

This article is the first to describe in detail and illustrate spore assemblages recovered from the ‘Lower Old Red Sandstone’ ‘Basement Beds’ of the Berriedale, or Ousdale–Badbea, Outlier of the Northern Highlands of Scotland. It represents a further contribution to a series of papers describing dispersed spore assemblages from Upper Silurian–Lower Devonian ‘Lower Old Red Sandstone’ deposits of Scotland (Wellman Reference Wellman1993a, Reference Wellmanb, Reference Wellman1994, Reference Wellman2006, Reference Wellman2010; Wellman & Richardson Reference Wellman and Richardson1993, Reference Wellman and Richardson1996; Lavender & Wellman Reference Lavender and Wellman2002). The aim is to: (i) biostratigraphically age date and stratigraphically correlate these deposits; (ii) use this information to improve understanding of the complex geological relationships of the ‘Lower Old Red Sandstone’ deposits of this region (particularly regarding their relationship to the assembly of the Old Red Sandstone continent); and (iii) shed light on the palaeoenvironments and biotas of these deposits.
1. Geological setting
The Devonian ‘Old Red Sandstone’ deposits of the Orcadian Basin straddle the Great Glen Fault System and, as such, outcrop on both the Northern Highlands to the northwest and the Grampian Highlands to the southeast. This laterally and vertically extensive sequence rests unconformably on an ancient landscape consisting largely of Moine schists. Over most of the area, there is a thin basal unit (originally termed the ‘Basement Group’ or ‘Barren Group’) that is generally considered to be ‘Lower Old Red Sandstone’ of Early Devonian age. The ‘Basement Group’ is overlain by ‘Middle Old Red Sandstone deposits’ of Mid Devonian age. The contact is either unconformable (e.g., the Berriedale Outlier) or disconformable (e.g. the Sarclet Outlier). The ‘Basement Group’ of the Northern Highlands north of the Great Glen Fault System consists of a series of discrete outcrops (Fig. 1):
(i) the Sarclet Outlier of the Sarclet Dome;
(ii) the “northern Lower Old Red Sandstone” (sensu Trewin & Thirlwall Reference Trewin, Thirlwall and Trewin2002), outcropping in a strip from Braemore northwards to Shurrery, and including the smaller Strathy, Ben Griam, Kirtomy, Roan Island and Tongue Outliers to the west of this;
(iii) the Badbea Basin (Berriedale, or Ousdale–Badbea, Outlier), located north of the Helmsdale Granite (Dec Reference Dec1992);
(iv) the Golspie Basin (Brora Outlier), located south of the Helmsdale Granite (Dec Reference Dec1992);
(v) the Meall Odhar or Crask Outlier;
(vi) the Strath Rannoch Outlier;
(vii) Struie–Strathpeffer;
(viii) the Den Siltstone Formation (Fletcher et al. Reference Fletcher, Auton, Highton, Merritt, Roberston and Rollin1996); and
(ix) the Mealfuarvonie Outlier (Mykura & Owens Reference Mykura and Owens1983).

Figure 1 Map of the eastern part of the Northern Highlands of Scotland, showing the distribution of Lower Devonian ‘Lower Old Red Sandstone’ deposits.
The ‘Lower Old Red Sandstone’ deposits of the Badbea Basin (sensu Dec Reference Dec1992) are often referred to as the Berriedale, or Ousdale-Badbea, Outlier. They are mapped on the British Geological Survey 1:50 000 Series Scotland Sheets 109 and 110. The geology of these deposits is described in the associated Memoirs of the Geological Survey of Scotland (Crampton & Carruthers Reference Crampton and Carruthers1914; Read Reference Read1931). More recently, the geology of the deposits has been considered by Westoll (Reference Westoll, House, Richardson, Chaloner, Allen, Holland and Westoll1977), Friend & Williams (Reference Friend and Williams1978), Dec (Reference Dec1992), Trewin (Reference Trewin, Trewin and Hurst1993, Reference Trewin, Trewin and Hurst2009a) and Trewin & Thirlwall (Reference Trewin, Thirlwall and Trewin2002) (Figs 2 & 3).

Figure 2 Geological map of the Berriedale Outlier, illustrating the location of sampled exposures. The numbers 1–4 refer to roadcut and roadside quarry localities in Trewin (Reference Trewin, Trewin and Hurst1993; Reference Trewin, Trewin and Hurst2009a) and the letters A–C refer to exposures in Ousdale Burn (see Table 2).
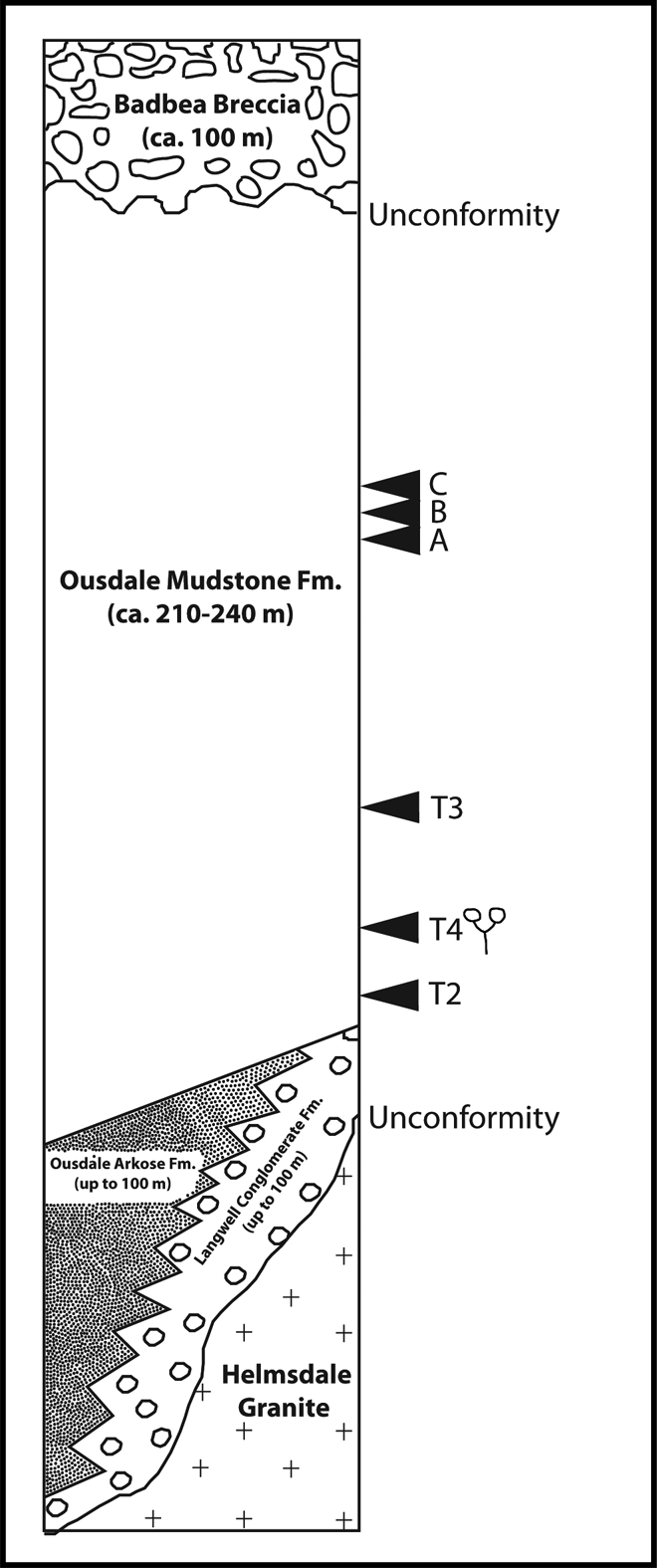
Figure 3 Stratigraphical succession in the Berriedale Outlier. The approximate position of productive palynological samples was calculated based on strike/dip information and measured distances using British Geological Survey 1:50,000 Series Scotland Sheets 109 and 110. T2–T4 refer to roadcut and roadside quarry localities in Trewin (Reference Trewin, Trewin and Hurst1993; Reference Trewin, Trewin and Hurst2009a) and A–C refer to exposures in Ousdale Burn. Productive samples are: T4 (AOU7, BOU4, BOU5); T3 (BOU3); A (BOU6, BOU7, BOU8); B (BOU9); C (BOU10, BOU11) (see Table 2). The location of plant remains discovered at locality T4 is indicated by the sketch of a Lower Devonian fossil plant.
The Berriedale Outlier is separated from the main strip of “northern Lower Old Red Sandstone” (sensu Trewin & Thirlwall (Reference Trewin, Thirlwall and Trewin2002), see (ii) above) by a narrow gap across the Scaraben–Newport ridge of basement rocks. The ‘Lower Old Red Sandstone’ deposits lie on an irregular surface consisting of the Helmsdale Granite, which is intruded into the Moine schists. Basal deposits consist of a regolith of Helmsdale Granite and fossil screes that are overlain by red-brown indurated sandstones. The thin impersistent basal conglomerate is called the Langwell Conglomerate Formation. It passes laterally into the Ousdale Arkose Formation. These are in turn overlain by the Ousdale Mudstone Formation, which consists predominantly of brownish siltstones (but with some thin green and grey layers) with some sandier layers. The ‘Lower Old Red Sandstone’ sequence is unconformably overlain by the ‘Middle Old Red Sandstone’ Badbea Breccio–Conglomerate Member. The sequence is interpreted as a low energy floodplain deposit, with drainage to the northeast, interrupted by occasional sheet-wash deposits of very local origin (Friend & Williams Reference Friend and Williams1978; Trewin Reference Trewin, Trewin and Hurst1993, Reference Trewin, Trewin and Hurst2009a).
The Ousdale Mudstone Formation has yielded scales of the fish Porolepis (Collins & Donovan Reference Collins and Donovan1977), plant fragments (“Psilophyton”) and nematophytes (Pachytheca) (Collins & Donovan Reference Collins and Donovan1977), dispersed spore assemblages (Richardson Reference Richardson1967; Collins & Donovan Reference Collins and Donovan1977) and trace fossils (Trewin Reference Trewin, Trewin and Hurst1993, Reference Trewin, Trewin and Hurst2009a). Together, these fossils remains have been interpreted to suggest an age high in the Early Devonian (Westoll Reference Westoll, House, Richardson, Chaloner, Allen, Holland and Westoll1977; Friend & Williams Reference Friend and Williams1978; Trewin Reference Trewin, Trewin and Hurst1993, Reference Trewin, Trewin and Hurst2009a).
2. Palynology
2.1. Previous palynological investigation
Richardson (Reference Richardson1967) reported relatively poorly preserved and highly coalified spore assemblages recovered from the Ousdale “mudstones” of the Basement Group of Berriedale. He noted that the assemblage consisted entirely of azonate spores that in the most part could be assigned to Retusotriletes, ?Apiculiretusispora and Emphanisporites (rotatus type). Richardson (Reference Richardson1967) suggested that the spore assemblage was most likely of Early Devonian age. Subsequently, Collins & Donovan (Reference Collins and Donovan1977) reported on a moderately well preserved but slightly coalified spore assemblage recovered from the Ousdale Mudstones from road cuttings on the A9. They listed the species present and their relative abundance (Table 1). They suggested an early Emsian age based on the spores.
Table 1 Spore taxa and their abundance reported by Collins & Donovan (Reference Collins and Donovan1977). P=Present (<1 %); C=Common (1–10 %); A= Abundant (>10 %).

2.2. Methods
During the course of two fieldtrips, 18 siltstone horizons were sampled from throughout the ‘Lower Old Red Sandstone’ sequence of the outlier (Table 2). They were processed using standard palynological techniques: HF-HCl-HF acid maceration followed by heavy liquid separation using zinc chloride. Recovered organic residues were sieved using a 20 μm mesh, oxidised for varying periods of time using Schultz solution, and strew mounted for light microscope analysis. Of the 18 samples, ten were productive and all came from the Ousdale Mudstone Formation. The assemblages are rich in palynomorphs that are of good to excellent preservation and moderate thermal maturity (Thermal Alteration Index 3- to 3 from the chart provided in Traverse Reference Traverse2007). Oxidation with Schultz solution (a maximum of 30 minutes was required) cleared the spores to a workable translucent dark orange–pale brown colour. All materials (samples, residues, slides) are housed in the collections of the Centre for Palynology of the University of Sheffield.
Table 2 Details of samples (listed in approximate stratigraphical order from oldest to youngest); see Figs 2–3. B=barren; P=productive.

2.3. Description of spore assemblages
All of the productive samples essentially yield an identical palynomorph assemblage (see Table 1; Figs 4–6). They contain only dispersed spores and palynodebris. The dispersed spores are dominated by laevigate retusoid spores (Retusotriletes spp.) (Fig. 4a, b) and apiculate retusoid spores, with a sloughing extra-exosporal layer that bears the ornament (Apiculiretusispora spp.) (Fig. 4c–e). Many of the Retusotriletes have thickenings (triangular or annular) associated with the trilete mark and extra-exosporal material adhering to them (Fig. 4a, b). Similarly, some of the Apiculiretusispora have a triangular thickening associated with the trilete mark (Fig. 4d, e). Other apiculate retusoid spores have a biform sculpture (Dibolisporites spp., including D. ardchoircii, D. echinaceus and D. eifeliensis) (Fig. 4f–k). The genus Dictyotriletes is notably under-represented, with only very rare examples (e.g., Fig. 5c). Laevigate crassitate (Ambitisporites spp.) (Fig. 4b) and patinate (Archaeozonotriletes chulus) (Fig. 6g, h) spores are relatively uncommon. Ornamented crassitate forms are present, including apiculate forms (Aneurospora sp. A) (Fig. 5d, e), verrucate forms (Verrucosisporites polygonalis) (Fig. 6k) and foveolate forms placed with Brochotriletes spp. (Fig. 6a–c, j), including B. bellatulus (Fig. 6b, c). Ornamented patinate forms are extremely rare (e.g., Cymbosporites spp.) (Fig. 6i, j). The highly distinctive spore Amicosporites streelii (Fig. 6d) is identified. Spores with proximal radial ribbing are rare, but present in all samples, and include examples of E. annulatus, E. erraticus, E. rotatus, E. schultzii and E. zavallatus? (Fig. 5f, i–n). Zonate spores are present and include Camptozonotriletes caperatus (Fig. 6p), a new species of Samarisporites (Fig. 6l, m) and a solitary possible example of Breconisporites (Fig. 6o). Very rare cryptospores are present and are represented by the laevigate hilate spores (Laevolancis divellomedium) (Fig. 6q) and the permanent tetrad Tetrahedraletes medinensis (Fig. 6n). A complete list of identified spore taxa is provided in Table 3.

Figure 4 Light microscope images of dispersed spores from the Ousdale Mudstone Formation of the Berriedale Outlier: (a) Retusotriletes cf. triangulatus (Streel) Streel, Reference Streel1967, slide BOU3/1, E.F. No. (H28/1); (b) Retusotriletes rotundus (Streel) Streel, Reference Streel1967, slide BOU3/1, E.F. No. (J44); (c) Apiculiretusispora plicata (Allen) Streel, Reference Streel1967, slide BOU3/1, E.F. No. (J24/1); (d) Apiculiretusispora brandtii Streel, Reference Streel1967, slide BOU9/1, E.F. No. (D47/4); (e) Apiculiretusispora brandtii Streel, Reference Streel1967, slide BOU11/1, E.F. No. (J50/3); (f) Dibolisporites eifeliensis (Lanninger) McGregor, Reference McGregor1973, slide BOU3/1, E.F. No. (T46/2); (g) Dibolisporites ardchoircii Wellman & Richardson, Reference Wellman and Richardson1996, slide BOU3/1, E.F. No. (F44); (h) Dibolisporites cf. ardchoircii Wellman & Richardson, Reference Wellman and Richardson1996, slide BOU4/1, E.F. No. (H55/1). Note that this specimen has some sculptural elements that are spatulate; (i) Dibolisporites echinaceous (Eisenack) Richardson, Reference Richardson1965, slide BOU10/1, E.F. No. (T26); (j) Dibolisporites eifeliensis (Lanninger) McGregor, Reference McGregor1973, slide AOU7/4, E.F. No. (J38/1); (k) Dibolisporites eifeliensis (Lanninger) McGregor, Reference McGregor1973, slide AOU7/2, E.F. No. (R50/1). Scale bar=20 µm.

Figure 5 Light microscope images of dispersed spores from the Ousdale Mudstone Formation of the Berriedale Outlier: (a) Dibolisporites sp., slide BOU11/1, E.F. No. (F35/2); (b) Ambitisporites sp., slide BOU8/1, E.F. No. (M29); (c) ?spore of Dictyotriletes-type, slide BOU9/1, E.F. No. (G48/4); (d) Aneurospora sp. A, slide BOU11/1, E.F. No. (G40); (e) Aneurospora sp. A, slide BOU3/1, E.F. No. (Q46/1); (f) Emphanisporites rotatus McGregor, Reference McGregor1961, slide BOU7/1, E.F. No. (N26); (g) spore with distinctive reticulate ornament, slide BOU4/1, E.F. No. (E25); (h) spore with ornament of distinctive proximal striae and distal coni, slide BOU8/1, E.F. No. (Q42); (i) Emphanisporites rotatus McGregor, Reference McGregor1961 (robustus-type), slide BOU9/1, E.F. No. (S42); (j) Emphanisporites zavallatus? Richardson et al., Reference Richardson, Streel, Hassan and Steemans1982, slide BOU3/1, E.F. No. (M24); (k) Emphanisporites schultzii McGregor, Reference McGregor1973, slide BOU3/1, E.F. No. (V39/1); (l) Emphanisporites annulatus McGregor, Reference McGregor1961, slide AOU7/3, E.F. No. (K45); (m) Emphanisporites erraticus (Eisenack) McGregor, Reference McGregor1961, slide BOU3/1, E.F. No. (Q23); (n) Emphanisporites annulatus McGregor, Reference McGregor1961, slide BOU10/1, E.F. No. (K49/2). Scale bar=20 µm.

Figure 6 Light microscope images of dispersed spores from the Ousdale Mudstone Formation of the Berriedale Outlier: (a) Brochotriletes foveolatus? Naumova, Reference Naumova1953, slide BOU3/1, E.F. No. (F51/3); (b) Brochotriletes bellatulus Steemans, Reference Steemans1989, slide BOU3/1, E.F. No. (G39); (c) Brochotriletes bellatulus Steemans, Reference Steemans1989, slide BOU3/1, E.F. No. (T46/2); (d) Amicosporites streelii Steemans, Reference Steemans1989, slide BOU5/1, E.F. No. (Q40/1); (e) distinctive patinate spore, slide BOU9/1, E.F. No. (G47); (f) Brochotriletes rarus Arkangelskaya, 1978, slide BOU3/1, E.F. No. (D47); (g) Archaeozonotriletes chulus (Cramer) Richardson & Lister, Reference Richardson and Lister1969, slide BOU3/1, E.F. No. (N51/3); (h) Archaeozonotriletes chulus (Cramer) Richardson & Lister, Reference Richardson and Lister1969, slide BOU9/1, E.F. No. (F32); (i) Cymbosporites sp., slide BOU6/1, E.F. No. (L33); (j) Cymbosporites sp., slide BOU7/1, E.F. No. (R49); (k) Verrucosisporites polygonalis Lanninger, Reference Lanninger1968. Note that there is considerable integradation between two characters in the Scottish population of this species: verrucae vary from rounded to polygonal and structure varies from retusoid to crassitate. Slide BOU8/1, E.F. No. (L26); (l) Samarisporites sp. A, slide AOU7/3, E.F. No. (J44/2); (m) Samarisporites sp. A. This new taxon has a laevigate zona but the distal surface below the inner body has an ornament of spines around the margin that merge to form a reticulum over the inner body. Slide AOU7/3, E.F. No. (X28); (n) Tetrahedraletes medinensis (Strother & Traverse Reference Strother and Traverse1979 emend. Wellman & Richardson Reference Wellman and Richardson1993), slide BOU9/1, E.F. No. (C35/1); (o) Breconisporites sp., slide BOU3/1, E.F. No. (C32/4); (p) Camptozonotriletes caperatus McGregor, Reference McGregor1973, slide BOU5/1, E.F. No. (T28/2); (q) Laevolancis divellomedium (Chibrikova) Burgess & Richardson, Reference Burgess and Richardson1991, slide BOU11/1, E.F. No. (H33). Scale bar=20 µm.
Table 3 Identified spore taxa and their occurrence

2.4. Age determination
The described spore assemblages are correlated with the Emphanisporites annulatus–Camarozonotriletes sextantii Spore Assemblage Biozone (AS SAB) of Richardson & McGregor (Reference Richardson and McGregor1986), based on the presence of one of the nominal species (E. annulatus) and the general characteristics of the assemblage in terms of morphotypes present and their general abundances. The preceding and succeeding zones are precluded by, amongst other observations, the presence of E. annulatus and the absence of spores with grapnel-tipped spines, respectively. The age of the AS Spore Assemblage Biozone is Early Devonian Emsian (but not earliest Emsian nor latest Emsian). In the spore zonation scheme of Streel et al. (Reference Streel, Higgs, Loboziak, Riegel and Steemans1987), the AS Spore Assemblage Biozone is equivalent to the annulatus–bellatulus and foveolatus–dubia Oppel Zones (AB OZ and FD OZ). Both nominal species of the AB OZ are present: E. annulatus and B. bellatulus. Neither of the nominal or other characteristic species of the FD Oppel Zone is present. Thus, the spore assemblage may belong to the AB OZ, constraining the age to early Emsian. However, one must be cautious when applying the Ardennes–Rhenish scheme to the ‘Lower Old Red Sandstone’ of Scotland because: (i) there is a large distance between northern Scotland and the Ardennes–Rhenish region; (ii) the two regions represent different facies (intermontaine basin versus coastal plain); and (iii) many of the taxa present in the diverse spore assemblages of the coastal plain deposits of the Ardennes-–Rhenish region are not present in the depauperate spore assemblages of the inland intermontane deposits of Scotland (see below).
2.5. Palynofacies analysis and palaeoenvironmental interpretation
The palynomorph assemblages contain only land-derived forms. These consist predominantly of disarticulated plant parts (cuticles, including some with stomata, and conducting tissues) and spores derived from the plants. There are also rare fungal hyphae, rare banded tubes similar to those produced by nematophytes, and a single fragment interpreted as the organic remnants of a fish scale. The nature of the palynomorph assemblages suggests accumulation in a non-marine environment. This strongly supports previous interpretations of these deposits, based on sedimentological observations, as accumulating in typical ‘Lower Old Red Sandstone’ terrestrial fluviatile–lacustrine floodplain environments (e.g., Trewin Reference Trewin, Trewin and Hurst1993, Reference Trewin, Trewin and Hurst2009a).
3. Geological implications
Understanding the Caledonian Orogeny and determining how and when the various Scottish terranes were emplaced are long standing problems (e.g. Trewin & Thirlwall Reference Trewin, Thirlwall and Trewin2002). The new biostratigraphical information suggests that the age of the ‘Basement Group’ of the Berriedale Outlier is Early Devonian Emsian (but not earliest Emsian nor latest Emsian), with an early Emsian age most likely. This has a number of implications relevant to these debates.
The Caledonian Orogeny involved the closing of the Iapetus Ocean and collision between Avalonia, Laurentia and Baltica. During closure, there was basement thrusting, uplift and exhumation due to deep weathering. Closure was oblique, with significant sinistral strike-slip movement along a number of NE-trending faults, including the Great Glen Fault System (e.g., McClay et al. Reference McClay, Norton, Coney and Davis1986; Norton et al. Reference Norton, McClay and Way1987). During the Late Silurian–Early Devonian ‘Lower Old Red Sandstone’, facies deposits began to be deposited on the southeast margin of the newly-created Old Red Sandstone continent. Modern theories regarding Devonian sedimentation in the Orcadian Basin revolve around three main models (summarised in Trewin Reference Trewin, Trewin and Hurst2009b): (i) sedimentation was controlled by extensional collapse of the Caledonian crust and occurred in half-grabens, with unconformities resulting from footwall uplift (e.g. Norton et al. Reference Norton, McClay and Way1987); (ii) strike-slip was important, with extension occurring in transtensional basins, with sediment deriving from erosion of transpressional highs (e.g., Trewin Reference Trewin1989); (iii) a combination of the above, with extensional collapse superimposed on compressional structures relating to earlier strike-slip (e.g., Underhill & Brodie Reference Underhill and Brodie1993).
The new early Emsian (ca. 403–407 Ma) age provides a constraint for the commencement of sedimentation in the Orcadian basin and, in particular, on the Northern Highlands to the northwest of the Great Glen Fault System. The Helmsdale Granite has been dated as ca. 420 Ma (Pidgeon & Aftalion Reference Pidgeon, Aftalion, Bowes and Leake1978). Thus, the new age constraint indicates a long period of ca. 15 million years for granite weathering on the post-Caledonian land surface (based on ages provided in Gradstein et al. Reference Gradstein, Ogg, Schmitz and Ogg2012). Trewin (Reference Trewin, Trewin and Hurst2009b) estimates that at least 3 km of rock was weathered, unroofing the Helmsdale Granite and Moine metamorphics. Mendum & Noble (Reference Mendum, Noble, Law, Butler, Holdsworth, Krabbendam and Strachan2010) estimate ca. 12–15 km of uplift, with exhumation rates of between 2 and 4 mm per annum, based on dating of events associated with the Rosemarkie Inlier. Interestingly, there are no ‘Lower Old Red Sandstone’ deposits identified as older than early Emsian associated with the Orcadian Basin, although beyond the Orcadian Basin on the Grampian Highlands there are ‘Lower Old Red Sandstone’ deposits of Pragian–?earliest Emsian age in the Rhynie Outlier (Wellman Reference Wellman2006), of Lochkovian age at Glen Coe (Wellman Reference Wellman1994) and of latest Silurian–earliest Devonian age at Lorne (Wellman & Richardson Reference Wellman and Richardson1996). These older deposits probably represent an early onset of ‘Lower Old Red Sandstone’ sedimentation on the post-Caledonian land surface, and it is possible that somewhere in the Orcadian Basin, volcanic collapse structures and half-grabens preserving pre-Emsian deposits are concealed by overstep.
The new age also suggests that in the Berriedale Outlier, the unconformity between ‘Lower Old Red Sandstone’ (early Emsian) and ‘Middle Old Red Sandstone’ (late Eifelian) deposits represents an estimated 16 million years in duration. Presumably, however, an unknown thickness of ‘Lower Old Red Sandstone’ sequence could have been removed during this time. Thus, the basin-wide ‘Lower Old Red Sandstone’/‘Middle Old Red Sandstone’ unconformities/disconformities are likely variable in duration and are almost certainly due to minor, local rather than large-scale, regional tectonism. However, Mendum & Noble (Reference Mendum, Noble, Law, Butler, Holdsworth, Krabbendam and Strachan2010) suggest that deformation in ‘Lower Old Red Sandstone’ deposits on the Northern Highlands (e.g., at Struie) may have resulted from a Mid Devonian Acadian event.
The juxtapositioning of Orcadian Basin deposits across the Great Glen Fault System suggests that there was little-or-no syn-depositional strike-slip movement along this fault during Devonian ‘Old Red Sandstone’ deposition (Rogers et al. Reference Rogers, Marshall and Astin1989; Marshall & Hewett Reference Marshall, Hewett, Evans, Graham, Armour and Bathurst2003). Rogers et al. (Reference Rogers, Marshall and Astin1989) suggest that the early history of the Great Glen Fault System included late Caledonian sinistral motion, but that this had ceased by the late Emsian and the fault system was not active transcurrently during ‘Old Red Sandstone’ deposition. The new age date provided herein helps refine the dating of movement.
Adjacent offshore ‘Lower Old Red Sandstone’ deposits are only present southeast of the Great Glen Fault System (Marshall & Hewett Reference Marshall, Hewett, Evans, Graham, Armour and Bathurst2003). These accumulated in extensional half-graben systems and are genetically related to onshore deposits of the Grampian Highlands, such as those of the Turriff Basin (Richards Reference Richards1985; Sweet Reference Sweet1985). These deposits are highly interesting because they are surprisingly thick (in excess of 1000 m in thickness in boreholes 12/27-1, 12-27-2 and 12/29-2). Marshall & Hewitt (Reference Marshall, Hewett, Evans, Graham, Armour and Bathurst2003) suggest that different dynamics in different half-graben systems were responsible for such significant variation in thickness of ‘Lower Old Red Sandstone’ deposits in the Orcadian Basin.
4. Palaeobotanical implications
Coeval spore assemblages are known from elsewhere in Scotland and adjacent areas: (i) the Midland Valley deposits of the Strathmore Basin (Richardson Reference Richardson1967; Ford Reference Ford1971; Wellman Reference Wellman2010); (ii) the lowland floodplain deposits of southern England (Chaloner Reference Chaloner1963; Richardson & Rasul Reference Richardson and Rasul1978a, Reference Richardson and Rasulb), although these are probably slightly younger and equate to the FD OZ of mid Emsian age of the spore zonation of Streel et al. (Reference Streel, Higgs, Loboziak, Riegel and Steemans1987); (iii) the coastal plain deposits of the Ardennes–Rhenish region (Streel et al. Reference Streel, Higgs, Loboziak, Riegel and Steemans1987). Although all three locations share many common spore taxa, indicating similarities in the flora, both the Scottish and southern English assemblages appear rather depauperate compared to those from the Ardennes–Rhenish region, and indeed from other localities on the Old Red Sandstone continent such as Gaspé, eastern Canada (McGregor Reference McGregor1973, Reference McGregor1977). AS SAB/AB OZ spore assemblages from both the Ardennes–Rhenish region and Gaspé yield in the region of 50 taxa. As discussed by Richardson & Rasul (Reference Richardson and Rasul1978b), these differences most likely reflect ecological, or perhaps facies, effects.
The absence from Scotland of several well-known and distinctive species is interesting in that it suggests that these plants may not have inhabited the inland uplands. Notable absences are Acinosporites lindlarensis and Camarozonotriletes sextantii. Acinosporites lindlarensis is the spore of the lycopsid Leclercqia (Richardson et al. Reference Richardson, Bonamo and McGregor1993; Wellman et al. Reference Wellman, Gensel and Taylor2009) and C. sextantii is the microspore of the unclassified, probably heterosporous plant, Chaleuria (Andrews et al. Reference Andrews, Gensel and Forbes1974; McGregor & Camfield Reference McGregor and Camfield1976). However, C. sextantii is known to have patchy distribution (see discussion in Richardson & McGregor Reference Richardson and McGregor1986, pp 13–14).
Deposits belonging to the AS SAB/AB OZ from Scotland, southern England and the Ardennes–Rhenish region have all yielded plant megafossils. At present, the Scottish flora is limited to the zosterophyll Sawdonia ornata and lycopsid Drepanophycus spinaeformis (Rayner Reference Rayner1983, Reference Rayner1984), and the southern English flora to the zosterophyll S. ornata and trimerophyte Dawsonites arcuatus (Chaloner et al. Reference Chaloner, Hill and Rogerson1978). It is evident that the Ardennes–Rhenish plant assemblages are much more diverse (e.g. Kräusel & Weyland Reference Kräusel and Weyland1930; Stockmans Reference Stockmans1939; Leclercq Reference Leclercq1942; Schweitzer Reference Schweitzer1983; Fairon-Demaret Reference Fairon-Demaret1985; Gerrienne Reference Gerrienne1993) and include rhyniophytes, zosterophylls (including the possible barinophyte Krithodeophyton), lycopsids, trimerophytes and a possible sphenopsid (Estinnophyton) and cladoxyle (Foozia). To some extent, this almost certainly reflects collector bias, with the Ardennes–Rhenish region extremely well exposed and highly researched, and the southern English sequences only exposed in the subsurface. However, the Scottish deposits are reasonably well explored (e.g., Henderson Reference Henderson1932; Lang Reference Lang1932; Rayner Reference Rayner1983, Reference Rayner1984) and it does seem likely that the floras of the intermontaine basins of Scotland were less diverse than those from the floodplains. Nonetheless, in this respect the lack of diversity of both spores and plant megafossils in southern English floodplain deposits is puzzling. It is noteworthy that spore assemblages from all of these regions are dominated by Retusotriletes spp. and Apiculiretusispora spp. that were most likely produced in the main by various zosterophylls, lycopsids and trimerophytes. However, many distinctive spore morphotypes are still of unknown affinity, suggesting that plant megafossil diversity is not entirely reflective of the regional floras as a whole.
5. Acknowledgements
This research was funded by the NERC.









