



Southwest coastal Ecuador was a hearth of innovation in the prehistoric New World. Plant domestication began before 8500 BC, with squash, gourd, and llerén identified at Las Vegas (Site 80), and maize introduced by 5800 BC (Piperno and Pearsall Reference Piperno and Pearsall1998; Stothert et al. Reference Stothert, Piperno and Andres2003; all dates calibrated). Site 80 was a base camp and burial site for a seasonally mobile population. By 4400 BC, life in southwestern Ecuador had become sedentary village life, and ceramic production, agriculture, polished stone tool production, and figurine ideology came together to form the Early Formative Valdivia culture (Zeidler Reference Zeidler, Silverman and Isbell2008). Valdivia culture was originally identified at the Valdivia type-site and investigated in the 1950s and 1960s by Emilio Estrada, Betty Meggers, and Clifford Evans. Some 100 Valdivia sites are now known to archaeologists (Marcos Reference Marcos, Raymond and Burger2003). By the Middle Valdivia (2800–2400 BC), one village, Real Alto, was transformed into a town, with large houses, packed neighborhoods, and ceremonial mounds (Clark et al. Reference Clark, Gibson, Zeidler, Bandy and Fox2010).
In this article we investigate the foodways of Middle Valdivia Real Alto and consider how they reflect and inform the community's social relationships and economy. We discuss new research that enables us to evaluate the importance of different domesticated plants, including maize, through microfossils recovered from stone tools from residential and ceremonial structures and from human dental calculus. We demonstrate that maize played a role in both the domestic and ceremonial lives of the Real Alto population. It was an important component of daily meals and ceremonial foods but only one of the crops making up Valdivia dry tropical forest agriculture. Contextualizing the preparation, consumption, and discarding of maize and other foods within the site—and the site within the agricultural system of the region—enhances our understanding of the development of agriculture in southwestern Ecuador and social transformations at Real Alto.
Real Alto in Perspective
Real Alto is located on a low ridge between the seashore and the Río Verde, near the transition between xerophytic coastal plain and inland deciduous tropical forest (Figure 1; Neill Reference Neill, Jørgensen and León-Yánez1999). This location provided access to riverine and marine resources and to the Chanduy estuary 4 km downriver. Real Alto was excavated from 1974 to 1975 by the late Donald W. Lathrap and students (Figure 2; Lathrap et al. Reference Lathrap, Marcos and Zeidler1977; Marcos Reference Marcos1978, Reference Marcos1988a, Reference Marcos1988b; Pearsall Reference Pearsall1979; Zeidler Reference Zeidler1984), with excavations in 1977, 1984, and 1986 by Damp (Reference Damp1979, Reference Damp1984a); in 1984 by faculty and students of Centro de Estudios Arqueológicos y Antropológicos of the Escuela Superior Politécnica del Litoral (ESPOL), Guayaquil; and most recently by Russian archaeologists working with ESPOL (Tabarev et al. Reference Tabarev, Kanomata, Marcos, Popov and Lazin2016).
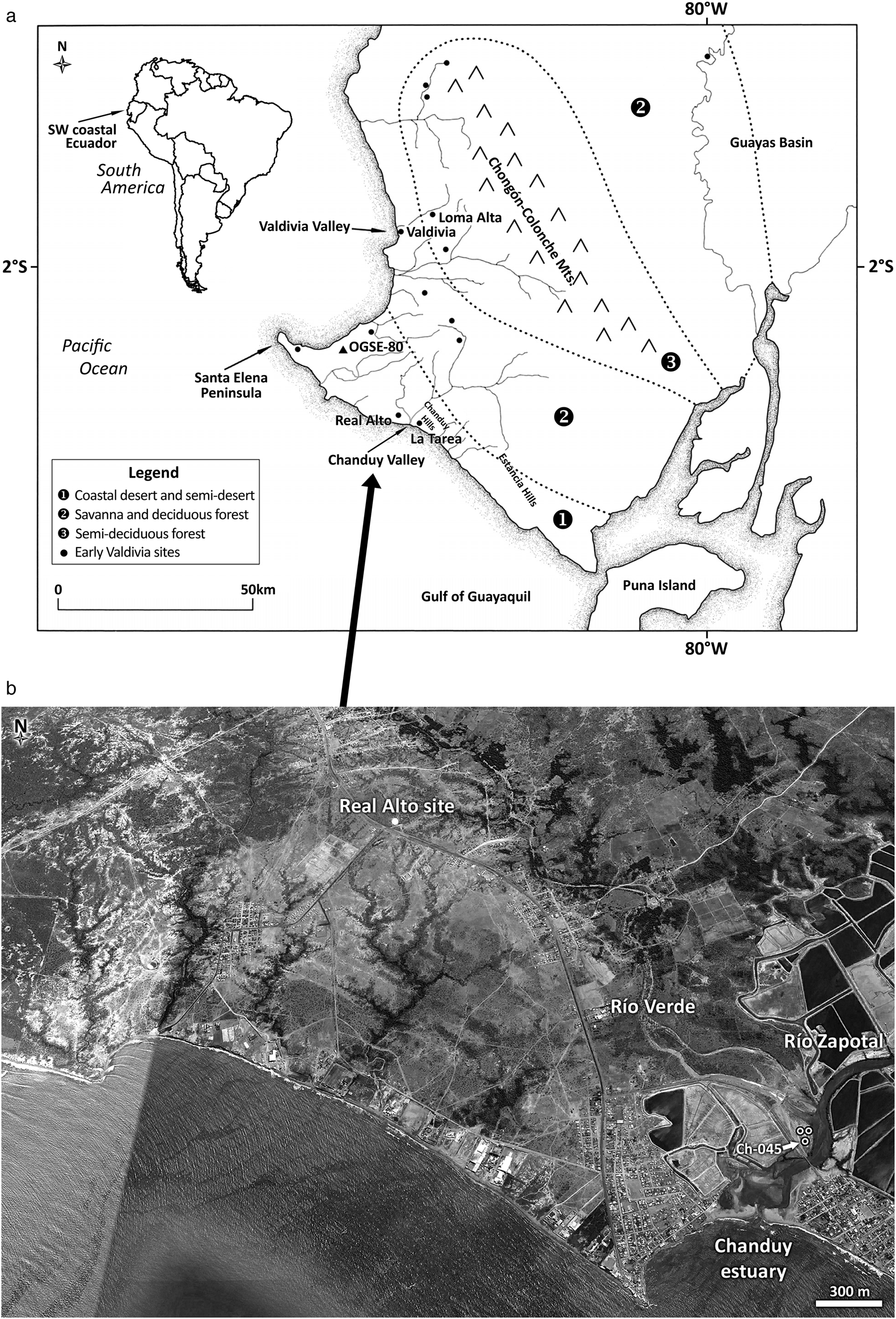
Figure 1. Location of the Real Alto site in southwestern coastal Ecuador and the relationship of this site to vegetation zones, rivers, archaeological sites mentioned in the text, and the Chanduy estuary.

Figure 2. Excavations at Real Alto, 1974–1975. (a) Site setting today, looking across the Río Verde; (b) view across the inner plaza, looking toward the Charnel House Mound; (c, d) excavations reveal densely packed domestic structures. Note the structure of the wall trenches (c, foreground and left) and floor excavations in progress (d).
Real Alto was occupied during Valdivia Phases 1–7 (4400–1800 BC; see Table 1; Marcos and Michczynski Reference Marcos and Michczynski1996; Tabarev et al. Reference Tabarev, Kanomata, Marcos, Popov and Lazin2016; Zeidler Reference Zeidler, Raymond and Burger2003). During this time it grew from a U-shaped village of 12–15 houses grouped around a small plaza (Early Valdivia, Phases 1a–2b) to a 12.4 ha town formed by a ring of larger domestic structures around a central ceremonial precinct with two mounds (Middle Valdivia, Phases 3–5; see Figure 3), until contracting in size and population (Late Valdivia, Phases 6–7) and remaining an active ceremonial center until it was abandoned (Clark et al. Reference Clark, Gibson, Zeidler, Bandy and Fox2010; Lathrap et al. Reference Lathrap, Marcos and Zeidler1977; Marcos Reference Marcos1978, Reference Marcos1988a, Reference Marcos1988b, Reference Marcos, Raymond and Burger2003; Zeidler Reference Zeidler1984, Reference Zeidler1991; Zeidler and McEwan Reference Zeidler, McEwan, Cockrell, Hoopes and McEwan2020).

Figure 3. The Phase 3 Real Alto town, with excavation trenches (A, B, C), ceremonial mounds (Feasting or Fiesta Mound, Funerary or Charnal House Mound), and sampled structures indicated.
Table 1. Chronological Chart of Valdivia Cultural Sequence and Ceramic Phases.

Notes: Calibrated radiocarbon dates and thermoluminescence determinations used to determine the ceramic phases and calibrated ranges. Based on Marcos and Michcynski (Reference Marcos and Michczynski1996:Table 4; see also Zeidler Reference Zeidler, Raymond and Burger2003).
The transformation of Real Alto during the Middle Valdivia involved significant changes in community and household size and configuration (Clark et al. Reference Clark, Gibson, Zeidler, Bandy and Fox2010; Marcos Reference Marcos1978, Reference Marcos, Raymond and Burger2003; Zeidler Reference Zeidler1984, Reference Zeidler1991). The settlement grew to four times the size of the earlier village, incorporating both domestic (outer ring of densely packed houses) and ceremonial/public (inner plaza with two mounds) spaces. Two mounds—the Fiesta or Feasting Mound, named for numerous drinking bowl fragments found in structures built on it, and the Charnel House Mound, named for burials in the mound-top structure, Structure 7—faced each other, dividing a formerly single communal space into two unequal spaces. These spatial relationships and the differences in the mounds suggest that paired complementarities in social structure existed (Lathrap et al. Reference Lathrap, Marcos and Zeidler1977; Zeidler Reference Zeidler1984; Zeidler and McEwan Reference Zeidler, McEwan, Cockrell, Hoopes and McEwan2020). Rituals had become formalized within structures, with different kinds of rituals on the mounds.
Mean house sizes increased from 14.4 m2 to 49.10 m2 between Phases 1 and 3, and houses were in tight clusters of five to six (Clark et al. Reference Clark, Gibson, Zeidler, Bandy and Fox2010; Zeidler Reference Zeidler1984). From this arrangement it is inferred that much larger groups (likely extended families) lived within houses and related families (descent groups) lived near each other. Two size groupings of houses were identified (40–50 m2, 60–70 m2) suggesting differences in the relative social standing of families. These changes in community and household structure, among other indicators of evolving social conditions, indicate the emergence of a nascent social hierarchy by the Middle Valdivia, with higher-status individuals identified by burial in the Charnel House Mound (Lathrap et al. Reference Lathrap, Marcos and Zeidler1977; Marcos Reference Marcos1978, Reference Marcos, Raymond and Burger2003; Zeidler Reference Zeidler1984, Reference Zeidler1991; Zeidler and McEwan Reference Zeidler, McEwan, Cockrell, Hoopes and McEwan2020).
Archaeobotanical and faunal remains were recovered systematically. Phytolith samples, flotation-recovered and in situ macroremains from the 1974–1975 excavations, and in situ and fine-sieved samples from Damp's excavations were analyzed by Pearsall (Reference Pearsall1979, Reference Pearsall, Raymond and Burger2003; Damp and Pearsall Reference Damp and Pearsall1994; Damp et al. Reference Damp, Pearsall and Kaplan1981). Faunal remains recovered in quarter-inch mesh excavation screens during the 1974–1975 excavations were later analyzed by Byrd (Reference Byrd, Reitz, Newsom and Scudder1996) and Stahl (Reference Stahl, Raymond and Burger2003; Stahl and Zeidler Reference Stahl, Zeidler, Saunders and de Montmollin1988, Reference Stahl and Zeidler1990). Charred wood fragments were the only macroremains recovered in quantity using the flotation method in 1974–1975. There were few other charred remains, and the ones recovered were mostly unidentifiable. Fine-sieving led to the recovery of less fragmented nonwood remains from Early Valdivia contexts. Economic plants identified in macrobotanical and phytolith records in Early to Middle Valdivia contexts included maize (Zea mays), canna (Canna), arrowroot (Maranta), squash/gourd (Cucurbitaceae), cotton (Gossypium), and jack bean (Canavalia). Charred wood fragments (Pearsall Reference Pearsall2019:Supplemental Table 1 [doi:10.6067/XCV8448928]) documented a focus on xerophytic-zone legume trees for fuel throughout the Real Alto occupation, with no indication of cutting of inland deciduous forests. Commonly used vertebrate fauna from Middle Valdivia contexts included marine catfish (Ariidae), drum (Sciaenidae), cartilaginous fish (Chondrichthyes), sea bass (Serranidae), sting ray (Dasyatidae), jack and pompano (Carangidae), and white-tail deer (Odocoileus virginianus). Other marine and estuarine fish—snook (Centropomus), lookdown (Selene), snapper (Lutjanus), croaker (Micropogon), and mullet (Mugil)—occurred in low numbers. Invertebrate fauna included ark shell (Anadara), thorny oyster (Spondylus), horn shell (Cerithidea), and tun shell (Malea), indicating the use of intertidal, mangrove, and shallow to moderately deep marine habitats.
Some 100 human skeletons were recovered from Real Alto. Klepinger (Reference Klepinger1979) constructed an abridged life table based on 72 Phase 3 individuals. Demographic data were comparable to other early skeletal series and indicated a natural population profile. Ubelaker (Reference Ubelaker, Raymond and Burger2003) examined the entire population and compared it to other Ecuadorian skeletal series. Life expectancy at birth was slightly lower at Real Alto than during preceramic Las Vegas or the subsequent Late Formative/Regional Developmental period. Stature was similar to mean values from these periods. Ten individuals showed trauma, a higher proportion than observed earlier or later. The frequencies of infection increased over time, likely correlated to increasing population densities. Rates of dental hypoplasia (1.7%) and dental caries (8.5%) were higher at Real Alto than at Las Vegas (1.0% and 3.0%, respectively). The markedly higher Real Alto caries rate was consistent with either higher rates of consumption of cariogenic foods such as maize, changes in food production that increased caries formation, or both (Ubelaker Reference Ubelaker, Raymond and Burger2003). In comparison to the population at Las Vegas, that at Middle Valdivia Real Alto had a slightly higher birthrate and family size, as well as a marked increase in the ratio of juveniles and elderly to productive population members (Clark et al. Reference Clark, Gibson, Zeidler, Bandy and Fox2010). Overall fertility increased to a level consistent with Neolithic populations in other parts of the world (Bocquet-Appel Reference Bocquet-Appel2002), providing evidence for a Neolithic Demographic Transition for coastal Ecuador between Las Vegas and Middle Valdivia (Clark et al. Reference Clark, Gibson, Zeidler, Bandy and Fox2010).
This earlier research at Real Alto enhanced and even transformed our understanding of Early Formative Valdivia culture. Questions nevertheless remained regarding the nature of food production at the community and the role of agriculture in the emergence of social complexity during the Formative period (Pearsall Reference Pearsall, Raymond and Burger2003). Although earlier research had identified a suite of domesticated plants, including maize, it remained challenging to evaluate maize's role in the domestic economy, the provisioning of elites, or ceremonial life and to understand the interplay of environmental and social factors in the development of agricultural societies in the region.
These issues were addressed through a program of new research incorporating (1) analysis of microfossil residues from previously excavated stone tools and human skeletal remains from Real Alto and (2) extraction and analysis of sedimentary cores from estuaries in the region. The coring study (Pearsall et al. Reference Pearsall, Duncan, Jones, Freidel, Veintimilla and Neff2016) informs this article's discussion of Real Alto in its regional context. The microfossil residue research was accomplished in stages: formalizing and testing maize cob diagnostic phytoliths (Pearsall et al. Reference Pearsall, Chandler-Ezell and Chandler-Ezell2003), identifying maize phytolith and starch residues on Structure 20 tools (Pearsall et al. Reference Pearsall, Chandler-Ezell and Zeidler2004), formalizing diagnostic phytoliths for underground storage organs and other edible plant parts of economic plants and then investigating the occurrence of roots and fruits on Structure 20 tools (Chandler-Ezell et al. Reference Chandler-Ezell, Pearsall and Zeidler2006), and identifying plant residues on stone tools from three domestic and two ceremonial structures and in dental calculus samples (as discussed later).
Residue Study
Sample Contexts
We targeted floor deposits within three Phase 3 domestic structures (Structures 1, 10, 20) as a means of inferring food preferences and food preparation and consumption activities within households that sheltered extended families. Artifact and ecofact deposition in floors can provide valuable insights into the “life rhythms” of household members and their use of dwelling space (Boivin Reference Boivin2000). To compare domestic foodways and the use of food in ritual, we analyzed tools from the Phase 3 Charnel House, Structure 7, and a Phase 2 ceremonial structure—S-CHM-1/Feature 306—underlying Structure 7. These contexts are briefly described here; for detailed treatment of the domestic dwellings, see Zeidler (Reference Zeidler1984, Reference Zeidler1991, Reference Zeidler1998), and for ceremonial structures, see Marcos (Reference Marcos1978, Reference Marcos1988a, Reference Marcos1988b). Figure 3 shows the structure locations.
Domestic Structures
Structure 1 was a large (11.5 × 8.5 m) oval-shaped house with a 62.28 m2 floor area at the large end of the Phase 3 house size continuum (Figure 4). Structure 10 was a small (8.55 × 7.40 m) oval-to-round house located near Structure 1 (Figure 5). Its floor area (35.65 m2) fell well below the Phase 3 mean. Structure 20 was a somewhat larger (8.0 × 6.5 m; 39.14 m2 floor area) oval-shaped house located at the northwestern end of the town (Figure 6). Like many Middle Valdivia houses, these structures were defined by deep, encircling wall trenches and a small entryway at one end (not visible in Structure 10). Floor deposits were consolidated sediments 20–25 cm thick that were replete with discarded and trampled artifacts, including ceramic sherds, chipped stone and debitage, ground and polished stone fragments, and faunal remains (Stahl and Zeidler Reference Stahl, Zeidler, Saunders and de Montmollin1988, Reference Stahl and Zeidler1990).

Figure 4. Structure 1. Locations of tools within 1 × 1 m floor units. Units A-2 and A-3 are from site trenching that revealed the structure. Adapted from Zeidler (Reference Zeidler1984).

Figure 5. Structure 10. Locations of tools within the section of floor deposits revealed in unit A-11. Adapted from Zeidler (Reference Zeidler1984).

Figure 6. Structure 20. Locations of tools within 1 × 1 m floor units. Units C-14, C-15, and C-16 are from site trenching that revealed the structure. Adapted from Zeidler (Reference Zeidler1984).
Excavation of Structure 1 and Structure 20 floors proceeded in 1 × 1 m units, in 5 cm layers, with the exception of larger units that were part of site trenching through which the structures were discovered. Structure 1 contained numerous post molds (for partition walls and furnishings), a stone hearth in the center front, several shallow pits, a marine shell cache, and a wall trench burial (Burial 3, Feature 16) at the back of the structure. Based on entryway placement, Structure 1 was oriented to the southeast. Floor deposits yielded two radiocarbon determinations of 4050 ± 185 RCYBP (#GX-7429) and 3845 ± 240 RCYBP (#GX-7430), which after calibration place it within Phase 3. Structure 20 was oriented to the southwest and had a circular hearth to the right center of the floor.
Structure 10 was one of six Phase 3 dwellings arrayed in a superimposed cluster, with Structures 28 and 29 lying below Structure 10, which is then partially superimposed by Structures 2, 22, and 40. Because of this dense superimposition, only a small portion of Structure 10—the portion that fell within unit A-11 of excavation Trench A—was excavated. Excavation proceeded in 5 cm layers, and all cultural materials were piece-plotted in situ.
Ceremonial Structures
Structure 7 is interpreted as a charnel or funerary structure (Marcos Reference Marcos1978, Reference Marcos1988a, Reference Marcos1988b) used during Phase 3 for the interment of highly ranked kin groups (Figure 7a; Zeidler and McEwan Reference Zeidler, McEwan, Cockrell, Hoopes and McEwan2020). Although oval shaped (9.5 × 6.0 m), it differed from domestic structures in having a wide doorway (2.30 m) on one side, rather than on one end. The wall trench on either end of the oval was “pinched,” forming narrower floor spaces at each end of the building. Structure 7 is also noteworthy in that its entryway was oriented northeast at an azimuth consistent with the June solstice sunrise (Zeidler Reference Zeidler1998; Zeidler and McEwan Reference Zeidler, McEwan, Cockrell, Hoopes and McEwan2020). Individuals were interred in two areas: (1) at the entryway threshold—a 35-year-old female was interred in a unique stone-lined crypt in association with stacked secondary burials of males, and (b) primary and secondary interments and surficial skeletal remains were found in the floor of the north half of the structure (Marcos Reference Marcos1988a). Two radiocarbon dates place Structure 7 at the earlier end of Phase 3 (#ISGS-439, 4100 ± 80; #ISGS 446, 4270 ± 80; Liu et al. Reference Liu, Riley and Coleman1986:106). The floor deposits were similar to those of domestic houses, with dense artifact concentrations and abundant faunal remains. Floor depth was shallower, consisting of a single 10 cm level. The floor was gridded into 1 × 1 m units, and materials were piece-plotted before being excavated as a single level within each unit (Marcos Reference Marcos1988a).

Figure 7. Ceremonial structures. (a) Locations of tools within 1 × 1 m floor units of Structure 7, Phase 3 Charnel House; (b) locations of tools in Structure S-CHM-1/Feature 306, Phase 2 ceremonial structure beneath Structure 7. Adapted from Marcos (Reference Marcos1978).
Underlying Structure 7 was a smaller burned structure designated S-CHM-1, of ceremonial (but not funerary) function dating to Phase 2 (Figure 7b). It was the first structure associated with mound building at Real Alto (Marcos Reference Marcos1978, Reference Marcos1988a, Reference Marcos1988b). Archaeologically it is represented by Feature 306, located 120 cm below Structure 7, but the spatial extent of this feature (2.4 × 6.4 m) was larger than the mapped extent of Feature 306. S-CHM-1 was a burned wattle-and-daub building interpreted as a feasting or meeting house. In addition to burned daub, the floor contained shellfish, mammal bone, ground-stone fragments, and 29 Phase 2 potsherds. A small ramp leading to the mound top contained three dedicatory offerings: a conch shell (Strombus peruvianus) trumpet, a Phase 2 tetrapod bowl (Feature 287), and a Phase 2 tripod bowl (Feature 282). A single radiocarbon assay dates this early mound-building context to 4700 ± 300 RCYBP (#ISGS-452; Liu et al. Reference Liu, Riley and Coleman1986:106). Although the large error range results in a wide spread after calibration, it falls within Phase 2 and predates the Phase 3 occupation of Structure 7. Figure 8 shows the chronological relationships between Structures 1, 7, and S-CHM-1/F306.

Figure 8. Calibrated dates (calBC) for five radiocarbon assays derived from Structures 1, 7, and S-CHM-1/Feature 306 at Real Alto. Calibration was carried out using OxCal v4.3.2 software (Bronk Ramsey Reference Bronk Ramsey2017), and the r.5 IntCal13 atmospheric calibration curve was developed by Reimer and colleagues (Reference Reimer, Bard, Bayliss, Warren Beck, Blackwell, Ramsey, Buck, Cheng, Lawrence Edwards, Friedrich, Grootes, Guilderson, Haflidason, Hajdas, Hatté, Heaton, Hoffmann, Hogg, Hughen, Felix Kaiser, Kromer, Manning, Niu, Reimer, Richards, Marian Scott, Southon, Staff, Turney and van der Plicht2013). The two Geochron Laboratory (GX) assays were previously published in Zeidler (Reference Zeidler1984), and the three Illinois State Geological Survey (ISGS) assays were previously published in Liu and colleagues (Reference Liu, Riley and Coleman1986:106–107), with commentary by Donald W. Lathrap (see also Marcos Reference Marcos1988a, Reference Marcos1988b).
Methods and Approaches
Methods for sampling stone tools for microfossil residues are described in Pearsall and colleagues (Reference Pearsall2004); see Supplemental Text 1 for a summary and a description of dental calculus sampling. Phytolith scanning focused on economic taxa. Identifications were based on the University of Missouri (MU) Phytolith database (http://phytolith.missouri.edu; Pearsall Reference Pearsall2000) and comparative collection. The following economic taxa were identified by phytoliths: maize, yuca, arrowroot, llerén, and members of the genera, family, and order of these species; canna; squash and squash family and tribe; and bean family (see Table 2 for their scientific names). All Marantaceae and Zingiberales phytoliths were considered to represent economic taxa because members of these groups do not occur naturally in the xerophytic zone (see Supplemental Text 1 for further discussion). Silicified transport elements were classified as fruity or rooty following criteria established by Chandler-Ezell and others (Reference Chandler-Ezell, Pearsall and Zeidler2006), rooty parenchyma was also tallied, and the charcoal particles noted.
Table 2. Ubiquity (Percentage Presence) by Context of Microfossil Remains from Stone Tools, Sediment Samples, and Dental Calculus.

Notes: S = starch; P = phytoliths; CH = chipped; CO = cobble; G = ground stone.
All starch grains were identified to the extent possible using the MU comparative collection. The following plants were identified by starch: maize, yuca, possible yam, arrowroot, canna, chili pepper, and possible heat-altered chili pepper. Most samples were also examined for the presence of gelatinized starch.
Different pathways can lead to the deposition of phytoliths, starch, and charcoal on tools and in calculus. These pathways were investigated by Pearsall and colleagues (Reference Pearsall2004) in the context of maize occurrence in Structure 20. That study concluded that kernel grinding was the primary source of maize starch on tools, with fewer maize cob phytoliths originating from grinding than from sediments adhering to tools that contained phytoliths from cob decay. Based on comparisons to control samples, particulate charcoal was a component of in situ microfossil deposition on tool surfaces (see Supplemental Text 1 for further discussion). In the case of Structures 1, 7, S-CHM-1, and 10, field washing of artifacts removed most loose floor sediments, potentially reducing phytolith contributions from that source.
We compared occurrences of economic plants among sampled contexts using percentage presence (ubiquity; see Table 2), with separate calculations for different specificities of identifications (e.g., separately for arrowroot, Maranta, likely Maranta, and Marantaceae). For ease of comparisons, we calculated the ubiquity of all Marantaceae, combined (see Supplemental Tables 2–6 for sample-by-sample phytolith and starch tallies and Supplemental Figures 1–6 for microfossil images [Pearsall Reference Pearsall2019:doi:10.6067/XCV8448921]).
Results
Domestic Structures
Phytoliths of economic plants were recovered from all 15 tools from Structure 1; 13 (87%) had economic starch.Footnote 1 Maize phytoliths were absent, but maize starch was found on 80% of the tools and occurred in relatively large quantities on two (13%; see Table 2 and Supplemental Table 8). In terms of root-tubers, one tool (7%) had yuca phytoliths, and another one (7%) had Maranta and likely Maranta, llerén, Calathea, and Zingiberales. Two other tools (13%) preserved Calathea phytoliths, and 40% preserved Marantaceae. Overall, 47% of the tools processed some kind of Marantaceae. Three (20%) had Canna phytoliths. Most tools (80%) preserved generalized rooty tissues. In terms of fruits, squash phytoliths occurred on two tools (13%) but were not large enough for us to confirm that they were domesticated. Five tools (33%) preserved evidence for chili or heat-altered chili starch. Four of these (27%) also had maize starch. Phytoliths produced by Fabaceae fruits occurred on 60% of tools, and 73% had generalized fruity silicified tissues. Gelatinized starch occurred on all tools; 53% had charcoal present. Two tools (13%) had notably diverse residues.
Phytoliths from economic plants were recovered from all six tools from Structure 10, of which five (83%) had economic starch. Maize, gelatinized starch, Fabaceae, and generalized rooty tissues occurred on all tools. Charcoal and fruity tissues were present on 50% of tools and chili starch on 33%. Yuca, possible yam, Marantaceae, and Zingiberales each occurred on a single tool (17%). One tool (17%) stood out in having abundant maize starch and diverse residues.
Maize starch was found on all 17 tools from Structure 20; maize phytoliths occurred on 59%. Evidence for root-tuber use included arrowroot starch or phytoliths on 41% of tools and likely Maranta phytoliths on one tool (6%),; Calathea on 59% of tools and likely Calathea on 35%, Marantaceae on 24%, Zingiberales on 65%, Canna phytoliths or starch on 71% of tools, possible yam starch on two tools (12%), and yuca phytoliths or starch on 35%. In addition, rooty tissues occurred on 82% of the tools. Overall, 46% of tools preserved evidence of some kind of Marantaceae. In terms of fruits, chili or heat-altered chili starch occurred on two tools (12%), Cucurbita phytoliths on one (6%), Cucurbitaceae on 29%, Fabaceae on one (6%), and fruity transport elements on 47%. Gelatinized starch occurred on all but one tool (94%) and charcoal on every one. Several tools preserved abundant maize and diverse residues.
Floor sediment samples from Structure 20 contained phytoliths of maize, yuca, likely arrowroot, Calathea, Marantaceae, Zingiberales, and Canna. No starch grains were observed. Maize phytoliths were commonly recovered (from 77% of samples), as was evidence of some kind of Marantaceae (88%).
Ceremonial Contexts
Microfossils of economic plants were recovered from all seven tools from Structure 7. Maize was present on every tool; maize starch was abundant on two (29%; see Table 2 and Supplemental Table 8). In terms of root-tubers, rooty tissues were present on all tools, yuca on 43%, Calathea and likely Calathea on 29%, and arrowroot, Canna, and possible yam on one tool (14%) each. Overall, 29% of tools preserved some kind of Marantaceae. In terms of fruits, Cucurbita, Cucurbitaceae, and Cucurbitae occurred on one tool (14%) each. Fruity transport tissues were present on 57% of tools. Gelatinized starch was ubiquitous. Although maize co-occurred with rooty indicators on all tools, no tools had notably diverse residues.
Tools from Structure S-CHM-1 preserved evidence of maize (80% of tools), yuca (20%), Calathea and likely Calathea (40% combined), chili (60%), squash (20%), and Fabaceae (20%). Evidence for some kind of Marantaceae occurred on 60% of tools; rooty tissues were ubiquitous, as was gelatinized starch. No tools had notably diverse residues.
Dental Calculus
Calculus from all dentitions preserved gelatinized starch, which is evidence of the consumption of cooked plant foods (see Table 2). Silicified transport or parenchyma tissues from root-tuber foods and fruits were represented in 48% and 50% of individuals, respectively. Such tough, stringy tissues would likely become stuck in teeth or driven into the gum line. Indicators of maize, yuca, possible yam, Calathea or Maranta, Canna, domesticated squash, Fabaceae, Marantaceae, and Zingiberales were identified in the calculus of a few individuals, but identifiable microfossils of economics were sparse.
Silicified tissues from edible portions of foods (e.g., rooty and fruity transport tissues, Fabaceae seed parenchyma and fibers, Calathea/Maranta rhizome spindles, and Zingiberales spheres) can potentially be preserved in cooked and raw foods and then become incorporated into dental calculus. By contrast, identifiable starch is less likely to be preserved in cooked foods, although unaltered granules are sometimes present, such as in toasted maize (Chandler-Ezell et al. Reference Chandler-Ezell, Pearsall and Zeidler2006). Silicified tissues from inedible portions of foods (e.g., maize crosses and rondels, yuca secretory bodies, and Cucurbita and Canna spheres) could become incorporated into cooked foods by cooking unpeeled roots or fruits or maize on the ear. It is unlikely that phytoliths from inedible portions of foods became deposited on teeth after burial: comparing the phytolith collection from Burial 84 (FS#2122) to a sediment sample that accompanied the remains (FS#2012) shows virtually no overlap (Pearsall Reference Pearsall2019:Supplemental Table 6 [doi:10.6067/XCV8448933]).
Discussion
Insights into Foodways from Domestic Structures
The initial study of Structure 20 tools demonstrated that maize was commonly processed and that tools were used for a variety of foods in addition to maize, including root-tubers and fruits. The common occurrence of gelatinized starch and charcoal fragments indicated that cooked foods, as well as raw ingredients, came into contact with tools. Phytoliths from inedible portions of foods (e.g., maize cobs, squash rinds, and yuca peels) also became deposited on tools, likely via two pathways: from decayed debris present in floor sediments and through food processing. Many tools were multipurpose and preserved evidence of a diverse cuisine.
New microfossil data from Structure 1 and Structure 10 presented here provide additional insights into domestic foodways at Real Alto. The most striking finding is the strong similarity among households in the plant foods used. Maize, yuca, Marantaceae, Zingiberales, chili, and Fabaceae, in addition to generalized rooty and fruity tissues and gelatinized starch, were identified in all households. In addition, arrowroot or Maranta, llerén or Calathea, and squash were documented in Structures 1 and 20. Phase 3 households had access to the same basic foodstuffs and a nutritionally rich diet. There is no evidence for specialization in food production among households; the less diverse residues from Structure 10 may result from the smaller number of tools studied and the lower microfossil counts.
Domestic households also show similar patterning in the ubiquity of different foods. Maize, rooty tissues, and gelatinized starch were each ubiquitous or nearly so. Maize and root-tubers were commonly prepared in households, using the same multipurpose tools, and residues from food preparation were preserved. The presence of gelatinized starch and charcoal fragments indicates that cooked foods were commonly given additional processing using the same tools. Fabaceae was commonly processed in Structure 1 and Structure 10, less so in Structure 20. Fruity tissues were present on 47%–73% of the tools in households.
Residues of chili, yuca, and Marantaceae occurred on fewer tools in each house. Chili starch, for example, was present on 6%–33% of tools by household, yuca starch or phytoliths on 7%–35%. This indicates less common use or less frequent deposition or preservation of residues of these foodstuffs. For chili, starch is not abundant in domesticated peppers (Perry et al. Reference Perry, Dickau, Zarrillo, Holst, Pearsall, Piperno, Berman, Cooke, Rademaker, Ranere, Scott Raymond, Sandweiss, Scaramelli, Tarble and Zeidler2007) and so is likely underrepresented on tools. Processing raw yuca should leave abundant, identifiable starch on tools, whereas processing cooked yuca should leave little recognizable starch (Chandler-Ezell et al. Reference Chandler-Ezell, Pearsall and Zeidler2006). Yuca phytoliths are not abundant in roots and are likely underrepresented. The low percentage presence of yuca starch on tools suggests that raw yuca was not commonly processed; as shown by the presence of yuca phytoliths, cooked yuca likely contributed to the gelatinized starch and rooty tissues found on tools.
Arrowroot occurred on 41% of tools from Structure 20 and llerén on 7% from Structure 1. Considering all Marantaceae combined suggests the equally common use of this root-tuber group in these households (46%–47% of tools) and its presence in Structure 10. Family-level diagnostic phytoliths (nodular spheres) are commonly produced in stem, root, tuber, and rhizome tissues (Chandler-Ezell et al. Reference Chandler-Ezell, Pearsall and Zeidler2006) and are likely to become deposited on tools if Marantaceae were processed. Nodular spheres occurred on tools in all households (Pearsall Reference Pearsall2019:Supplemental Table 2 [doi:10.6067/XCV8448929], Supplemental Table 3 [doi:10.6067/XCV8448930], Supplemental Table 4 [doi:10.6067/XCV8448931]). Genus- and species-level diagnostics are less commonly produced and are likely underrepresented. The large starch grains produced by arrowroot and llerén also may be underrepresented; arrowroot starch was found only on the unwashed tools of Structure 20.
In summary, microfossils from stone tools and floor sediments from domestic structures provide insights into the foodways of Phase 3 Real Alto. The plant component of the cuisine was broad, incorporating maize and root-tubers (yuca, yam, arrowroot, llerén, canna) equally commonly. Fabaceae provided protein (note that jackbean [Canavalia] macroremains were identified in earlier contexts [Damp et al. Reference Damp, Pearsall and Kaplan1981]); squash, chili, and potentially other fruits and seeds contributed to food diversity. There were striking similarities among households in the combinations of foods processed on tools, as well as how commonly different foods were prepared, suggesting an overall equality in access to basic foodstuffs.
We have inferred that yuca was roasted and pounded on tools and that chili (raw and roasted) and maize were ground. What foods were prepared from maize flour? Valdivia pottery does not include griddles for flour toasting. Early Valdivia ceramic collections are dominated by bowls and jars with tall or short necks (Lathrap et al. Reference Lathrap, Collier and Chandra1975). Elaboration of vessel forms characterizes Middle and Late Valdivia. Bowls were used to serve food and drink; jars show charring/soot deposits from being used for cooking. Microfossil residues on Early Valdivia cooking jar sherds from Loma Alta show that ground maize was cooked in water with chili, yuca, arrowroot, and jack bean, making a nutritionally balanced soup or stew (Zarrillo et al. Reference Zarrillo, Pearsall, Scott Raymond, Tisdale and Quon2008). Perhaps jars were also used to steam maize dough wrapped in leaves.
Some ceramic vessels may have been used to ferment maize flour or yuca roots for beer (Marcos Reference Marcos1978, Reference Marcos1988a, Reference Marcos1988b; Zeidler Reference Zeidler1984). One unique form was recovered from Structure 1: it is a large, spheroidal- to ovoidal-shaped jar with a high shoulder, restricted opening, and cambered rim that aided binding of a cloth or leaf to cover its opening (Zeidler Reference Zeidler1984:Figures 15c and 59). Other forms potentially used for fermentation are the wide-mouth, bell-shaped jar and the folded rim (“pie-crust”) jar commonly decorated with continuous maize kernel impressions (Zeidler Reference Zeidler1984:Figures 57 and 58). Sherds of folded rim jars were broadly dispersed across Structure 1, whereas bell-shaped jars were clustered on its west side (Zeidler Reference Zeidler1984).
Domestic and Ceremonial Foodways at Real Alto
Microfossils from tools from Charnel House Mound structures reveal striking similarities between plant foods processed in ceremonial and domestic contexts. These similarities extend to the foods present and their relative rankings as measured by percentage presence. Maize and root-tubers were ubiquitous, or nearly so, in ceremonial contexts and were prepared using the same tools. Both raw and cooked foods contacted tools, but no tools from Structures 7 and S-CHM-1 exhibited diverse residues as did some tools from domestic structures. In addition, fewer microfossils of economic plants were recovered per tool from ceremonial structures than from two of three domestic houses. These differences suggest that the use-lives of Charnel House Mound tools were shorter and more focused.
A variety of jar forms occurred in the ceremonial precinct of Real Alto (Marcos Reference Marcos1978, Reference Marcos1988a, Reference Marcos1988b), including large, restricted-neck jars suitable for containing maize or yuca beer. Such jars were found in refuse pits within ceremonial structures in association with broken drinking bowls. Grinding stones occurred in floor deposits and in burial pits. Maize phytoliths were present in Structure 7 floor sediments (Pearsall Reference Pearsall2019:Supplemental Table 7 [doi:10.6067/XCV8448934]). The remains of feasting activities (drinking vessels, food remains) and feasting preparation (microfossil residues on grinding stones and in floor sediments, brewing vessels) were interred together on the Charnel House Mound: thus the actions of creating food and drink—not just its consumption—were ceremonial. If beer was indeed produced at Real Alto, microfossil residues and distributions of brewing jars indicate that its production took place in both domestic and funerary contexts. Similar evidence supports feasting preparation and consumption within Fiesta Mound structures, which were used through the Late Valdivia (Marcos Reference Marcos1978, Reference Marcos1988a, Reference Marcos1988b). Five refuse pits in Fiesta Mound structures dating to the Late Valdivia preserved maize leaf/husk phytoliths in association with drinking bowls (Pearsall Reference Pearsall1979).
In summary, we found no evidence for differentiation or specialization in plant-based foods processed in Phase 3 households for domestic use or ceremonial occasions. Food preparation, presentation, consumption, and disposal took place in both domestic and ceremonial contexts; the same foods nourished the Middle Valdivia population daily and their ancestors in funerary ritual. Households had equal access to annual crops, such as maize, and longer-growing root-tubers. This lack of differentiation in foodways parallels patterning in artifact collections, which have shown no evidence for specialized craft production or significant accumulation of prestige goods during Phase 3—although the presence of Spondylus shell beads, ceramic figurines, and a miniature ceramic “shaman's stool” interred with infant and juvenile burials in Structure 7 is noteworthy (Marcos Reference Marcos1978; Zeidler and McEwan Reference Zeidler, McEwan, Cockrell, Hoopes and McEwan2020). It is likely that Valdivians traded the sacred marine shell dyad of Spondylus and Strombus to inland localities during the Middle Valdivia, but long-distance exchange of worked Spondylus artifacts probably dated to the Late or Terminal Valdivia (Zeidler Reference Zeidler1991). Given that the presence of two modal sizes of houses suggests some degree of economic inequality—some households or descent groups had access to more labor (Zeidler Reference Zeidler1984, Reference Zeidler1991)—we next consider how access to labor affected Valdivia agriculture and the ability of groups to produce more or different kinds of plant products.
Real Alto Agriculture: Contextualizing New Results
Root-tuber crops and maize were central components of Middle Valdivia foodways. Given the complexities of phytolith production and starch survival, it is unlikely we can establish an absolute ranking of these foods in their diet. Maize, yuca, llerén, arrowroot, canna, and yam are all carbohydrates that can be made into an array of foods; all have been grown by tropical forest agriculturalists. It is beyond the scope of this discussion to review cultivation practices for these crops in detail. We therefore focus on one intriguing issue concerning Real Alto agriculture arising from our findings: How did cultivation of longer-season crops like yuca, arrowroot, and llerén (in addition to perennial cotton) differ from that of maize and other short-season annuals?
Real Alto is located above the Río Verde, which joins the Río Zapotal at the Chanduy estuary (see Figure 1). Pedestrian surveys of these river floodplains and adjacent uplands identified 12 Valdivia sites (Damp Reference Damp1979, Reference Damp1984b; Zeidler Reference Zeidler and Marcos1986). A more intensive survey was done within 5 km of Real Alto, with less systematic coverage farther away. Only Real Alto and Cerro Centinela, located 26 km inland, were occupied in the Early and Middle Valdivia and afterward. All other Valdivia sites dated to the Late or Terminal Valdivia. Systematic surveying beyond those zones by Kreid (Reference Kreid1986; Kreid et al. Reference Kreid, Garcia and Vedova1985) identified numerous hamlets dating to the Terminal Valdivia (Phase 8). Thus, as inhabitants of the only settlement in the Chanduy area dating to the Early and Middle Valdivia, the population of Real Alto had exclusive access to alluvial lands along the middle and lower reaches of both rivers and at their confluence.
Naturally watered alluvial terraces would have been prime agricultural lands. Rainfall is seasonal and limited on the Santa Elena Peninsula, falling from January through April; it is highest in the Chongón-Colonche mountains (400–700 mm/yr) and decreases to the west/southwest to 100–200 mm/yr (Hydrotechnics 1974). The drainage basins of the Río Verde and Río Zapotal fall within the interior lowlands of the peninsula (see Figure 1); rivers that arise there, rather than in the mountains, are characterized by seasonal flows. Today, annuals like maize are planted along streams with the first rains and harvested as rainfall tapers off. As discussed earlier, indirect evidence that Real Alto agriculture was also oriented toward naturally watered alluvium included the site location and the absence of fuel wood cut from upland forests.
Direct evidence that crops were grown in alluvial settings came from core Ch-045 in the mangrove of the Chanduy estuary (Pearsall et al. Reference Pearsall, Duncan, Jones, Freidel, Veintimilla and Neff2016). Pollen was well preserved in strata radiocarbon dated 3236 BC (base of the core) to 502 BC (275 cm below the surface). The locality was then a freshwater swamp. Maize pollen was abundant at the base of the core and present throughout the sequence in quantities that indicated fields were located nearby. Other herbs and cultigens identified were Canna-type, Manihot (yuca genus), and Malvaceae (cf. Gossypium, cotton genus). High percentages of pollen from weedy plants indicated intensive human activity near the locality. Phytolith concentrations were low, but weedy taxa and economics, including palm, Marantaceae, and Canna, were identified. Core Ch-045 documented that farmers planted maize and other crops near the Chanduy estuary during the Early Valdivia, when Real Alto was a village; they continued to do so as it became a town and for long afterward. Alluvium-based cropping was a reliable agricultural system in this dry, seasonal environment.
How did farmers leverage 100–200 mm of rain falling over four months to grow yuca, arrowroot, llerén, and perennial cotton, which have longer growing seasons and higher water requirements? For example, arrowroot requires 10–11 months to mature and rainfall of 1,500–2,000 mm/year; llerén, 10–18 months and rainfall of 640–2,820 mm/year (see Pearsall [Reference Pearsall and Minnis2014] for Marantaceae overview and primary sources). We suggest three possibilities: developing shorter-season crop varieties, extending the growing season by watering, and growing long-season crops over two rainy seasons.
Shorter-season varieties of yuca are grown today in coastal Ecuador. In the Jama Valley, for example, some farmers grow three-month yuca intercropped with maize (Pearsall Reference Pearsall2004). There is no mention of shorter-season varieties of arrowroot and llerén in the limited literature on these crops (Pearsall Reference Pearsall and Minnis2014). There are 64 Calathea species and 4 Maranta species in Ecuador (Jørgensen and León-Yánz Reference Jørgensen and León-Yánez1999). Perhaps Valdivia farmers developed now extinct short-season cultivars from this diverse group.
Although the rainy season is only four months long on the peninsula and stream flow is limited to that season, woody plants thrive along streams and springs by using groundwater. Perennial cotton could have been grown in these settings by Valdivia farmers. Long-season crops could also have been irrigated. Because there is no evidence for canal irrigation, Pearsall (Reference Pearsall1979) proposed pot irrigation from wells dug into dry streambeds as a way of extending the growing season.
Numerous earthen water-catchment features (albarradas) occur on the peninsula, some dated as early as the Terminal Valdivia and Chorrera (Marcos Reference Marcos2004; Marcos and Tobar Reference Marcos, Tobar and Marcos2004; Stothert Reference Stothert1995). Coring in La Tarea (LT-041), an albarrada east of Chanduy, recovered maize phytoliths in the upper 2 m (Pearsall et al. Reference Pearsall, Duncan, Jones, Freidel, Veintimilla and Neff2016). Marantaceae and Canna were also present in fewer samples. Unfortunately, all radiocarbon dates were stratigraphically below these microfossils. Although LT-041 maize could not be dated, its presence further documents the strong association of maize and albarradas observed by Veintimilla (Reference Veintimilla and Marcos2004), who identified maize phytoliths in all albarradas excavated by Marcos and colleagues. Canna, squash, and bean phytoliths occurred infrequently; Marantaceae was not present in those albarradas.
Perhaps Real Alto farmers grew long-season root crops over two rainy seasons. Crops would be planted at the beginning of the rains, go dormant during the dry season, and then be harvested the next rainy season. In the Jama Valley yuca is routinely left in the ground until needed, sometimes for several years (Pearsall Reference Pearsall2004). Whether arrowroot and llerén can be grown this way is unclear. Detailed accounts of traditional cultivation practices are lacking for these crops; most information comes from small-scale commercial production (Pearsall Reference Pearsall and Minnis2014). Arrowroot is propagated from rhizome tips, and plants readily sprout the following rainy season from pieces remaining in the soil. This suggests that unharvested plants allowed to go dormant could regrow and develop starchy rhizomes. Llerén is also propagated from rhizome segments, but the utilized portion are tubers that form as rains decline and growth stops. What controls tuber formation is unknown.
In summary, Real Alto farmers had exclusive access to alluvial lands in the Chanduy region. Rainfall was sufficient to grow maize and other short-season crops—or three-month yuca—seasonally on alluvial terraces. Different cultivation practices were likely required for arrowroot and llerén, given that both are grown today under higher water regimes and longer growing seasons. In the absence of short-season varieties, hand watering could have extended the growing season, or crops may have been grown over two rainy seasons. Either approach would require increased labor—in terms of time spent watering or planting fields for every-other-year production—and multiyear access to plots and plantings. Because agriculture was rain fed, with prime lands spread out along streams, there was no point of control of agricultural production. Producing a surplus (e.g., maize and yuca for beer/feasting foods) would be a matter of having the necessary labor in a household or descent group to put more fields into production. As Zeidler (Reference Zeidler1991:257) noted,
The tendency toward larger household size at Real Alto could have come about as a result of complex scheduling requirements in Valdivia subsistence strategies, such that scheduling incompatibilities made it advantageous to maintain larger households (or a greater number of productive members in the household) . . . so as to effectively reduce the “dependency ratio” of producers to non-productive consumers and, thus, substantially increase the productive capacity of the domestic group, i.e., its allocative efficiency.
Conclusions
From the earliest research at Real Alto, Donald Lathrap was convinced that Valdivia culture was based on maize agriculture, was tropical forest in origin, and represented emergent social and political complexity. Lathrap based his conclusions on site location, size, and inferences on community structure drawn from tropical forest societies. Others subsequently contributed insights into lifeways at Real Alto from their study of domestic structures, mound building, pottery, regional settlement patterns, and botanical, faunal, and human skeletal remains.
This research transformed our understanding of Valdivia culture. Questions, however, still remained about the nature of food production at Real Alto and the role of agriculture—and of maize—in the emergence of social complexity during the Early Formative. To address these issues we undertook a new program of research that incorporated environmental coring and analysis of microfossil residues from previously excavated stone tools and human remains from Real Alto.
Lathrap was correct: Real Alto was an agricultural community whose inhabitants also fished, hunted, and gathered. The suite of crops present by the Middle Valdivia—maize, root-tubers (yuca, arrowroot, llerén, canna, and yam), jackbean, squash, gourd, chili pepper, and cotton—represented tropical forest agriculture. Residues of maize, chili, and root-tubers were found on the same processing tools, representing a truly mixed diet.
There were not different ceremonial and everyday foods at Real Alto: the actions of food (and perhaps beer) preparation, presentation, consumption, and disposal were both domestic and ceremonial, depending on the cultural context. Domestic households processed a similar array of raw and cooked plant foods, and rankings of foods by ubiquity within households were similar. Maize was ubiquitous or nearly so and occurred with indicators of root-tuber use. Residues of fruits and seeds were present on fewer tools. Gelatinized starch and charcoal were commonly recovered, indicating the processing of cooked foods. Dental calculus residues confirmed the common consumption of cooked foods, fruits, and root-tubers. The same plants were present, and in similar quantities, on tools from Charnel House Mound structures, and both raw and cooked foods were processed there as well. There were no tools notable for high food diversity in ceremonial contexts, indicating shorter, more focused use-lives for tools used there.
Real Alto farmers had exclusive access to alluvial lands in the Chanduy region and grew maize and other crops there. Cultivating crops with different water and growing season requirements necessitated diverse practices, potentially including developing short-season varieties, hand watering, and growing crops over two rainy seasons. The latter two approaches would have required increased labor inputs and multiyear access to plantings. The transformation of Real Alto from village to town during the Middle Valdivia represented significant changes in the social relationships of a growing population, reflected in settlement layout, domestic structures, public buildings, mortuary practices, and artifacts (Clark et al. Reference Clark, Gibson, Zeidler, Bandy and Fox2010; Zeidler and McEwan Reference Zeidler, McEwan, Cockrell, Hoopes and McEwan2020). The small Early Valdivia plaza was transformed into a ceremonial precinct with two platform mounds, one the site of feasting and the other of interment of selected individuals. Individual small houses were replaced by tight clusters of extended family dwellings of two modal sizes. Although this size difference suggests economic inequality, significant accumulations of prestige goods are lacking. Nevertheless, larger domestic groups would have had a considerable competitive advantage over smaller ones in terms of intensifying production (Zeidler Reference Zeidler1991). Access to more labor—to grow more maize and to maintain long-season plantings—was likely a key element supporting status differences.
Acknowledgments
Support was provided by a University of Missouri Research Board grant to Pearsall and a Harvard University Dumbarton Oaks research fellowship to Zeidler. Thomas Emerson helped arrange the loan of Real Alto stone tools curated at the University of Illinois (UI), Urbana. Linda Klepinger facilitated access to human skeletal remains curated at UI; Jorge Marcos, who discovered Real Alto and excavated the ceremonial structures, and Cesar Veintimilla facilitated access to human skeletal remains curated at ESPOL, Guayaquil. Figures were composed in final form by Howard Wilson. Permits were not required for this work. No financial conflicts of interest exist.
Data Availability Statement
Data in digital form are in Supplementary Tables 2–6; images are in Supplementary Figures 1–6 (doi:10.6067/XCV8448921). Lab forms and microscope slides are curated at the Museum Support Center of the Museum of Anthropology, University of Missouri, Columbia.
Supplemental Materials
Supplemental Figures 1–6 and Supplemental Tables 1–7 are available at The Digital Archaeological Record (tDAR id: 44892), doi:10.6067/XCV8448921. The rest of the supplemental files are available at https://doi.org/10.1017/laq.2019.96.
Supplemental Text 1. Methods and Approaches.
Supplementary Figure 1. Structure 1 images.
Supplementary Figure 2. Structure 10 images.
Supplementary Figure 3. Structure 20 images tools.
Supplementary Figure 4. Structure 20 images sediments.
Supplementary Figure 5. Charnal House Mound images.
Supplementary Figure 6. Dental Calculus images.
Supplementary Table 1. Pearsall 1979 wood data.
Supplementary Table 2. Structure 1 microfossil data.
Supplementary Table 3. Structure 10 microfossil data.
Supplementary Table 4. Structure 20 microfossil data.
Supplementary Table 5. Charnal House Mound microfossil data.
Supplementary Table 6. Dental calculus microfossil data.
Supplementary Table 7. Pearsall 1979 phytolith data.
Supplemental Table 8. Microfossil Occurrences on Selected Tools.












