


Introduction
Substratum specificity is a strong feature in Fuscidea. However, there have been occasional reports of corticolous specimens of mainly saxicolous species. For example, F. recensa (Stirt.) Hertel is capable of inhabiting both rock and bark in Scandinavia (Tønsberg Reference Tønsberg1992) but corticolous specimens have not been formally recognized (Nordin et al. Reference Nordin, Moberg, Tønsberg, Vitikainen, Dalsätt, Myrdal and Ekman2010). In Great Britain and Ireland, species of Fuscidea inhabit rock or, more rarely, bark, occasionally wood, and 9 of the 11 species are either exclusively saxicolous (8 spp.) or exclusively corticolous/lignicolous (2 spp.), with F. cyathoides (Ach.) V. Wirth & Vězda and F. recensa (Stirt.) Hertel, V. Wirth & Vězda capable of inhabiting both rock and bark (Gilbert et al. Reference Gilbert, Purvis, Skjolddal and Tønsberg2009; B. Coppins, pers. comm.). Substratum ecology and the presence/absence of soredia have been suggested as important characters for formal recognition of taxonomic entities of F. cyathoides at the species (Hafellner & Türk Reference Hafellner and Türk2001; Hafellner Reference Hafellner2002) and varietal levels (Fries Reference Fries1831; Magnusson Reference Magnusson1925; Zschacke Reference Zschacke1927) (Table 1).
Table 1 Overview of Fuscidea cyathoides nomenclature

Magnusson (Reference Magnusson1925) discussed seven saxicolous forms of F. cyathoides (as Lecidea rivulosa Ach.) and introduced var. infuscata H. Magn., separated from var. cyathoides based on habitat and thallus colour (see Supplementary Material Table S1, available online). None of the F. cyathoides forms are still recognized and Oberhollenzer & Wirth (Reference Oberhollenzer and Wirth1984) synonymized var. infuscata with var. cyathoides (see Taxonomy below).
In F. cyathoides, corticolous material has been attributed taxonomic rank at both infraspecific and specific levels. According to Fries (Reference Fries1831), F. cyathoides var. corticola (Fr.) Kalb (as Biatora rivulosa b. corticola Fr.) is distinct from var. cyathoides in possessing a different thallus colour (i.e. black-brown when dry and greenish when wet vs. grey when dry and umber brown when wet in var. cyathoides). Although some authors (e.g. Oberhollenzer & Wirth Reference Oberhollenzer and Wirth1984; Gilbert et al. Reference Gilbert, Purvis, Skjolddal and Tønsberg2009) consider the corticolous variety as merely F. cyathoides on bark, others (e.g. Inoue Reference Inoue1981; Santesson et al. Reference Santesson, Moberg, Nordin, Tønsberg and Vitikainen2004) recognize this taxon as F. cyathoides var. corticola.
Zschacke (Reference Zschacke1927) recognized the absence of a black prothallus, the larger and flatter thallus as well as the larger apothecia as diagnostic characters for distinguishing F. fagicola (as Lecidea fagicola Zschacke) from F. cyathoides (as L. rivulosa). When comparing the so-called Fagus-type of apothecia of var. corticola (i.e. apothecia from specimens growing on Fagus in southern Europe) with those on Betula, the so-called Betula-type, Oberhollenzer & Wirth (Reference Oberhollenzer and Wirth1984) did not find any significant variation. Based on this result they concluded that L. fagicola most certainly is synonymous with var. corticola. Hafellner & Türk (Reference Hafellner and Türk2001) transferred L. fagicola to Fuscidea and placed F. cyathoides var. corticola in synonymy without any explanatory discussion.
Hafellner (Reference Hafellner2002) made the combination F. stiriaca (A. Massal.) Hafellner based on the basionym Biatora stiriaca A. Massal., which was treated as a synonym of var. cyathoides by Magnusson (Reference Magnusson1925) (as L. rivulosa var. corticola (Fr.) Jatta) and by Vainio (Reference Vainio1934) (as L. rivulosa f. corticola (Fr.) Vain.), and he synonymized F. fagicola with F. stiriaca. The sorediate form, var. sorediata (H. Magn.) Poelt, is saxicolous and rare. It was, for example, accepted by Santesson et al. (Reference Santesson, Moberg, Nordin, Tønsberg and Vitikainen2004) and Gilbert et al. (Reference Gilbert, Purvis, Skjolddal and Tønsberg2009).
Molecular approaches changed the concept of species delimitation (as discussed in Resl et al. Reference Resl, Mayrhofer, Clayden, Spribille, Thor, Tønsberg and Sheard2016) and provided a new approach to assess the status of sorediate lichens. In the studies of Pseudevernia furfuracea (L.) Zopf by Ferencová et al. (Reference Ferencová, Del Prado, Pérez-Vargas, Hernández-Padrón and Crespo2010), Mycoblastus alpinus (Fr.) Kernst./M. affinis (Schaerer) Schauer by Spribille et al. (Reference Spribille, Klug and Mayrhofer2011), several species of Dirina Fr. by Tehler et al. (Reference Tehler, Ertz and Irestedt2013) and Rinodina degeliana Coppins/R. subparieta (Nyl.) Zahlbr. by Resl et al. (Reference Resl, Mayrhofer, Clayden, Spribille, Thor, Tønsberg and Sheard2016) no taxonomic relevance was given to the presence of soredia. For example, Spribille et al. (Reference Spribille, Klug and Mayrhofer2011) confirmed the hypothesis of Tønsberg (Reference Tønsberg1992) that M. alpinus and M. affinis are conspecific using a combined matrix of two protein coding (EF1-α, MCM7) and ITS genes. These two species differ in their morphologies (esorediate, richly fertile vs. sorediate, sterile or sparingly fertile) and chemistries (usnic acid absent/thallus grey vs. usnic acid present in the (yellowish) soralia).
Here we aim to revise the taxonomy of F. cyathoides s. lat., providing a morphological, chemical and phylogenetic investigation of all three currently recognized varieties, and clarify the taxonomy of F. cyathoides, including the related F. fagicola and F. stiriaca.
Material and Methods
Taxon sampling
Herbarium material studied was provided by BG, HO, MSC, LD, UPS, TUR and H-Ach, as well as from private collections. As Fuscidea cyathoides var. sorediata is scarce in Europe, we managed to obtain only one fresh specimen.
Morphology
To determine morphological differences between varieties, the apothecia and thalli were examined by light microscopy in hand-cut sections mounted in water with 10% KOH using a Carl Zeiss Axioskop 2 microscope. Twenty specimens of F. cyathoides, including all three varieties, were investigated. The following morphological characters were studied: overall diameter of the apothecia and the areoles, height of the epihymenium and hymenium, length and width of the ascospores, and colour of the thalli (Table 2). The ratio between length and width of spores was calculated. Characters were examined using an unconstrained linear ordination, Principal Components Analysis (PCA), to explore the morphological variation. We performed the analysis with centering and standardization of characters in CANOCO 5 (Ter Braak & Šmilauer Reference Ter Braak and Šmilauer2012).
Table 2 Overview of morphological characters measured on the varieties of F. cyathoides, given as (smallest values–)arithmetic mean±SD(–largest values). Numbers in square brackets are the number of replicates

Secondary chemical compounds
Lichen substances were analyzed by thin-layer chromatography (TLC), using the methods of Culberson & Kristinsson (Reference Culberson and Kristinsson1970), Culberson (Reference Culberson1972) and Menlove (Reference Menlove1974). All three solvents (A, B´ and C) were used, with glass plates in solvent C for the detection of fatty acids. Selected specimens were also run in solvent G for a detailed study of β-orcinol depsidone fumarprotocetraric acid and possible occurrences of the related substances protocetraric and succinprotocetraric acids (see Culberson et al. Reference Culberson, Culberson and Johnson1981).
DNA extraction, PCR amplification and sequencing
DNA from 10 specimens of Fuscidea cyathoides were analyzed together with six other Fuscidea species for three genes. Altogether we generated 36 new sequences, in addition to sequences of Fuscidea downloaded from GenBank (Table 3). DNA was extracted from apothecia or soredia with thallus using the DNeasy Plant Mini Kit (Qiagen). Although phylogenies based on ITS alone have been considered sufficient for infraspecific taxonomic investigations (e.g. Davydov et al. Reference Davydov, Peršoh and Rambold2010; Solheim et al. Reference Solheim, Torp and Hietala2013), we constructed a concatenated data set of three markers from two different genomes (mtSSU, nuITS and nuLSU). The mtSSU fragment was made with the primers mrSSU1 and mrSSU3R (Zoller et al. Reference Zoller, Scheidegger and Sperisen1999), while ITS and LSU were amplified by ITS1F (Gardes & Bruns Reference Gardes and Bruns1993), ITS4 (White et al. Reference White, Bruns, Lee and Taylor1990) and nu-nuLSU-1125-3´ (Vilgalys & Hester Reference Vilgalys and Hester1990). The PCR master mix included: 1× Buffer II GeneAmp® 10×PCR (Applied Biosystems), 2·5 µM MgCl2 (Applied Biosystems), 20 µM dNTPs (Promega), 0·6 µM of each primer, 0·036 U AmpliTaq® DNA Polymerase (Applied Biosystems), 5·0 µl of genomic DNA extract and distilled water to a total volume of 25 µl.
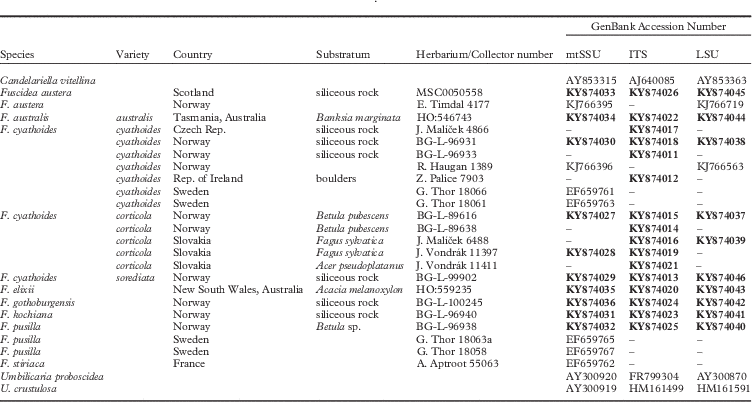
Table 3 List of voucher specimens with their collection details and GenBank Accession numbers, in addition to sequences included from GenBank. Newly generated sequences are indicated in bold
The PCR reactions were performed using the C1000TM Touch thermal cycler (Bio-Rad Laboratories). The following programs were used for mtSSU: initial denaturation at 94°C for 5 min, touchdown of six cycles (94°C for 30 s, 62–56°C for 30 s, and 72°C for 1 min 45 s), followed by 34 cycles of 94 °C for 30 s, 56°C for 30 s, 72°C for 1 min 45 s, and a final elongation at 72°C for 10 min. The same were used for LSU and ITS, except for the annealing temperature where the touchdown ranged from 63–57°C for six cycles, ending at 57°C for 34 cycles.
PCR products were visualized on a 1% Red Gel-stained agarose gel under UV light and purified using Exosap-IT (GE Healthcare). The PCR products were sequenced using the PCR primers with the BigDye Terminator Cycle Sequencing Kit (Applied Biosystems) and run on an ABI Prism 3700XL DNA analyzer (Applied Biosystems) at the DNA Sequencing Laboratory, University of Bergen, Norway. The sequences were assembled in SeqMan II version 4.05 (DNASTAR).
Phylogenetic analyses
Geneious version 8.1.8 (Biomatters Ltd.) was used to align the mtSSU, LSU and ITS sequences with 65% similarity option on (Gap penalty=14·5, Gaps extension penalty=5), followed by manual adjustment. Candelariella vitellina (Hoffm.) Müll. Arg. was used as outgroup and two sequences, Umbilicaria proboscidea (L.) Schrad. and U. crustulosa (Ach.) Frey, as a sister group to Fuscidea.
To identify suitable substitution models for all fragments (i.e. mtSSU, LSU, ITS1, 5.8S and ITS2), a likelihood ratio test (Huelsenbeck & Crandall Reference Huelsenbeck and Crandall1997) was performed using the software jModelTest version 2.1.7 (Posada Reference Posada2008). The model GTR+G was selected for mtSSU, GTR+I+G for LSU, SYM+G for ITS1, K80+I for 5.8S, HKY+G for ITS2 and GTR+I+G for the concatenated data set.
To detect potential conflicts between the data sets, we inspected the internodes of the phylogenetic trees with bootstrap values >70%. These were generated using the neighbour-joining model with a maximum likelihood distance (e.g. Reeb et al. Reference Reeb, Lutzoni and Roux2004). Bootstrap scores were calculated using 2000 non-parametric replicates in the Jukes-Cantor distance model implemented in Geneious version 8.1.8 (Biomatters Ltd.).
Phylogenetic relationships were estimated from the data sets, both from each gene separately and the concatenated data, using MrBayes version 3.2.1 (Ronquist & Huelsenbeck Reference Ronquist and Huelsenbeck2003) to sample trees using a Markov chain Monte Carlo (MCMC) method in the Bayesian inference (BI). Tree sampling performed under the MCMC analysis was run for 4 000 000 generations with four parallel chains starting from a random tree, using the default setting with temperature of 0·2. Gaps were coded as a fifth character state. Sampling frequency of trees was every 10th generation, including branch lengths. The first 40 000 trees (i.e. 10% of the total number) were deleted as burn-in. A majority-rule consensus tree with average branch lengths was constructed from 360 000 trees and visualized in Geneious (Biomatters Ltd.). Significant posterior probabilities were ≥95%.
Weighted maximum parsimony (MP) and maximum likelihood (ML) analyses were carried out in PAUP*4.0b10 (Swofford Reference Swofford2002) to construct MP and ML trees with bootstrap support. A first heuristic search was run to find MP trees using random sequence additions with 500 replicates, and tree bisection-reconnection branch swapping (TBR). The MulTrees and steepest descent options were on, and the collapse zero-length branches option was off. Gaps were coded as a fifth character state. To estimate the branch support for the MP trees, 1000 bootstrap replicates with 10 random additions of the taxa were performed. A second heuristic search with 500 replicates under the ML criterion and the selected substitution model was run using the MP trees from the previous heuristic search as starting trees. Branch support for the ML trees was estimated by 100 bootstrap replicates with 10 random additions of the taxa. High bootstrap support was considered to be ≥70%.
Results
Morphological examination
This study showed that only corticolous specimens had greenish to green thalli and that saxicolous specimens varied from grey to brown (Table 2). Corticolous specimens developed tuberculate apothecia more frequently (90% of specimens examined) than saxicolous thalli (20% of specimens examined). Var. sorediata, represented by only one specimen, had smaller and fewer apothecia (Table 2). The ascospores of all three varieties were bean-shaped. However, those of the corticolous specimens were narrower (4·53±1·82 µm) than saxicolous thalli (4·79±1·21 µm), but were similar in length (i.e. 10·17±4·79 µm and 10·24±3·11 µm, respectively) (see Table 2).
PCA based on morphological characters of F. cyathoides did not separate corticolous and saxicolous specimens along the two first ordination axes representing 28·77% and 24·11% of the variation, respectively (Fig. 1). One character, namely the ratio between the length and width of ascospores, had a larger range for var. cyathoides than var. corticola. The heights of the hymenium and epihymenium, and width of ascospores, were all positively correlated with the size of areoles and tuberculate apothecia.

Fig. 1 Biplot of the first two principal component axes, showing morphological variation of Fuscidea cyathoides identified in this study. Key to variables: Apom=diam. (mm) of apothecia; Apotub=diam. (mm) of tuberculate apothecia; Aream=diam. (mm) of areoles; Hym=height (μm) of hymenium; Epi=height (μm) of epihymenium; Lspore=length (μm) of ascospores; Wspore=width (μm) of ascospores; Lwspore=ratio of Wspore:Lspore.
Secondary chemical compounds
Analysis of secondary chemical compounds in the fumarprotocetraric acid chemosyndrome did not reveal any chemical differences between the specimens. The major and diagnostic constituent was fumarprotocetraric acid; a trace of the satellite substance protocetraric acid was present in most specimens, whereas succinprotocetraric acid was not detected in any of the specimens tested.
Phylogeny of Fuscidea cyathoides
As no conflicts were detected between the data sets for different genes, they were combined and the final aligned sequence matrix comprised 26 taxa with 2187 characters, of which 1618 were constant and 361 informative. The GenBank Accession numbers are given in Table 3. The majority-rule consensus tree from the BI is displayed in Fig. 3. The average −ln likelihood of the tree was 8081·83 and the average standard deviation of split frequencies was 0·0025, indicating that two independent runs of the Markov chain search converged. The calculated likelihood parameters of the MCMC analysis are summarized in Table S2 (see Supplementary Material, available online).
A heuristic search using the parsimony criterion resulted in 100 MP trees of length 1041 with consistency index=0·7080, homoplasy index=0·2920, retention index=0·6984 and rescaled retention index=0·4945. A second heuristic search under the ML criterion and the GTR+I+G model using the MP trees as starting trees resulted in three equally best ML trees (−lnL=8104·6499). The consensus ML tree was incongruent with the BI tree in the position of F. kochiana (Hepp) V. Wirth & Vězda, and five specimens (A. Aptroot 55063, M. Zahradníková MZ05 (BG-L-96931), G. Thor 18066, G. Thor 18061 and R. Haugan 1389) within the F. cyathoides group. All incongruences are indicated with a circle in Fig. 3.
All the samples of Fuscidea included here formed a monophyletic group. All three varieties of F. cyathoides were clustered in one subgroup with PP=0·99, MP=97%, ML=99% support. Within this subgroup, no clear classification in the varieties of corticolous and saxicolous specimens was discovered. The specimens from Central Europe (i.e. the Czech Republic and Slovakia) formed a separate group to those from North-West Europe (i.e. Norway and the Republic of Ireland).
Discussion
Neither the chemistry nor the molecular data show evidence for differentiation within F. cyathoides. Our results suggest that the bean-shaped spores becoming brownish when mature and the production of fumarprotocetraric acid are the only diagnostic characters for the recognition of F. cyathoides.
Our findings agree with Bylin et al. (Reference Bylin, Arnerup, Hogberg and Thor2007), where corticolous and saxicolous specimens of F. cyathoides were grouped together, but with less sampling and MP bootstrap support lower than 80%. Moreover, Fuscidea stiriaca was clustered with var. cyathoides (MP=100%).
The included representatives with apothecia of both the Fagus- and Betula-types show no morphological or genetic differences. The observed variation between these apothecium types is not significant (see Figs 1 & 3), confirming the report of Oberhollenzer & Wirth (Reference Oberhollenzer and Wirth1984). In the PCA, the Betula-type (BG-L-89616) and the Fagus-type (JV 11397, JV 11411 and JM 6488) are not separated from each other; furthermore, specimens BG-L-89616 and JV 11411 are found to overlap. We consider the colour and the presence of crystals in the apothecia as adaptations to localities with direct light exposure. It should be noted that Fahselt (Reference Fahselt1981) found that levels of perlatolic and fumarprotocetraric acids in populations of Cladonia stellaris (Opiz) Pouzar & Vězda and C. rangiformis Hoffm., respectively, were influenced by light intensity. Massalongo (Reference Massalongo1852) suggested that the bean-shaped spores and tuberculate apothecia were diagnostic for F. stiriaca (as Biatora stiriaca) (see Fig. 1). This cannot be supported since both characters are also present in var. cyathoides.
In the present study var. sorediata has the smallest apothecia (see Table 2), but this feature is considered to be a result of a biological energy saving strategy (see Tønsberg Reference Tønsberg1992) and should not be used as a diagnostic character for species forming species pairs sensu Poelt (Reference Poelt1970, Reference Poelt1972).
To conclude, no significant genetic difference between specimens reflecting the morphological and ecological variations was found in F. cyathoides. Therefore, we synonymize var. corticola and var. sorediata with the typical form. Fuscidea fagicola and F. stiriaca are synonymized with F. cyathoides.
Taxonomy
Fuscidea cyathoides (Ach.) V. Wirth & Vězda
Beitr. nat. kdl. Forsch. in Südwestdtschl. 31: 92 (1972).—Lichen cyathoides Ach., Lichenographiae Suecicae Prodromus: 62 (1798); type: Sweden, in saxis et rupibus (H-Ach 273 F & G—lectotypus [!] in Oberhollenzer & Wirth, Beih. Nova Hedwigia 79: 552 (1984)).—Lecidea cyathoides (Ach.) Ach., Methodus qua Omnes Detectos Lichenes: 51 (1803).—Biatora cyathoides (Ach.) Oxner, Flora of Lichens of the Ukraine 2: 78 (1968).
Lecidea rivulosa Ach., Methododus qua Omnes Detectos Lichenes: 38 (1803); type: Sweden, in saxis et rupibus (H-Ach 273 C—lectotypus [!] in Oberhollenzer & Wirth, Beih. Nova Hedwigia 79: 553 (1984)).—Biatora rivulosa (Ach.) Fr., Kongliga Vetenskaps Academiens Nya Handlingar: 269 (1822).—Microlecia rivulosa (Ach.) Choisy, Bull. Mens. Soc. Linn. Lyon 18: 151 (1949).
Lecidea rivulosa Ach. var. infuscata H. Magn., Kongliga Götheborgska Vetenskaps Samhällets Handlingar, Vetenskskaps Afd. 29: 27 (1925); type: Norway, Hordaland: Mosterhavn, Aug. 1910, Havaas, Havaas, Lich. Norv. Occ. 43 (UPS—lectotypus [!] in Oberhollenzer & Wirth, Beih. Nova Hedwigia 79: 554 (1984)).
Lecidea subrivulosa Vain., Acta Soc. Fauna Fl. Fenn. 57: 316 (1934); type: Russia [Finlandia]: in Somerikonvuoret in Suursaari v. Hoglandia, in rupe porphyrica, 1875, Vainio (TUR-Vainio 24352—holotype [!]).—Fuscidea subrivulosa (Vain.) P. James, Poelt & May. Inoue, Hikobia Supplement 1: 179 (1981).—Fuscidea subrivulosa (Vain.) P. James, Poelt & V. Wirth, Biblioth. Lichenol. 16: 154 (1981), nom. inval., Art. 41.4 (Melbourne).
Biatora rivulosa b. corticola Fr., Lichenographia Europaea Reformata: 272 (1831); type: Sweden, Småland: Femsjö, on bark, E. Fries: Exs. Lich. Suec. no. 39 (1818) (UPS—lectotypus [!] in Inoue, Hikobia Supplement 1: 178 (1981) as “holotypus”).—Fuscidea cyathoides var. corticola (Fr.) Kalb, Herzogia 4: 57 (Reference Kalb1976). Syn. nov.
Lecidea fagicola Zschacke, Verhandlungen des Botanischen Vereins der Provinz Brandenburg 69: 11 (1927); type: Frankreich, Corsica: Vizzavona, H. Zschacke (B—holotypus [lost, see Oberhollenzer & Wirth, Beih. Nova Hedwigia 79: 554 (1984)]; Frankreich, Corsica, Distr. Evissa: Silva Aitone, in valle rivi Aitone, c. 1300 m, Fagicola, 30 June 1969, J. Lambinon, Y. Rondon, A. Vězda (neotype [probably lost] designated by Oberhollenzer & Wirth, in Beih. Nova Hedwigia 79: 554 (1984)).—Biatorinella fagicola (Zschacke) Deschâtres & Werner, B. Soc. Bot. Fr. 121: 305 (1974).—Fuscidea fagicola (Zschacke) Hafellner & Türk, Stapfia 76: 152 (2001). Syn. nov.
Biatora stiriaca A. Massal., Ricerche sull´ autonomia del licheni crostosi: 125 (1852); type: Italia, vive sui faggi nelle Stiria, legit. Welwic. (VER—holotype [!]).—Lecidea stiriaca (A. Massal.) Jatta, Sylloge Lichenum Italicorum 39: 328 (1900).—Fuscidea stiriaca (A. Massal.) Hafellner, Fritschiana 33: 42 (2002). Syn. nov.
Lecidea rivulosa var. sorediata H. Magn., Göteborgs Kunglige Vetenskaps- och Vitterhets-Samhälles Handlingar, Ser. 4 29: 29 (1925); type: Sweden, Västergötland: par. Frölunda, Näset, on sunny boulder, 24 August 1924, A. H. Magnusson 9237 A (UPS, L-763155—lectotype, designated here).—Fuscidea cyathoides var. sorediata (H. Magn.) Poelt, Norw. J. Bot. 25: 127 (1978). Syn. nov.

Fig. 2 Fuscidea cyathoides.; A, saxicolous and esorediate specimen (Norway, TT 46572, BG-L-99904); B, saxicolous and sorediate specimen (Norway, TT 46570, BG-L-99902). C & D, corticolous; C, on Fagus sylvatica, Slovakia (JV 11397); D, on Alnus incana, Norway (TT 26205, BG-L-70280). Scales: A, B & D=2 cm; C=0·5 cm. Photographs by Kim Abel.
Thallus crustose, very variable, rimose-cracked to reticulate, delimited, occasionally sorediate; overall colour in saxicolous habitats from light grey to dark grey or brown, in corticolous habitats greyish or brownish green to olive-green. Areoles discrete, irregular, convex, highly variable in size, becoming secondarily cracked. Soralia rarely present, yellowish, sometimes tinged with brown, bursting from the apices of the areoles. Prothallus distinct, dark brown or black visible, ramifying the thallus, often forming mosaics. Photobiont Apatococcus fuscideae A. Beck & Zahradn., green, coccoid, globose to broadly ellipsoid.

Fig. 3 Phylogenetic relationships of esorediate and sorediate, saxicolous and corticolous specimens of Fuscidea cyathoides, shown here as a 50% majority-rule consensus tree of a B/MCMC analysis based on the concatenated data set (–ln=8081·83) of mtSSU, LSU and ITS. Posterior probabilities (PP) are displayed above, and MP and ML bootstrap values are displayed below, the branches; asterisks indicate a value of 100%. A circle indicates incongruent topology with the ML tree.
Apothecia immersed to sessile, constricted at base, roundish, up to 1·4 mm diam., to 1·9 mm when tuberculate, dark grey-brown to black; margin paler or concolorous with disc, rounded to strongly flexuous; disc black, mostly flat. Epihymenium brown; hymenium pale or faintly brownish; hypothecium hyaline. Asci clavate, of the Fuscidea-type. Ascospores simple, colourless, sometimes elliptical when young, bean-shaped when mature, brownish (6·0–)10·0–11·0(–14·5)×(3–)4–5(–7) μm.
Pycnidia abundant, brown, immersed to emergent with a thin thalline rim. Conidia bacilliform 3–4×1·5–2·0 μm.
Chemistry. Fumarprotocetraric acid (major), protocetraric acid (trace, usually present). Spot tests: K+ orange-yellow, Pd+ rust red, UV−.
Distribution and ecology. Fuscidea cyathoides is mainly saxicolous on coarse-grained, nutrient-deficient, siliceous rocks; occasionally it is corticolous on trunks and branches of Acer, Alnus, Betula, Castanea, Fagus, Ilex (B. Coppins, pers. comm.), Quercus and Sorbus.
The typical form (saxicolous esorediate) of Fuscidea cyathoides has been reported from Austria (Hafellner & Türk Reference Hafellner and Türk2001), Belgium and Luxembourg (Diederich & Sérusiaux Reference Diederich and Sérusiaux2000), the British Isles (Hawksworth et al. Reference Hawksworth, James and Coppins1980; Gilbert et al. Reference Gilbert, Purvis, Skjolddal and Tønsberg2009), China (Wei Reference Wei1991), Croatia (Partl Reference Partl2009), Czech Republic (Vězda & Liška Reference Vězda and Liška1999), Denmark (Søchting & Alstrup Reference Søchting and Alstrup2008), Estonia (Randlane & Saag Reference Randlane and Saag1999), Finland (Nordin et al. Reference Nordin, Moberg, Tønsberg, Vitikainen, Dalsätt, Myrdal and Ekman2010), France (Roux Reference Roux2012), Germany (Wirth Reference Wirth1987), Greenland (Thomson Reference Thomson1997), Italy (Puntillo Reference Puntillo1996), Morocco (Egea Reference Egea1996), Norway (Nordin et al. Reference Nordin, Moberg, Tønsberg, Vitikainen, Dalsätt, Myrdal and Ekman2010), Poland (Faɫtynowicz Reference Faɫtynowicz1993), Portugal (van den Boom & Giralt Reference van den Boom and Giralt1999; Llimona & Hladun Reference Llimona and Hladun2001), Romania (Ciurchea Reference Ciurchea1998), Russia (Urbanavichus & Andreev Reference Urbanavichus and Andreev2010), Serbia (Savić & Tibell Reference Savić and Tibell2006), Slovakia (Pišút et al. Reference Pišút, Guttová, Lackovičová and Lisická1998), Slovenia (Suppan et al. Reference Suppan, Prügger and Mayrhofer2000), Spain (Llimona & Hladun Reference Llimona and Hladun2001), Sweden (Nordin et al. Reference Nordin, Moberg, Tønsberg, Vitikainen, Dalsätt, Myrdal and Ekman2010), Switzerland (Clerc Reference Clerc2004), Turkey (Yildiz et al. Reference Yildiz, John and Yurdakul2002) and Ukraine (Kondratyuk et al. Reference Kondratyuk, Dymytrova and Nadyeina2010). The saxicolous sorediate form is known from the British Isles (Hawksworth et al. Reference Hawksworth, James and Coppins1980), Denmark (Søchting & Alstrup Reference Søchting and Alstrup2008), France (Roux Reference Roux2012), Poland (Faɫtynowicz Reference Faɫtynowicz1993), Norway (Poelt & Buschardt Reference Poelt and Buschardt1978; and material to be published), and Sweden (Nordin et al. Reference Nordin, Moberg, Tønsberg, Vitikainen, Dalsätt, Myrdal and Ekman2010) (see Fig. 4). The records from North America and Tasmania (Richardson & Richardson Reference Richardson and Richardson1982; Egan Reference Egan1987) were later rejected as they were based on misidentifications (Kantvilas Reference Kantvilas2001; Fryday Reference Fryday2008).

Fig. 4 Distribution of saxicolous, esorediate and sorediate forms of Fuscidea cyathoides based on the material examined and the literature.
The corticolous form has been reported from Albania (Svoboda et al. Reference Svoboda, Bouda, Malíček and Hafellner2012), Austria (Hafellner & Türk Reference Hafellner and Türk2001), Belgium and Luxembourg (Diederich & Sérusiaux Reference Diederich and Sérusiaux2000), Bosnia-Herzegovina (Christensen Reference Christensen1994), the British Isles (Gilbert et al. Reference Gilbert, Purvis, Skjolddal and Tønsberg2009; B. Coppins, pers. comm.), Croatia (Partl Reference Partl2009), Denmark (Søchting & Alstrup Reference Søchting and Alstrup2008), France (Roux Reference Roux2012), Germany (Cezanne et al. Reference Cezanne, Eichler and Wirth2004), Italy (Tretiach & Nimis Reference Tretiach and Nimis1994), Poland (Faltynowicz 1993), Portugal (van den Boom & Giralt Reference van den Boom and Giralt1999; Llimona & Hladun Reference Llimona and Hladun2001), Montenegro (Knežević & Mayrhofer Reference Knežević and Mayrhofer2009), Norway (Nordin et al. Reference Nordin, Moberg, Tønsberg, Vitikainen, Dalsätt, Myrdal and Ekman2010), Russia (Urbanavichus & Andreev Reference Urbanavichus and Andreev2010), Slovakia (Bielczyk et al. Reference Bielczyk, Lackovičová, Farkas, Lőkös, Breuss and Kondratyuk2004), Slovenia (Suppan et al. Reference Suppan, Prügger and Mayrhofer2000), Spain (Llimona & Hladun Reference Llimona and Hladun2001), Sweden (Nordin et al. Reference Nordin, Moberg, Tønsberg, Vitikainen, Dalsätt, Myrdal and Ekman2010), Switzerland (Clerc Reference Clerc2004) and Ukraine (Coppins et al. Reference Coppins, Kondratyuk, Khodosovtsev, Zelenko and Wolseley2005) (see Fig. 5), as well as from Taiwan (Aptroot & Sparrius Reference Aptroot and Sparrius2003).

Fig. 5 Distribution of corticolous forms of Fuscidea cyathoides based on the material examined and the literature.
Specimens examined (saxicolous, esorediate). Czech Republic: Central Bohemia: Distr. Beroun, Brdy Mts, Neřežín - Malá Víska, upper part of Krkavčina Mt., forested (Picea, Betula, Larix etc.) rocky hill, 49°45'55''N, 13°53'36''E, alt. 570–600 m, on siliceous boulder, 2012, J. Malíček 4916; Distr. Beroun, Brdy Mts, Neřežín, Jindřichova skála Mt., 1 km SE of Malá Víska, rock with E-exposed boulder scree, 49°46'05''N, 13°52'55''E, alt. 550–580 m, on siliceous boulder, 2012, J. Malíček 4928. Western Bohemia: Distr. Rokycany, Brdy Mts, Strašice - Lipovsko Mts (651 m), 3 km SE of town, rock with boulder scree on S-exposed slope, 49°42'53''N, 13°47'11''E, alt. 620–640 m, on siliceous rock, 8 xi 2012, J. Malíček (4866). Moravský kras: Mohelno, 49°06'08·80''N, 16°11'05·20''E, alt. 344 m, in deciduous forest on shaded siliceous rock, 2011, J. Halda 662/2011 (JHP/13294).—Norway: Hordaland: Fjell, Sotra, SW of Landro, Ingholet, 100 m from cemetery, 60°25'6''N, 4°58'31·2''E, alt. 35–45 m, saxicolous on SW-facing vertical siliceous stone wall, 2011, M. Zahradníková MZ 30 (BG-L-96933); Fjell, Sotra, W of the road between Skålvik and Sekkingstad, S of road jct to Algrøyna, 60°20'06·6''N, 4°59'42·0''E, alt. 40–70 m, saxicolous on siliceous rock wall in coastal heath, 2010, M. Zahradníková MZ 5 (BG-L-96931). Nordland: Nesna, Tomma Island, Valhaugen, 66°17'44·63''N, 12°49'15·31''E, alt. 35 m, saxicolous on bedrock in open, treeless situation, 2016, T. Tønsberg 46572 & A. Botnen (BG-L-99904).—Ireland: Co. Kerry: Macgillycuddy’s Reeks, Gaddagh River valley NE of Carrauntoohil (Corrán Tuathail) [1039 m], c. 14 km WSW of Killarney, 52°00'50·0''N, 9°42'49·0''W, alt. 225 m, on boulders near the brook, 2003, J. Halda & Z. Palice 7903.—Great Britain: Scotland: V.C. 92, South Aberdeenshire: Braemar, Invercauld Estate, Craig Leek, NE-E facing crags, partly limestone, 57°01'24·0''N, 3°39'60·0''W, alt. 425 m, on siliceous rock in pasture below crags, 2005, A. M. Fryday 9012 (MSC0050557).—Sweden: Skåne: S. Mellby, Stenshuvud, på block i strandskogen, 1987, S. Ekman 265 (LD–1132977).
Specimens examined (corticolous): Norway: Rogaland: Forsand, N side of Mt. Uburen by Forsandåna, alt. 60–80 m, on Betula pubescens in boulder field, 29 viii 2001, J. I. Johnsen (BG-L-89616); Sokndal, S of Årstad, alt. 60–80 m, on Betula pubescens, 25 viii 2010, J. I. Johnsen (BG-L-89638).—Slovakia: Bukovské Mts: Nová Sedlica, protected area Stužica, valley of Stužická Rieka, natural deciduous forest, 49°04'23''N, 22°32'25''E, alt. 608–700 m, on bark of Fagus sylvatica, 2013, J. Malíček & J. Vondrák 6488; Nová Sedlica, beech forest on the crest Čiertáž - Hrúbky - Kremenec, 49°05'34·2''N, 22°22°31'36·6''E, alt. 1110 m, on bark of Fagus, 2004, Z. Palice & J. Šárová 9629; Nová Sedlica, protected area Stužica, 49°4'24''N, 22°32'35''E, alt. 600–1200 m, on bark of Acer pseudoplatanus, 2013, J. Vondrák & J. Malíček 11411; Ulič, Nová Sedlica, protected area Stužica, 49°04'24''N, 22°32'35''E, alt. 600–1200 m, on bark of Fagus sylvatica, 2013, J. Vondrák & J. Malíček 11397; Ulič, Nová Sedlica, protected area Stužica, 49°04'24''N, 22°32'35''E, alt. 1000–1200 m, on bark of Fagus sylvatica, 2013, J. Vondrák & J. Malíček 11476. Muránska planina: Nová Maša, alder stand along unnamed stream in parallel with Rácov brook, 48°48'45–50''N, 20°01'45''E, alt. 770–780 m, on bark of Alnus incana, 1999, A. Guttová, V. Orthová & Z. Palice 4642.—Sweden: Skåne: Vittsjö, N end of Vittsjö, on roadside Fagus near bridge, 1987, U. Arup & S. Ekman L035 (LD–1157864); N Åkarp, 2·5 km S Bjärnum, c. 500–700 m SSW of Lake Agnsjön, W of road, on Fagus, 1987, U. Arup & S. Ekman L036 (LD–1157444); Tåssjö par., Hålskutt, 1988, U. Arup & S. Ekman L130 (LD–1131717); S. Mellby, Stenshuvud, på rönn i Ö-branten, 1987, S. Ekman L146 (LD–1133157).
Specimens examined (saxicolous, sorediate): Norway: Nordland: Nesna, Tomma, Valhaugen, 66°17'44·77''N, 12°49'15·31''E, alt. 35–40 m, saxicolous in shallow crevice (with seeping water) in bedrock in open, treeless situation, 2016, T. Tønsberg 46570 (BG-L-99902).
We thank the curators of HO, LD, UPS, H-Ach, and MSC for the loan of material, Zdeněk Palice, Jan Vondrák, Jiří Malíček (all from the Academy of Science of the Czech Republic) and Josef Halda (Museum and Gallery of Orlické hory Mts., Czech Republic) for providing fresh material from Central Europe, M. R. D. Seaward (University of Bradford, England) for language improvements, Louise Lindblom, Beate Helle and Per M. Jørgensen (all University of Bergen, Norway) for technical help with the molecular work (LL), technical help with the preparation of the distribution maps (BH) and help with botanical nomenclature (PMJ), respectively, and Kim Abel (Røyken, Norway) for taking the photographs. The project was funded by the University Museum of Bergen, University of Bergen, including grants from the Grolle Olsen fund. This work is part of the doctoral thesis of MZ. The molecular work was carried out in the Biodiversity Laboratories at the University of Bergen.
Supplementary material
For supplementary material accompanying this paper visit https://doi.org/10.1017/10.1017/S0024282917000524










