



Introduction
Outcrops of Paleozoic rocks from the Santiago Ixtaltepec-Nochixtlán region, in Oaxaca State, contain one of the most diverse and abundant marine faunas from the Carboniferous of Mexico. The sequence was originally divided into two formations, the Santiago formation, an informal unit by homonymy that was considered as Mississippian and the Ixtaltepec Formation, thought to be Late Mississippian and Pennsylvanian in age (Pantoja-Alor, Reference Pantoja-Alor1970; Torres-Martínez and Sour-Tovar, Reference Torres-Martínez and Sour-Tovar2012). Several components of the invertebrate fauna from Ixtaltepec Formation have been published, including Pennsylvanian bivalves (Quiroz-Barroso, Reference Quiroz-Barroso1995; Quiroz-Barroso and Perrilliat, Reference Quiroz-Barroso and Perrilliat1997, Reference Quiroz-Barroso and Perrilliat1998), trilobites (Morón-Ríos and Perrilliat, Reference Morón-Ríos and Perrilliat1988), bryozoans (González-Mora and Sour-Tovar, Reference González-Mora and Sour-Tovar2014), and Mississippian and Pennsylvanian crinoids (Villanueva-Olea et al., Reference Villanueva-Olea, Castillo-Espinoza, Sour-Tovar, Quiroz-Barroso and Buitrón-Sánchez2011; Villanueva-Olea and Sour-Tovar, Reference Villanueva-Olea and Sour-Tovar2015).
In these Carboniferous strata, brachiopods are the most abundant group. Lingulides (Torres-Martínez and Sour-Tovar, Reference Torres-Martínez and Sour-Tovarin press), strophomenides (Sour-Tovar and Quiroz-Barroso, Reference Sour-Tovar and Quiroz-Barroso1989; Sour-Tovar, Reference Sour-Tovar1994), chonetidines (Sour-Tovar and Martínez-Chacón, Reference Sour-Tovar and Martínez-Chacón2004), productidines (Torres-Martínez and Sour-Tovar, Reference Torres-Martínez and Sour-Tovar2012; Torres-Martínez, Reference Torres-Martínez2014), and neospiriferins (Torres-Martínez et al., Reference Torres-Martínez, Sour-Tovar and Pérez-Huerta2008) have been described. The order Productida is particularly well represented at all levels of the Ixtaltepec Formation and the productides are a very important component in all the communities that characterize different stratigraphic levels. In this context, the present work is focused on the description of the productoids of the Ixtaltepec Formation to analyze their distribution in the sedimentary sequence and to interpret their stratigraphic implications for the Carboniferous sequence.
Material and geological setting
Brachiopods described in this work were collected from the “Arroyo de las Pulgas” type section of Ixtaltepec Formation located between 17°32'–17°33'N and 97°06'–97°07'W, 500 m north of the town of Santiago Ixtaltepec (Fig. 1). The entire region belongs to the Municipality of Nochixtlán, Oaxaca State, and is notable because it contains one of the most complete Paleozoic sections in Mexico. The basement is the Oaxacan Complex, composed of Proterozoic pegmatites, schists, paragneiss and orthogneiss with ages of 900–1100 million years (Fries et al., Reference Fries, Schmitter, Damon and Livingstone1962; Solari et al., Reference Solari, Keppie, Ortega-Gutiérrez, Cameron, López and Hames2003). Overlying the basement rocks are the Tiñú Formation, a Cambrian-Ordovician marine succession of calcareous and argillaceous rocks with abundant trilobites and other invertebrate fossils (Robison and Pantoja-Alor, Reference Robison and Pantoja-Alor1968; Landing et al., Reference Landing, Westrop and Keppie2007, Reference Landing, English and Keppie2010). The Santiago Formation is 165 m thick and unconformably overlies the Tiñú Formation. It has been informally divided into two members. The lower member consists of 65 m of sandstone, limestone and calcareous sandstone whose age has been established as Osagean (Early Mississippian) constrained by brachiopods (Navarro-Santillán et al., Reference Navarro-Santillán, Sour-Tovar and Centeno-García2002) and rostroconchs (Quiroz-Barroso et al., Reference Quiroz-Barroso, Pojeta, Sour-Tovar and Morales-Soto2000). The upper member is composed of approximately 100 m of shale and fine-grained sandstone; the age of the middle part of this member has been established as Meramecian (Middle Mississippian) based on the presence of Goniatites crenistria, Beyrichoceras hornerae and Prolecanites americanus (Cephalopoda) (Castillo-Espinoza, Reference Castillo-Espinoza2013).

Figure 1 Geographic location of the Ixtaltepec Formation type section, where specimens of productoid brachiopods have been collected.
The Ixtaltepec Formation, with a thickness of 430 m at the Arroyo de las Pulgas type section conformably overlies the Santiago formation. The basal part includes 90 m of interbedded siltstone, sandstone, sandy-limestone and shale. The rest of the succession is composed mainly of shale and minor fine-grained sandstone. The Ixtaltepec Formation has been divided, from the base to the top, into eight fossiliferous units named API–1 to API–8, each one characterized by distinct fossil associations (Fig. 2). The brachiopods studied in the present work were collected in units API–1, 2, 3, 6, 7, and 8.

Figure 2 Stratigraphy of Paleozoic outcrops from Santiago Ixtaltepec area. The continuous lines on each taxa indicated stratigraphic levels of the Ixtaltepec Formation where the species of the superfamily Productoidea were found.
Unconformably overlaying the Ixtaltepec Formation is the Yododeñe Formation, a conglomerate with calcareous clasts whose age is estimated to be Permian to Jurassic. Throughout the region, the Paleozoic sequence is covered by Cretaceous marine rocks.
Discussion
The Ixtaltepec Formation was initially assigned to the Bashkirian-Moscovian (Early-Middle Pennsylvanian) by the presence of Anthracospirifer occiduus throughout the unit (Pantoja-Alor, Reference Pantoja-Alor1970); however, subsequent studies (e.g., Butts, Reference Butts2007) have expanded the stratigraphic range of that species downward, to the Viséan-Serpukhovian (=Chesterian). In previous work, some species of different groups referred to the Late Mississippian (Chesterian) have been reported from the Ixtaltepec Formation, such as the bivalves (Quiroz-Barroso and Perrilliat, Reference Quiroz-Barroso and Perrilliat1998) and crinoids (Villanueva-Olea et al., Reference Villanueva-Olea, Castillo-Espinoza, Sour-Tovar, Quiroz-Barroso and Buitrón-Sánchez2011; Villanueva-Olea and Sour-Tovar, Reference Villanueva-Olea and Sour-Tovar2015). Moreover, Orbiculoidea caneyana, Echinoconchella elegans, Marginovatia minor, and Ovatia muralis have been referred to the basal stratigraphic levels of the formation (Torres-Martínez and Sour-Tovar, Reference Torres-Martínez and Sour-Tovar2012; Torres-Martínez and Sour-Tovar, Reference Torres-Martínez and Sour-Tovarin press).
The presence in levels API–1 and API–2 of the characteristic Viséan-Serpukhovian taxa Semicostella sp., Keokukia? sp. and Inflatia inflata corroborate the Mississippian age of the base of the Ixtaltepec Formation. Buxtonia websteri a Moscovian taxon (Beus and Lane, Reference Beus and Lane1969) occurs in levels API–6, API–7 and API–8 and the Moscovian-Gzhelian taxon Reticulatia cf. R. huecoensis (King, Reference King1931; Muir-Wood and Cooper, Reference Muir-Wood and Cooper1960; Hoare, Reference Hoare1961; Sturgeon and Hoare, Reference Sturgeon and Hoare1968), occurs in level API–7. These ocurrences confirm the Pennsylvanian (Bashkirian-Moscovian) age for the upper levels of the formation, and are internally consistent with previous assessments based on the presence of Orbiculoidea missouriensis, Orbiculoidea capuliformis, Neochonetes granulifer, Echinaria knighti, Linoproductus prattenianus, Linoproductus platyumbonus, Marginovatia aureocollis, Marginovatia pumila, and Neospirifer dunbari (Sour-Tovar and Martínez-Chacón, Reference Sour-Tovar and Martínez-Chacón2004; Torres-Martínez et al., Reference Torres-Martínez, Sour-Tovar and Pérez-Huerta2008; Torres-Martínez and Sour-Tovar, Reference Torres-Martínez and Sour-Tovar2012; Torres-Martínez and Sour-Tovar, Reference Torres-Martínez and Sour-Tovarin press).
It must be emphasized that the presence of Inflatia and Flexaria in levels API–7 and API–8 from the Ixtaltepec Formation leaves the possibility that their stratigraphic range might extend to the Middle Pennsylvanian.
Paleobiogeographic implications
The existence of a strong affinity between the Carboniferous faunas of the Santiago Ixtaltepec area with the different faunas from the Mississippian or Pennsylvanian localities of the Midcontinent and western United States has been noted previously (Sour-Tovar, Reference Sour-Tovar1994; Quiroz-Barroso and Sour-Tovar, Reference Quiroz-Barroso and Sour-Tovar1996; Sour-Tovar et al., Reference Sour-Tovar, Quiroz-Barroso, Centeno-García, Navarro-Santillán and Ortiz-Lozano1997; Quiroz-Barroso and Perrilliat, Reference Quiroz-Barroso and Perrilliat1998; Sour-Tovar, Reference Sour-Tovar, Alvarez and Martínez-Chacón2005; Navarro-Santillán et al., Reference Navarro-Santillán, Sour-Tovar and Centeno-García2002; Torres-Martínez et al., Reference Torres-Martínez, Sour-Tovar and Pérez-Huerta2008; Torres-Martínez and Sour-Tovar, Reference Torres-Martínez and Sour-Tovar2012; Torres-Martínez and Sour-Tovar, Reference Torres-Martínez and Sour-Tovarin press). For example, of the previously brachiopods, O. caneyana, O. missouriensis, O. capuliformis, N. granulifer, I. inflata, R. huecoensis, B. websteri, E. knighti, L. prattenianus, L. platyumbonus, M. minor, M. aureocollis, M. pumila, O. muralis, and N. dunbari have been reported from Carboniferous strata in Missouri (Hoare, Reference Hoare1961); Ohio (Sturgeon and Hoare, Reference Sturgeon and Hoare1968); Nevada (Beus and Lane, Reference Beus and Lane1969); Oklahoma, Arkansas (Girty, Reference Girty1909; Gordon and Henry, Reference Gordon and Henry1990; Gordon et al., Reference Gordon, Henry and Treworgy1993); Nebraska, Kansas (Dunbar and Condra, Reference Dunbar and Condra1932; Spencer, Reference Spencer1967); Wyoming (Gordon, Reference Gordon1975); or Idaho (Butts, Reference Butts2007) and their presence in Oaxaca confirms the similarity between the fauna of brachiopods from Ixtaltepec with faunas referred to the Midcontinent. Other invertebrate taxa, including mollusks and crinoids from the Ixtaltepec Formation have similar pattern of paleogeographic distribution (Quiroz-Barroso and Perrilliat, Reference Quiroz-Barroso and Perrilliat1997, Reference Quiroz-Barroso and Perrilliat1998; Quiroz-Barroso et al., Reference Quiroz-Barroso, Pojeta, Sour-Tovar and Morales-Soto2000; Villanueva-Olea et al., Reference Villanueva-Olea, Castillo-Espinoza, Sour-Tovar, Quiroz-Barroso and Buitrón-Sánchez2011). This taxonomic distribution suggests that during the Carboniferous, the area of Nochixtlán was connected to an epicontinental sea that extended from the northern part of the Midcontinent to the current region of the southeast of Mexico.
Systematic paleontology
All studied specimens are deposited in the collections of the Museo de Paleontología, Facultad de Ciencias, Universidad Nacional Autónoma de México. Type and figured specimens are designated in the descriptions by the prefix FCMP. The classification and morphological terminology above the species level follow the descriptions of Muir-Wood and Cooper (Reference Muir-Wood and Cooper1960), Brunton et al., (Reference Brunton, Lazarev, Grant and Yu-Gan2000) and Brunton (Reference Brunton2007). The material is preserved as internal and external molds of both valves; some specimens have permineralized shells. In many cases, the internal molds of ventral valves are composite molds where it is just possible to see the external morphology.
Phylum Brachiopoda Duméril, Reference Duméril1805
Subphylum Rhynchonelliformea Williams, Carlson, Brunton, Holmer and Popov, Reference Williams, Carlson, Brunton, Holmer and Popov1996
Class Strophomenata Williams, Carlson, Brunton, Holmer and Popov, Reference Williams, Carlson, Brunton, Holmer and Popov1996
Order Productida Sarytcheva and Sokolskaya, Reference Sarytcheva and Sokolskaya1959
Suborder Productidina Waagen, Reference Waagen1883
Superfamily Productoidea Gray, Reference Gray1840
Family Productellidae Schuchert, Reference Schuchert1929
Subfamily Plicatiferinae Muir-Wood and Cooper, Reference Muir-Wood and Cooper1960
Tribe Semicostellini Nalivkin, Reference Nalivkin1979
Genus Semicostella Muir-Wood and Cooper, Reference Muir-Wood and Cooper1960
Type species
Avonia oklahomensis Snider, Reference Snider1915, from the Chester Group (Serpukhovian) of Oklahoma, USA.
Semicostella sp.
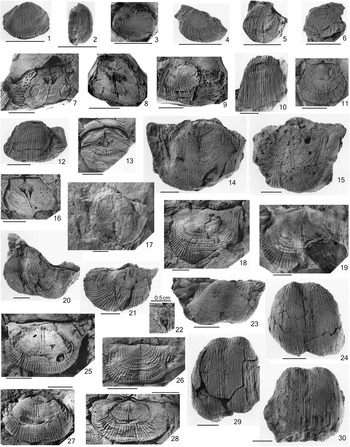
Figure 3 Semicostella sp., (1–3) FCMP 1013, ventral internal mold in ventral and lateral view, and cast; (4) FCMP 1014, ventral internal mold; (5) FCMP 1015, ventral internal mold; (6) FCMP 1016, ventral internal mold; Weberproductus donajiae n. gen. n. sp., (7–9) FCMP 1018 (holotype), dorsal internal mold with part of ventral internal and external molds (7), rubber cast of dorsal internal mold (8), and rubber cast of ventral internal mold (9); (10) FCMP 1023, ventral internal mold; (11) FCMP 1024, dorsal external mold; (12) FCMP 1017, ventral internal mold; (13) FCMP 1026, dorsal internal mold; (16) FCMP 1027, dorsal internal mold; Antiquatonia sp., (14) FCMP 1028, ventral internal mold; (15) FCMP 1029, ventral internal mold; (18, 19) FCMP 1030, dorsal external mold and rubber cast of dorsal internal mold; Keokukia? sp., (17) FCMP 1031, ventral valve; (20) FCMP 1032, ventral internal mold; (21) FCMP 1033, ventral internal mold; Dictyoclostus transversum n. sp., (22) FCMP 1043, x4.0, fragment of dorsal valve, interior; (23) FCMP 1035, ventral valve, posterior view; (25) FCMP 1034 (holotype), ventral internal mold; (26) FCMP 1044, dorsal internal mold; (27) ventral valve, rubber mold of FCMP 1039; (28) FCMP 1040, dorsal external mold; Inflatia inflata (McChesney, Reference McChesney1860), (24) FCMP 1045, ventral valve, anterior view; (29) FCMP 1046, ventral valve, anterior view; (30) FCMP 1047, ventral valve, anterior view. Scale bars represent 1.0 cm.
Occurrence
Ixtaltepec Formation, Arroyo las Pulgas type section, level API–2, Upper Mississippian.
Description
Small-sized productellid, subquadrate in outline. Ventral valve highly convex, sulcus absent. Short trail separated from visceral disc by a cincture. Umbo massive, incurved. Triangular ears, flattened. Greatest width at hingeline. Flanks steep, at 90° angle. Cardinal extremities angular. Ornamented by costae low, rounded, 9 to 11 per 5 mm at venter, beginning at 2 mm of the beak, well developed at anterior margin, costae initiating bifurcations on the middle of visceral disc, curved on flanks. Narrow concentric rugae on visceral disc. Spines (1) concentrically arranged on the trail, and (2) one or two spines near of hinge on the ears; spine bases scattered on visceral disc. Dorsal valves unknown.
Materials
Four ventral internal molds (FCMP 1013–1016) and a ventral external mold (FCMP 1013).
Remarks
The genus Semicostella was proposed by Muir-Wood and Cooper (Reference Muir-Wood and Cooper1960), as referenced to the presence of a trail usually separated from visceral disc by subparallel cincture, costae developed at anterior end of visceral disc, narrow rugae on visceral disc and scattered spines. Semicostella sp. presents these traits thus corroborating the generic assignment. The specimens is distinguished from Semicostella oklahomensis (Snider, Reference Snider1915), of Moorefield Limestone from Viséan-Serpukhovian of Oklahoma by its smaller size (S. oklahomensis=23 mm of length and 23 mm of width; Semicostella sp.=10.3 mm of length and 9.0 mm of width), lower convexity, slightly less transverse (length/width ratio of 1:1 in S. oklahomensis; length/width ratio of 1:0.8 in Semicostella sp.) and greater number of costae. The subparallel cincture of Semicostella sp. is observed somewhat slightly posterior in comparison with the cincture of S. oklahomensis (e.g., Muir-Wood and Cooper, Reference Muir-Wood and Cooper1960, pl. 62, fig. 6). This is the first occurrence of the genus in Mexico.
Family Productidae Gray, Reference Gray1840
Subfamily Productinae Gray, Reference Gray1840
Tribe Productini Gray, Reference Gray1840
Genus Weberproductus new genus
Type species
Weberproductus donajiae n. gen. n. sp. from the upper Ixtaltepec Formation (Bashkirian-Moscovian) of Oaxaca, Mexico.
Diagnosis
Ventral valve with or without broad and shallow sulcus. Surface ornamented by subequal costae, bifurcating at visceral disc, weak to absent near ears. Narrow rugae confined to umbonal region and ears. Scattered spines over valve, cluster of numerous spines on the umbonal slopes and near ears; rugae more pronounced at dorsal valve; single diaphragm around of the visceral disc. Cardinal process small, bilobate above and trilobate posteriorly; median septum stout posteriorly, narrowing suddenly between adductor muscle scar and extending across diaphragm; brachial ridges separated 2.5 mm of the anterior edge, crossed in the base by the diaphragm; cardinal ridges moderately strong, parallels to hingeline; posterior pair of adductor muscles dendritic and the anterior pair smooth; small endospines anterior of the visceral disc.
Etymology
In honor of Reinhard Weber Göbel, in recognition of the valuable information that contributed to the knowledge of Paleozoic in Mexico.
Occurrence
All specimens described are from Ixtaltepec Formation, Arroyo las Pulgas type section, level API–7, Middle Pennsylvanian.
Remarks
The material described is assigned to the tribe Productini based on characteristically fine costae; thin spines that are more numerous on the ears; presence of a diaphragm and adductor scars on a platform (Brunton et al., Reference Brunton, Lazarev, Grant and Yu-Gan2000). Ixtaltepec specimen have certain structural similarities with other taxa within the tribe; however, do not belong to any of the previously described genera. Weberproductus is similar to Productus Sowerby, 1814, by the medium size and the presence of a diaphragm restricted to one third of the anterior region; however Productus is dissimilar by their thinner costae, long and broad trail, dorsal valve geniculated, median septum does not cross the diaphragm, and lateral ridges diverging from the hingeline. Diaphragmus Girty, Reference Girty1910, differs from Weberproductus in the series of trails with the exterior the longer; the spines more numerous and grouped on the flanks; posterior reticulation more pronounced and dorsal disc flat. The new genus resembles Companteris Lazarev, Reference Lazarev1981 in the presence of a diaphragm with two trails and subcircular shape; however Companteris is dissimilar to Weberproductus in its smaller size; more convex ventral valve, fewer spines, similarly distributed on ears and flanks, and adductor scars overhang laterally. Dowhatania Waterhouse (in Waterhouse and Gupta), Reference Waterhouse and Gupta1979, differs from Weberproductus in the numerous spines on ears and flanks; and by the presence of more than one dorsal diaphragm. Lopasnia Ilkhovsky (in Lazarev), Reference Lazarev1990, differs from the new genus in its smaller size; transverse shape; very thick costae and cardinal process supported by adductor ridges. Marginirugus Sutton, Reference Sutton1938, is different from Weberproductus by its larger size, short trails, thinner costae, rugae posterolaterally, no reticulation, without scattered spines, and a row of stout spines near the hingeline.
Weberproductus resembles Carlinia Gordon, Reference Gordon1971, in the presence of a group of abundant spines on the ears, posterior reticulation almost absent and a wide area of diaphragms; however, the new genus is dissimilar to Carlinia being subcircular in outline, with costae of the same thickness; a larger number of spines on the ears and flanks of the umbo, a single diaphragm with two trails and anterior endospines in the dorsal valve.
According to Gordon (Reference Gordon1971), Carlinia has an elongated shape, costae of different thicknesses, fewer spines in the ears, a series of multiple trails, and no endospines.
Weberproductus donajiae new species
Diagnosis
Ventral valve strongly curved posteriorly, broad and shallow sulcus, surface ornamented by subequal costae; 40–42 total near anterior margin, averaging about 13 per 10 mm; bifurcation starting at visceral disc; costae weak to absent near ears; narrow rugae, confined to umbonal region and ears; scattered spines covering valve, with a cluster of 28 to 32 on the umbonal slopes and near of ears; rugae more notorious at dorsal valve, mainly in diaphragm area. Single concentric diaphragm; cardinal process short, bilobate; cardinal ridges parallel to hingeline; adductor muscle scars well differentiated, elevated on prominent platforms; small endospines.
Occurrence
Ixtaltepec Formation, Arroyo las Pulgas type section, level API–7, Middle Pennsylvanian.
Description
Shell moderate-sized for the tribe, concavo-convex, subcircular in outline with short trail. Ventral valve strongly curved posteriorly, decreasing gradually from umbo to anterior margin. Flanks steep. With a broad shallow sulcus. Narrow umbo, rounded, ending approximately 2.8 mm beyond hinge. Greatest width at the mid-length. Short ears, slightly convex. Ornamented by low costae, rounded, subequal, 40–42 near anterior margin, averaging about 13 per 10 mm, bifurcating on visceral disc, weak to absent on ears. Narrow rugae, inconspicuous, confined to umbonal region and ears. Spines on the costae (1) scattered on venter, anterior and lateral slopes, and (2) on umbonal slopes and ears in a group of 28 to 32 fine spines. Ventral interior not observed. Dorsal valve gently concave median-posteriorly. With shallow fold, corresponding to sulcus of ventral valve. One concentric diaphragm with two trails, the external trail is ornamented with thin radial costae. Concentric rugae, developed mainly in visceral disc. Dorsal interior with wide diaphragm area. The diaphragm is concentric around the visceral disc. Cardinal process short, bilobate internally and trilobate posteriorly. Median septum stout posteriorly, narrowing suddenly between adductor muscle scar and extending across diaphragm. Brachial ridges 2.5 mm apart of the anterior edge, crossed in the base by the diaphragm. Cardinal ridges moderately strong, run parallel to hingeline. Adductor muscle scars well differentiated, posterior pair dendritic and anterior pair smooth, elevated on prominent platforms. Small endospines in front of the visceral disc. Measurements in millimeters and indices in Table 1.
Table 1 Measurements of Weberproductus donajiae n. gen. n. sp.

LE, length; LA, length along arc; WH, width at hinge; GW, greatest width; units, millimeters; Costae, number of costae per 10 mm on the anterior margin; e, estimated, sample incomplete.
Etymology
The name refers to the legend of the Princess Donají, a story of great significance for the people of Oaxaca, where two ethnic cultures are linked by the relationship from the Zapotec Princess Donají and the Mixtec Prince Nucano.
Materials
A dorsal internal mold with part of ventral internal and external molds (FCMP 1018) (holotype), seven ventral internal molds (FCMP 1017, 1019–1023), and four dorsal internal molds (FCMP 1024–1027).
Remarks
Besides the previously described differences for the genus, Weberproductus donajiae differs from Carlinia species by several characteristics. Carlinia amsdeniana Gordon, Reference Gordon1971, is reported to the Amsden Formation from the Serpukhovian of Wyoming (Gordon, Reference Gordon1971, Reference Gordon1975) and is different to W. donajiae because it is larger, with fewer costae on the ventral valve and fewer wrinkles in the dorsal visceral disc. Carlinia phillipsi (Norwood and Pratten, Reference Norwood and Pratten1855), from the Serpukhovian of Chainman Shale and Manning Canyon Shale in Utah (Gordon, Reference Gordon1971) is similar in size, but differs from the specimens of Ixtaltepec in its more elongated shape, with thinner costae and a higher total number of costae in the valve. Carlinia diabolica Gordon, Reference Gordon1971, from the Serpukhovian of Chainman Shale and Manning Canyon Shale in Utah (Gordon, Reference Gordon1971) is different from W. donajiae in being larger, with longer ears not well differentiated and spines that have a specific arrangement. In particular, W. donajiae is easily differentiated by the presence of the 28 to 32 spines cluster located on the ears and the flanks of the umbo.
Tribe Retariini Muir-Wood and Cooper, Reference Muir-Wood and Cooper1960
Genus Antiquatonia Miloradovich, Reference Miloradovich1945
Type species
Productus antiquatus Sowerby, 1821, from Cloghran (Viséan) of Derbyshire, England.
Antiquatonia sp.
Occurrence
Ixtaltepec Formation, Arroyo las Pulgas type section, level API–3, Upper Mississippian.
Description
Medium-sized, concavo-convex productid. Subrectangular in outline, greatest width at hingeline. Large shells up to 45 mm in length and 55 mm in width. Ventral valve inflated, greatest convexity at venter. Flanks steep. Umbo moderately convex. Ears well developed, slightly arched, subquadrate to subtriangular, separated from umbonal slopes by a shallow concave flexure. Ornamented by numerous costae, entire, rounded, 8–10 occurring in space of 10 mm over anterior margin. Spines occurring (1) in curved row up flanks, complementary to internal lateral ridges, and (2) rarely scattered over the valve; the rows of spines near hinge or flanks are not visible. Rugae numerous on visceral disc, forming a reticulation with the costae. Dorsal valve concave with shallow fold. Similar ornament to ventral valve, without spines. Interior with small trilobate cardinal process, median lobe ventrally sulcate. Median septum extending to two-thirds length of valve. Muscle field small, adductor scars dendritic, elongate. Lateral ridges strong, diverging slightly from hinge in the lateral margins.
Materials
Two ventral internal molds (FCMP 1028, 1029) and a dorsal external and internal mold (FCMP 1030).
Remarks
Ixtaltepec specimens exhibit characteristics of the genus, such as shape, posterior reticulation, ventral spine ridge flanking ears with internal complimentary lateral ridges, ventral spines thick, adductor scars dendritic and enlarged (Muir-Wood and Cooper, Reference Muir-Wood and Cooper1960; Brunton et al., Reference Brunton, Lazarev, Grant and Yu-Gan2000). The row of spines at the base of flanks placed on a ridge was not observed but the previous morphological features confirm that the material of Oaxaca belongs to the genus Antiquatonia. However, it is not possible to relate the material to any previously described species because of the poor conservation.
Genus Keokukia Carter, Reference Carter1990
Type species
Keokukia sulcata Carter, Reference Carter1990, from lower Warsaw Formation (Viséan) of Illinois, USA.
Keokukia? sp.
Occurrence
Ixtaltepec Formation, Arroyo las Pulgas type section, level API–2, Upper Mississippian.
Description
Medium-sized, subovate in outline, greatest width at the hingeline, trail of moderate length. Subtriangular ears of medium size. Large shells up to 45 mm in length and 50 mm in width. Ventral valve strongly convex, most inflated in posterior region, not geniculate, with narrow sulcus, moderately deep, originated on visceral disc; flanks steep, moderately convex; ears delimited by concave flexures; massive umbo; ornament consisting of fine costae, rounded, 10–12 per 10 mm on anterior margin, increased by bifurcation; weak rugae on visceral disc. Spines unobserved. Interior not observed.
Materials
Ventral valve (FCMP 1031), two ventral internal molds (FCMP 1032, 1033).
Remarks
Ixtaltepec specimens presents certain features described by Carter (Reference Carter1990) for Keokukia as their transversely to longitudinally subovate outline, ventral valve strongly convex, not geniculate, most convex in umbonal region, greatest width at hingeline, ears well separated by a sulcus, moderate reticulation, with entire surface finely costate and nearly regular rugae on the visceral disc; however, our specimens are larger than the previously described species and the spines were not observed, which along with the lack of material, poor preservation and lack of dorsal valves, prevent us from a reliable assignment.
Subfamily Dictyoclostinae Stehli, Reference Stehli1954
Tribe Dictyoclostini Stehli, Reference Stehli1954
Genus Dictyoclostus Muir-Wood, Reference Muir-Wood1930
Type species
Anomites semireticulatus Martin, Reference Martin1809, from Yorkshire (Viséan) of Derbyshire, England.
Dictyoclostus transversum new species
Diagnosis
Strongly transverse shell with large ears, with length/width ratio of 1:1.5, long trail. Ventral valve most convex in the middle of the total length. With or without shallow median sulcus. Flanks with a slope of 70°. Ornamented by 11–13 costae in space of 10 mm on anterior slope. Spines on visceral disc, flanks and trail, clustered at ears and three spines small on each side of the slopes of umbo. Dorsal valve with the strongest rugae on visceral disc. Cardinal process trilobate, short; median septum extended forward about two-thirds of the length of visceral disc; lateral ridges parallel to hingeline; muscle field small.
Occurrence
Ixtaltepec Formation, Arroyo las Pulgas type section, levels API–7 and API–8, Middle Pennsylvanian.
Description
Medium sized shell, concavo-convex, strongly transverse, with length/width ratio of 1:1.5, subrectangular in outline, with long trail. Large and slightly convex ears, delimitated by concave flexures. Ventral valve strongly convex, most inflated in the middle of the length. A shallow and narrow sulcus, when it is present, begins in the anterior part of visceral disc and extends to end of trail. Flanks moderately steep, with a slope of 70°. Thick and rounded umbo. Ornament consisting of numerous rounded costae, entire, weak on flanks and ears, about 11–13 in space of 10 mm on anterior slope. Rugae numerous, weak and continuous on visceral disc. Erect spines occurring (1) scattered on visceral disc, flanks, and trail; (2) clustered at ears, increased in thickness as they approach to the lateral margins; and (3) three small spines on each side of the slopes of umbo. Interior having a large muscle area, diductor muscle scars longitudinally ridged, elongated. Dorsal valve concave, principally on visceral disc. Short trail. Gentle median fold, corresponding to sulcus of ventral valve. Ornament is similar to the ventral valve. Rugae is most conspicuous on the visceral disc. Spines not observed and presumably absent. Interior with small, internally trilobate cardinal process, with a high middle lobe and posteriorly concave lateral lobes. Median septum extended forward about two-thirds of the length of visceral disc. Lateral ridges short, weak, parallel to hingeline. Muscle field small, adductor scars dendritic, trigonal, slightly elevated, positioned close to hinge. Measurements in millimeters and indices in Table 2.
Table 2 Measurements of Dictyoclostus transversum n. sp.

LE, length; WD, width; TK, thickness; units, millimeters; Costae, number of costae per 10 mm on the anterior margin; e, estimated, sample incomplete.
Etymology
From Latin transversus, the name refers to the distinctive transverse shape of the specimens.
Materials
A ventral internal mold (FCMP 1034) (holotype); four ventral internal molds (FCMP 1035–1038), a ventral external mold (FCMP 1039), a fragment of dorsal internal mold (FCMP 1043), and four dorsal external molds (FCMP 1040–1042, 1044).
Remarks
Taking into account the great number of genera with semireticulate shell, the inclusion of the species in the Tribe Dictyoclostini is because their medium size; trail long, simple; ribbing complete with reticulation posteriorly; ventral spines, dorsal spines absent and dorsal adductor scars positioned close to hingeline (Brunton, Reference Brunton2007); the specimens from Oaxaca are included in the genus Dictyoclostus because their ginglymus absent, spines clustered at ears, otherwise weak and cardinal ridges weak, not extended as ear baffles (Brunton et al., Reference Brunton, Lazarev, Grant and Yu-Gan2000). Dictyoclostus transversum shows a transverse shape, trait shared with Dictyoclostus richardsi (Girty, Reference Girty1927), and Dictyoclostus simplex Campbell, Reference Campbell1957. Dictyoclostus richardsi of the Heath Formation from the Serpukhovian of Montana (Easton, Reference Easton1962) differ from D. transversum by having a smaller size; width slightly greater than length; short ears and visible rugae on the ventral visceral disc. D. simplex of the lower Burindi Group from Tournaisiano (Osagean) of Wales (Campbell, Reference Campbell1957) is dissimilar to the new species by their larger shell, greater convexity in the posterior region, sulcus anterior to visceral disc and greater number of costae.
Subfamily Yakovleviinae Waterhouse, Reference Waterhouse1975
Tribe Yakovleviini Waterhouse, Reference Waterhouse1975
Genus Inflatia Muir-Wood and Cooper, Reference Muir-Wood and Cooper1960
Type species
Productus inflatus McChesney, Reference McChesney1860, from Chester Group (Viséan-Serpukhovian) of Indiana, USA.
Inflatia inflata (McChesney, Reference McChesney1860)
1860 Productus inflatus Reference McChesneyMcChesney, p. 40, pl. 6, figs. 1a–c.
1914 Productus inflatus; Reference WellerWeller, p. 111, pl. 10, figs. 1–6.
1926 Productus inflatus; Reference ButtsButts, p. 188, pl. 61, fig. 13.
1938 Dictyoclostus inflatus; Reference SuttonSutton, p. 563.
1960 Inflatia inflata; Reference Muir-Wood and CooperMuir-Wood and Cooper, p. 227, pl. 55, figs. 1–15.
1962 Inflatia inflata; Reference BransonBranson, p. 243.
1993 Inflatia inflata; Reference Gordon, Henry and TreworgyGordon et al., p. 6, figs. 5.5–5.13.
2007 Inflatia cf. I. inflata; Reference ButtsButts, p. 52, figs. 4.7–4.12.
Occurrence
Ixtaltepec Formation, Arroyo las Pulgas type section, levels API–1 and API–2, Upper Mississippian.
Description
Medium to moderately large sized shell, elongate, long trail. Deep corpus cavity. Greatest width at hingeline. Large shells up to 72 mm in length along arc and 44 mm in width. Ventral valve strongly gibbous over posterior region. Narrow sulcus, slightly deep, originating from umbonal region. Slightly convex umbo. Short ears, subquadrate, delimitated by concave flexures. Ornamented with numerous costae, entire, rounded, separated by slightly narrower intercostal furrows, with few bifurcations over trail, 9–13 per 10 mm over anterior margin. Spines occurring (1) scattered on the trail and visceral disc, and (2) in single row of five or six along hinge margin. Rugae numerous on posterior region, strongest on ears. Dorsal valve moderately concave over visceral disc. One median fold, corresponding to sulcus of ventral valve. Ornamented by costae similar to ventral valve, but the rugae are more developed. No spines present. Interiors not observed.
Materials
Three permineralized shells (FCMP 1045–1047).
Remarks
The specimens have all the features mentioned by Gordon et al., (Reference Gordon, Henry and Treworgy1993, p. 7) for I. inflata, but the specimens of Ixtaltepec present a slightly larger size. The species occurs in the Upper Mississippian of United States in Montana (Brewster, Reference Brewster1984), Oklahoma, Arkansas (Gordon et al., Reference Gordon, Henry and Treworgy1993), and Idaho (Butts, Reference Butts2007), as well as in Egypt (Kora, Reference Kora1995) and Australia (Roberts, Reference Roberts1975). The genus Inflatia is described for the Middle Carboniferous from Brazil, Egypt, Australia and North America. This is the first report of the genus in Mexico.
Inflatia coodzavuii new species

Figure 4 Inflatia coodzavuii n. sp., (1) FCMP 1050, ventral internal mold; (2) dorsal valve interior, rubber mold of FCMP 1054; (3) FCMP 1054 (holotype), dorsal internal mold; (4) FCMP 1057, dorsal internal mold; (5) FCMP 1053, ventral internal mold; (6) FCMP 1052, ventral internal mold; (7) FCMP 1058, dorsal external mold; Reticulatia cf. R. huecoensis (King, Reference King1931), (8) FCMP 1061, dorsal external mold; (9) FCMP 1059, ventral internal mold; (10) FCMP 1062, dorsal external mold; (11) FCMP 1060, ventral internal mold; Buxtonia websteri Beus and Lane, Reference Beus and Lane1969, (12) FCMP 1064, ventral internal mold; (13) FCMP 1065, ventral internal mold; (14) dorsal valve interior, rubber mold of FCMP 1067; (15) FCMP 1066, dorsal internal mold; Buxtonia inexpletucosta n. sp., (16) FCMP 1076 (holotype), dorsal external mold; (17) FCMP 1078, dorsal external mold; (18) FCMP 1079, dorsal internal mold; (19) FCMP 1077, dorsal valve, external mold; (20) rubber mold of FCMP 1078; (21) FCMP 1070, ventral valve, inner mold; (24) rubber cast of dorsal internal mold of FCMP 1080; (25) dorsal valve, rubber mold of FCMP 1079; Flexaria magna n. sp., (22, 23) dorsal valve interior, rubber mold of FCMP 1088; (26) FCMP 1085, composed mold, with part of dorsal external mold and part of ventral internal mold; (27) FCMP 1086 (holotype), composed mold, with part of dorsal external mold and part of ventral internal mold; (28) cardinal process strongly recurved dorsally, viewed ventrally, rubber mold of FCMP 1090; (29) FCMP 1091, dorsal external mold; (30) FCMP 1092, dorsal internal mold; (31) FCMP 1084, ventral internal mold; (32) FCMP 1085, ventral internal mold. Scale bar represents 1.0 cm.
Diagnosis
Shell subquadrate in outline, widest in hingeline. Moderately small ears. Sulcus narrow, beginning about 4 mm anterior to beak and continuing until anterior margin; umbo protruding 2 mm across hingeline; 14–16 costae in the space of 10 mm on anterior slope. Some spines over visceral disc, four spines as much as 1.4 mm of thickness horizontally arranged over trail, and simple row of two or four spines parallel to hingeline. Median septum extends from cardinal process to midlength. Brachial ridges curving to the height of median septum, initiating at low angle parallel to hinge.
Occurrence
Ixtaltepec Formation, Arroyo las Pulgas type section, levels API–3, API–7, and API–8, Upper Mississippian and Middle Pennsylvanian.
Description
Concavo-convex shell, small- to medium-sized, subquadrate in outline. Greatest width at hingeline, length generally greater than width; strongly curved in lateral profile. Ventral valve rather deep; visceral disc highly arched in profile. Strongly steep lateral slopes. Small ears, slightly arched, subquadrate to subtriangular, separated from umbonal slopes by a shallow concave flexure. Narrow median sulcus shallow to obsolete, extending from roughly four millimeters of beak to end of trail. Umbonal region rounded, umbo protruding 2 mm across hingeline. Ornamented by rounded costae, entire, 14–16 occurring in the space of 10 mm on anterior slope. Narrow, discontinuous, and irregular rugae on visceral disc and ears. Erect spines above costae occurring (1) four on anterior slope of roughly 1.4 mm in diameter, (2) in posterior row of one or two spines of small diameter parallel to hingeline, and (3) scattered over visceral disc. Interior having narrow ginglymus, muscle area large, diductor muscle scars longitudinally ridged, occupying subcircular depressions at either side. Concave dorsal valve, having weak median fold corresponding to sulcus of ventral valve. Ornamented by costae similar to the ventral valve. Concentric rugae on visceral disc. Aspinose. Interior having straight lateral ridges; median septum extends from cardinal process to midlength. Dendritic posterior pair of adductor scars, in subtriangular platforms; the anterior pair smooth. Brachial ridges curving to the height of median septum, initiating at low angle parallel to hinge. Endospines on trail. Measurements in millimeters and indices in Table 3.
Table 3 Measurements of Inflatia coodzavuii n. sp.

LE, length; LA, length along arc; WD, width; TK, thickness; units, millimeters; Costae, number of costae per 10 mm on the anterior margin; e, estimated, sample incomplete.
Etymology
The specific nomination is conferred by the personal name ‘Coo Dzavui,’ one of the principal deities of the Mixtec culture; major ethnic group in the northwestern region of Oaxaca.
Materials
A dorsal internal mold with ventral internal mold (FCMP 1054) (holotype); eight ventral internal molds (FCMP 1048–1053, 1055, 1056), a dorsal valve internal mold (FCMP 1057), and a dorsal valve external mold (FCMP 1058).
Remarks
Inflatia coodzavuii is similar in size and shape to Inflatia lovei Gordon, Reference Gordon1975, of the Amsden Formation from Serpukhovian of Wyoming (p. 36, pl. 3, figs. 1–9), but it differs from I. lovei by an increased convexity to a half of the total length; absence of spines on the lateral flanks; presence of only four spines on the trail and a shallow depression that differentiates the ears. Inflatia inflata (McChesney) of Fayetteville Shale from Serpukhovian of Arkansas (Gordon et al., Reference Gordon, Henry and Treworgy1993) differs from I. coodzavuii by larger size; greatest number of spines parallel to the hinge; most spines on the trail; strongly concave dorsal valve and concentric rugae on the trail. Inflatia cherokeensis (Drake, Reference Drake1897), of Fayetteville Shale from Serpukhovian of Arkansas (Gordon et al., Reference Gordon, Henry and Treworgy1993) is different from I. coodzavuii by the subrectangular form, greater number of spines on the visceral disc, most number of spines in a row parallel to the hinge, spines absent on trail, prominent ears, and ventral sulcus that starts 4 mm of the beak. Inflatia gracilis Gordon, Henry, and Treworgy, Reference Gordon, Henry and Treworgy1993, of Fayetteville Shale from Serpukhovian of Oklahoma is distinguished from I. coodzavuii by the strongly curved ventral valve; with a beak that extends 1 mm past hinge; most costae in the anterior margin; most spines on visceral disc and trail; greater number of spines parallel to the hingeline. Inflatia pusilla Gordon, Henry, and Treworgy, Reference Gordon, Henry and Treworgy1993, of Fayetteville Shale from Serpukhovian of Oklahoma is dissimilar to I. coodzavuii in its smaller size; subcircular in outline; greater number of costae in the anterior margin; greater number of spines in row parallel to the hingeline and most number of spines scattered on visceral disc and trail.
Tribe Reticulatiini Lazarev, Reference Lazarev2000
Genus Reticulatia Muir-Wood and Cooper, Reference Muir-Wood and Cooper1960
Type species
Productus huecoensis King, Reference King1931, from Gym Formation (Gzhelian) of Texas, USA.
Reticulatia cf. R. huecoensis (King, Reference King1931)
1931 Productus huecoensis Reference KingKing, p. 68, pl. 11, figs. 7–8.
1938 Dictyoclostus newelli Reference KingKing, p. 272, pl. 39, figs. 1–4.
1960 Reticulatia huecoensis; Reference Muir-Wood and CooperMuir-Wood and Cooper, p. 284, pl. 104, fig. 1–5, pl. 105, figs. 1–8.
1961 Reticulatia huecoensis; Reference HoareHoare, p. 50, pl. 5, fig. 8–10.
1968 Reticulatia huecoensis; Reference Sturgeon and HoareSturgeon and Hoare, p. 49, pl. 16, figs. 1–4.
2001 Reticulatia huecoensis; Reference Olszewski and PatzkowskyOlszewski and Patzkowsky, p. 666.
Occurrence
Ixtaltepec Formation, Arroyo las Pulgas type section, level API–7, Middle Pennsylvanian.
Description
Shell large, subrectangular in outline, with greatest width at the hinge. Large ears, triangular, slightly convex. Large shells up to 28 mm in length and 43 mm in width. Ventral valve convex. Shallow sulcus, extending from the beak to the edge of trail. Steep flanks. The beak protruding around of 4 mm from hingeline, umbo convex. Surface ornamented by rounded costae, entire, 8 or 10 in 10 mm along the anterior margin, with numerous closely spaced concentric rugae; crossed on posterior third of shell with the costae, forming a strong reticulation. Spines of small diameter on trail and lateral margins in arranged quincuncially. Dorsal valve concave, geniculated, well-developed trail. Ornamentation similar to the ventral valve, without spines. Interior not observed.
Materials
Two ventral internal molds (FCMP 1059, 1060) and three dorsal external molds (1061–1063).
Remarks
Lazarev in 2000 restricts the genus Reticulatia to the Permian of North America, establishing that the Carboniferous taxa belong rather to the genus Admoskovia, located in Eurasia and mid-Asia (Brunton, Reference Brunton2007). Admoskovia can be differentiated from Reticulatia for their ears strongly defined, reticulation in discs relatively weak, spines on ears strongly developed (Brunton, Reference Brunton2007); features not shared with the Ixtaltepec specimens. For these differences and the similarity with the diagnosis made by Muir-Wood and Cooper (Reference Muir-Wood and Cooper1960, p. 284) for Reticulatia, we consider these carboniferous specimens belong to the latter. The specimens of Reticulatia cf. R. huecoensis from Oaxaca are similar to the Carboniferous material figured in Sturgeon and Hoare (Reference Sturgeon and Hoare1968, pl. 16, fig. 1–4) for Ohio, but the poor preservation of our material and the absence of spines parallel to hingeline does not allow us to identify it with more certainty.
Subfamily Buxtoniinae Muir-Wood and Cooper, Reference Muir-Wood and Cooper1960
Tribe Buxtoniini Muir-Wood and Cooper, Reference Muir-Wood and Cooper1960
Genus Buxtonia Thomas, Reference Thomas1914
Type species
Anomites scabriculus Martin, Reference Martin1809, from Yorkshire (Viséan) of Derbyshire, England.
Buxtonia websteri Beus and Lane, Reference Beus and Lane1969
1969 Buxtonia websteri Reference Beus and LaneBeus and Lane, p. 994, pl. 119, figs. 9, 14, 17, 19.
Occurrence
Ixtaltepec Formation, Arroyo las Pulgas type section, levels API–6, API–7 and API–8, Middle Pennsylvanian.
Description
Large shell, subquadrate, with greatest width at the hinge, large ears. Large shells up to 47 mm in length along arc and 40 mm in width. Ventral valve strongly convex; beak small and acute; median sulcus wide and shallow; flattened ears separated from umbonal slopes by a shallow concave flexure; surface covered with incomplete costae, 10–11 per 10 mm over anterior margin. Prominent rugae on the ears and umbonal slopes; inconspicuous across the visceral disc and trail. Spines occurring (1) in row on posterior slope of rugae on ears and umbonal slopes, and (2) scattered on the visceral disc and trail. Interior not observed. Dorsal valve slightly flattened, with broad median fold; concentric rugae on visceral disc and ears; narrow spines scattered on the valve. Interior with a long cardinal process, bilobate, a pit between the converging buttress plates is present at the base. The median septum is long, anteriorly narrow and broadly rounded posteriorly. The lateral ridges are parallel to hinge, laterally indistinct. Scars of adductor muscles are dendritic, low, and suboval in outline.
Materials
Two ventral compound molds (FCMP 1064, 1065) and two dorsal compound molds (FCMP 1066, 1067).
Remarks
The specimens from Santiago Ixtaltepec resembles with the material of Beus and Lane (Reference Beus and Lane1969) from Nevada. In plate 119, the figures 9 and 19 show the convex profile of the ventral valve; in the case of interior and exterior dorsal valve of our material, it is similar to the pictures seen in the plate 119 (figures 14, 17); however, the specimens of Oaxaca are smaller than those from Nevada.
Buxtonia inexpletucosta new species
Diagnosis
Suboval shell, ventral valve convex, median sulcus shallow. The beak protrudes five millimeters beyond hinge. Irregularly developed costae with indistinct bifurcations, 9–11 occurring in a space of 10 mm over anterior margin, absent on ears. Spines on the costae set in concentric rows arranged in quincunx. Dorsal valve geniculate in trail. Flattened ears, not clearly differenced from the umbonal region. Cardinal process bilobate, with an antron in the base; median septum long; adductor scars extending from the middle of antron to point about two-thirds of the total length of the septum.
Occurrence
Ixtaltepec Formation, Arroyo las Pulgas type section, level API–7, Middle Pennsylvanian.
Description
Medium-sized concavo-convex buxtoniid. Suboval in outline.Large trail, small ears, with greatest width at the hingeline.Ventral valve convex. Umbo rounded, narrow. Beak projecting 5 mm beyond hinge. Median sulcus shallow, initiating anteriorly to the visceral disc; broad in its front. Flattened ears, separated from umbonal slopes by a shallow concave flexure. Ornamented by irregularly developed costae with indistinct bifurcations, 9–11 per 10 mm over anterior margin; absent on ears. Spines on the costae set in concentric rows arranged in quincunx on the valve, larger in lateral margins and trail. Concentric faint rugae on the visceral disc; prominent on the ears and lateral margins. Interior unknown. Dorsal valve is flat to slightly concave in the visceral disc. A median fold is developed corresponding to the ventral sulcus, initiating at mid-length of the valve, extending to anterior margin with geniculated trail. Flattened ears, not clearly demarcated from the umbonal region. The ornamentation is similar to that of the ventral valve, but the rugae are prominent. Interior with a bilobate cardinal process, posteriorly trilobate. The median septum is long, broadly rounded posteriorly, narrower and overhanging anteriorly. A distinctive antron present at the base of the process. The lateral ridges are slightly divergent from the hinge. The adductor scars are dentritic, elongated, extending from the middle of antron to about two-thirds of the total length of the septum. Measurements in millimeters and indices in Table 4.
Table 4 Measurements of Buxtonia inexpletucosta n. sp.

LE, length; WH, width at hinge; GW, greatest width; units, millimeters; Costae, number of costae per 10 mm on the anterior margin; e, estimated, sample incomplete.
Etymology
The name is assigned from the Latin words inexpletus and costae, referring to the constant interruption of the ribs through the entire shell.
Materials
A dorsal external mold (FCMP 1076) (holotype); seven ventral internal molds (FCMP 1068–1074), two dorsal internal molds (FCMP 1079, 1080), four dorsal external molds (FCMP 1075–1078).
Remarks
Buxtonia scabriculoides (Paeckelmann, Reference Paeckelmann1931) of the Los Santos de Maimona locality from Viséan-Serpukhovian (Serpukhovian) of Spain (Martínez-Chacón and Legrand-Blain, Reference Martínez-Chacón and Legrand-Blain1992) is distinguished from B. inexpletucosta by their largest size, transverse shell, ventral valve strongly convex and more numerous costae in the anterior margin. Buxtonia arizonensis Hernon, Reference Hernon1935, of the Alaska Beach locality from Serpukhovian-Bashkirian of Montana (Easton, Reference Easton1962) is dissimilar of the new species by its greater size; with less numerous costae on the anterior margin and lateral ridges parallel to the hinge. Buxtonia websteri Beus and Lane, of the Bird Spring Formation from Moscovian of Nevada (Beus and Lane, Reference Beus and Lane1969) differs from B. inexpletucosta by its larger size; more convex ventral valve; long ears; rugae are more noticeable that the costae and dorsal valve with fold most notorious. Buxtonia suttoni Elias, of the Redoak Hollow Formation from Serpukhovian of Oklahoma (Elias, Reference Elias1957) is different from B. inexpletucosta in its smaller size; an alveolus in the base of the cardinal process and one median septum reaches two-thirds of the total length. Flexaria magna n. sp. of the Ixtaltepec Formation is dissimilar to B. inexpletucosta by its larger size; different number of costae; with a cardinal process very curved dorsally; flattened visceral disc and larger ears.
Genus Flexaria Muir-Wood and Cooper, Reference Muir-Wood and Cooper1960
Type species
Productus arkansanus Girty, Reference Girty1910, from Fayetteville Shale (Viséan-Serpukhovian) of Arkansas, USA.
Flexaria magna new species
Diagnosis
Medium to large-sized shell. Irregularly developed costae on the visceral disc; on the trail the costae are complete; 9–11 occurring anteriorly in a space of 10 mm. Spines on the costae, scattered over both valves, forming concentric rows on the trail. Concentric rugae on visceral disc and ears. Dorsal valve with flattened visceral disc, geniculate trail. Cardinal process bilobate, long, strongly recurved dorsally; long median septum with an alveolus in the base; the adductor scars are subtriangular, extending to about two-thirds of the total length of the septum; lateral ridges parallel with the hingeline.
Occurrence
Ixtaltepec Formation, Arroyo las Pulgas type section, levels API–7 and API–8, Middle Pennsylvanian.
Description
Medium to large-sized productid. Subrectangular in outline. Short trail and ears. With greatest width at the hinge. Ventral valve convex, more so in the posterior region. Deep corpus. Median sulcus is narrow, shallow, initiating at the anterior of the visceral disc until the anterior margin. Steep flanks. Flattened and subtrigonals ears. Broad umbo; the beak slightly overhanging hinge. The ornamentation consists of irregularly developed costae in the posterior region; in the trail, the costae are complete; absent in the ears; 9–11 occurring in a space of 10 mm over anterior area, with irregular bifurcations, separated by narrow and rounded intercostal grooves. Spines are on the costae, scattered over valve; larger spines and numerous on trail where forming concentric rows. Concentric rugae on visceral disc; more notorious and continuous in the ears. Interior not observed. Dorsal valve with flattened visceral disc. Strong geniculation rounded the visceral disc. Median fold weakly broad, corresponding to sulcus of ventral valve. Ornamentation is similar to opposite valve, but the spines are large and more numerous in the ears and anterior region, shorter on the visceral disc. The rugae are more prominent on the visceral disc and ears. Interior with a long cardinal process, narrow, anteriorly bilobate and posteriorly trilobate, strongly recurved dorsally. Cardinal process is attached to a long median septum, with an alveolus in the base. The adductor scars are subtriangular, dendritic, enlarged, extending to a point about two-thirds of the total length of the septum. The lateral ridges are parallel with the hingeline, well developed in the base of the cardinal process, but imperceptible in the lateral margins. Measurements in millimeters and indices in Table 5.
Table 5 Measurements of Flexaria magna n. sp.

LE, length; WH, width at hinge; GW, greatest width; units, millimeters; Costae, number of costae per 10 mm on the anterior margin; e, estimated, sample incomplete.
Etymology
From Latin magnus, the name refers to the large size of this species compared to other forms described for the genus.
Materials
A composed mold, with part of dorsal external mold and part of ventral internal mold (FCMP 1086) (holotype); a composed mold, with part of dorsal external mold and part of ventral internal mold (FCMP 1085), five ventral internal molds (FCMP 1081–1085), three dorsal internal molds (FCMP 1088, 1090, 1092), and three dorsal external molds (FCMP 1087, 1089, 1091).
Remarks
Unlike the previously described species of Flexaria the specimens of Santiago Ixtaltepec are easily distinguished by having a larger size. Flexaria floydensis (Sutton, Reference Sutton1942), of the Crawfordsville region from Osagean of Indiana (Lane et al., Reference Lane, Matthews, Driscoll and Yochelson1973) is dissimilar to F. magna by the absent ventral sulcus; more number of costae in the anterior margin and anterior spines slightly developed. Flexaria arkansana (Girty, Reference Girty1910), of Fayetteville Shale from Serpukhovian of Arkansas (Girty, Reference Girty1910) differs from the new species in the absence of ventral sulcus; more numerous costae on the anterior margin and absence of rugae on the ears. Equally, F. magna are distinguished of B. websteri of the Ixtaltepec Formation by the presence of a curved dorsally cardinal process; visceral disc flattened and rugae most notorious on visceral disc and ears. This is the first report of Flexaria in Mexico.
Acknowledgments
The authors thank S. Butts for critically reviewing the manuscript and her help in the revision of specimens from the collection of the Peabody Museum, Yale University; to M.L. Martínez-Chacón, L. Benedetto, and B. Pratt for critically reviewing the manuscript and for their valuable comments; to J. Hagadorn for reviewing the text; to H. Hernández for his photographic work and to D. Navarro and L. Martin for their field work and their technical assistance. Torres-Martínez received support from the Schuchert and Dunbar Grants in Aid, Yale Peabody Museum. This work is part of the Ph.D. thesis of M. A. Torres-Martínez entitled “Braquiópodos carboníferos del área de Santiago Ixtaltepec, Oaxaca. Implicaciones paleoambientales, estratigráficas y paleobiogeográficas,” sponsored by CONACyT with scholarship 174854 and by the Dirección General de Asuntos del Personal Académico (DGAPA), UNAM through PAPIIT project IN213710 and IN215013.









