


Introduction
The pre-Pliocene fossil record of North American newts (Salamandridae, Tarichina) is sparse, especially west of the Rocky Mountains, yet it extends far back into the Oligocene (32–33 Ma). Previous morphological comparisons with extinct and extant North American salamanders recognized at least two species of fossil salamanders from Oregon’s early Oligocene deposits: Taricha oligocenica of the Mehama Formation in western Oregon (Van Frank, Reference Van Frank1955), and Taricha lindoei of the John Day Formation in eastern Oregon (Naylor, Reference Naylor1979). Van Frank’s T. oligocenica and Naylor’s T. lindoei are found in beds assigned to the Rupelian Stage (30–33 Ma; early Oligocene; dates by Retallack et al., Reference Retallack, Bestland and Fremd2000, Reference Retallack, Orr, Prothero, Duncan, Kester and Ambers2004), suggesting correspondence with the Orellan NALMA of the early Oligocene for T. oligocenica and T. lindoei (Albright et al., Reference Albright, Woodburne, Fremd, Swisher, MacFadden and Scott2008). Naylor (Reference Naylor1982) described another Taricha specimen from the early Oligocene (32–33 Ma) Gumboot Mountain strata in Washington, but to genus only. Farther east, vertebrae from a third species, Taricha miocenica, have been uncovered in Montana’s late Oligocene (Arikareean; 24–26 Ma) Middle Cabbage Patch Beds (Tihen, Reference Tihen1974; Rasmussen, Reference Rasmussen1977; Rasmussen and Prothero, Reference Rasmussen and Prothero2003), while fossil trackways from the late Miocene of California and the late Miocene of Kansas have also been identified as Taricha (Peabody, Reference Peabody1959; Holman, Reference Holman2006). Although we did not examine any fossils of T. miocenica in this study, Tihen (Reference Tihen1974) proposed the grouping of T. oligocenica and T. miocenica into the subgenus Palaeotaricha (not recognized in the latest classifications; Estes, Reference Estes1981; Holman, Reference Holman2006; Dubois and Raffaëlli, Reference Dubois and Raffaëlli2009). If the latest classifications are correct, then T. miocenica would represent one of the two occurrences of Taricha (along with the Kansas Miocene trackways) east of the Rocky Mountains.
Several previously described, partial to nearly complete skeletons from Oligocene localities in the John Day and Mehama formations of Oregon represent excellently preserved, post–metamorphic individuals of the genus Taricha, also known as Pacific (or Western) newts (Van Frank, Reference Van Frank1955; Naylor, Reference Naylor1979; Holman, Reference Holman2006; Fremd, Reference Fremd2010; McClaughry et al., Reference McClaughry, Wiley, Ferns and Madin2010). Additional undescribed specimens include fossils from new localities near Gray Butte, Oregon and Lyons, Oregon, and a large individual from Goshen, Oregon. This study examines the new specimens and redescribes previously examined material. We also investigate the evolutionary relationships of the family Salamandridae in North America, because the migration of salamandrids to North America and their subsequent divergence into the genera Taricha and Notophthalmus is poorly understood (Tihen, Reference Tihen1974; Naylor, Reference Naylor1979; Naylor and Fox, Reference Naylor and Fox1993).
We identify Oregon’s fossil salamanders and examine diagnostic skeletal characters for use in future phylogenetic analyses with extinct and extant North American salamandrids. Furthermore, we examine previously described fossil Taricha to determine whether these specimens are truly separate species, morphological variations within a single species, or if they represent a distinct genus from extant Pacific newts. We incorporate new, previously undescribed specimens from various Oregon localities, along with the previously described specimens of T. oligocenica (Van Frank, Reference Van Frank1955) and T. lindoei (Naylor, Reference Naylor1979). By pursuing these goals, we aim to answer the following questions: Are previous identifications and descriptions of the Oligocene fossil salamanders of Oregon adequate? How do these fossil salamanders relate to each other and to extant taxa? Can we gain any insight on the functional morphology of these newts? Finally, what can the paleoecology and distribution of these salamanders tell us about the distribution and evolution of North American salamandrids?
Geologic setting
The southern Willamette Valley of Oregon is an alluvial basin bordered to the west by the Coast Range and to the east by the Western Cascades physiographic provinces. The basin is within the seismically active Puget–Willamette Lowland, and is situated along the Cascadia subduction zone, where the Juan de Fuca plate has been subducting obliquely beneath the North American continental plate for much of the Cenozoic (McClaughry et al., Reference McClaughry, Wiley, Ferns and Madin2010). This has resulted in a complicated forearc basin that has locally accumulated >7,000 m of sedimentary and volcanic strata since the Paleocene (Retallack et al., Reference Retallack, Orr, Prothero, Duncan, Kester and Ambers2004; McClaughry et al., Reference McClaughry, Wiley, Ferns and Madin2010). The exposed rocks of the southern Willamette Valley include a diverse series of Paleogene marine sedimentary rocks with submarine lavas, Paleogene and Neogene terrestrial volcanic and sedimentary rocks and intrusions, and Quaternary surficial deposits (Retallack et al., Reference Retallack, Orr, Prothero, Duncan, Kester and Ambers2004; McClaughry et al., Reference McClaughry, Wiley, Ferns and Madin2010). The Mehama Formation of the Little Butte Volcanics, within which the Willamette and Lyons floras and all Western Oregon salamander fossils are preserved, is locally fossiliferous and represents generally nondescript volcaniclastic sedimentary rocks that occur between the more distinctive features of the Little Butte Volcanics, such as mapped formations, marker beds, and volcanic centers (McClaughry et al., Reference McClaughry, Wiley, Ferns and Madin2010).
Along with the latest Eocene–earliest Oligocene Goshen flora (34.85–32.8 Ma, 40Ar/39Ar dates), the early Oligocene Willamette flora (>30.64 Ma) is regarded as a critical indicator of climatic and vegetational shift across the Eocene–Oligocene transition (Retallack et al., Reference Retallack, Orr, Prothero, Duncan, Kester and Ambers2004; Retallack, Reference Retallack2008). Thick lahars separate the dated ash from the lake beds of the Willamette flora, which is found between Spores Point tuff (31.3±0.6 Ma) and a Little Butte Volcanics tuff (30.6 ± 0.5 Ma; Myers et al., Reference Myers, Kester and Retallack2002; Retallack et al., Reference Retallack, Orr, Prothero, Duncan, Kester and Ambers2004; McClaughry et al., Reference McClaughry, Wiley, Ferns and Madin2010). The presence of thick andesitic debris-flow deposits suggests that the Willamette Flora (and the salamanders found within it) grew in an unstable environment adjacent to the increasingly active Western Cascades (Myers et al., Reference Myers, Kester and Retallack2002).
The dark gray to whitish-tan shales of the Mehama Formation produce vertebrate specimens alongside numerous plant fossils. The presence of fossil Taricha agrees with the interpretation of the bedded shales as lake-margin deposits because this genus and the majority of this family feed in a freshwater environment; species of the genus Taricha prefer slow-moving or still water conditions (Wake and Özeti, Reference Wake and Özeti1969; Goin et al., Reference Goin, Goin and Zug1978; Naylor, Reference Naylor1978a, Reference Naylor1978b, Reference Naylor1979; Retallack et al., Reference Retallack, Orr, Prothero, Duncan, Kester and Ambers2004; Pollett et al., Reference Pollett, MacCracken and MacMahon2010).
The Lyons flora, located in northwestern Oregon (southeast of the town of Lyons, Oregon in the upper Thomas Creek area), is comprised of at least 24 plant taxa, 12 of which are identifiable to species, while the other 12 are identifiable to genus. The rocks within which the leaves are preserved are composed of thinly laminated, tuffaceous material that has been silicified (Meyer, Reference Meyer1973). The stratification of the beds, abundance of fossil leaves, and the presence of a water plant (Nymphoides) in the fossil record suggest a shallow, quiet lacustrine environment (Meyer, Reference Meyer1973). Retallack et al. (Reference Retallack, Orr, Prothero, Duncan, Kester and Ambers2004) have listed the Lyons flora among the forested Oligocene lacustrine floras. The Lyons flora has produced one definite salamander fossil, consisting of part and counterpart, thus far.
The John Day Formation is a complex series of strata with a unique record of the long-term change (~20 Myr) of climate and biotas for the Pacific Northwest of North America (Fremd, Reference Fremd2010). The John Day Formation occurs within a number of basins in north-central and central Oregon, all within the Blue Mountains physiographic province (Fremd, Reference Fremd2010).
The Big Basin Member of the lower John Day Formation has distinctive red and tan claystones and contains excellent paleobotanical localities, which also produce the vast majority of caudate fossils from eastern Oregon. Radiometric dates indicate that the Big Basin Member preserves ~9 Myr of deposition, from ~30–39 Ma (Bestland and Retallack, Reference Bestland and Retallack1994; Retallack et al., Reference Retallack, Bestland and Fremd2000). The Bridge Creek Flora type localities, which are near the Eocene-Oligocene boundary, are within the lacustrine shales of the Big Basin Member in the Painted Hills Unit of John Day Fossil Beds National Monument, where fish and amphibian fossils have also been recovered (Meyer and Manchester, Reference Meyer and Manchester1997; Retallack et al., Reference Retallack, Bestland and Fremd2000; Dillhoff et al., Reference Dillhoff, Dillhoff, Dunn, Myers and Strömberg2009; Fremd, Reference Fremd2010). The Bridge Creek flora’s age is well dated (Bestland and Retallack, Reference Bestland and Retallack1994; Retallack et al., Reference Retallack, Bestland and Fremd2000) because it is sandwiched between the underlying Biotite Tuff (32.99±0.11 Ma) and the overlying Overlook Tuff (32.66±0.03 Ma).
The fossil salamanders found in the Big Basin Member of the John Day Formation have been unearthed at the Wheeler County High School exposures in the town of Fossil, Wheeler County, Oregon (Naylor, Reference Naylor1979). Plant, vertebrate, and invertebrate fossils preserved in these rocks are consistent with a cooler temperate climate than found in the Willamette flora, with fewer Eocene subtropical holdovers present (Meyer and Manchester, Reference Meyer and Manchester1997; Retallack, Reference Retallack2008; Dillhoff et al., Reference Dillhoff, Dillhoff, Dunn, Myers and Strömberg2009). Salamander fossil-bearing beds are between 32.66 Ma and 32.99 Ma, placing these fossils solidly in the Orellan NALMA (Bestland and Retallack, Reference Bestland and Retallack1994; Retallack et al., Reference Retallack, Bestland and Fremd2000; Albright et al., Reference Albright, Woodburne, Fremd, Swisher, MacFadden and Scott2008).
The smallest fossil salamander, and most recent acquisition for this study, is from a new locality for vertebrate fossils. The area, near Gray Butte and the town of Prineville in central Oregon, has produced a limited fossil flora in the past. Based on the work of Thormahlen (Reference Thormahlen1984) and Smith et al. (Reference Smith, Manchester, Ashwill, McIntosh and Conrey1998), the location of this specimen coincides with the Trail Crossing flora, which to date has produced Acer, Alnus, Fagus, and Quercus fossils from ashy, tan to yellow, massive to stratified, highly silicified lacustrine siltstone and sandstone beds. These beds are consistent with the temperate, hardwood-dominated Bridge Creek flora of the Oligocene. The fossils from Gray Butte are between 32–33 Ma because the overlying alkali-olivine basalt is 40Ar/39Ar-dated at 32.49 ± 0.30 Ma, and the lithology of the Gray Butte area aligns with the middle Big Basin Member of the western facies of the John Day Formation.
Materials and methods
Systematic paleontology and comparative morphology
For this study, we re-examined all the previously described specimens of fossil salamanders from the Oligocene of Oregon, as well as a number of previously undescribed specimens and the skeletons of several extant salamandrids. This included comparing them to modern salamander taxa in order to understand caudate evolutionary relationships during a period of climatic cooling, floral transition, and perhaps increased seasonality during the early Oligocene in the Pacific Northwest of North America. The specimens are in collections at the University of Oregon Museum of Natural and Cultural History (UOMNH), University of California-Berkeley Museum of Paleontology (UCMP), and John Day Fossil Beds National Monument (JODA).
Table 1 contains a list of the specimens examined. We examined skeletons and isolated bones of eight Taricha granulosa, a single Taricha sierrae, seven Taricha torosa, 12 Taricha rivularis, and four Notophthalmus viridescens, in addition to the fossils of five T. oligocenica, 12 T. lindoei, and one fossil identified to genus Taricha. For all examined specimens, we recorded morphologic details of the scapulocoracoid, phalangeal count, and vertebral column, especially the atlas and trunk vertebrae, since these features have been noted as useful for diagnosing fossil salamanders in the past (Estes, Reference Estes1981; Holman, Reference Holman2006). We paid special attention to details of the skull when possible because it is rare to find a complete or mostly complete salamander skull in the fossil record, where skull fragments and postcrania (particularly trunk vertebrae) are much more common than articulated specimens. The systematic descriptions provide details of the morphology of these structures. We coded several of these details into the phylogenetic analysis as characters.
Table 1 List of specimens with collection numbers. Dagger symbol (†) indicates extinct taxa.

Because each fossil preserves different areas of the skeleton, establishing useful measurements is a complex task. For example, the vertebral measurements used in Tihen (Reference Tihen1974) are difficult to apply to fossils that are articulated skeletons, impressions, or flat (as are so many of the relatively complete salamander fossils). Instead, this study uses a variety of measurements, as applied by Good and Wake (Reference Good and Wake1992) to the morphometrics of the genus Rhyacotriton and by Kuchta (Reference Kuchta2007) to the morphometrics of T. torosa and T. sierra, whenever possible. While the small sample size and limited variety of age classes for fossils make it unlikely that we can perform a morphometric analysis in the manner of Good and Wake (Reference Good and Wake1992) and Kuchta (Reference Kuchta2007), these measurements are still useful for determining body proportions and predicting the size of individual specimens. These measurements include snout-vent length (SVL), tail length (TL), snout-gular length (SG), head width (HW), eye width (EW), distance between the eyes (E-E), distance from eye to nostril (E-N), axilla-groin length (AG), trunk width (TW), forelimb length (FLL), hind limb length (HLL), and foot length (FL). We used digital calipers or a metric ruler at the millimeter scale for all attainable measurements for each fossil. Some of these measurements (SVL, SG, and AG) use soft-tissue reference points (Good and Wake, Reference Good and Wake1992; Kuchta, Reference Kuchta2007), and we adjust these to use osteological landmarks that represent corresponding measurements for fossil taxa. Here, the anterior edge of the premaxilla to the posterior edge of the pelvic girdle is SVL, the anterior edge of the premaxilla to the anterior portion of the atlas is SG, and the length of the trunk between the forelimb and the hind limb is AG.
We took all University of Oregon (UO) and John Day Fossil Beds National Monument (JODA) fossil salamander photos with a Nikon D-90 at a focal length of 90 mm. We photographed all University of California–Berkeley specimens using either a Nikon optiphot2-pol with a Nikon digital sight ds-fi2 attachment, or a Nikon Coolpix L24 digital camera.
Phylogenetic analysis
Our matrix expands upon the matrix of Marjanović and Witzmann (Reference Marjanović and Witzmann2015), with the addition and/or adjustment of a few characters (Supplementary Data Sets 1, 2). Many of the characters in our matrix also use characters (Supplementary Data Set 1) first established in Wake and Özeti (Reference Wake and Özeti1969), Titus and Larson (Reference Titus and Larson1995), Venczel (Reference Venczel2008), and Schoch and Rasser (Reference Schoch and Rasser2013). Naylor (Reference Naylor1978b) made a few adjustments to the morphology-based phylogeny of Wake and Özeti (Reference Wake and Özeti1969), some of which are adopted in this analysis. We also added a few characters of our own to test their usefulness for analyzing North American taxa specifically (specimen availability did not allow us to make clear observations on taxa outside of North America for the purpose of coding these characters). We coded other characters relating to non-North American taxa according to the observations of Wake and Özeti (Reference Wake and Özeti1969), Titus and Larson (Reference Titus and Larson1995), Venczel (Reference Venczel2008), Schoch and Rasser (Reference Schoch and Rasser2013), Naylor (Reference Naylor1978b), and Marjanović and Witzmann (Reference Marjanović and Witzmann2015). We included a variety of characters, comprising skeletal, behavioral (including reproductive), soft-tissue, and geographic characters in this matrix, and we coded soft-tissue and behavioral characters as unknown for fossil taxa.
We assembled the matrix of this study in Mesquite 2.75 (Maddison and Maddison, Reference Maddison and Maddison2011), then ran a heuristic analysis using the Phylogenetic Analysis Using Parsimony program Version 4.0a152 (PAUP; Swofford, Reference Swofford2002) to produce simple parsimony trees. We elected to use simple parsimony for our analyses because they contained multistate, ordered, and unordered characters. We analyzed a total of 108 characters and a total of 40 taxa in a matrix using all characters, including both unconstrained and molecularly constrained (via consensus data assembled in Marjanović and Witzmann, Reference Marjanović and Witzmann2015) from Zhang et al. (Reference Zhang, Papenfuss, Wake, Qu and Wake2008) and Pyron (Reference Pyron2014) analyses. With 108 characters (33 ordered), this analysis is the largest morphological matrix used for analyzing salamandrid evolutionary relationships to date. In another round of heuristic analyses, we removed fossil newt taxa known only from vertebrae (Koalliella, Taricha miocenica, Notophthalmus crassus [in this case, including the fossils sometimes assigned to Notophthalmus slaughteri; Holman, Reference Holman2006], and Notophthalmus robustus) prior to running unconstrained and molecularly constrained analyses. We coded characters inapplicable or unknown for a taxon (e.g., soft tissue characters when coding fossil taxa) with question marks. Bracketed numbers indicate a character’s original number from its referenced source (Supplementary Data Sets 1, 2). We performed a heuristic search with 1000 repetitions of stepwise addition with random addition sequence using the sectorial search algorithm setting, and TBR (tree bisection and reconnection), with all characters weighted equally. We calculated the Consistency Index, Retention Index, and Homoplasy index, as well as strict consensus trees, in PAUP.
Ambystoma and Dicamptodon are outgroups for our analyses. Within Taricha, T. sierrae was combined with T. torosa because there was not enough morphological character data available, and few positively identified skeletal specimens available to distinguish the species from T. torosa. As such, T. sierra as presently understood scores identically to T. torosa in our morphological phylogenies. Because of our lack of data, we did not include Notophthalmus slaughteri because it may be synonymous with Notophthalmus crassus, and we have no direct observations of either potential species (Holman, Reference Holman2006).
We modified the assigned character states for taxa in a few cases for previously used characters based on personal observations and/or previously published literature; this is particularly true for Tylototriton in character 71, and character 61 for extant species of Taricha. Character 71 refers to the presence of ribs either as short rods (0) or as long as three vertebral centra (1), yet in at least Tylototriton verrucosus, the ribs do not quite exhibit character state 1 (based on Nussbaum and Brodie, Reference Nussbaum and Brodie1982) because they are closer to the length of two vertebral centra, while Tylototriton asperrimus was initially assigned to Echinotriton partially because their trunk ribs fit character state 1 (Nussbaum et al., Reference Nussbaum, Brodie and Datong1995). We assigned both states 0 and 1 to Tylototriton as a genus. The length of the ribs in Taricha oligocenica, and to a lesser extent, Taricha lindoei, are visually similar to those of Tylototrion verrucosus, both in terms of length and in the number, structure, and orientation of the epipleural processes, which we discuss briefly later. We therefore assigned a character state of 0 to these taxa, but recommend further investigation of this character as a whole, because the ribs of Tylototriton verrucosus and Taricha oligocenica do not quite fit either character state (longer than simple, short rods, but shorter than described in state 1).
Repositories and institutional abbreviations
JODA, John Day Fossil Beds National Monument; UALVP, University of Alberta Laboratory for Vertebrate Paleontology; UCMP, University of California – Berkeley Museum of Paleontology; UCMP, University of California – Berkeley Museum of Vertebrate Zoology; UOMNH, University of Oregon Museum of Natural History.
Systematic paleontology
Class Amphibia Gray, Reference Gray1825
Subclass Lissamphibia Haeckel, Reference Haeckel1866
Order Caudata Scopoli, Reference Scopoli1777
Suborder Salamandroidea Fitzinger, Reference Fitzinger1826
Family Salamandridae Goldfuss, Reference Goldfuss1820
Genus Taricha Gray, Reference Gray1850
Type species
Taricha torosa (Rathke, Reference Rathke1833).
Diagnosis
Osteologic characters of extant Taricha include a low to moderately high neural spine without an extensive pitted dermal cap (Wake and Özeti, Reference Wake and Özeti1969; Naylor, Reference Naylor1978b). The genus Taricha also possesses concave inter-prezygapophyseal neural arch margins (Boardman and Schubert, Reference Boardman and Schubert2011). The premaxillae are fused, nasals are separated, the rib processes do not extend past the body wall (no costal grooves), anterior caudal ribs (caudosacral ribs of Estes, Reference Estes1981) are absent, and the cotyles of the vertebrae appear horizontally oval (Estes, Reference Estes1981). The hyobranchium possesses a cartilaginous, mineralized first basibranchial and second ceratobranchial, but lacks the second basibranchial and anterior radii. Taricha also has a reduced interradial cartilage (Estes, Reference Estes1981).
Remarks
Taricha’s closest relative is Notophthalmus, the only other North American newt genus. It is important to note that extinct species Taricha oligocenica (Van Frank, Reference Van Frank1955) and Taricha miocenica (Tihen, Reference Tihen1974) possess pitted spine tables on the dorsal side of their vertebrae, leading to questions about the ancestral state of North American newts. Weaver (Reference Weaver1963) determined that the shape of the vomerine (then incorrectly thought to be “prevomerine”; Atkins and Franz-Odendaal, Reference Atkins and Franz-Odendaal2015; Marjanović and Witzmann, Reference Marjanović and Witzmann2015) tooth row of extant Taricha is distinguishably different between Taricha granulosa, which displays the plesiomorphic V-shaped arrangement, and Taricha torosa, Taricha sierra, and Taricha rivularis, which all display a Y-shaped pattern.
Taricha oligocenica Van Frank, Reference Van Frank1955

Figure 1 Dorsal view of UOMNH F-5405, type specimen of T. oligocenica as a photograph (1) and a line drawing (2).

Figure 2 Possible juveniles of T. oligocenica. UOMNH F-36412: (1) line drawing and (2) photograph; (3) photograph of UOMNH F-30648.

Figure 3 Ventral view of UOMNH F-55196, the largest individual of T. oligocenica. (1) Close-up of the trunk, showing the diagnostic epipleural rib processes; (2) right pes (left side), which is fully ossified, a condition not commonly seen in extant Taricha; (3) full ventral view of UOMNH F-55196; (4) line drawing of right ribs, ventral view, with epipleural processes clearly visible; (5) line drawing of fully ossified left pes.

Figure 4 Nearly complete skeleton of UOMNH F-59812 A–B, found in association with the Lyons flora. (1) Full view of part and counterpart; (2) view of the trunk of F-59812 showing soft-tissue preservation and preservation of rib processes; (3) fully ossified right pes of F-59812 A; (4) line drawing of right pes of F-59812 A.

Figure 5 Line drawings for comparison of trunk (1, 2) and atlas (3, 4) morphology between T. oligocenica and T. torosa in lateral view, anterior to the left. (1) Taricha oligocenica trunk vertebra; (2) T. torosa trunk vertebra; (3) atlas of T. oligocenica; (4) atlas of T. torosa.
1955 Palaeotaricha oligocenica Van Frank in Reference Van FrankVan Frank, p. 4, pl. 1–3.
1974 Taricha (Palaeotaricha) oligocenica Van Frank in Reference TihenTihen, p. 215.
1981 Taricha oligocenica Van Frank in Reference EstesEstes, p. 85.
Holotype
UOMNH F-5405, a mostly complete skeleton, early Oligocene, Mehama Formation, Lane County, Oregon, UO 11026 Goshen.
Diagnosis
Possesses a broad, rectangular, expansive dorsal spine table on the neural spines of the vertebrae (Fig. 5.1, 5.2; Tihen, Reference Tihen1974; Estes, Reference Estes1981; Holman, Reference Holman2006). Taricha oligocenica exhibits three apparently unique characteristics for the genus: (1) the atlantal neural crest of T. oligocenica is tall and extends laterally on the dorsal side of the vertebra towards the anterior end of the atlas, before forming a short anteroventral slope, followed by an abrupt ventral drop to the condylar facets (Fig. 5.3, 5.4); (2) T. oligocenica has an anteriorly narrowing, dorsal expansion on the top of the atlas (Fig. 5.3); and (3) T. oligocenica exhibits a narrow scapular portion of the scapulocoracoid (Van Frank, Reference Van Frank1955; Naylor, Reference Naylor1979, Reference Naylor1982; Holman, Reference Holman2006). Taricha oligocenica also possesses sculpturing in the dermal bones of the skull roof, particularly the frontals, parietals, squamosals, and frontosquamosal arch. Taricha oligocenica possesses elongate, distinct epipleural rib processes.
Occurrence
Early Oligocene, Oregon
Description
The posterior extension of the maxillae of UOMNH F-5405 and UOMNH F-30648 project nearly to the quadrate, similar to extant Taricha and much farther than in extant Notophthalmus. Preparation performed on the type specimen (UOMNH F-5405) after its initial publication revealed that the maxillary and quadrate elements do not fuse or suture in T. oligocenica. The premaxillae are fused and the paroccipital processes project strongly, abutting the posterior side of the squamosals. The bony frontosquamosal arches in all specimens appear to be more robust than in modern Taricha of similar size, and are complete; the skull and the frontosquamosal arches (best preserved in the type; Fig. 1) are also more sculptured than typically seen in extant Taricha. As in all Western newts, the opening of the cavum internasale is large (Van Frank, Reference Van Frank1955; Wake and Özeti, Reference Wake and Özeti1969).
The vertebral features are also listed in the diagnosis. The vertebrae are opisthocoelous with laterally extensive rectangular dorsal expansions that are broad, flat, and somewhat sculptured (Fig. 5.1). These spine tables extend over the entire dorsal surface of the trunk vertebrae, including the atlas, and abut each other both anteriorly and posteriorly (Figs. 4, 5). While previously only the type specimen preserved these spine tables, this character appears to be similarly expansive and identical in shape in the newer UOMNH F-59812 A-B (Fig. 4) specimen. The neural spines are high and elongate, and there are 14 presacral vertebrae. The atlas differs significantly from that of recent Taricha.
The ribs of T. oligocenica possess long epipleural processes that decrease in size towards the tail (Figs. 3.1, 3.4, 4.1, 4.2). Van Frank (Reference Van Frank1955) noted that the number of these epipleural rib processes varied in number by individual, but that these processes were consistently much shorter and less robust in extant Taricha than in the type of T. oligocenica (UOMNH F-5405); Naylor (Reference Naylor1979) noted that T. lindoei shares these long processes with T. oligocenica. These long processes are easily distinguishable in UOMNH F-55196 and UOMNH F-59812 A–B (Figs. 3.1, 3.4, 4.1, 4.2), more so than in F-5405, which was embedded in epoxy during preparation, obscuring the view of this character. There is no variation in this character in the sample examined.
Comparison of the forelimbs and hind limbs of T. oligocenica to extant taxa reveals little difference morphologically, and Van Frank (Reference Van Frank1955) asserted that limb proportions were the same between the type specimen and extant skeletons. However, the degree of ossification apparent in the specimens with limbs preserved (UOMNH F-5405, F-55196, and F-59812 A-B) is different than seen in extant Taricha, especially in the previously undescribed specimens. Specifically, T. oligocenica preserves fully ossified tarsals (Figs. 3.2, 3.5, 4.3, 4.4), which differs from extant Taricha, where the tarsals fully ossify only rarely, even in older individuals (Shubin et al., Reference Shubin, Wake and Crawford1995). This difference in tarsal ossification may indicate that UOMNH F-5405, F-55196, and F-59812 A–B are old individuals, or may represent a legitimate character differentiation in T. oligocenica. It seems unlikely that all specimens preserving the tarsals (UOMNH F-5405, F-55196, and F-59812 A–B) would be as old as extant Taricha individuals would have to be to preserve this pattern, which suggests that T. oligocenica ossifies this region differently than modern members of the genus. It can be alternatively argued that the lack of tarsal preservation in the smaller individuals (UOMNH F-36412 and F-30648; Fig. 2.2, 2.3) suggests a lack of ossified tarsals. The phalangeal count of extant Taricha is 1-2-3-2 (manus) and 1-2-3-3-2 (pes). This holds true in the fossils as well, as exhibited in F-5405, F-55196 (pes only; Fig. 3.2, 3.5), and F-59812 A–B (Fig. 4.3, 4.4). Van Frank (Reference Van Frank1955) described the pattern of bones in the tarsus (tarsal formula) in the type specimen of T. oligocenica as having an arrangement of 1, 2, 3, 4+5. We are unsure of his meaning on this, but we find that there appears to be a union of adjacent distal carpals four and five (dt 4 + 5); however, epoxy covering the ventral side of the specimen has yellowed with age and obscures that side, making it difficult to confirm if this is an original arrangement or a result of taphonomy. If Van Frank’s (Reference Van Frank1955) assessment is taken to portray our observations, and if neither are a result of taphonomy, this tarsus arrangement contrasts with the pattern typically seen in extant Taricha, which possesses a basale commune, with distal tarsal three, four, and five as separate elements. Our observation is, however, within variation for at least extant Taricha granulosa, even within a single population (Shubin et al., Reference Shubin, Wake and Crawford1995). While there is a preferred arrangement within a species (Shubin et al., Reference Shubin, Wake and Crawford1995), a large sample size is required to describe variation and determine a most common arrangement. With only two other specimens of T. oligocenica partially preserving bones of the tarsus (UOMNH F-55196 and F-59812 A–B), the sample size is not nearly large enough to undertake such a study at this time, and even within this sample, UOMNH F-59812 appears to have the formula seen in extant Taricha (Fig. 4.3). Given the variation found in extant populations and the small sample size of fossil specimens, this study does not use the tarsus bone arrangement as a character for use in phylogenetic analyses. The arrangement of the carpus bones is the same in both extant Taricha and specimens UOMNH F-5405, F-55196, and F-59812 A–B of T. oligocenica. The scapular portion of the scapulocoracoid is long and narrow compared to extant Taricha, as noted by Van Frank (Reference Van Frank1955), Naylor (Reference Naylor1979), Estes (Reference Estes1981), and Holman (Reference Holman2006).
In T. oligocenica, the total body lengths of the more complete fossils (F-5405, F-59812 A–B) appear to be between ten times the skull width and ten times the snout-gular length (Table 2). The type for T. oligocenica, which measures 120 mm, but is missing a little over a centimeter of the tail, is between 130–135 mm in total length. For this same specimen, the skull width is 13 mm, while the snout-gular length is 14 mm. Naylor (Reference Naylor1979) made similar calculations, determining that the total body length in extant Taricha was ~12 times the skull width, but these calculations were done with specimens that still possessed soft tissues, and therefore appear to over-predict the total body length of specimens with only skeletal material remaining.
Table 2 Measurements of extinct Taricha. Some specimens are too poorly preserved to perform accurate measurements. Approximate length calculated as between 10x skull width and 10x snout-gular length. Instances in which only partial measurements were available, such as when part of the tail is missing, are listed in the table with a > symbol to denote that the structure is partial and therefore larger than the available measurement. Abbreviations: head width (HW), snout-gular length (SG), snout-vent length (SVL), axilla-groin length (AG), tail length (TL), hind limb length (HLL), foot length (FL), forelimb length (FLL), trunk width (TW), distance between the eyes (E-E), eye width (EW), and distance from eye to nostril (E-N). All measurements in millimeters.

Materials
Mehama Formation, Lane County, Oregon, UO 11026 Goshen: UOMNH F-5405, a nearly complete skeleton (Fig. 1); UOMNH F-36412, a carbonized imprint preserving a few highly carbonized vertebrae representing a young, but metamorphosed individual (Fig. 2.1, 2.2); UOMNH F-30648, a partial skeleton preserving the dorsal side of the skull and anterior vertebral column representing a young adult (Fig. 2.3); UOMNH F-55196, partial skeleton preserving the ventral side of the salamander, but missing the skull, atlas, and left forelimb (Fig. 3). Mehama Formation, Linn County, Oregon, UO 4356 Thomas Creek Drive: UOMNH F-59812 A-B, a nearly complete but crushed skeleton in part and counterpart (Fig. 4).
Remarks
Van Frank (Reference Van Frank1955) originally described the type specimen of Taricha oligocenica (Fig. 1) as a new genus and species, Palaeotaricha oligocenica, citing several differences he observed from extant Taricha, including: (1) a maxilla/quadrate suture or fusion; (2) a slightly different arrangement of tarsal bones; (3) extensive, expanded and sculptured spine tables on the neural arches, (4) long epipleural rib processes, (5) an unusually large, knob-like prepubic process in the pelvis; and (6) one additional trunk vertebra (14 as opposed to 13). The latter of these traits is highly variable within species (Holman, Reference Holman2006). It should be noted that Van Frank (Reference Van Frank1955) was unclear on whether he believed the maxilla and quadrate to be truly fused, or simply sutured, because the terms may have been used interchangeably. This practice is no longer recommended, since fusion should indicate a lack of a detectable suture (Irmis, Reference Irmis2007). Later, additional preparation of the type led to elimination of some differences (maxilla-quadrate suture/fusion, prepubic process morphology), therefore Tihen (Reference Tihen1974) reclassified the type, relegating Palaeotaricha to the subgeneric level within Taricha, and assigned Taricha miocenica to the Palaeotaricha subgenus.
Naylor (Reference Naylor1979), in his description of the Taricha lindoei type, saw no reason to eliminate the subgeneric separation of Palaeotaricha and modern Taricha, even noting differences (atlas shape, height of the trunk vertebrae) that Tihen (Reference Tihen1974) and later, Estes (Reference Estes1981) apparently did not take into account. Estes (Reference Estes1981) did not recognize the Palaeotaricha subgenus because he was unconvinced that there was enough evidence to differentiate any of the fossils from extant taxa at a subgeneric level. Curiously, the long epipleural rib processes are not something seen in extant Taricha, as recognized by Van Frank (Reference Van Frank1955), not discussed in Tihen (Reference Tihen1974), and reestablished by Naylor (Reference Naylor1979). These features were not discussed when the status of the Palaeotaricha was first changed by Tihen (Reference Tihen1974), then later, when the subgenus was not recognized at all (Estes, Reference Estes1981; Dubois and Raffaëlli, Reference Dubois and Raffaëlli2009). Holman (Reference Holman2006) provided only a summary of the previous work done on the type specimen (UOMNH F-5405), and used the diagnosis and classification of Estes (Reference Estes1981). Unlike Estes (Reference Estes1981), Holman (Reference Holman2006) noted the presence of elongate epipleural rib processes. The most recent taxonomy of family Salamandridae in Dubois and Raffaëlli (Reference Dubois and Raffaëlli2009) listed all fossil Taricha species as “incertae sedis” at the subgeneric level, likely intended to indicate the uncertainty of their position in the absence of a study subsequent to the original descriptions of these species (in which T. oligocenica and T. lindoei are described from one specimen each).
Taricha lindoei Naylor, Reference Naylor1979

Figure 6 UOMNH F-59813, the only specimen of T. lindoei from Gray Butte. This specimen is not only the smallest articulated individual of Taricha in the fossil record, but also preserves a soft-tissue outline for much of the specimen’s body.

Figure 7 Taricha lindoei: (1) UOMNH F-110577, (2) UOMNH F-30616, (3) UOMNH F-37883A, and (4) JODA 1230. All specimens were found associated with the early Oligocene Bridge Creek Flora in Wheeler County, Oregon.
Holotype
UALVP 13870, partial skeleton impression, Oligocene, Big Basin Member, John Day Formation, Wheeler County, Oregon.
Diagnosis
Taricha lindoei lacks sculptured spine tables on the neural spines of the vertebrae, but has a very slight expansion of the posterior neural crest. The skull of T. lindoei is elongate and narrows anteriorly; however, the posterior region of the skull remains wide and possesses minor sculpturing. Taricha lindoei also possesses a broad scapular region of the scapulocoracoid, prominent epipleural pectoral rib processes, and concave inter-prezygapophyseal margins.
Occurrence
Early Oligocene, Oregon.
Description
The vertebrae of Taricha lindoei are opisthocoelous, as in all salamandrids, with long, thin, low neural crests that do not possess expansive, sculptured spine tables, and concave inter-prezygapophyseal neural arch margins. The type specimen (UALVP 13870), UOMNH F-35553, and UOMNH F-59813 (Fig. 6) all possess 14 trunk vertebrae, as also seen in T. oligocenica (Van Frank, Reference Van Frank1955; Naylor, Reference Naylor1979); extant Taricha typically possess 13 trunk vertebrae (Van Frank, Reference Van Frank1955; Naylor, Reference Naylor1978b, Reference Naylor1979). However, the number of trunk vertebrae is variable within the genus, and therefore ineffective in differentiating species of Taricha without a larger sample size of the fossil taxa (T. oligocenica and T. lindoei) and an analysis of intraspecific variation (Naylor, Reference Naylor1978b, Reference Naylor1979). The type specimen of T. lindoei exhibits the general proportions of modern Taricha in the neural arches and zygapophyses (Naylor Reference Naylor1979), but differ from the proportions of T. oligocenica and Notophthalmus (Naylor, Reference Naylor1978b, Reference Naylor1979). Furthermore, T. lindoei has a similar vertebral length and width to Taricha miocenica described in Tihen (Reference Tihen1974), although the neural spine is much higher in the latter (Naylor, Reference Naylor1979; Estes, Reference Estes1981). The atlas of T. lindoei is robust and slopes anteriorly, and the scapular part of scapulocoracoid is broad, both characters as in extant Taricha and unlike in T. oligocenica (Naylor, Reference Naylor1979; Holman, Reference Holman2006). The epipleural processes of the pectoral ribs are longer than those of extant Taricha species, as best seen in UOMNH F-59813 (Fig. 6; Naylor, Reference Naylor1979; Estes, Reference Estes1981; Holman, Reference Holman2006). As in all members of the genus Taricha, the skull of T. lindoei is broad posteriorly; however, the snout of T. lindoei narrows anteriorly, and the frontosquamosal arch is bony and complete. The premaxilla is fused (Figs. 6, 7). Dermal sculpturing of the skull and frontosquamosal arch is greater than in extant Taricha, but less than in T. oligocenica and Notophthalmus; the overall amount of sculpturing is minor. The preservation of the vomerine tooth row reveals a V-shape, similar to the shape exhibited in T. granulosa.
Materials
UO 2783 Gray Butte, John Day Formation, Crook County, Oregon: UOMNH F-59813, an imprint/compression of a near complete skeleton and body impression of a young adult, except the distal limbs and girdles (Fig. 6). UO 10744 Fossil locality, John Day Formation, Wheeler County, Oregon: UOMNH F-35553, an impression of a nearly complete skeleton, except for the distal parts of limbs, posterior portion of the tail, most of the ribs, and the pectoral and pelvic girdles; UOMNH F-30616, a partial skeleton impression lacking girdles, distal ends of limbs, and posterior end of tail (Fig. 7.2); UOMNH F-109709, a partial skeleton imprint; UOMNH F-109710, part and counterpart of a flattened partial skeleton; UOMNH F-110577, impression of partial skeleton including skull and several vertebrae (Fig. 7.1); UOMNH F-111395 A–B, flattened partial skeleton in part and counterpart; UOMNH F-37883 A–B, part and counterpart of a flattened skeleton, and additional flattened partial skull, and a flattened vertebra (Fig. 7.4); JODA 10429 A–B, flattened skull and first several vertebrae compressed onto block; JODA 1230, a block with skeletal impressions (Fig. 7.3). UCMP John Day Formation locality V5636, Fossil, Wheeler County, Oregon: UCMP 137466, an anterior salamander impression/compression; UCMP 137464, partial skeleton compression.
Remarks
Naylor (Reference Naylor1979) first described Taricha lindoei as the earliest representative of the subgenus Taricha, which at the time contained all extant members of the genus Taricha. The new taxonomy of Dubois and Raffaëlli (Reference Dubois and Raffaëlli2009) has since placed T. rivularis as the lone member of the subgenus Twittya based on iris color, ventral coloration, egg deposition, and habitat (flowing water as opposed to standing or mildly flowing water), while maintaining the earlier caution of Estes (Reference Estes1981) by labeling all Oligocene salamanders assigned to the genus Taricha as Incertae sedis. Estes (Reference Estes1981) stated that T. lindoei may very well represent the earliest member of the subgenus Taricha, but did not find the characterization of multiple subgenera of Taricha in the fossil record to be adequate, given one described individual each for T. lindoei and T. oligocenica, and fewer than 40 individual skeletal elements in T. miocenica. Without further evidence, it was not possible for Naylor (Reference Naylor1979) or Estes (Reference Estes1981) to determine if the combination of smaller size, narrower skull, and absence of sculptured spine tables represented ontogenetic features of one taxon or a taxonomic difference; however, the existence of T. miocenica, which was similar in size to T. lindoei, but possessed less extensive expansion of the spine table than T. oligocenica, was enough for Estes to maintain them as separate species. With the additional specimens now available, we can confirm that even the largest individual assigned to this species (UOMNH F-37883 A–B) bears no dermal cap on the vertebra, as in modern Taricha, while also possessing the broad scapular region of the living group and a narrower rostrum. Additionally, none of the specimens representing this species appears to be larval or juvenile in age; they instead represent young to mature adults. The possibility of T. lindoei and T. oligocenica belonging to the same species and differing in only age at death and preservation is therefore unlikely.
The long epipleural rib processes resemble those seen in T. oligocenica, best seen in (Naylor, Reference Naylor1979). Interestingly, the shape of the vomerine tooth row revealed in T. lindoei bears a V-shape similar to that seen in extant Taricha granulosa. Taricha lindoei consistently presents an anterior narrowing the skull not seen in other members of the genus.
It is possible that the body size of T. lindoei is smaller than T. oligocenica, given that even the most mature specimens do not reach the same size as mature T. oligocenica (Table 2). Notably, the vertebrae of fully adult T. miocenica are also smaller than those of T. oligocenica, and are around the same size as adult T. lindoei (Estes, Reference Estes1981). The fact that salamandrids are known to be variable in size (Estes, Reference Estes1981; Duellman and Trueb, Reference Duellman and Trueb1986; Holman, Reference Holman2006), combined with the small sample sizes of T. lindoei and T. oligocenica, does not currently permit accurate assessment of these differences, and therefore cannot be used as a diagnostic tool for species identification. Nonetheless, it is worth noting that, while salamanders possess the potential for indeterminate growth, in reality, body size is limited by a number of factors, especially age of first reproduction (Bruce, Reference Bruce2010) and climatic conditions such as temperature and precipitation (Reading, Reference Reading2007; Caruso et al., Reference Caruso, Sears, Adams and Lips2014). Hence, the difference in stresses between the cooler, drier, seasonal environment preserved in the Big Basin Member versus the wetter, milder environment preserved in the Mehama Formation (due to proximity to the Pacific Coast) may also explain the apparent segregation of species and the difference in body sizes for two fossil newts of similar age.
Phylogenetic analyses
Results
Because our focus is on the North American newts, specifically the extant genus Taricha and the fossil species attributed to that genus, most of our discussion of these results will focus specifically on the North American taxa. Marjanović and Witzmann (Reference Marjanović and Witzmann2015) produced a detailed and authoritative discussion of the taxa outside of North America, and with few specimens availabile, we have little to add to their discussion of those taxa beyond a few additional details and an adjustment to character 71 for Tylototriton, in which the length of the trunk ribs differs between species.
The unconstrained analysis with all 40 taxa produced 420 most parsimonious trees, with a length of 635 steps, a consistency index of 0.501, a retention index of 0.633, and a homoplasy index of 0.753. The strict consensus (Fig. 8) is not exceptionally well resolved, although the outgroups (Ambystoma and Dicamptodon), Salamandrinae (Salamandra and Chioglossa), Pleurodelini (Tylototriton, Echinotrition, and Pleurodeles), and Molgini (including a European and North American clade) form monophyletic groups as in molecular studies (Zhang et al., Reference Zhang, Papenfuss, Wake, Qu and Wake2008; Pyron, Reference Pyron2014). Unfortunately, the Molgini, along with Salamandrina, form a massive polytomy in the strict consensus tree as presently coded. This polytomy includes uncertain placements of Notophthalmus viridescens and the fossil Notophthalmus crassus, as well as several European fossil taxa, Cynops, and Paramesotriton. We also retain a stepped relationship of Lissotriton species (along with Carpathotriton) and a clade including Neurergus, Ommatotriton, Triturus, and Ichthyosaura. Finally, we recovered a clade including all living and extinct species of Taricha, along with fossil species Notophthalmus robustus in our analysis.

Figure 8 Strict consensus tree of 420 most parsimonious trees (length = 635 steps) from our unconstrained analysis with all 40 taxa. Extinct taxa are in bold.
The constrained analysis of all 40 taxa produced 760 most parsimonious trees, with a length of 667 steps, a consistency index of 0.4768, a retention index of 0.5956, and a homoplasy index of 0.7646. The strict consensus of these trees (Fig. 9) forms a large polytomy within the Salamandrinae. Within this polytomy, the strict consensus retains the entirety of the North American clade, but retains only minor parts of the Pleurodelini and European newts. Within the North American clade, the fossil species (except for N. crassus) form a polytomy alongside the group of extant Taricha species, while living N. viridescens and extinct N. crassus form a polytomy with the larger North American newt group.

Figure 9 Strict consensus tree of 760 most parsimonious trees (length = 667 steps) from our molecularly constrained analysis of all 40 taxa. Extinct taxa are in bold.
Our unconstrained analysis without the “vertebrae-only” fossil taxa produced 32 most parsimonious trees, with a length of 630 steps, a consistency index of 0.503, a retention index of 0.629, and a homoplasy index of 0.751. The strict consensus tree of this analysis (Fig. 10) retains the outgroup, the Salamandrinae, and the Pleurodelini-Molgini sister groups. Within the Molgini, we have a trichotomy of extinct and extant Taricha, Notophthalmus viridescens, and all other newts within Molgini, including the fossil taxa. Within Taricha, the fossil taxa T. oligocenica and T. lindoei form a separate branch from extant species of Taricha, which have additional soft tissue and reproductive character data available.

Figure 10 Strict consensus tree of 32 most parsimonious trees (length=630 steps) from our unconstrained analysis without the “vertebrae-only” fossil taxa. Extinct taxa are in bold.
Finally, our constrained analysis without the “vertebrae-only” fossil taxa produced 16 most parsimonious trees, with a tree length of 662 steps, a consistency index of 0.4789, a retention index of 0.5907, and a homoplasy index of 0.7628. The strict consensus tree (Fig. 11) portrays the outgroup, Salamandrina, the Salamandrinae, and a North American clade of Taricha and Notophthalmus well, but overall reduces the newts into a large polytomy, removes Pleurodeles and several fossil taxa to one step outside of the Pleurodelini, and struggles to separate the taxa into a distinct Pleurodelini-Molgini sister-group relationship. Notophthalmus viridescens forms a sister group to living and fossil species of Taricha, while the non-North American Molgini also form a single large clade with fair similarity to molecular trees.
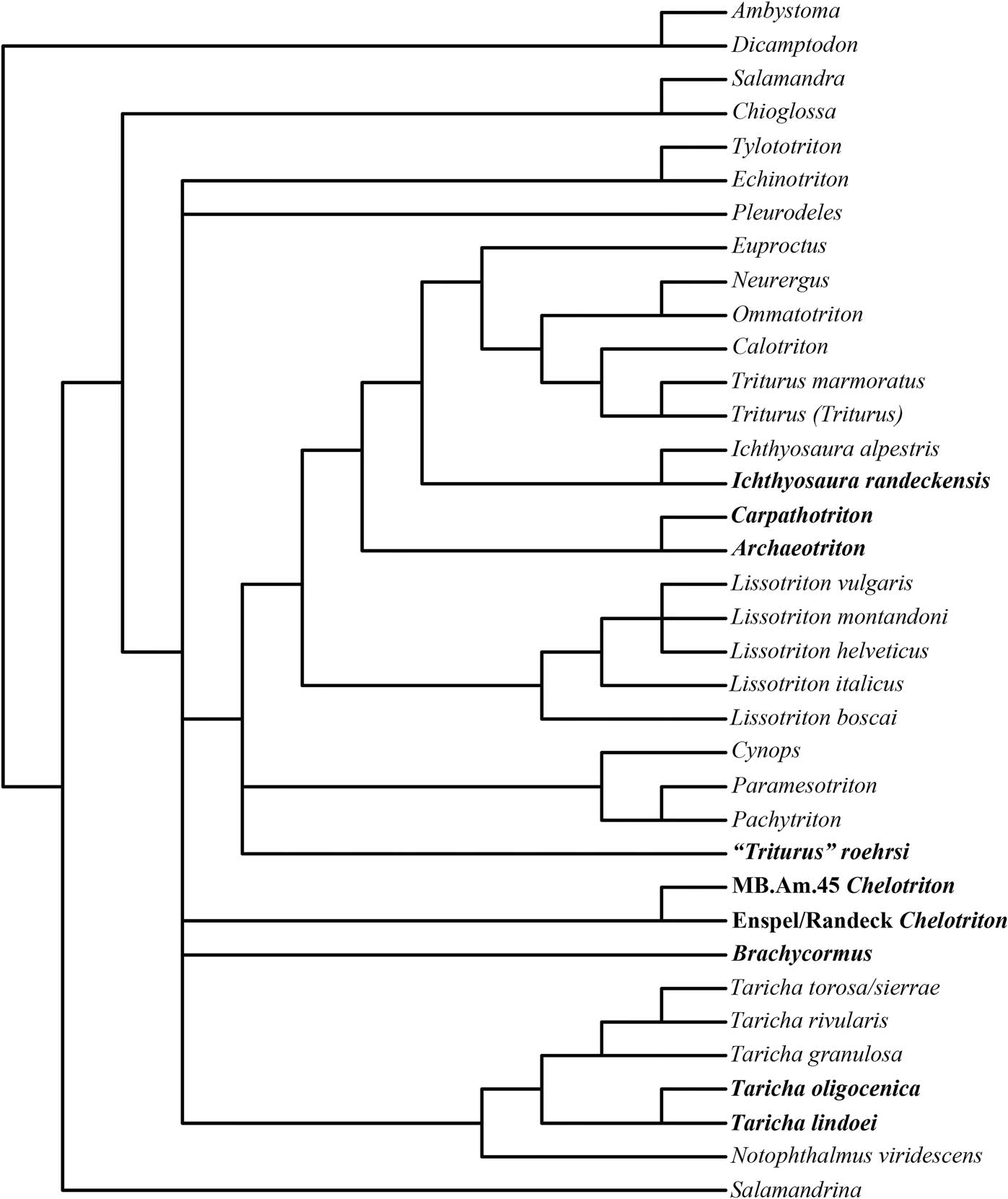
Figure 11 Strict consensus tree of 16 most parsimonious trees (length = 662 steps) from our molecularly constrained analysis without the “vertebrae-only” fossil taxa. Extinct taxa are in bold.
Discussion
This paper focuses on North American taxa, although our analysis enables us to make a few observations about non-North American taxa in addition to those found in Marjanović and Witzmann (Reference Marjanović and Witzmann2015). Both of our constrained analyses (Figs. 9, 11) form a large polytomy including the newt taxa while not recovering a monophyletic Pleurodelini, fossil Chelotriton (both the Enspel/Randeck and MB.Am.45 specimens), and Pleurodeles. The analysis reflects a North American clade including all living and fossil North American species. Removal of the “vertebrae-only” taxa before analysis greatly reduced the number of trees (from 760 to 16), suggesting that the current status of vertebral characters in Salamandridae as a whole is insufficient to properly understand several of the taxa, perhaps because of a lack of data and/or low samples sizes, homoplasy in vertebral characters, poor description of character states, or some combination of the above. One possible example of this issue pertains to the width and dorsal sculpture of the neural crest of the vertebrae (character 61; character 61 in Marjanović and Witzmann, Reference Marjanović and Witzmann2015; characters 2 and 3 in Buckley and Sanchiz, Reference Buckley and Sanchiz2012). Using Taricha as an example, Venczel (Reference Venczel2008) assigned state 0 (neural crest not expanded into spine table), whereas Buckley and Sanchiz (Reference Buckley and Sanchiz2012) and Marjanović and Witzmann (Reference Marjanović and Witzmann2015) assigned Taricha state 2 (dorsal surface of broadened spine table sculptured). Examination of extant species of Taricha (Table 1) reveals a mix of character states even within a single vertebral column. Fully mature individual skeletons typically exhibit both state 1 (unsculptured expansion of the neuropophysis into spine table) and the previously mentioned state 2, apparently across all living species in our analysis. We have coded the extant species as such. Taricha oligocenica and Taricha miocenica both appear to exhibit only state 2, though with different shapes for the spine table, while Taricha lindoei appears to exhibit state 1 as seen in some extant Taricha, although this may be an artifact of nearly two-dimensional preservation.
In addition to the difference in observations even within a single newt genus for this character, we suspect that the current list of character states is insufficient for describing the morphology of the neural crest and the spine table, when present. Taxa such as Taricha, for example, only show a broadening spine table at the posterior-most part of the neural crest (like an elongate, narrow, upside-down funnel), and may or may not exhibit sculpture, while taxa such as Echinotriton and Tylototriton have overall broader, near-triangular, extensive neural crest spine tables, and appear to exhibit sculpture throughout a mature individual’s trunk vertebrae. We suggest investigating differences of shape in the spine table in the future, and perhaps either adding these data as an additional character, or using them to better define or increase the number of character states for this character in future analyses. Other vertebral characters may similarly produce such issues, but we feel that the amount of expansion and sculpture of the neural crest exhibits the problem best.
Our unconstrained analyses (Figs. 8, 10) recovered a monophyletic Pleurodelini group, unlike in our constrained analyses; however, inclusion of all 40 taxa resulted in a massive polytomy between the Pleurodelini and various parts of what is molecularly recognized as the Molgini. Just as in the constrained analyses, the removal of “vertebrae-only” taxa greatly reduced the number of most parsimonious trees (from 420 to 32), and provided better resolution for the Molgini, separating out the non-North American Molgini, Taricha, and Notophthalmus. One major issue with the unconstrained analyses is that Notophthalmus viridescens and, when included, Notophthalmus crassus do not form a group with Notophthalmus robustus, nor do they form a sister-group relationship with Taricha such that a North American Molgini clade is formed. Instead, N. viridescens and N. crassus form a polytomy with all other North American newts when all taxa are included, while N. viridescens forms a trichotomy with Taricha and the non-North American Molgini. We suggest that our unconstrained morphological analyses fail to show a consensus North American clade as the result of a lack of characters for the post-cranial skeleton of Notophthalmus, combined with a deep split of the Taricha and Notophthalmus lineages, resulting in significant autapomorphy evolution that likely began in either the late Oligocene or early Miocene.
In all four of our strict consensus trees (Figs. 8–11), we recover a group including all extant and fossil species of Taricha, the vertebrae-only Taricha miocenica, as well as, curiously, Notophthalmus robustus, when they are included. Our strict consensus trees also did not place any fossil or extant North American newts within the Eurasian Molgini. Estes (Reference Estes1981) noted that N. robustus is apparently less derived than other known species, and differs more from at least the extant species than those species do from each other. While T. miocenica was placed with other species of Taricha, its exact placement within the group is uncertain, possibly because it relies entirely on our vertebral characters; analyses including N. robustus and N. crassus face a similar problem. As such, we suggest a reexamination of the three “vertebrae-only” North American taxa because we did not investigate those taxa personally when assigning character states. We furthermore suggest the addition of extant taxa Notophthalmus meridionalis and Notophthalmus perstriatus to future analyses that look to investigate North American taxa specifically, especially when including fossil taxa. Clearly, the current state of unconstrained morphological analyses struggle with Notophthalmus overall; the addition of all species and species-level characters may improve the resolution in future studies.
We also suggest deeper investigation into the vertebral characters of the Salamandridae overall because the current state of characters may not necessarily explain the morphological similarities and differences within the group. We were unable to address this in our current study, and were also unable to code some of our original characters for non-North American taxa, because of insufficient specimen availability to account for possible variation. Future studies should build on our current analysis and incorporate more data, as available, while studying multiple individuals of each taxon.
Overall, we find support for T. oligocenica, T. lindoei, and T. miocenica as members of the Taricha group with a long history of >32 Ma. The curious and consistent placement of N. robustus warrants a reexamination of the taxon and the character states assigned to it, and we recommend the same for the remaining North American taxa (including the questionable taxon N. slaughteri) for the sake of confirming their morphologies. Improving the characters and character states for the vertebrae and ribs of salamandrid phylogenies should also be a priority for future analyses; other characters, such as the single geographic character, may also be possible to improve on when advancing these studies.
Functional morphology
Discussion
The overall impression of Taricha oligocenica is that of a sturdy or robust, moderate-to-large-sized newt with a mosaic of defensive adaptions present in the Salamandridae. First is the frontosquamosal arch, which consists of an anteromedial projection from the squamosal and a posterolateral projection from the frontal that has been considered a defensive adaptation in the family Salamandridae (Naylor, Reference Naylor1978a; Brodie, Reference Brodie1983; Duellman and Trueb, Reference Duellman and Trueb1986), the function of which is to lessen injury during predatory attack by increasing the skull’s resistance to crushing. The frontosquamosal arch also increases the difficulty of swallowing for predators similar to Thamnophis, which attempt to swallow their prey headfirst (Brodie, Reference Brodie1968a, Reference Brodie1983; Naylor, Reference Naylor1978a; Duellman and Trueb, Reference Duellman and Trueb1986), or predators such as Lithobates and some fishes, which also attempt to swallow their prey whole (Brodie, Reference Brodie1968a; Naylor Reference Naylor1978a). Although not all salamandrids possess a complete, bony arch (some exhibit a narrow gap, or no frontosquamosal arch), this structure is only found in the Salamandridae (Wake and Özeti, Reference Wake and Özeti1969; Naylor, Reference Naylor1978a; Duellman and Trueb, Reference Duellman and Trueb1986). The ridges and grooves of the frontosquamosal arch, continuous with the relatively robust and rigid skull seen in salamandrids (compared to other salamanders), effectively strengthen the skull, especially when resisting lateral pressure, such as that experienced when being swallowed by predators, including bullfrogs and snakes (Naylor, Reference Naylor1978a). Additionally, the paired arches can form a protective shelf over the eyes of newts; this shelf can also be observed when these newts swallow or take a defensive posture (Brodie, Reference Brodie1977). As mentioned above, the extensive pitting of the skull and frontosquamosal arches of T. oligocenica is beyond that of living members of Taricha, but less than found in Echinotriton or Tylototriton.
The most glaring differences between T. oligocenica and extant Taricha involve the heavy ossification of different parts of the skeleton in T. oligocenica, specifically the trunk region. As Naylor (Reference Naylor1978a) has pointed out, only newts with complete bony frontosquamosal arches possess expanded spine tables on the vertebral neural crests (examples include Notophthalmus, Cynops, and Tylototriton; Brodie, Reference Brodie1983; Duellman and Trueb, Reference Duellman and Trueb1986). Although not all newts with bony arches possess extensive sculpture on these expansions, others may vary in degree of sculpture even within an individual, such as in Taricha (Naylor, Reference Naylor1978a, Reference Naylor1978b; Brodie, Reference Brodie1983; this study). These expansions are most likely defensive in nature (Naylor, Reference Naylor1978a; Brodie, Reference Brodie1983; Duellman and Trueb, Reference Duellman and Trueb1986) because they have no muscle attachments and form interlocking series along the usually vulnerable spine of these salamanders. The amount of protection provided by these expansions likely differs by how extensively they cover the spine (Van Frank, Reference Van Frank1955). Taricha oligocenica stands out in that its expansions are as extensive and sculptured as those of extant Tylototriton and Echinotriton, perhaps even more so, as the spine tables of those taxa narrow anteriorly, while T. oligocenica has somewhat rectangular expansions (Van Frank, Reference Van Frank1955, Estes, Reference Estes1981; this study).
The elongate epipleural rib processes of T. oligocenica and T. lindoei are here proposed to provide a defensive role similar to the epipleural processes of Tylototriton. Like Tylototriton, T. oligocenica and T. lindoei possess singular, robust processes, directed dorsally and distally, on somewhat elongate (beyond the length of a simple rod) ribs, both of which terminate in blunt ends (Nussbaum and Brodie, Reference Nussbaum and Brodie1982; Brodie et al., Reference Brodie, Nussbaum and DiGiovanni1984; Heiss et al., Reference Heiss, Natchev, Salaberger, Gumpenberger, Rabanser and Weisgram2009). Unlike in Echinotriton, Pleurodeles, and possibly fossil salamanders such as Chelotriton (Marjanović and Witzmann, Reference Marjanović and Witzmann2015; Schoch et al., Reference Schoch, Poschmann and Kupfer2015), which possess even longer, pointed ribs, often with one or more pointed epipleural processes that function to pierce the skin and inject toxins into predators if compressed, these structures are unlikely to have protruded through the skin, and may have been used for muscle attachment and to apply pressure to toxin-containing granular gland clusters (risen on the skin of Tylototriton) when compressed without piercing the skin or to help project toxin-concentrated parts of the body towards potential antagonists. The increased muscle attachment and overall robustness of the structures could also have provided extra resistance against crushing forces when attacked. Unfortunately, testing this latter hypothesis would involve significant biomechanical testing, which is beyond the scope of this study. It should also be noted that epipleural processes are present throughout fossil and extant salamandrids (and even other salamanders) to some extent, with various types and levels of functionality (Nussbaum and Brodie, Reference Nussbaum and Brodie1982; Brodie et al., Reference Brodie, Nussbaum and DiGiovanni1984; Marjanović and Witzmann, Reference Marjanović and Witzmann2015). It is therefore difficult to project the function of epipleural processes in fossil Taricha with complete confidence outside of their similarity to those of extant Tylototriton verrucosus.
The skin of all newts is toxic and sometimes fatal to potential predators; it is especially potent in the genus Taricha (Wakely et al., Reference Wakely, Fuhrman, Fuhrman, Fischer and Mosher1966; Brodie, Reference Brodie1968a, Reference Brodie1968b; Brodie et al., Reference Brodie, Hensel and Johnson1974; Hanifin, et al., Reference Hanifin, Brodie and Brodie2004). Given the phylogenetic bracketing of T. oligocenica and T. lindoei with other toxic newt species (including both North American genera), the fossil species were also likely to have been toxic, although it is impossible to discern how potent the toxins actually were in these organisms. Naylor (Reference Naylor1978a) was the first to propose a connection between the frontosquamosal arch and newt toxins on the basis that it is decidedly advantageous if an organism with toxic (not to mention distasteful) skin secretions also has defensive mechanisms that allow it to survive and decrease injury from predators experiencing the poison. In a now-famous encounter, one bullfrog (Lithobates catesbeiana) voluntarily ate an individual of T. granulosa (Brodie, Reference Brodie1968a). The defensive advantages of a robust, hardy body and toxic skin secretions became clear when the frog died within ten minutes and the newt emerged unscathed shortly after (Brodie, Reference Brodie1968a). While the toxicity of the fossil newts in this study is unknown, the skeleton of T. oligocenica in particular is more armored than that of T. granulosa (Van Frank, Reference Van Frank1955), and is likely able to resist easy consumption from predators, as seen in extant Taricha (Brodie, Reference Brodie1968a, Reference Brodie1977; Naylor, Reference Naylor1979).
Conclusions
Our four phylogenetic analyses provided mixed results when compared to the molecular phylogenies of Zhang et al. (Reference Zhang, Papenfuss, Wake, Qu and Wake2008) and Pyron (Reference Pyron2014), yet consistently nested Taricha oligocenica, Taricha lindoei, Taricha miocenica, and Notophthalmus robustus within a clade of North American newts. Future studies should attempt to find additional osteological characters for all fossil and extant species of North American newts for better resolution of morphological analyses, particularly for Notophthalmus, which appears to be problematic in morphological analyses.
Unfortunately, the morphological characters in this analysis did not resolve the positions of the fossil taxa relative to extant species of the genus, so their exact classification within the North American clade is uncertain. The use of muscular and behavioral characters does not help resolve fossil species, while a lack of species-level osteological characters, the presence of homoplasy in some potential characters, and the possibility of cryptic species complicate the ability of our analyses to discriminate between taxa.
A detailed morphological study of T. oligocenica and T. lindoei supports their classification as separate species more similar to Taricha than to any other known genus of newt. As in the phylogenetic analysis, this conclusion suggests that the genus Taricha has deep roots into at least the early Oligocene, where ancestors to living species may have diverged 32–33 Ma. Functionally, T. lindoei is similar to, though possibly smaller than, all extant taxa, and existed in a seasonal temperate environment similar to the environments in which Pacific newts are found today. Taricha oligocenica, on the other hand, represents a large, robust newt that was considerably more armored than extant Taricha, and lived in a milder climate nearer to the Oligocene coast.
Because we did not directly examine any specimens of Taricha miocenica or of fossil Notophthalmus outside of published literature (Tihen, Reference Tihen1974; Estes, Reference Estes1981; Holman, Reference Holman2006), it is of particular importance to examine and compare these fossils in detail with additional fossils of Taricha and Notophthalmus, along with skeletons of extant members of these genera, to examine their evolutionary relationships with modern North American salamandrids. The fact that T. miocenica and all fossils of Notophthalmus are only preserved as isolated vertebrae (Tihen, Reference Tihen1974; Estes, Reference Estes1981) enhances the difficulty of such determinations, and our analyses and observations indicate further development of vertebral characters is clearly required. Despite the complications, only by performing such studies can we hope to draw concrete conclusions on the phylogenetic positions North American fossil newts relative to each other and to extant Taricha and Notophthalmus.
Acknowledgments
We thank E. Davis (UOMNCH), G. Retallack (UOMNCH), J. Samuels (East Tennessee State University), P. Holroyd (UCMP), K. Padian (UCMP), and D. Wake (UCMVZ) for access to the specimens under their care. We are grateful to E. Davis (UOMNCH), G. Retallack (UOMNCH), J. Samuels (ETSU), and the past and present members of the Hopkins-Davis Lab and other students from the University of Oregon for productive discourse and support. Special thanks to E. White (UOMNCH), for contributing line drawings for Figures 1 and 5, and N. Famoso (JODA), for photographs in Figures 1, 2.1, 7.2, and 7.3. We give credit to the Museum of Comparative Zoology and Harvard University for permission to use copyrighted material to help make the line drawings in Figure 5. We also thank the editor and reviewers of this paper for their contributions to the quality of this work, particularly D. Marjanović for his insight on non-North American newts. Finally, we acknowledge funding from a Geological Society of America graduate student research grant, the University of Oregon Thomas Condon Fellowship, and the UCMP Doris O. and Samuel P. Welles Research Fund for making this research possible.
Accessibility of supplemental data
Data available from the Dryad Digital Repository: http://dx.doi.org/doi:10.5061/dryad.8m9t1













