INTRODUCTION
Many lakes in tropical Africa are known for their endemic fishes and for environmental histories that are preserved in the sediments beneath them. The topography of the West African tropics is generally less conducive to the formation of lakes than the East African rift region, but Lake Bosumtwi, Ghana, which formed after a bolide impact ca. 1 million yr ago (Koeberl et al., Reference Koeberl, Bottomley, Glass and Storzer1997; Peck et al., Reference Peck, Green, Shanahan, King, Overpeck and Scholz2004), has yielded detailed records of late Quaternary climate (Russell et al., Reference Russell, Talbot and Haskell2003; Peck et al., Reference Peck, Green, Shanahan, King, Overpeck and Scholz2004; Shanahan et al., Reference Shanahan, Overpeck, Anchukaitis, Beck, Cole, Dettman, Peck, Scholz and King2009). An additional 39 crater lakes of volcanic or phreatic origin occur in and around the highlands of western Cameroon (Kling, Reference Kling1987). Lowland crater lake Barombi Mbo, also ca. 1 million yr old, is home to 11 endemic species of cichlid fish (Schliewen et al., Reference Schliewen, Tautz and Pääbo1994) and has yielded a detailed pollen record of climate and rain forest history spanning the last 33 ka (Trewavas et al., Reference Trewavas, Green and Cobet1972; Cornen et al., Reference Cornen, Bande, Giresse and Maley1992; Giresse et al., Reference Giresse, Maley and Brenac1994; Maley and Brenac, Reference Maley and Brenac1998; Schliewen and Klee, Reference Schliewen and Klee2004; Lebamba et al., Reference Lebamba, Vincens and Maley2012). The geochemical and microfossil records of Bosumtwi and Barombi Mbo represent the most widely cited paleoenvironmental histories in equatorial West Africa, and the endemic cichlids of Barombi Mbo have been the focus of intensive studies by evolutionary biologists (Trewavas et al., Reference Trewavas, Green and Cobet1972; Schliewen et al., Reference Schliewen, Tautz and Pääbo1994, Reference Schliewen, Rassmann, Markmann, Markert, Kocher and Tautz2001; Schliewen and Klee, Reference Schliewen and Klee2004; Martin, Reference Martin2012; Musilová et al., Reference Musilová, Indermaur, Nyom, Tropek, Martin and Schliewen2014; Martin et al., Reference Martin, Cutler, Friel, Dening, Coop and Wainwright2015). However, other lesser known lakes from this region are also potentially informative.
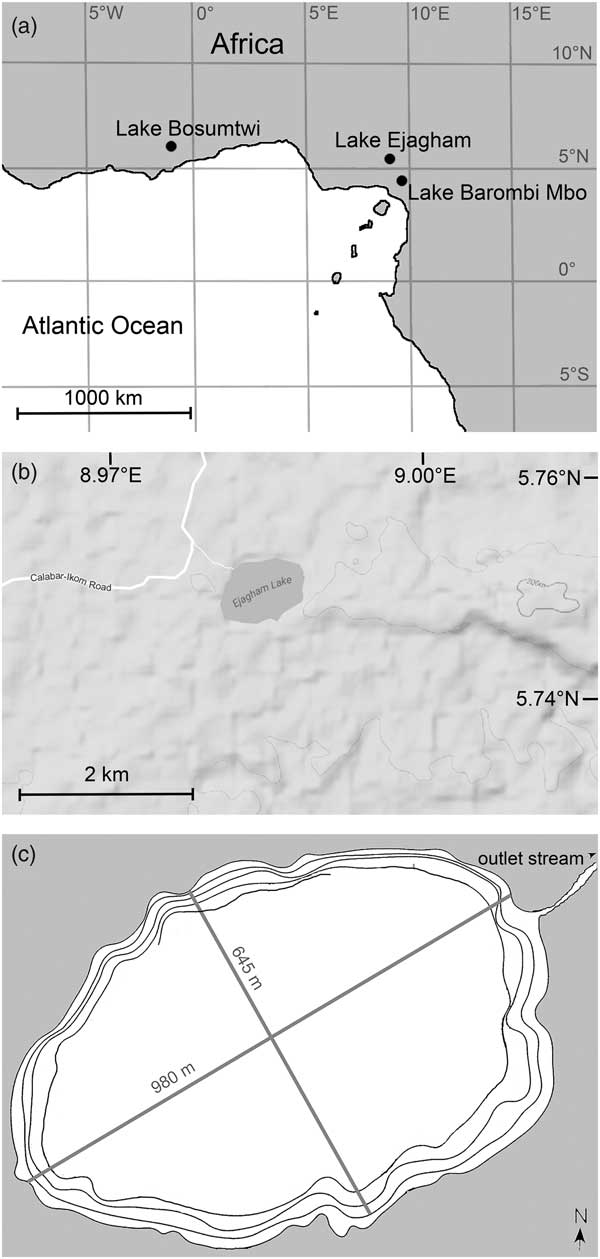
One of those additional water bodies is Lake Ejagham, Cameroon (Fig. 1). Unlike most West African lakes, Ejagham is of enigmatic origin, and it supports two endemic radiations of endemic cichlids (Schliewen et al., Reference Schliewen, Rassmann, Markmann, Markert, Kocher and Tautz2001). Some sources have suggested that it is a chemical solution lake (Schliewen et al., Reference Schliewen, Rassmann, Markmann, Markert, Kocher and Tautz2001), but the late Daniel A. Livingstone (DAL) speculated (personal communication, 1985) with the first author that it may be a bolide impact crater, as is Bosumtwi. He collected two cores (EJ1 and EJ2) from the center of the lake in 1985 in order to reconstruct local climate history and investigate the environmental context for the evolution of the cichlids. However, the cores were not dated, and the only analyses performed on them since 1985 were measurements of organic matter and carbonate content on the longer core, EJ2. Upon the closure of DAL’s laboratory at Duke University in 2015, the first author transferred both cores to Paul Smith’s College, New York, for subsampling and analysis.
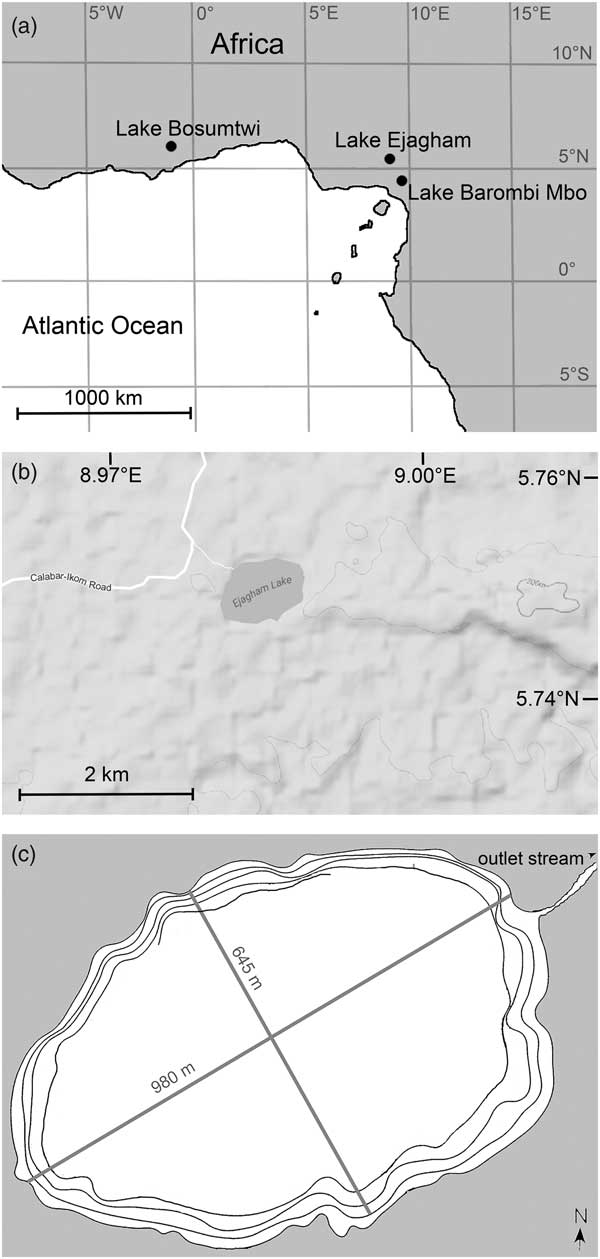
Figure 1 Site map. (a) Regional map of West Africa. (b) Detail of the vicinity of Lake Ejagham. (c) Bathymetric map of Lake Ejagham. Each contour represents 3 m. Only partial representation of the 12 m contour is currently available.
In this article, we present the results of radiocarbon, geochemical, mineralogical, and diatom analyses on the EJ1 and EJ2 cores that offer new insights into the age and origin of Lake Ejagham and its endemic fishes. We use those data to demonstrate that both the lake and its endemic fish species are no more than 9000 yr old. We also compare the EJ1 diatom record to a high-resolution oxygen isotope record from Bosumtwi, which represents hydroclimate variability associated with the West African monsoon system during the Holocene. Finally, we review the possible origins of the lake and assess the evidence for an extraterrestrial bolide impact.
Study site
Lake Ejagham is located in a sparsely populated region of evergreen rain forest near the village of Eyumojok in southwestern Cameroon (Fig. 1a; surface area 0.5 km2; ~1 km maximum diameter; elevation ~200 m above mean sea level; 5°45.00′N, 8°59.20′E; Schliewen et al., Reference Schliewen, Rassmann, Markmann, Markert, Kocher and Tautz2001; Dunz and Schliewen, Reference Dunz and Schliewen2010). The West African summer monsoon system produces a rainy season in the region between April and October with mean local rainfall of ~3000 mm (Offem et al., Reference Offem, Ayotunde, Ikpi, Ada and Ochang2011). Air temperatures in the region can range from ~20°C to ~33°C, but monthly mean temperatures are typically within the 25°C–28°C range (ftp://ftp.atdd.noaa.gov/pub/GCOS/WMO-Normals/TABLES/REG__I/KM/64890.TXT [accessed February 6, 2017]).
Lake Ejagham lies on the eastern margin of the Benue Trough, a Mesozoic depositional basin atop crystalline Precambrian basement adjacent to the northeast-to-southwest-trending Cameroon Volcanic Line, where most of the region’s crater lakes are located (Kling, Reference Kling1987; Tokam et al., Reference Tokam, Tabod, Nyblade, Julià, Wiens and Pasyanos2010). Two major basalt formations associated with the Mamfe basin, a rift extension of the Benoue Trough, lie several miles to the east of Lake Ejagham and are 65–82 Ma and 9–11 Ma old (Fairhead et al., Reference Fairhead, Okerere and Nnange1991). The lake itself lies on Cretaceous fluviolacustrine sedimentary deposits within the Mamfe basin that obscure the underlying geology (Ndougsa-Mbarga et al., Reference Ndougsa-Mbarga, Manguelle-Dicoum, Campos-Enriquez and Atangana2007; Ephraim, Reference Ephraim2012; Njoh et al., Reference Njoh, Nforsi and Datcheu2015).
Lake Ejagham is shallower than most Cameroon crater lakes (maximum depth 17 m; Kling, Reference Kling1987). It is oval in shape with a low rim (~10 m; Kling, Reference Kling1987), a single small surface outlet, a broad, flat bottom of fine-grained organic sediment offshore, and a narrow belt of sandy shallows that dip steeply from the shoreline (Fig. 1c; Offem et al., Reference Offem, Ayotunde, Ikpi, Ada and Ochang2011). The pH of the lake water is circumneutral, and its moderately elevated conductivity of 86–120 µS/cm measured in 2008 and 2009 might reflect inputs from saline springs that are common in the Mamfe basin (Eseme et al., Reference Eseme, Abanda, Agyingi, Foba-Tendo and Hannigan2006; Offem et al., Reference Offem, Ayotunde, Ikpi, Ada and Ochang2011; Njoh et al., Reference Njoh, Nforsi and Datcheu2015). Local soils are sandy, and the bedrock of the watershed includes lenses of salt and marl interbedded with shales, sandstones, and other sedimentary rocks (Ephraim, Reference Ephraim2012; Njoh et al., Reference Njoh, Nforsi and Datcheu2015). The terrain surrounding the lake is generally flat and devoid of obvious karst features (Fig. 1b).
In 2008 and 2009, the predominant phytoplankton during both wet and dry seasons were reported to be cyanobacteria including Oscillatoria and Microcystis with lesser amounts of Cyclotella, Melosira, Rhizosolenia, Synedra, and Tabellaria diatoms (Offem et al., Reference Offem, Ayotunde, Ikpi, Ada and Ochang2011). The lake also supports five endemic cichlid species belonging to two distinct African cichlid lineages, Sarotherodon and Coptodon (Schliewen et al., Reference Schliewen, Rassmann, Markmann, Markert, Kocher and Tautz2001; Dunz and Schliewen, Reference Dunz and Schliewen2010; Neumann et al., Reference Neumann, Stiassny and Schliewen2011). In addition, a subspecies of killifish, Aphyosemion gardneri lacustri, has been described from the lake along with two widespread species, the killifish Procatopus aberrans and the cyprinid Barbus cf. callipterus (Schliewen et al., Reference Schliewen, Rassmann, Markmann, Markert, Kocher and Tautz2001). In 2000–2001, a local town council member introduced large numbers of catfish (and possibly other species), resulting in a breeding population of Parauchenoglanis sp. that was still present in the lake as of 2016 (Martin, Reference Martin2012; Martin, C.H., personal observation).
METHODS
In 1985, DAL obtained two rod-driven piston cores, EJ1 (2.5 m long) and EJ2 (6 m long), from a raft anchored in the approximate center of the lake. No indication of overlaps or gaps between the core subsections was recorded in DAL’s field notes. The steel coring tubes were capped and transported to Duke University where the two segments of core EJ2 were subsampled in 10 cm increments during the 1980s. In 2015, the EJ2 core was found to be severely desiccated and lacking structural integrity. The sediments were therefore unsuitable for diatom analysis, but subsamples from the lowermost meter were removed for radiocarbon dating and mineralogical analysis.
Core EJ1 was collected in three steel core tubes that remained unopened until 2016, when they were extruded by the first two authors at Paul Smith’s College and subsampled in 10 cm increments for diatom analysis and radiocarbon dating.
In addition to the two EJ cores, two gravity cores of <1 m length were collected in 1985 by George Kling and JCS from DAL’s coring raft and stored at Duke University along with the other cores. The diatom assemblages in the top and bottom organic sediments from those cores were examined for this study in 2016, as were surface samples that were hand collected from the shallow littoral zone in 2016 by CHM.
Radiocarbon age model
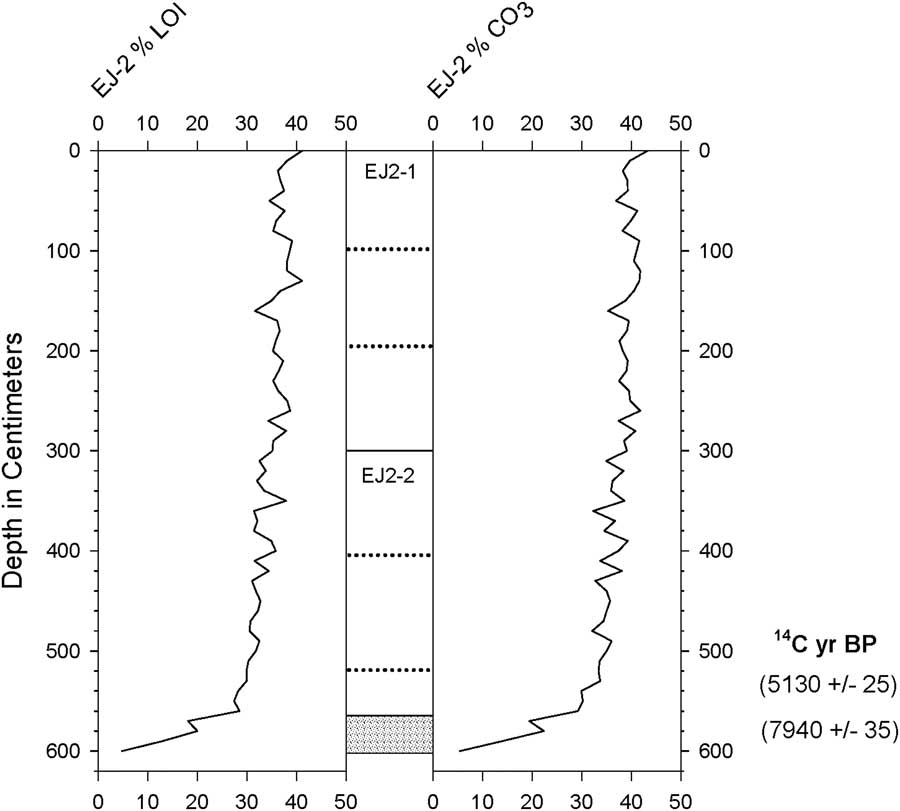
Two radiocarbon ages were obtained by accelerator mass spectrometry on subsamples from the lowest meter of EJ2 (Table 1, Fig. 2). Because the core was in poor condition, the stratigraphic position of the upper subsample of fine-grained sediment could only be estimated (524–528 cm; described here as 526 cm). At the base of the core (580 cm), a fragment of wood was also removed from coarse-grained sandy sediment for dating (Fig. 3).
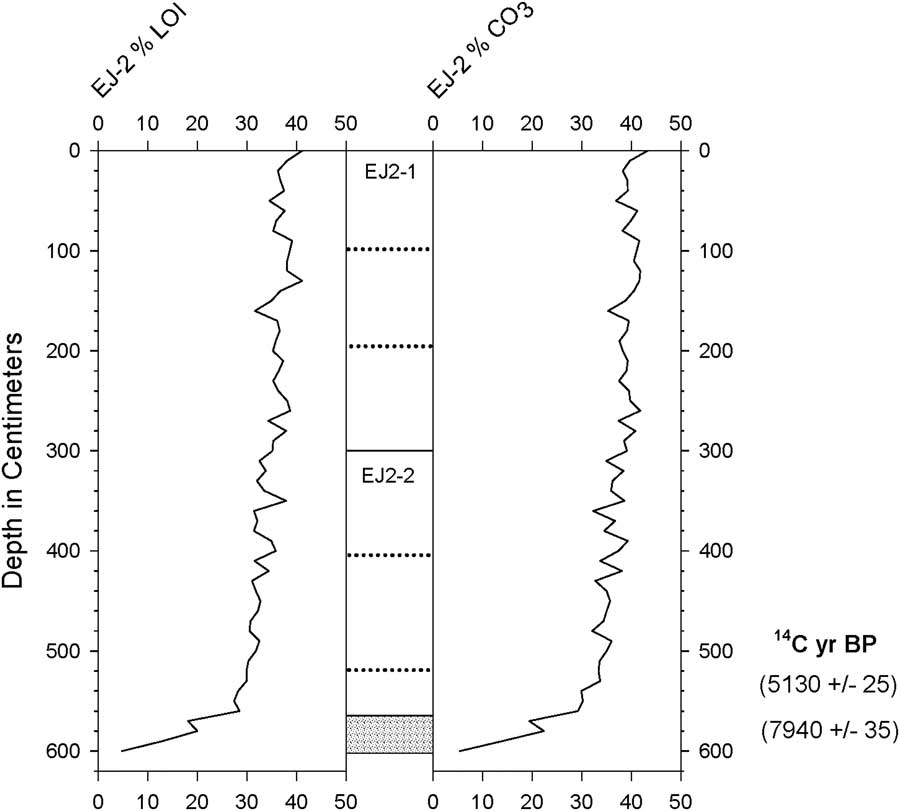
Figure 2 Estimated organic matter and carbonate content (by percent weight loss on ignition [LOI]) of core EJ2. Gray shading indicates coarse-grained, disaggregated sediment.

Figure 3 (color online) Coarse-grained, disaggregated sediments containing wood fragments at the base of core EJ2.
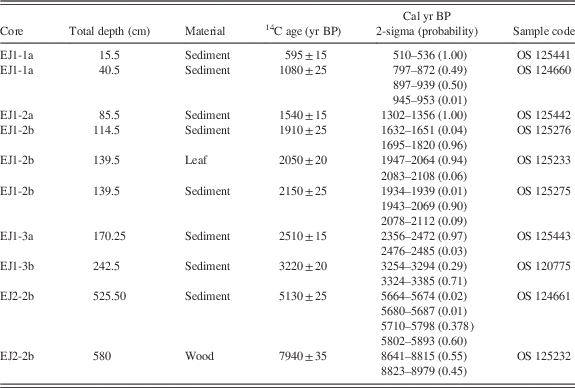
Table 1 Radiocarbon and calendar age ranges for samples from Lake Ejagham cores EJ1 and EJ2. All calibrated ages were determined with CALIB version 7.1 (Stuiver and Reimer, Reference Stuiver and Reimer1993) after subtracting 100 yr from the radiocarbon ages of all bulk sediment samples (see text).
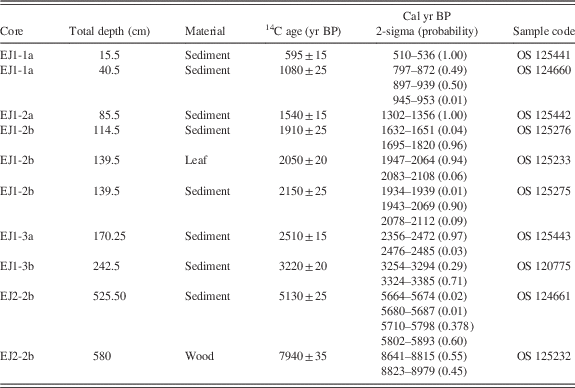
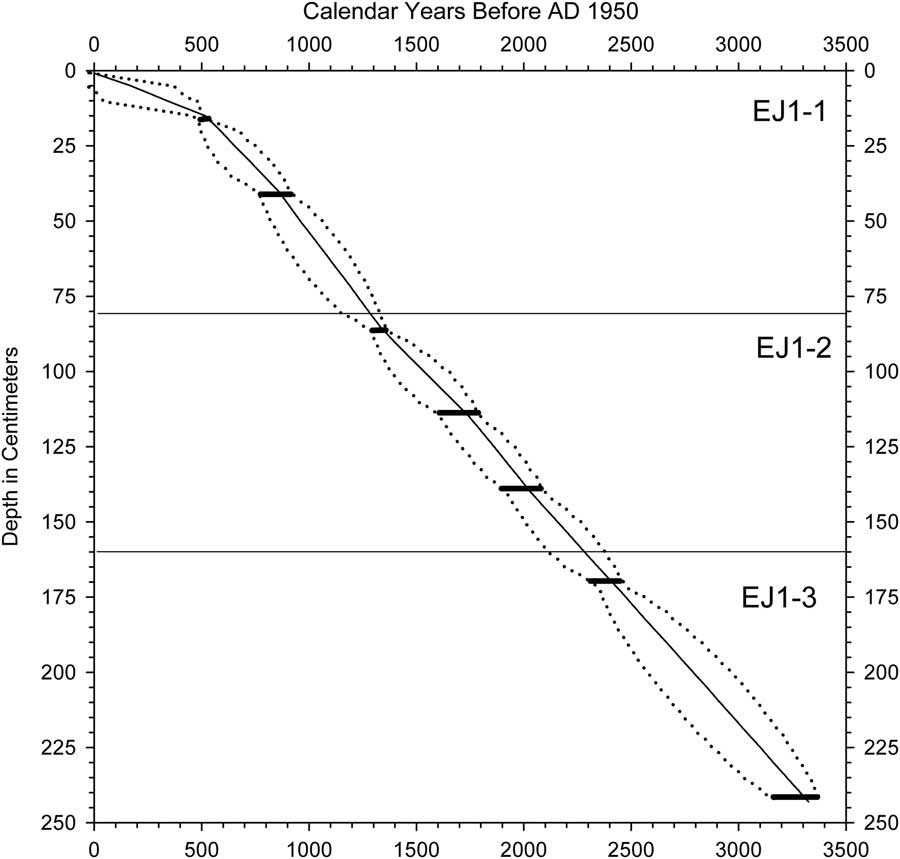
Eight radiocarbon ages were obtained for bulk sediments and a leaf in shorter core EJ1 (Table 1, Fig. 4). Radiocarbon ages were converted to 2-sigma calendar year ranges with CALIB version 7.1 (Stuvier and Reimer, 1993). The discontinuous nature of the EJ1 core because of multiple sampling drives limits the reliability of a composite age model for it, but a likely age-depth relationship in EJ1 was constructed using Bayesian age modeling with the Bacon modeling package in R (Blaauw and Christen, Reference Blaauw and Christen2011). A provisional age-depth relationship using the mean values of the Bacon age model (Fig. 5) was applied to the diatom record for comparison to a high-resolution oxygen isotope record from Lake Bosumtwi (Shanahan et al., Reference Shanahan, Overpeck, Anchukaitis, Beck, Cole, Dettman, Peck, Scholz and King2009; Fig. 6; see Discussion).

Figure 4 Thin-section photomicrographs of mineral grains from core EJ2. Scale bar represents 50 μm. (a) Sample from 578–582 cm depth under crossed-polarized light. Note the angular nature of the grains. (b–d) A single quartz grain from 566–569 cm depth showing possible impact effects (see text). This polycrystalline, subangular quartz grain contains curviplanar fractures with abundant submicron fluid inclusions. (b) Under crossed-polarized light. (c) Toasting, a browning effect, is evident along the fractures in the plane light view. Undulose extinction is evident in the cross-polarized view. (d) Detail from inset in panel c, showing apparent toasting. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
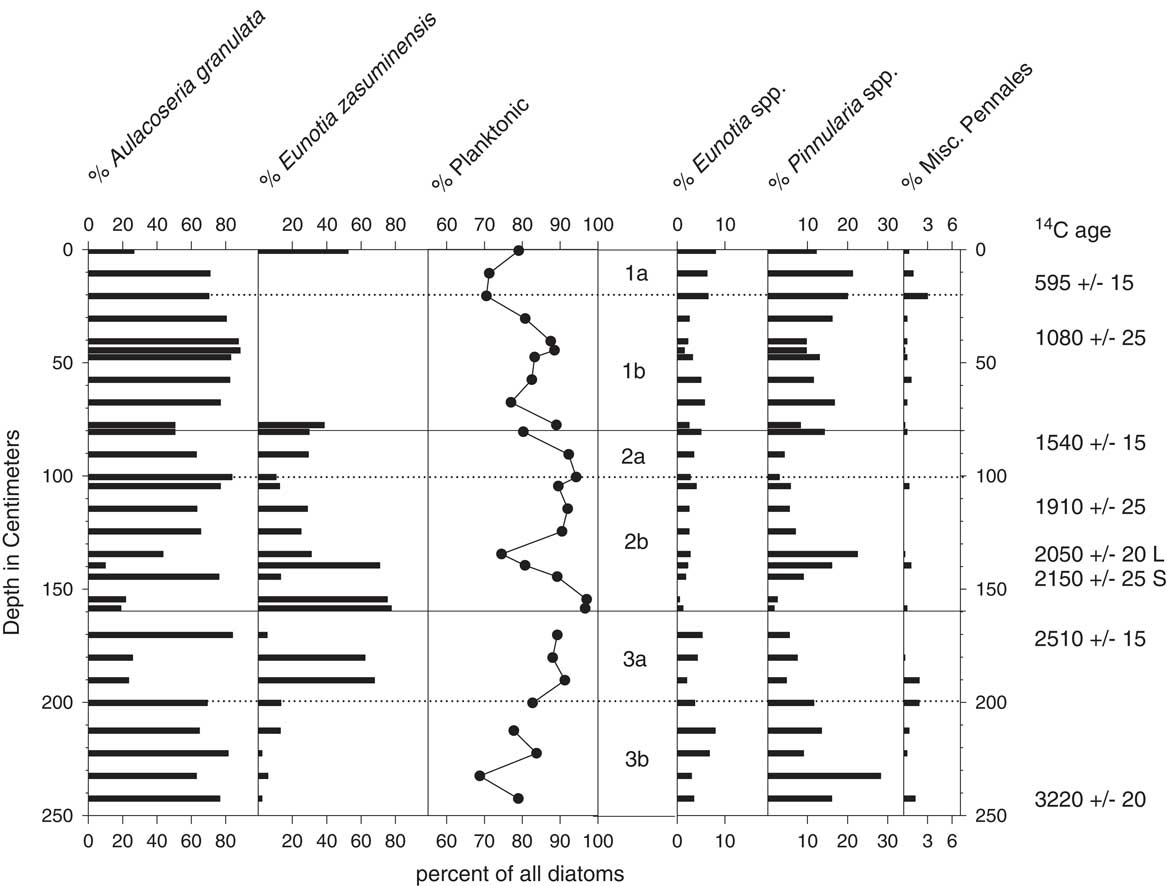
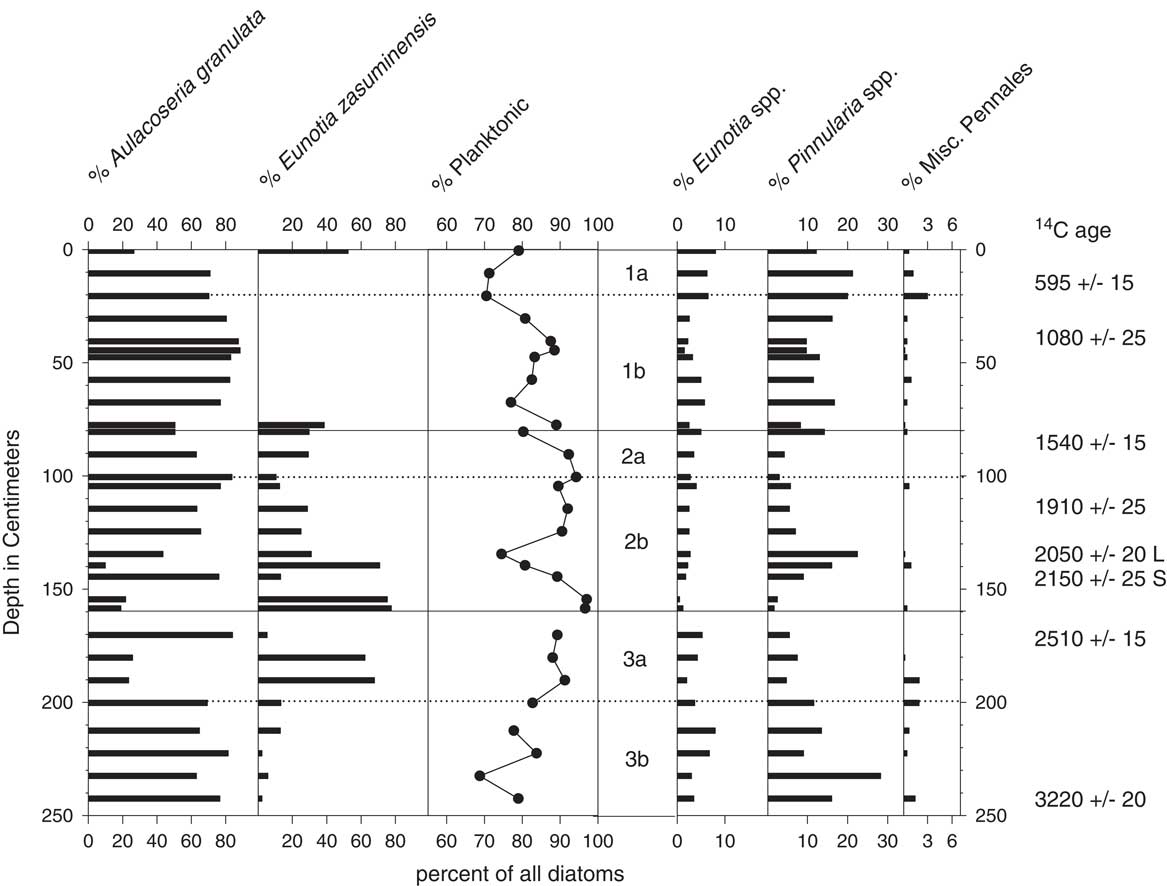
Figure 5 Diatom stratigraphy for core EJ1. Middle panel: the three main subsections of the core are delineated by solid lines; dotted lines indicate locations where the subsections were cut into smaller segments for storage. Radiocarbon ages to the right include two ages obtained for a leaf (L) and bulk organic sediment (S) from the same depth in the core.
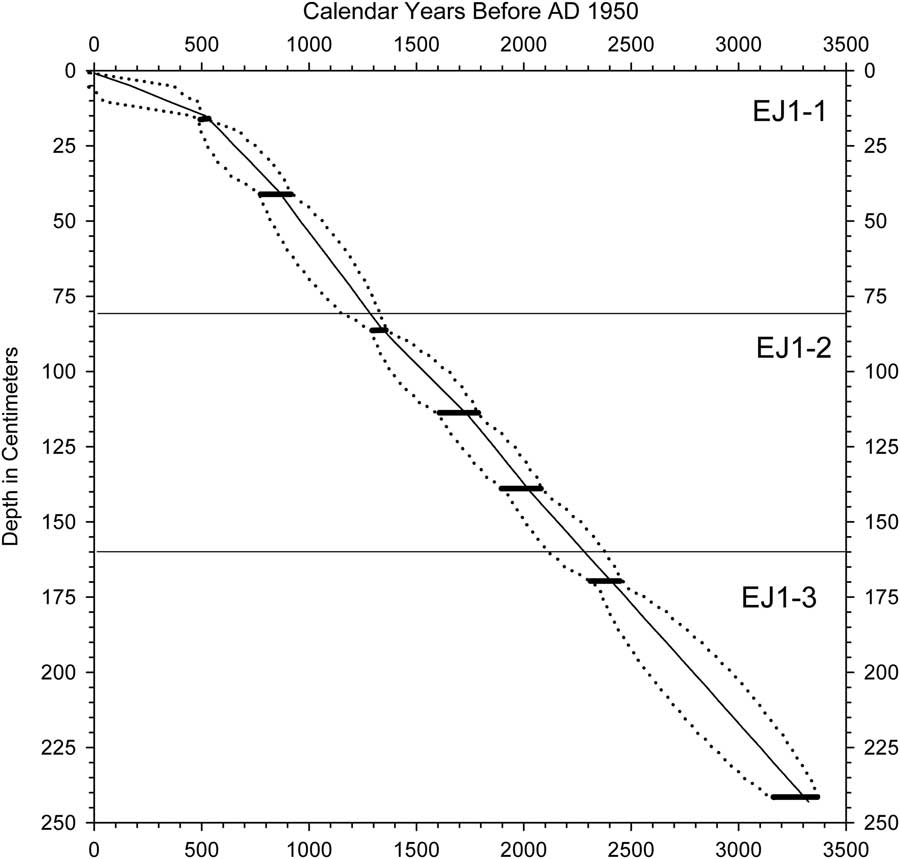
Figure 6 Age model for core EJ1 developed for comparison to the Bosumtwi oxygen isotope record (see text). Horizontal bars indicate radiocarbon age ranges after conversion to calendar years. Dotted lines represent maximum and minimum age limits. Solid line represents the mean ages, which were used to construct an age model for the EJ1 diatom record.
Diatom analyses
Diatom assemblages in EJ1 were sampled by mixing raw sediment with distilled water and applying the slurry to coverslips, which were affixed to glass slides with Permount mounting medium. The fine texture of the dispersed sediment particles and lack of coagulated organic matter made the diatom valves plainly visible without chemical treatment, thereby reducing the risk of damage to the frustules, which were already highly fragmented in most samples. At least 400 valves were enumerated per sample at 1000× magnification by KA, using taxonomy based primarily on Gasse (Reference Gasse1986) and Lange-Bertalot et al. (Reference Lange-Bertalot, Bak and Witkowski2011).
Sedimentologic analyses
Organic matter and carbonate contents for sediments in EJ2 were estimated by weight loss on ignition at 500°C and 900°C, respectively (Sutherland, Reference Sutherland1998), and the data were recorded in a notebook that was retrieved along with the cores by JCS upon the closure of DAL’s lab in 2015.
Mineralogical analyses were performed on two samples from the 566–569 cm and 578–582 cm intervals in core EJ2 (Fig. 2). Mineral content, grain shapes, surface textures, and grain sizes were determined under a binocular dissecting microscope before thin sectioning. Roundness of the grains was categorized as rounded, subrounded, subangular, or angular (Powers, Reference Powers1953). Both samples were then separated into two subsamples according to size by handpicking the coarse sand grains and granules (1–4 mm diameter; size terms of Wentworth, Reference Wentworth1922), leaving behind the smaller grains mixed with brown-gray clay and silt matrix. The subsamples were placed into small plastic containers, which were then filled with epoxy for thin sectioning. Thin sections of the embedded samples (30 µm thickness) were mounted on glass slides and examined under a petrographic microscope equipped with cross polarizers. Minerals were identified in thin sections using standard petrographic techniques. The quartz and feldspar grains in the thin sections were examined by DTK and LWP to determine their provenance or origin and to look for impact-related shock-metamorphic effects or evidence of melting.
RESULTS
Radiocarbon dating
The wood fragment from the 580-cm interval at the base of core EJ2 yielded a 2-sigma age between 8641 and 8979 cal yr BP (Table 1, Fig. 2). This result establishes a maximum age of ca. 9 ka for the lake. Fine-grained sediment at an estimated depth of 526 cm in the core yielded an age between 5664 and 5893 cal yr BP (Table 1). If the core top included the mud-water interface, then the mean temporal span of a centimeter increment in the upper 526 cm of the core was ca. 11 yr. The ca. 3 ka age difference between the upper and lower dated subsamples suggests that the basal half-meter of the record may have been discontinuous or compressed.
In contrast to EJ2, the sediments of core EJ1 were moist and apparently undisturbed. There was a difference of ca. 100 yr between the radiocarbon ages of a leaf and bulk sediment from the 34–35 cm interval in subsection EJ1-2b (114–115 cm composite depth; Table 1, Figs. 5 and 6). We therefore subtracted 100 yr from each radiocarbon age for bulk sediments before determining the calendar year age ranges (Table 1). The EJ1 core appears to represent the last ca. 3.3 ka of lake record (Fig. 4), with sediment age increments ranging from 6 to 16 yr/cm within the three subsections of the core (Figs. 4 and 5).
Diatom analyses
The dominant taxa in the EJ1 core were Aulacoseira granulata var. muzzanensis (F. Meister) Simonsen and small, humerus-shaped Eunotia zasuminensis (Cabejszekówna) Körner, which are planktonic to tychoplanktonic in both tropical and temperate lakes (Gasse, Reference Gasse1986; Kociolek, Reference Kociolek2005; Lange-Bertalot et al., Reference Lange-Bertalot, Bak and Witkowski2011; Nicholls and Carney, Reference Nicholls and Carney2011). They were also found in surface sediments from Cameroon crater lakes Nyos and Monoun in 1985 by JCS. Combined percentages of the two planktonic species fluctuated between 69% and 97%, with the highest values occurring within the 154–159 cm interval and the lowest values occurring within the lowermost 10 cm and uppermost 20 cm of the core (Fig. 5). Littoral pennate taxa belonged to the genera Eunotia, Pinnularia, and Navicula, in addition to lesser numbers of other Pennales.
The most common diatom taxa in the uppermost samples of the two gravity cores were A. granulata and E. zasuminensis, along with minor occurrences (<20%) of littoral taxa, primarily Pinnularia and bottom-dwelling Eunotia spp. The basal samples (core lengths not recorded) contained similar relative abundances of Aulacoseira and pennate diatoms, but E. zasuminensis was rare to absent.
The percentages of planktonic diatom taxa were much higher in midlake samples of the sediment-water interface than in samples from the littoral zone. Benthic Eunotia, including E. cf. pectinalis, were the most common taxa in sand-rich sediments collected by hand from the littoral zone in 2016, along with small percentages of A. granulata.
Sedimentologic analyses
The profiles of organic matter and carbonate content for EJ2 showed little variability (both averaging 35%–40%) except for the coarse-grained basal sediments below ~5.6 m depth, in which the values of both parameters were much lower (Fig. 2). The coarse particles were primarily composed of irregular mineral grains, but small fragments of wood were also present (Fig. 3).
Under microscopic examination, subsamples from 566–569 cm and 578–582 cm depths in core EJ2 were characterized by abundant gray-brown, clay-bearing detrital grains embedded in a gray-brown clay matrix (Fig. 3). The ratio of clay to detrital grains was approximately 50:50. The matrix was dark amber in thin section and contained about 30% finely macerated organic material, including some woody fragments but mainly consisting of small, opaque masses about 10–15 µm in diameter. The clay was almost entirely free of mica flakes.
Detrital grains consisted almost entirely of quartz and feldspar. The average detrital grain size was about 0.5 mm, which is on the boundary between medium and coarse sand (Wentworth, Reference Wentworth1922). The quartz grains were generally larger than the feldspars. There was a distinct bimodal size distribution among the detrital grains larger than 0.004 mm diameter. By volume, ~65% of the detrital grains were in the size range of 1 to 4 mm (very coarse sand to granule size), and the remainder were fine sand and silt (0.25 to 0.004 mm). The larger detrital grains commonly possessed a slight frosting or polishing on their surfaces, suggestive of chemical weathering and/or eolian activity prior to inclusion in the lake deposits.
Some of the quartz grains displayed undulose extinction and possible evidence of toasting, which is not typical evidence of shock metamorphism but could be indicative of extreme heat and pressure during an impact event (Fig. 4; see “Discussion”). Clear evidence of shock metamorphism was lacking, however, and no melt spherules or fragments were identified.
DISCUSSION
The transition from coarse, inorganic sediments to fine, organic mud near the base of core EJ2 represents the first development of lacustrine conditions at Lake Ejagham ca. 9 ka ago. The quartz and feldspar grains would have originated from weathering of Precambrian basement rocks (granites, gneisses, and migmatites; Fairhead et al., Reference Fairhead, Okerere and Nnange1991) and/or Lower Cretaceous sandstone formations that crop out in the Mamfe basin and Benue Trough (Fairhead et al., Reference Fairhead, Okerere and Nnange1991). The Precambrian source is more likely because the grains are generally angular to subangular (Fig. 4a) and the sandstones would be more likely to yield subrounded to rounded grains. Also, many of the quartz fragments are polycrystalline with interlocking grain domain boundaries, and some of them contain tiny needlelike rutile inclusions, both of which are indicative of a plutonic igneous source. The mineral grains in Lake Ejagham did not come from local basalts because basalt does not normally contain sizable quartz grains, and there are no basaltic rock fragments in the sediment.
The slightly greater radiocarbon age of bulk sediment compared with a leaf in core EJ1 may indicate contamination of the lake’s sediments or food web by local geologic deposits that include ancient carbon from local limestone, petroleum, and/or natural gas (Ndougsa-Mbarga et al., Reference Ndougsa-Mbarga, Manguelle-Dicoum, Campos-Enriquez and Atangana2007), as do relatively high carbonate contents in the EJ2 core (Fig. 2).
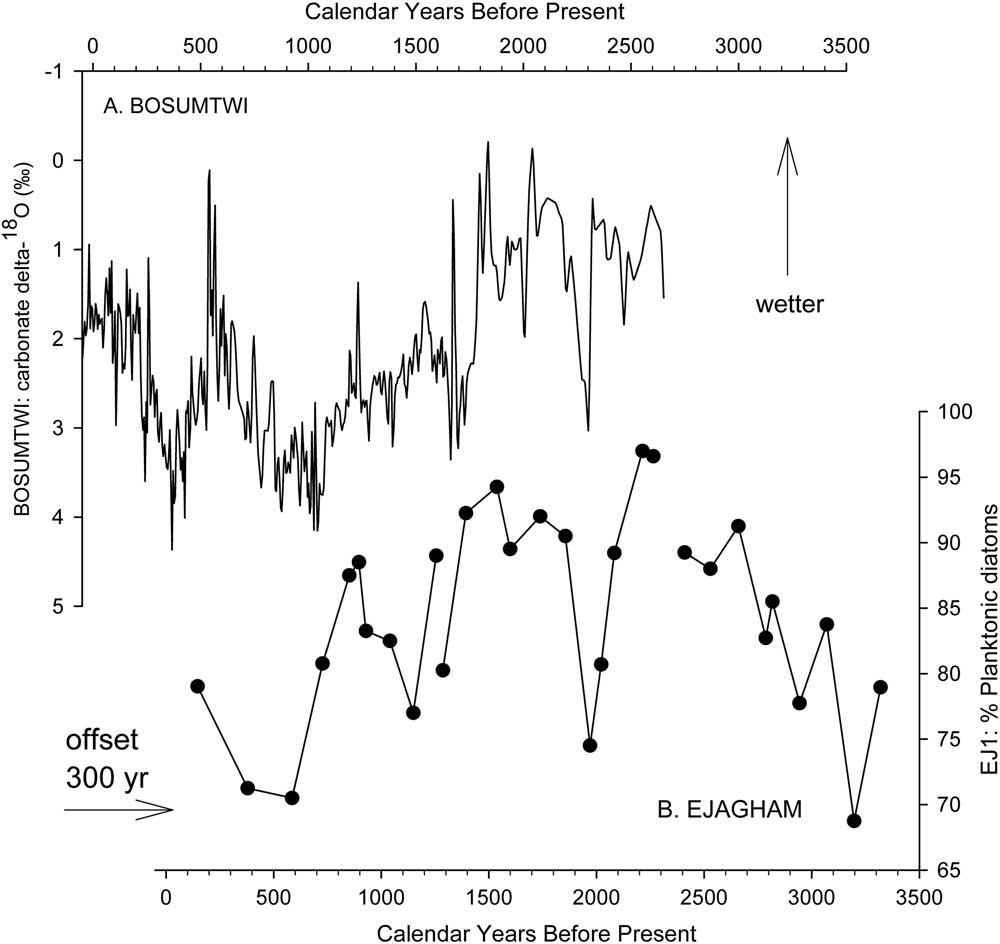
The association of high percentages of planktonic diatoms with deep-water habitats and/or wetter climatic conditions has been used for qualitative reconstructions of hydroclimate from fossil diatom assemblages with other lakes around the world (Stager et al., Reference Stager, Ryves, Cumming, Meeker and Beer2005, Reference Stager, Mayewski, White, Chase, Neumann, Meadows, King and Dixon2012, Reference Stager, Cumming, Laird, Garrigan-Piela, Pederson, Wiltse, Lane, Nester and Ruzmaikin2017; Laird et al., Reference Laird, Kingsbury and Cumming2010, Reference Laird, Kingsbury, Lewis and Cumming2011). In this study, high percentages of planktonic A. granulata and E. zasuminensis in the cores from the center of Lake Ejagham are taken to represent dilute, open-water conditions similar to those of today. The abundance of E. zasuminensis in the uppermost samples of EJ1 and its rarity in the interval immediately below them (Fig. 5) suggest that the youngest sediments of EJ1 were deposited shortly after relatively dry periods ca. 1000 or 500 yr ago, which were also registered at Lake Bosumtwi (Fig. 7; Shanahan et al., Reference Shanahan, Overpeck, Anchukaitis, Beck, Cole, Dettman, Peck, Scholz and King2009) and other Cameroon lakes (Ngeutsop et al., 2011; Lebamba et al., Reference Lebamba, Vincens and Maley2012).
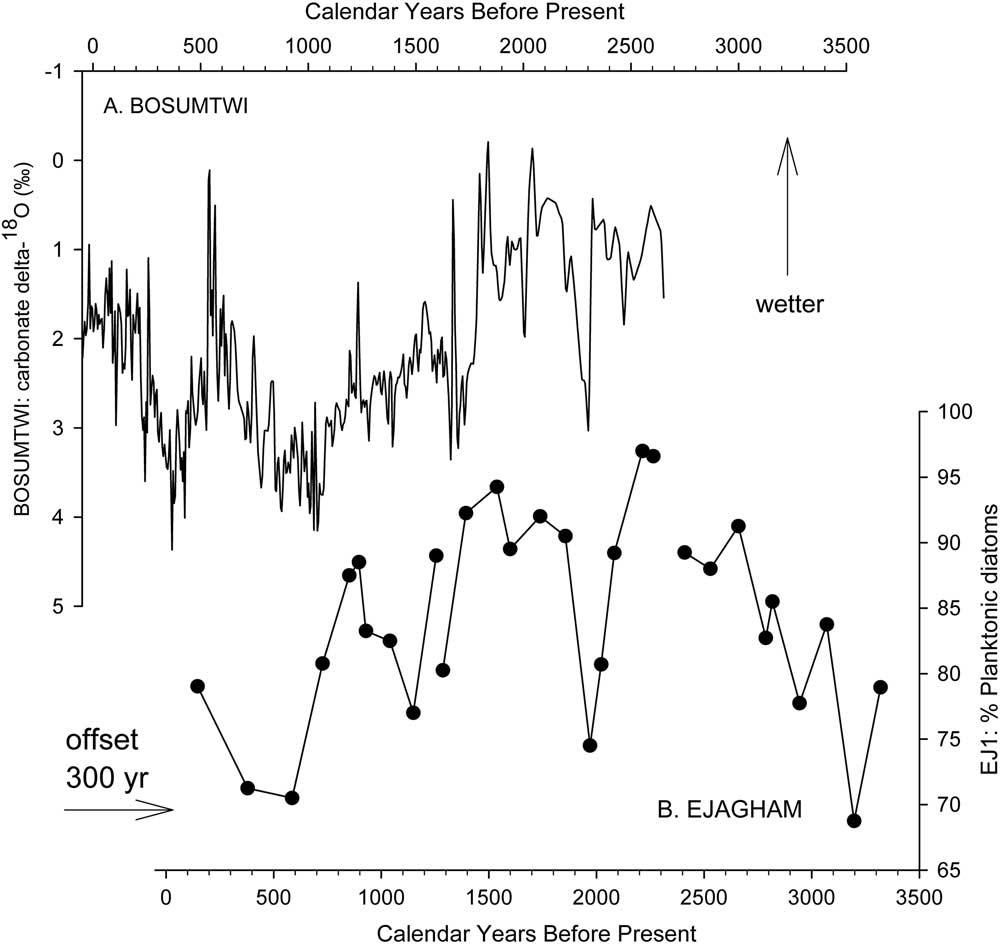
Figure 7 Comparison of (a) Lake Bosumtwi record of carbonate δ18O (Shanahan et al., Reference Shanahan, Overpeck, Anchukaitis, Beck, Cole, Dettman, Peck, Scholz and King2009) with composite planktonic diatom stratigraphy in the three subsections of core EJ1 (b). Shifting the provisional age model for EJ1 by 300 yr brings the diatom series into reasonably close alignment with the Bosumtwi isotope record. Both series indicate wetter climatic conditions upward on the figure.
The age of Lake Ejagham
Our results provide a maximum age of ca. 9 ka for Lake Ejagham. The radiocarbon age obtained for woody debris in the coarse sediments at the base of core EJ2 is unlikely to be contaminated with ancient carbon, as appears to be indicated for the lacustrine sediment in core EJ1. However, it is possible that the wood was reworked from older deposits.
The ca. 9 ka basal age of EJ2 is greater than would be expected from the mean sediment accumulation rate of ~25 cm/yr inferred for the upper 526 cm of the core, but it would be consistent with the mean sediment accumulation rate determined for EJ1. Downward linear extrapolation of the radiocarbon age of sediment at 580 cm depth in EJ2 suggests an expected age closer to 7 ka at the base of the core. Therefore, it appears that the sediment record between 526 cm and 580 cm depth is highly compressed or discontinuous, or (less likely) that the estimated depth of the upper subsample is incorrect because of the loss of structural integrity in the EJ2 core during storage.
The origin of Lake Ejagham
To our knowledge, there has never been a definitive geographic or geologic investigation of Lake Ejagham and its origin. It has been hypothesized to be a solution lake (Schliewen et al., Reference Schliewen, Tautz and Pääbo1994, Reference Schliewen, Rassmann, Markmann, Markert, Kocher and Tautz2001; http://www.lakepedia.com/lake/ejagham.html [accessed 2016]), but we consider this to be unlikely. Local geographic mapping reveals no formations with substantial soluble deposits. Thin layers of limestone, marl, and halite do contribute to high solute concentrations in local mineral springs (Fairhead et al., Reference Fairhead, Okerere and Nnange1991; Njoh et al., Reference Njoh, Nforsi and Datcheu2015), but in the vicinity of Lake Ejagham, those soluble lenses are interbedded with insoluble shales and sandstones that are likely to prevent the formation of large subterranean dissolution cavities (Njoh, O.A., personal communication, 2016). The lack of obvious karst solution features in the surroundings (Fig. 1b) further suggests that Ejagham is not a solution lake.
We also consider it highly improbable that Lake Ejagham formed as a result of climatic change 9 ka ago, as Lake Victoria did following an extreme drought ca. 17 ka ago (Stager and Johnson, Reference Stager and Johnson2007; Stager et al., Reference Stager, Ryves, Chase and Pausata2011). The hydrologic variability we infer from the EJ1 diatom record resembles that reflected in the Bosumtwi oxygen isotope record (Fig. 7; Shanahan et al., Reference Shanahan, Overpeck, Anchukaitis, Beck, Cole, Dettman, Peck, Scholz and King2009), and moderately reduced lake levels and/or rainfall ca. 2000 yr BP and ca. 1000 yr BP indicated by low percentages of planktonic diatoms in the EJ1 core and high δ18O in the Bosumtwi record have also been reported from Cameroonian volcanic crater lakes Barombi Mbo (Giresse et al., Reference Giresse, Maley and Brenac1994; Lebamba et al., Reference Lebamba, Vincens and Maley2012), Mbalang, and Ossa (Nguetsop et al., Reference Nguetsop, Servant-Vildary, Servant and Roux2010, Reference Nguetsop, Bentaleb, Favier, Martin, Bietrix, Giresse, Servant-Vildary and Servant2011). We therefore consider it probable that hydroclimates at Ejagham were similar to those elsewhere in the West African summer monsoon region during the early Holocene as well. The hydroclimate records of Bosumtwi (Peck et al., Reference Peck, Green, Shanahan, King, Overpeck and Scholz2004; Shanahan et al., Reference Shanahan, Overpeck, Anchukaitis, Beck, Cole, Dettman, Peck, Scholz and King2009) and Barombi Mbo (Giresse et al., Reference Giresse, Maley and Brenac1994; Lebamba et al., Reference Lebamba, Vincens and Maley2012) both indicate continuously wet conditions throughout the early Holocene because of insolation-driven strengthening of the West African summer monsoon. We therefore conclude that the transition from coarse material to lacustrine sediments at the base of the EJ2 core does not reflect recovery from a desiccation event. We also conclude that the lake basin itself formed no more than 9000 yr ago as well, because the region’s extremely wet monsoonal climates during preceding centuries of the early Holocene (Giresse et al., Reference Giresse, Maley and Brenac1994; Peck et al., Reference Peck, Green, Shanahan, King, Overpeck and Scholz2004; Shanahan et al., Reference Shanahan, Overpeck, Anchukaitis, Beck, Cole, Dettman, Peck, Scholz and King2009; Lebamba et al., Reference Lebamba, Vincens and Maley2012) would presumably have established lacustrine conditions within a preexisting depression.
Lake Ejagham lies more than 60 meters above the floodplains of the region’s major river systems, so a fluvial origin of the basin is unlikely despite a previous suggestion that the lake might have formed as a result of flooding (Offem et al., Reference Offem, Ayotunde, Ikpi, Ada and Ochang2011). The coarser grains (1–4 mm) in the basal section of core EJ2 are instead likely to have washed into the center of the lake from the shoreline through wave action or storm runoff when the lake was newly formed and biogenic deposits had not yet buried and immobilized most of the mineral sands within the lake.
The majority of lakes in Cameroon are of volcanic or phreatic origin. Most are much deeper than Ejagham, circular in shape, and confined within relatively high crater rims. Ejagham’s shallowness, elliptical shape, and lack of a high rim sets it apart from most other Cameroon lakes, but those features are not in themselves conclusive evidence against an explosive origin and are shared by several other lakes in the region (Kling, Reference Kling1987). Exposures of basaltic bedrock in the vicinity of the Mamfe basin (Tokam et al., Reference Tokam, Tabod, Nyblade, Julià, Wiens and Pasyanos2010) indicate previous volcanism there, so an explosive origin for the lake cannot be completely ruled out. However, those igneous deposits are apparently too old to be directly relevant to the origin of Lake Ejagham, which is millions of years younger.
It should be noted that, even if it arose from a volcanic or phreatic explosion, Lake Ejagham poses little or no risk of magmatic carbon dioxide bursts such as those that occurred in Lakes Monoun and Nyos during the 1980s (Kling et al., Reference Kling, Evans, Tanyileke, Kusakabe, Ohba, Yoshida and Hell2005) and that have been suggested as a possible threat to biodiversity at Ejagham as well (e.g., IUCN, 2016). The conditions in shallow, easily mixed lakes such as Ejagham are not conducive to the accumulation of large quantities of dissolved magmatic gases.
DAL and others (e.g., http://ww2.kqed.org/science/2015/07/10/nasas-cassini-spacecraft-reveals-mystery-lakes-on-saturns-moon-titan/ [accessed October 15, 2016]) have suggested that Lake Ejagham is the result of an impact from a small bolide. The presence of wood splinters at the base of EJ2 is consistent with forceful impact, yet we found no shocked mineral grains or melt spherules in the basal sediments of core EJ2. There is no known field of impact ejecta around Lake Ejagham, but the lack of such features does not necessarily exclude an extraterrestrial bolide origin (Schaller et al., Reference Schaller, Fung, Wright, Katz and Kent2016).
French’s (Reference French1998) review of impacts and the impactors that strike the earth indicates that an object 50–55 m in diameter can form a small, simple crater about the size of Lake Ejagham (~1 km diameter). The lake’s elliptical shape, which is not as commonly seen among impact craters as more nearly circular geometries, can nonetheless result from a low angle or oblique impact (Elbeshausen et al., Reference Elbeshausen, Wünnemann and Collins2013). However, definitive evidence of hypervelocity impact rests on petrographic and geochemical signals, including relatively high content of distinctive elements such as iridium, osmium, and chromium (French and Koeberl, Reference French and Koeberl2010). Isotopic analyses of the EJ2 samples were not performed because of lack of sufficient sample volumes, but the petrographic evidence we seek here is commonly used to document shock effects within quartz and feldspar grains and is generally the most definitive. The unusually high pressures and temperatures associated with impact events can leave highly diagnostic damage within grains including melting, intracrystalline shock damage (planar deformation structures), and high-pressure polymorphs of minerals (e.g., coesite). Less diagnostic features in grains affected by impact include orthogonal or parallel planar fractures and browning of quartz (a petrographic effect attributable to high-temperature effects on small fluid inclusions in the crystal, commonly called “toasting”). Toasting can be diagnostic for impact, but only when accompanied by other petrographic evidence, which is lacking in our study. It was found in abundance in quartz grains extracted from Lake Bosumtwi (Morrow, Reference Morrow2007), for example, as were quartz planar deformation features and silicate melt grains.
Toasting most commonly affects large areas of the altered quartz grain, so it is peculiar here that the apparent toasting found in a few quartz grains from core EJ2 (Fig. 4c and d) affected only the area immediately adjacent to fluid-inclusion-laden curviplanar fractures (Whitehead et al., Reference Whitehead, Spray and Grieve2002; Ferrière et al., Reference Ferrière, Koeberl, Reimold, Hecht and Bartosova2009). A few of the medium and fine quartz grains also exhibited parallel planar fractures (Fig. 4b). Planar fractures can be explained by other stresses besides impact and are only considered diagnostic where accompanied by planar deformation features, which were not seen in Lake Ejagham quartz particles examined for this study.
The morphological and petrographic evidence from this study is not conclusive, but it is suggestive of an impact origin for Lake Ejagham. We therefore propose that Lake Ejagham be considered a suspected but unproven impact crater that merits further investigation. The energy released by an impactor large enough to produce a crater the size of Lake Ejagham would have been on the order of 1016 J or equivalent to 10 to 15 megatons of TNT (French, Reference French1998) and therefore could cause significant local devastation. As this region was likely to have been occupied by people at the time the lake formed, we suggest that ethnographic or archaeological evidence of possible effects on the human community might also be investigated in order to independently assess the possible impact scenario as was done for the folk history of exploding lakes in Cameroon (Shanklin, Reference Shanklin2007).
Origin of the endemic fishes
The lack of major variability in the organic and water contents of the core sediments above the basal deposit in EJ2 indicates that no major erosion or desiccation events occurred in the lake since its origin. The absence of visually obvious changes in sediment composition in the EJ1 core, along with the continuous numerical dominance of a single assemblage of planktonic diatom taxa in the core, further supports that conclusion. If our interpretation of the age model is correct, then no more than the last five centuries are absent from the sediment record of EJ1, a period during which there were no exceptionally severe droughts indicated in the records of Barombi Mbo and Bosumtwi. Together, these results indicate that Lake Ejagham has been in continuous existence without major changes in water chemistry or depth since the early Holocene. Therefore, the evolution of the cichlids is also unlikely to have been interrupted or complicated by severe environmental disturbances in the lake during the last 9 ka.
This conclusion of environmental stability over 9 ka within Lake Ejagham strengthens the original argument (Schliewen et al., Reference Schliewen, Rassmann, Markmann, Markert, Kocher and Tautz2001) that the lake is a suitable setting for sympatric speciation to occur in. Sympatric speciation is one of the most controversial and fascinating processes within evolutionary biology, defined as the evolution of reproductive isolation barriers within a population through natural and sexual selection alone, without the benefit of geographic isolating barriers such as periodic changes in lake levels resulting in multiple isolated basins (Coyne and Orr, Reference Coyne and Orr2004; Bolnick and Fitzpatrick, Reference Bolnick and Fitzpatrick2007). Sympatric speciation was originally proposed by Darwin, but convincing examples remain scarce, in part because of the rarity of stable geographic settings in which all historical possibility of divided ranges or lake basins can be ruled out (Bolnick and Fitzpatrick, Reference Bolnick and Fitzpatrick2007; Martin, Reference Martin2012, Reference Martin2013). The case for sympatric speciation within Lake Ejagham is further strengthened by the fact that two genetically distinct lineages of cichlids have diversified within the confines of this tiny lake—in fact, the presence of multiple speciating cichlid lineages within one lake is unique to Ejagham among all known crater lake cichlid radiations (Seehausen, Reference Seehausen2006).
Finally, the divergence of each species flock is clearly recent based on their limited genetic and phenotypic differentiation (Dunz and Schliewen, Reference Dunz and Schliewen2010; Neumann et al., Reference Neumann, Stiassny and Schliewen2011; Martin et al., Reference Martin, Cutler, Friel, Dening, Coop and Wainwright2015), which is consistent with the young lake age determined in this study. We estimate that the Coptodon species flock in Lake Ejagham diverged within the last 3,600 yr based on genome-wide estimates of their genetic differentiation (F st = 0.165) and effective population size (Ne = 10,000; Poelstra, J.W., Martin, C.H., unpublished data) using the approximation discussed in Cavalli-Sforza and Bodmer (Reference Cavalli-Sforza and Bodmer1971). Nonetheless, strong assortative mating in the wild demonstrates that these are incipient species that are diverging along multiple independent phenotypic axes of coloration and trophic morphology (Martin, Reference Martin2013). A very young age for this species flock is consistent with the colonization of Lake Ejagham within the last 9000 yr.
The cichlids of Lake Ejagham and the other two Cameroon lakes with endemic cichlid radiations, Lake Bermin (9 endemic species) and Lake Barombi Mbo (11 endemic species), are among the clearest known examples of sympatric speciation in nature (Coyne and Orr, Reference Coyne and Orr2004; Schliewen et al., Reference Schliewen, Kocher, McKaye, Seehausen and Tautz2006; Bolnick and Fitzpatrick, Reference Bolnick and Fitzpatrick2007). However, ruling out all possible allopatric scenarios for speciation in these lakes also requires ruling out repeated colonization of the lakes by neighboring riverine cichlids. Each colonization could have resulted in a subsequent speciation event within the lake, as observed in other speciation model systems such as sticklebacks, Gasterosteus aculeatus (Schluter and McPhail, Reference Schluter and McPhail1992). Recent genome-wide sequencing studies in all known cichlid radiations in Cameroon lakes (including both lineages within Lake Ejagham) suggest some evidence for additional gene flow from neighboring rivers after the initial colonization of each lake. Thus, each lake was probably colonized by cichlids more than once, casting some doubt on the sympatric speciation hypothesis (Martin et al., Reference Martin, Cutler, Friel, Dening, Coop and Wainwright2015). However, it remains unknown how many times these lakes were colonized by cichlids and whether some speciation events occurred in sympatry while others were facilitated by the additional gene flow. Preliminary analyses suggest that two of the three Coptodon species in Ejagham (the benthivore C. deckerti and zooplanktivore C. fusiforme) may have resulted from secondary colonization, while the third species (the predator C. ejagham) may still have diverged in sympatry (Martin et al., Reference Martin, Cutler, Friel, Dening, Coop and Wainwright2015; Poelstra, J.W., Martin, C.H., unpublished data).
CONCLUSIONS
Radiocarbon dating of a coarse-grained deposit at the base of 6-m-long sediment core EJ2 from the center of Lake Ejagham suggests that the lake formed roughly 9 ka ago, possibly as a result of an extraterrestrial bolide impact. The continuous existence of the lake after its initial formation is indicated by (1) lack of variability in organic and carbonate content in the EJ2 core, (2) the similarity of diatom-based hydroclimate history from core EJ1 to other West African climate records that document a continuously humid Holocene, and (3) the absence of coarse-grained deposits in the younger sections of both cores. We therefore conclude that the maximum age of the endemic cichlid species flock in Lake Ejagham is also ca. 9 ka, a result that is consistent with genetic evidence as well.
Overall, the young age of Lake Ejagham provides a clear demonstration that sympatric speciation, perhaps facilitated by repeated bouts of riverine colonization, can occur in a remarkably short amount of time in different cichlid lineages within the same small lake. The young age of these species flocks is comparable to that of recent adaptive radiations of Cyprinodon pupfishes in Caribbean salt lakes (8–10 ka; Humphries and Miller, Reference Humphries and Miller1981; Martin and Wainwright, Reference Martin and Wainwright2011) and the youngest age estimates for the Lake Victoria cichlid radiation (12–15 ka; Seehausen, Reference Seehausen2002; Stager and Johnson, Reference Stager and Johnson2007).
Acknowledgments
DAL collected the EJ1 and EJ2 cores with the assistance of Ndoni Sangwa Paul. K. Beuning assisted DAL with the organic and carbonate content analyses for the EJ2 core. P. Palmer, R. Kendall, and J. Richardson assisted with transfer of the cores and notes from DAL’s lab in 2015, and O.A. Njoh and B. Burress provided valuable information about local geology and ethnography. This research was supported by a National Science Foundation grant to JCS (ATM-1358362) and by the Department of Biology, University of North Carolina at Chapel Hill to CHM. C. Dening and P. Enyong assisted with the collection of fish specimens. Chief Obi of Eyumojock village, the regional office of the Divisional Authority of Eyumojock, and the Ministry of Scientific Research and Innovation, Republic of Cameroon granted permission to conduct this research. N. Gonwuou assisted with obtaining permits. This paper is dedicated to the memories of D. Livingstone, J. Richardson, and J. Stager. Radiocarbon dating was performed by NOSAMS, Woods Hole, MA.