores; Lupo Reference Lupo2006; Lyman et al. Reference Lyman, Savelle and Whitridge1992). Por otro lado, la selección de partes esqueletarias puede relacionarse con actividades que no necesariamente sean de subsistencia (Lupo Reference Lupo2006). Tal es el caso del transporte de determinados elementos óseos considerando sus propiedades mecánicas y morfológicas en función de la confección de determinado instrumental (Christensen Reference Christensen2016), independientemente de su aporte energético directo a la dieta humana. Por ejemplo, en la región del canal Beagle se han identificado instrumentos óseos realizados sobre radios y ulnas de pinnípedos (Orquera y Piana Reference Orquera and Piana1999:68, Figura 30), dos elementos cuyos músculos asociados presentan un valor de %MUI bajo (19,7%).

Figura 8. Gráfico de dispersión entre los valores de %MAU del Segundo Componente de Imiwaia I y de Ajej I y los valores de %MMUI de A. australis.
Consideraciones Finales
Los índices de utilidad económica son herramientas importantes y de probado valor metodológico para acercarnos a un conocimiento más profundo de las estrategias de transporte, procesamiento, consumo y descarte de las carcasas puestas en práctica por las sociedades pretéritas. Por lo tanto, es fundamental el desarrollo de índices de utilidad económica específicos para las especies que dominan los conjuntos faunísticos sudamericanos. Tal como se muestra en el presente trabajo, en el caso de los pinnípedos, no sólo existen diferencias en la distribución muscular entre fócidos y otáridos (San Román Reference San Román, López, Cartajena, García and Mena2009), sino también entre dos especies de otáridos, ambas representadas en los yacimientos del sur del archipiélago fueguino a lo largo del Holoceno y en muchos registros zooarqueológicos del sector sur de la amplia región biogeográfica del neotrópico.
En términos generales, mientras los individuos de O. flavescens presentan mayor desarrollo muscular en el sector de las costillas, los A. australis lo hacen en las vértebras cervicales. Tal es así que la aplicación del índice desarrollado en el presente trabajo en conjuntos con predominio de especímenes de la especie A. australis ha mostrado un ajuste relativamente más adecuado respecto de las tendencias observadas usando los valores para O. flavescens. Específicamente, en la aplicación del %MMUI al conjunto del Segundo Componente de Imiwaia I se corrobora la puesta en práctica de una posible estrategia del tipo bulk (inversa). En el caso de Ajej I, únicamente se observó el surgimiento de una posible tendencia hacia la mayor abundancia de porciones de bajo y moderado valor económico; sin embargo, cabe la posibilidad de que estemos observando un caso de equifinalidad, el cual no había sido explicitado en trabajos previos. Dicho fenómeno debe ser un aspecto a evaluar en el desarrollo de futuros trabajos zooarqueológicos que se centren en el análisis de restos óseos de pinnípedos, considerando especialmente el uso combinado de distintos índices que permitan evaluar los perfiles anatómicos. También es necesario, por un lado, discutir y mejorar la forma en la que se cuantifican las unidades anatómicas de pinnípedos y, por otro lado, comenzar a aplicar modelos, como los generados para mamíferos terrestres, que tomen en consideración el potencial de supervivencia de los elementos óseos (ver Marean y Cleghorn Reference Marean and Cleghorn2003) correspondientes a mamíferos marinos.
Se considera fundamental seguir aportando al desarrollo de marcos de referencia específicos para nuestras áreas de estudio, ya que muchos de ellos fueron generados a partir de trabajos etnoarqueológicos y experimentales que no se corresponden necesariamente con las estrategias y presas seleccionadas por distintos grupos humanos que habitaron diferentes ambientes y que se enfrentaron con contingencias variables y particulares.
Agradecimientos
A todas las personas que colaboraron con el procesamiento del individuo y a su identificación: Natalia Dellabianca, Luciana Riccialdelli, Adrián Schiavini, Natalia Paso Viola, Daniela Alunni, Angélica Tivoli y Germán Pinto-Vargas. A Néstor del club AFASyN por haber recolectado el individuo aquí estudiado: su aporte fue más que fundamental para el desarrollo de este trabajo. A la Dra. Fiorito, por la identificación de la causa de muerte del espécimen. A Manuel San Román por los intercambios que tuvimos respecto del desarrollo de este índice. Al Museo Acatushún por permitirme acceder a las muestras comparativas allí alojadas. A Francisco Zangrando, Daniela Alunni, Martín Vázquez, Florencia Borella y María Gutiérrez por las lecturas realizadas sobre el manuscrito. A los evaluadores anónimos por las sugerencias fundamentales que realizaron. Este trabajo se realizó en el marco de los proyectos PICT 2013/1011 y PIP 0387.
Declaración de disponibilidad de datos
Repositorio institucional Centro Austral de Investigaciones Científicas (CADIC-CONICET), Bernardo Houssay 200, Ushuaia, Tierra del Fuego, Argentina.










