


1. Introduction
In the traditional mid Cambrian (or provisional Cambrian Epoch 3) marine platforms of southwestern Europe, for which palaeoenvironments and palaeogeography have been reconstructed by means of sedimentological (Álvaro & Vennin, Reference Álvaro and Vennin1997) and chemo- (Wotte et al. Reference Wotte, Álvaro, Shields, Brasier and Veizer2007; Álvaro et al. Reference Álvaro, Bauluz, Pierre, Subías and Vizcaïno2008a), event (Álvaro et al. Reference Álvaro, Ferretti, González-Gómez, Serpagli, Tortello, Vecoli and Vizcaïno2007) and sequence stratigraphical analyses (Álvaro, González-Gómez & Vizcaïno, Reference Álvaro, González-Gómez and Vizcaïno2003), generally clayey strata deposited in offshore settings display the most abundant and diverse fossil record. A well-documented biodiversity peak is represented by the Caesaraugustan–lowest Languedocian offshore shales of the Murero Formation (Iberian Chains; Álvaro, Vizcaïno & Vennin, Reference Álvaro, Vizcaïno and Vennin1999; Zamora & Álvaro, Reference Zamora and Álvaro2010), the Genestosa Member (Cantabrian Zone; Zamora, Reference Zamora2010) and the Coulouma Formation (Montagne Noire; Álvaro et al. Reference Álvaro, González-Gómez and Vizcaïno2003), where more than 30 trilobite, brachiopod and echinoderm species have been reported. Similarity of species composition is remarkable among coeval offshore-dominated benthic faunas along the Mediterranean margin of West Gondwana. Detailed descriptions of the fossil associations, recognized on the basis of autochthonous and parautochthonous modes of occurrence, were given in previous papers (Álvaro & Vennin, Reference Álvaro and Vennin1997; Álvaro, Vizcaïno & Vennin, Reference Álvaro, Vizcaïno and Vennin1999; Álvaro et al. Reference Álvaro, Elicki, Geyer, Rushton and Shergold2003; González-Gómez, Reference González-Gómez2005).
The aforementioned acme in benthic diversity was followed by a gradual decline and a barren interval associated with the onset of the mid Languedocian regression, well constrained throughout the western Mediterranean region: e.g. in the Borobia and Valdeorea formations of the Iberian Chains, the Adrados Member of the Cantabrian Zone and the Ferrals Formation of the Montagne Noire. In the mixed (siliciclastic-carbonate) platforms that recorded the re-establishment of mid-latitude shelly carbonate factories, such as those recognized in the Montagne Noire (Val d'Homs Formation), the aftermath of the regression was characterized in the late Languedocian by a major faunal turnover of trilobite, brachiopod and echinoderm families (Álvaro et al. Reference Álvaro, González-Gómez and Vizcaïno2003; Zamora & Álvaro, Reference Zamora and Álvaro2010). This is followed by a renewed Furongian–early Tremadocian radiation related to the stepwise immigration of invaders from East Gondwana, under persistent transgressive conditions (Courtessole, Pillet & Vizcaïno, Reference Courtessole, Pillet and Vizcaïno1988; Shergold, Feist & Vizcaïno, Reference Shergold, Feist and Vizcaïno2000; Álvaro et al. Reference Álvaro, Lefebvre, Shergold and Vizcaïno2001, Reference Álvaro, Elicki, Geyer, Rushton and Shergold2003, Reference Álvaro, Ferretti, González-Gómez, Serpagli, Tortello, Vecoli and Vizcaïno2007; Vizcaïno & Álvaro, Reference Vizcaïno and Álvaro2003; González-Gómez, Reference González-Gómez2005).
The Caesaraugustan–earliest Languedocian marine benthic assemblages of southwestern Europe represent ecologically analogous associations occurring in distinct biostratigraphical levels, where similar environmental conditions prevailed; however, this was not the case for the upper Languedocian transgressive shales. Correlation of faunal gradients with the discussed transgressive–regressive cycles in response to shifting environmental conditions, was, until recently, exclusively reported from the limestone interbeds of the Languedocian–Furongian transition (e.g. Val d'Homs Formation, Montagne Noire), whereas the fossil record in laterally equivalent shale strata (such as the Valtorres Formation, Iberian Chains) was stated to be dramatically impoverished (Schmitz, Reference Schmitz1971; Josopait, Reference Josopait1972; Shergold & Sdzuy, Reference Shergold and Sdzuy1991; Álvaro et al. Reference Álvaro, Ferretti, González-Gómez, Serpagli, Tortello, Vecoli and Vizcaïno2007).
The lack of late Languedocian faunas in clayey substrates has been explained (Álvaro et al. Reference Álvaro, González-Gómez and Vizcaïno2003, Reference Álvaro, Ferretti, González-Gómez, Serpagli, Tortello, Vecoli and Vizcaïno2007) by (i) the absence of carbonate substrates and their colonization by stenotopic benthic assemblages, (ii) taphonomic constraints recorded exclusively in the shale counterparts, or (iii) sampling biases. In order to test these possibilities, extensive sampling of the palaeogeographically distal parts of the upper Languedocian strata in the Iberian Chains, where the destructive effects of taphonomic agents were less marked, was undertaken. This resulted in the finding of the first reported siliceous concretions embedded in the Valtorres Formation. These are rife with trilobites, hyoliths, brachiopods and sponge spicules, and provide a taphonomic window containing vital information for interpreting the replacement of temperate-water, benthic communities in the aftermath of the mid Languedocian regression. This paper offers a re-assessment of the late Languedocian immigration of benthic communities both in mixed and siliciclastic substrates of southwestern Europe in the light of the fact that preservation and sampling biases dramatically control the knowledge of early Palaeozoic ecostratigraphical patterns.
2. Geological setting and stratigraphy
The pre-Variscan exposures of the Iberian Peninsula are traditionally subdivided into tectonostratigraphical zones, such as the Cantabrian, West Asturian-Leonese, Central-Iberian and Ossa-Morena zones (Fig. 1a). Their early Palaeozoic geodynamic evolution has been interpreted in terms of two distinct troughs: the Ossa-Morena and Cantabro-Iberian basins, the latter comprising some disconnected inliers, such as the Sierra de la Demanda and the Iberian Chains. The Cantabro-Iberian Basin was limited to the northeast by the Cantabro-Ebroan Land area, to the north of which a mosaic of Cambrian platforms are preserved in the Pyrenees, the Montagne Noire (southern Central Massif, France) and Sardinia (Italy).
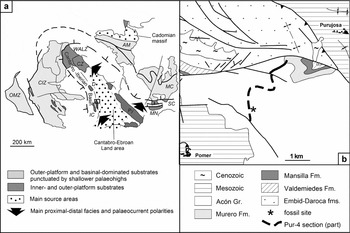
Figure 1. (a) Tentative Cambrian palaeogeographic reconstruction of the inner-platform/basin transects through the SW European margin of West Gondwana; modified from Álvaro et al. (Reference Álvaro, Monceret, Monceret, Verraes and Vizcaïno2010). OMZ – Ossa-Morena Zone; CIZ – Central-Iberian Zone; IC – Iberian Chains; D – Sierra de la Demanda; CZ – Cantabrian Zone; WALZ – West-Astrurian Leonese Zone; AM – Armorican massif; MC – Massif Central; SC – Southern Cévennes; MN – Montagne Noire; Py – Pyrenees. (b) Boxed area shown in (a) illustrating the study section in Purujosa, eastern Iberian Chain.
In the Cantabro-Iberian Basin, the Languedocian regional stage of the traditional middle Cambrian was formally erected by Álvaro & Vizcaïno (Reference Álvaro and Vizcaïno1998) to replace the informal intervals ‘Solenopleuropsidae-free’ previously proposed for the Iberian Peninsula (Sdzuy, Reference Sdzuy1971) and the ‘Barroubian’ (de Rouville, Delage & Miquel, Reference Rouville, Delage and Miquel1894; Miquel, Reference Miquel1905) for the Montagne Noire. In the latter area, the stratotype of the Languedocian was selected at Ferrals-les-Montagnes (Pardailhan nappe), whereas another section at Sallèles-Cabardès (Minervois nappe) was proposed as a parastratotype for its top. In both sections, a continuous succession spanning the Coulouma, Ferrals, La Gardie and Val d'Homs formations has yielded 36 trilobite species (Courtessole, Reference Courtessole1973, tables 3, 4; Courtessole, Pillet & Vizcaïno, Reference Courtessole, Pillet and Vizcaïno1988; Álvaro, Vizcaïno & Vennin, Reference Álvaro, Vizcaïno and Vennin1999). The base of the Languedocian was defined at the first appearance datum (FAD) of Manublesia thorali and its top at the FAD of the genus Bergeronites sensu Feist & Courtessole (Reference Feist and Courtessole1984), subsequently constrained to the species Palaeadotes latefalcata (Álvaro et al. Reference Álvaro, Vizcaïno, Kordule, Fatka and Pillola2004, Reference Álvaro2007). The top of the Languedocian does not coincide with the base of the Furongian, which has been recognized chemostratigraphically in the Montagne Noire, marked by the SPICE (Steptoean Positive Carbon Isotope Excursion) event (Álvaro et al. Reference Álvaro, Ezzouhairi, Ribeiro, Ramos and Solá2008a). The Languedocian Stage was subdivided into the following substages and zones: the M. thorali Interval Zone and the Solenopleuropsis Assemblage Zone (lower Languedocian), the Jincella? cf. prantli (Jincella convexa according to Álvaro et al. Reference Álvaro, Vizcaïno, Kordule, Fatka and Pillola2004) and Bailiella souchoni interval zones (middle Languedocian), and the Eccaparadoxides (Macrocerca) macrocercus Interval Zone (upper Languedocian). Languedocian trilobite and echinoderm species of the Montagne Noire have also been found in Morocco, the Iberian Peninsula, Sardinia, the Czech Republic, Germany and Turkey, facilitating correlation between coeval strata throughout the Mediterranean (biogeographic) subprovince (Álvaro et al. Reference Álvaro, González-Gómez and Vizcaïno2003; Geyer & Landing, Reference Geyer and Landing2004; Zamora & Álvaro, Reference Zamora and Álvaro2010).
Owing to preservation constraints, late Languedocian trilobites were not clearly recognized in the Iberian Chains, and several species similar to those described from the Montagne Noire were reported in open nomenclature (Schmitz, Reference Schmitz1971; Josopait, Reference Josopait1972). In the Iberian Chains, a single occurrence of trilobites in the upper part of the Valtorres Formation has so far provided the only evidence for the immigration of benthic faunas after the mid Languedocian regression; these trilobites are represented by endemic species of typical Furongian genera (Shergold & Sdzuy, Reference Shergold and Sdzuy1991). As a result, the barren lower part of the formation could not be tied to the global chronostratigraphical scheme. Investigation of available exposures in the vicinity of Purujosa (Fig. 1b), a locality palaeogeographically situated in the distal part of the Iberian platform (a part of the Cantabro-Ebroan Basin incompletely preserved in the Iberian Chains; Fig. 1a), has yielded a single occurrence of siliceous concretions within a c. 10 m thick interval. The concretions occur 6 m above the Encomienda/Valtorres contact (Fig. 2) and fill the gap in the present knowledge of the aftermath of the mid Languedocian regression in the Iberian Chains.
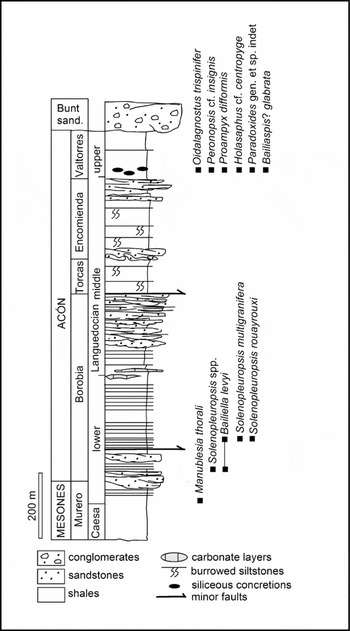
Figure 2. Languedocian (Cambrian Series 3) stratigraphical units recognized in the vicinity of Purujosa with setting of the described trilobite association; Caesa. – Caesaraugustan, Buntsand. – Buntsandstein.
3. Taphonomic features
One hundred and twenty fossiliferous concretions, from more than 300 split, were sampled in the Valtorres Formation of Purujosa. They are up to 10 cm in diameter and occur embedded with their long axes lying parallel to the bedding plane. They are uncompacted and stand out as small bosses on the bedding surface. Where affected by dolomitization, they become powdery. The contact with the surrounding shale is sharp where the silicification is more complete. The concretions contain a diverse shelly fauna, composed of trilobites, calcitic and linguliformean brachiopods, helcionellids, hyoliths, hexactinellid sponge spicules and subsidiary bradoriids (Fig. 3a, b). The host shale contains casts of flattened and poorly preserved shelly fossils and is irregularly burrowed.

Figure 3. Taphonomic features of the Guzhangian siliceous concretions of the Valtorres Formation at Purujosa. (a) Open concretion exhibiting internal and external moulds of the fossils; scale equals 10 mm. (b) Latex cast of hyolith accumulation; scale equals 5 mm. (c) Thin-section photomicrograph of trilobite sclerites showing iron-oxide (Fe) cores encrusted with siliceous crusts (Si) and embedded in a partly silicified matrix (m) rich in oxyhydroxides; scale equals 300 μm. (d) Disarticulated hexactinellid spicules replaced by iron oxyhydroxides; scale equals 2 mm.
The concretions consist of a mixture of terrigenous material (quartz and phyllosilicates such as illite/muscovite and chlorite) and biogenic components (calcite- and apatite-walled skeletons). Crystalline micro- and megaquartz (<20 μm and >20 μm, respectively) are the most abundant silica types (Fig. 3c). They occur both as replacement and as pore-lining and pore-filling cements in interparticle and mouldic porosities. Crystals are euhedral and equant, 10 to 150 μm in size, and display a mosaic texture similar to drusy mosaics. Silica cement shows an irregular poikilotopic texture due to pore-occluding cements. Where present, megaquartz represents the last pore-filling silica phase. The original high porosity was due to the morphology and arrangement of skeletal material.
Dolomite occurs as finely crystalline to microsparite, subhedral to anhedral crystals. The presence of clear dolomite rhombs, 20–30 μm in size, within the iron-oxide stained chert suggests that some dolomitization occurred before silicification. Pyrite is found as dispersed euhedral crystals, mainly pyritohedra and cubes, ranging from 15 to 50 μm in size, and represents less than 5% in volume of the components. Goethite and haematite also appear occasionally (particularly in the incompletely silicified concretions) as botryoidal networks or crusts, up to 4 mm thick and subsequently lined with silica.
Shelly accumulations are concentrated within the concretions (in the centre, at the margin or wholly dispersed). The shells are dominantly disarticulated and fragmented, although a few complete and enrolled trilobites have also been encountered. The shells and skeletons in the concretions do not show any deformation, contrary to those sparsely preserved in the host rock. The accumulations in the concretions do not develop laterally correlatable shell pavements, and represent relics of skeletal reworking by storm waves and the winnowing of fine particles.
Hexactinellid sponge spicules, originally of silica, are ubiquitous and have been completely replaced by ferroan dolomite and goethite (for a description of similar processes see Warnke, Reference Warnke1995 and Loi & Dabard, Reference Loi and Dabard2002). The spicules, mainly monaxons and triaxons bearing a central channel, and 0.1 to 4 mm long, are disarticulated (Fig. 3d). It is assumed that the silica building up the tiny authigenic quartz of the concretions was derived from dissolution of these spicules. The amount delivered by solution of detrital quartz grains in siltstone (tempestite) interbeds cannot be significant, as the grains are only slightly corroded, and volcaniclastic beds are absent (Álvaro et al. Reference Álvaro, Ezzouhairi, Ribeiro, Ramos and Solá2008b). The possibility that part of the silica was derived from other siliceous microfossils cannot be excluded, but relics of such forms are absent. The original silica was dissolved entirely, probably owing to slight changes in pH because of decaying organic matter. The solution produced silica-rich interstitial waters that reprecipitated as quartz, forming concretions that protected the hosted fossil skeletons from compaction and dissolution.
4. Systematic palaeontology
The basal part of the Valtorres Formation in Purujosa is richly fossiliferous, and the macrofauna has been systematically collected from the available concretions. All complete specimens and fragments identifiable to at least generic level were collected for preparation and further identification. More than 150 specimens have been studied from the concretions. The fossils are dominated by calcitic shells (94% of which are trilobites and hyoliths). The morphological terms used herein are mostly those defined by Whittington et al. (Reference Whittington, Chatterton, Speyer, Fortey, Owens, Chang, Dean, Jell, Laurie, Palmer, Repina, Rushton, Shergold, Clarkson, Wilmot, Kelly and Kaesler1997). Illustrated material from Purujosa is housed in the Palaeontological Museum of the Zaragoza University (abbreviation, MPZ).
Family Peronopsidae Westergård, Reference Westergård1936
Genus Peronopsis Hawle & Corda, Reference Hawle and Corda1847
Type species. Battus integer Beyrich, Reference Beyrich1845 from the Paradoxides gracilis Zone at Jince, Czech Republic, OD.
Peronopsis cf. insignis (Wallerius, Reference Wallerius1895)
Figure 4a–e
Material. Six cephala (MPZ 2012/26–31), two pygidia (MPZ 2012/32–33) and two enrolled specimens (MPZ 2012/34–35), preserved as internal and external moulds.

Figure 4. Agnostoids from the Guzhangian siliceous concretions of the Valtorres Formation at Purujosa. (a–e) Peronopsis cf. insignis (Wallerius, Reference Wallerius1895): (a, b) cephalon, internal mould, dorsal and lateral views, same scale, MPZ 2012/26; (c–e) complete enrolled specimen showing the pygidium; latex cast of external mould (c, d), dorsal and lateral views, same scale, and internal mould (e), MPZ 2012/34. (f–i) Oidalagnostus trispinifer Westergård, Reference Westergård1946: (f) cephalon, internal mould, MPZ 2012/36; (g–i) pygidia; internal mould (g, h) and latex cast of external mould (i), MPZ 2012/38–39. Scales of (a–e) equal 1 mm; scales of (f–i) equal 2 mm.
Description. Cephalon moderately convex, subcircular to subquadrate in outline, and subequal in length and width; glabella relatively convex, subrectangular, rounded anteriorly with anteroglabella narrower than M3, twice as long as its maximum width, surrounded by well-defined axial furrows, and relatively long (70% of total cephalic length) and wide (maximum glabellar width is 35% of maximum cephalic width); glabella unequally divided by transglabellar furrow (F3), straight or slightly curved forwards medially; F1 and F2 obsolete or absent; basal lobes subtriangular and simple, slightly wider than long, separated from glabella and genae by well-impressed furrows; genae smooth, convex, strongly down-sloping and confluent anteriorly (preglabellar median furrow absent); preglabellar field slightly shorter (sag.) than width (tr.) of posterior part of genae; border narrow, convex, defined by shallow border furrow and abrupt change in slope; sagittally, border and border furrow combined occupy 10% of total cephalic length; genal angle pointed, without spines; posterior part of cephalon not preserved.
Pygidium highly convex, subquadrate in outline; axis (excluding articulating half-ring) occupying 80% of total length (sag.), highly convex, unfurrowed, nearly parallel-sided to slightly tapering backwards, broadly rounded posteriorly, length of axis 1.4 times as long as its maximum width, and separated from posterior border furrow; F1 and F2 obsolete to effaced; median node or tubercle prominent, subcircular in outline; axis set off from pleural fields by well-defined dorsal furrows, effaced posteriorly; pleural fields smooth, strongly down-sloping, narrowest and confluent behind axis; border smooth, moderately wide, flat and widest posterolaterally, bearing a pair of tiny spines on posterolateral margins; border furrow marked by sharp change in slope; border and border furrow occupying 10% of total pygidial length (sag.); anterior border set off from pleural field by deeply incised furrow.
Comparison. Following Robison (Reference Robison1994), Peronopsis includes numerous species with plesiomorphic characters, such as confluent genae and a bipartite glabella. The material from the Valtorres Formation most closely resembles that of P. insignis from the Lejopyge laevigata Zone at Gudhem in Västergötland, south-central Sweden, except that the pygidial axis is wider, at least in the pygidium shown in Figure 4e; the Spanish cephala are incomplete but their preserved parts fall within the intraspecific variability shown by Westergård (Reference Westergård1946, pl. 3, figs 10–16). With the limited and poorly preserved material at hand, it is left under open nomenclature as P. cf. insignis.
Family Diplagnostidae Whitehouse, Reference Whitehouse1936
Subfamily Oidalagnostinae Öpik, Reference Öpik1967
Genus Oidalagnostus Westergård, Reference Westergård1946
Type species. Oidalagnostus trispinifer Westergård, Reference Westergård1946 from the Lejopyge laevigata Zone at Gudhem, Västergötland, Sweden, OD.
Oidalagnostus trispinifer Westergård, Reference Westergård1946
Figure 4f–i
2000 Oidalagnostus trispinifer Westergård, Reference Westergård1946; Peng & Robison, p. 56, figs 5.3, 42 (with previous synonymies).
Material. Two cephala (MPZ 2012/36–37) and three pygidia (MPZ 2012/38–40), preserved as internal and external moulds.
Description. Cephalon subquadrate in outline, and subequal in length and width; glabella slightly convex, nearly parallel-sided and rounded anteriorly, twice as long as its maximum width, surrounded by well-defined axial furrows; glabella relatively long (70% of total cephalic length) and wide (maximum glabellar width 35% of maximum cephalic width), unequally subdivided by nearly straight transglabellar furrow (F3); F1 and F2 absent; posteroglabella with narrow, elongated node in front of midpoint; basal lobes subtriangular, as wide as long, separated from glabella and genae by well-impressed furrows; genae slightly scrobiculate, gently convex; preglabellar median furrow well marked; border narrow, convex, defined by shallow and wide border furrow; border and border furrow of subequal length (sag.); sagittally, border and border furrow combined occupy 15% of total cephalic length; genal angle poorly preserved.
Pygidium subrectangular in outline, with length (sag.) 85% of pygidial width; axis long, posteriorly swollen, with weak ring furrows; posterior end of axis bluntly rounded; median node on M2 elongated, indenting F1 anteriorly and F3 posteriorly; posteroaxis divided by deep sulcus, curved slightly forwards medially; posteroaxis behind sulcus subrectangular; accessory lobules on acrolobe long, divided by a furrow, extending backwards and outwards from anterior part of posteroaxis, and terminating along a transverse line passing through rear part of posteroaxis; posterior margin trispinose.
Remarks. The material agrees in all essential features with Westergård's (Reference Westergård1946) detailed description and Robison's (Reference Robison1988) and Peng & Robison's (Reference Peng and Robison2000) diagnosis of the species. Oidalagnostus trispinifer is a geographically widespread species that has been recorded from lower Guzhangian strata in Sweden, Australia, China, Siberia, western Newfoundland and North Greenland (Robison, Reference Robison1988; Peng & Robison, Reference Peng and Robison2000 and references therein).
Family Agraulidae Raymond, Reference Raymond1913
Remarks. As stated by, among others, Geyer & Landing (Reference Geyer and Landing2001), the agraulids are difficult to characterize as a natural group, and the family is in need of careful revision. Harrington et al. (Reference Harrington, Henningsmoen, Howell, Jaanusson, Lochman-Balk, Moore, Poulsen, Rasetti, Richter, Richter, Schmidt, Sdzuy, Struve, Tripp, Weller, Whittington and Moore1959) established the presence of a preglabellar field, the small size of the palpebral lobes (situated opposite the anterior half of the glabella) and the presence of a spine on the librigenae as the only diagnostic characters of the cephalon. According to Geyer & Landing (Reference Geyer and Landing2001), members of the family are generally characterized by a slightly to moderately convex glabella with a subtrapezoidal outline and short, relatively faint glabellar furrows. They also noted that ‘internal glabellar moulds show a tendency to development of a crestline with nearly flat, lateral abaxial slopes’ (Geyer & Landing, Reference Geyer and Landing2001, p. 131). The remainder of the cephalon is moderately convex with shallow, almost effaced furrows and short palpebral lobes (Geyer & Landing, Reference Geyer and Landing2001).
As revised by Geyer & Landing (Reference Geyer and Landing2001), the Agraulidae is a small family including Agraulos Hawle & Corda, Reference Hawle and Corda1847, Proampyx Frech, Reference Frech1897 and Skreiaspis Růžička, Reference Růžička1946. Ahlberg & Bergström (Reference Ahlberg and Bergström1978), Bergström & Ahlberg (Reference Bergström and Ahlberg1981) and Ahlberg (Reference Ahlberg1984) assigned a number of species from the provisional Cambrian Series 2 of Scandinavia to Proampyx. These are currently assigned to Ornamentaspis Geyer, Reference Geyer1990 and other ellipsocephalid genera (e.g. Geyer, Reference Geyer1990; Geyer & Landing, Reference Geyer and Landing2004).
According to Harrington et al. (Reference Harrington, Henningsmoen, Howell, Jaanusson, Lochman-Balk, Moore, Poulsen, Rasetti, Richter, Richter, Schmidt, Sdzuy, Struve, Tripp, Weller, Whittington and Moore1959), Agraulos is a ptychopariid with practically effaced cephalic furrows and a long frontal area. Thus, members of the genus are typically smooth and almost featureless and their relation to other agraulid genera is still a matter of discussion. Fletcher et al. (Reference Fletcher, Theokritoff, Lord and Zeoli2005) described topotype material of the type species and emphasized the subtriangular outline of the cranidium, with a sharply rounded, pointed anterior margin and shallow furrows on external moulds. The cephalic furrows are more distinct on internal moulds. They also noted that the ornamentation consists of ‘three sizes of punctae scattered over a background of tiny granules, with fine terrace ridges along the anterior cranidial margin and posterior edge of the occipital ring, over narrow stretches’ (Fletcher et al. Reference Fletcher, Theokritoff, Lord and Zeoli2005, p. 330). Four pairs of lateral glabellar furrows are present, extending over about one-third of the glabella, and the occipital ring is short (sag.) and strongly curved backwards into a blunt point (and not into the small spine as drawn by Šnajdr, Reference Šnajdr1958; Fletcher et al. Reference Fletcher, Theokritoff, Lord and Zeoli2005, p. 330). As a result, the characters of the type species appear to exclude Skreiaspis and Proampyx as close relatives. According to Fletcher et al. (Reference Fletcher, Theokritoff, Lord and Zeoli2005), A. longicephalus (Hicks, Reference Hicks1872) may represent an early variant of A. ceticephalus with a slightly more prominent occipital spine, and A. tosali (Sdzuy, Reference Sdzuy1968; formerly assigned to Skreiaspis) may be a junior synonym of A. holocephalus (Matthew, Reference Matthew1891).
Possibly related species from Scandinavia are Proampyx aculeatus (Angelin, Reference Angelin1851), P. acuminatus (Angelin, Reference Angelin1851), P. difformis (Angelin, Reference Angelin1851) and P. anceps (Westergård, Reference Westergård1953), all from the Guzhangian Andrarum Limestone. These four species have a long (sag.) frontal area that extends into a distinct swelling, cusp or spine at the anterior margin, a subtrapezoidal glabella with a crestline and relatively long palpebral lobes (Westergård, Reference Westergård1953; Geyer & Landing, Reference Geyer and Landing2001). They differ from Agraulos and Skreiaspis in having considerably longer palpebral lobes (Ahlberg & Bergström, Reference Ahlberg and Bergström1978; Geyer & Landing, Reference Geyer and Landing2001) and, following Ahlberg & Bergström (Reference Ahlberg and Bergström1978) and Geyer & Landing (Reference Geyer and Landing2001), they are assigned to the genus Proampyx.
Skreiaspis has generally been considered as closely related to Agraulos. Harrington et al. (Reference Harrington, Henningsmoen, Howell, Jaanusson, Lochman-Balk, Moore, Poulsen, Rasetti, Richter, Richter, Schmidt, Sdzuy, Struve, Tripp, Weller, Whittington and Moore1959) distinguished the former by the presence of a shorter frontal area and relatively well-impressed axial furrows. However, the expression of the furrows is controlled by taphonomic processes and the preservation in shale or limestone. Subsequently, Sdzuy (Reference Sdzuy1968) erected the genus Asturiaspis based on the presence of a relatively long frontal area, well-marked cranidial furrows and an angular anterior margin. The differentiation between these three genera is in need of revision based on more characters than the presence/absence of an anterior border furrow, the relative length of the frontal area and the palpebral lobes, and the distinctiveness of the cranidial furrows.
Genus Proampyx Frech, Reference Frech1897
Type species. Proetus? difformis var. acuminatus Angelin, Reference Angelin1851 from the Andrarum Limestone Bed (Guzhangian Stage; Lejopyge laevigata Zone) at Andrarum, Scania, southern Sweden, OD.
Proampyx difformis (Angelin, Reference Angelin1851)
Figure 5a–f
1953 Agraulos difformis (Angelin, Reference Angelin1851); Westergård, pp. 4–5, pl. 1, figs 1–8 (with previous synonymies).
1953 Agraulos aculeatus (Angelin, Reference Angelin1851); Westergård, p. 5, pl. 1, figs 9–10 (with previous synonymies).
1978 Proampyx difformis (Angelin, Reference Angelin1851); Ahlberg & Bergström, p. 22, pl. 3, fig. 4.
1985 Agraulos difformis; Berg-Madsen, table 1.
1994 Proampyx difformis (Angelin, Reference Angelin1851); Rudolph, pp. 182–3, pl. 24, figs 8–11.
2008 Agraulos difformis; Kouchinsky et al., appendix table 5.

Figure 5. (a–f) Proampyx difformis (Angelin, Reference Angelin1851): (a) dorsal view of internal mould, MPZ 2012/41; (b, c) lateral and dorsal views of external mould, MPZ 2012/42; (d) dorsal view of latex cast of external mould, MPZ 2012/43; (e, f) dorsal views of internal moulds, MPZ 2012/44–45. (g–i) Holasaphus cf. centropyge Matthew, Reference Matthew1895: (g) pygidium, latex cast of external mould, MPZ 2012/55; (h, i) cranidia, internal moulds, MPZ 2012/52–53. Scales of (a–g) and (i) equal 2 mm; scale of (h) equals 5 mm.
Material. Eleven cranidia preserved as internal and external moulds; (MPZ 2012/41–51).
Diagnosis of the cranidium (modified after Westergård, Reference Westergård1953). A species of Proampyx having a subrectangular glabella with three pairs of faint glabellar furrows; S1 bigeniculate and S2 geniculate on internal moulds; anterior border convex, separated from flat to slightly depressed preglabellar field; anterior margin rounded to angulate; anterior border varying in length and outline, from semicircular to subtriangular; occipital ring widening adaxially, bearing a median node or spine.
Description. Cranidium smooth, subquadrate in outline, gently convex, with length 90–95% of cranidial width across palpebral lobes (ratio decreases during ontogeny); all cranidial furrows well impressed, broad and shallow; glabella slightly elevated above genal and frontal areas, subrectangular in outline, slightly tapering forwards, about 70–75% (including occipital ring) of cranidial length, and 42–48% of cranidial width across palpebral lobes; transverse glabellar vaulting increasing forwards, from quite low near occipital ring to a maximum at about two-thirds length, in front of which it decreases in convexity to preglabellar furrow; three pairs of lateral glabellar furrows, faintly impressed, broad and short, gradually decreasing in length from S1 to S3; S1 and S2 curved slightly backwards medially; axial furrows shallow and broad; flat to slightly depressed preglabellar field and convex anterior border distinctly separated by anterior border furrow; anterior border slightly convex (parabolic) anteriorly and in some specimens slightly expanded backwards (mimicking a plectrum) along sagittal line; anterior border up to three times longer (sag.) than preglabellar field, and whole frontal area about 20–30% of cranidial length (sag.); occipital furrow generally straight, prominent and slightly wider medially; occipital ring about 25–30% of glabellar length, with evenly curved posterior margin, and bearing a median node or spine (depending on the preservation as internal v. external moulds, such as in Westergård, Reference Westergård1953, pl. 1, figs 9–10); fixigena down-sloping abaxially; palpebral lobes relatively long (exsagittal length about 30–35% of cranidial length, in juveniles about 40–45%), wide and moderately curved, anterior tip in front of S3 and posterior tip behind S1; preocular field weakly convex to flat, separated from convexity of fixigenae by distinct (robust in holaspides) eye ridge, reaching axial furrows in front of S3; anterior branch of facial suture up to twice as long as posterior branch, and parallel (adults) to slightly diverging (juveniles) anteriorly (making an angle of up to 30° with sagittal line), posterior branch short, distinctly diverging posteriorly; anterior cranidial margin variable, ranging from parabolic to evenly curved.
Remarks. Westergård (Reference Westergård1953) noted that P. difformis displays a wide range of morphological variation, particularly with respect to the relative length (sag.) of the anterior border and its outline, varying from semicircular to subtriangular, and the degree of expression of the anterior border furrow, although the impression of the furrow depends on the type of preservation. As stated by Westergård (Reference Westergård1953, p. 5), the only difference between P. difformis and P. aculeatus is the presence/absence of a ‘stout and long occipital spine instead of a faint node’. The Iberian material shows, however, that the occipital node is extended into a prominent, posteriorly directed spine on external moulds (see latex casts in Fig. 5b–d). Hence, the distinction between the two species seems to be a preservational effect, and we consider P. aculeatus to be a synonym of P. difformis. Other features noted in the description above change during ontogeny, for instance, the convexity and the relative length of the anterior border. Skreiaspis brianensis Courtessole, Reference Courtessole1973 (redescribed by Courtessole, Pillet & Vizcaïno, Reference Courtessole, Pillet and Vizcaïno1988), from the upper Languedocian La Gardie Formation of the Montagne Noire, France, is a similar species bearing a stout occipital ring. However, poor preservation of its described specimens precludes any comparison.
Stratigraphical distribution. P. difformis occurs in the Andrarum Limestone Bed (Solenopleura? brachymetopa Zone of Westergård, Reference Westergård1946) of the Alum Shale Formation, Scandinavia (Westergård, Reference Westergård1953; Berg-Madsen, Reference Berg-Madsen1985), and the Anomocarioides limbataeformis Zone of the Upper Ust’-Brus and lower Labaz formations, Kulyumbe section, Siberia (Kouchinsky et al. Reference Kouchinsky, Bengtson, Gallet, Korovnikov, Pavlov, Runnegar, Shields, Veizer, Young and Ziegler2008). The S.? brachymetopa Zone is currently included in the Lejopyge laevigata Zone, and faunal evidence suggests that the Andrarum Limestone Bed should be assigned to the lower L. laevigata Zone (Axheimer & Ahlberg, Reference Axheimer and Ahlberg2003; Axheimer et al. Reference Axheimer, Eriksson, Ahlberg and Bengtsson2006). The first appearance of the cosmopolitan agnostoid L. laevigata (Dalman, Reference Dalman1828) coincides with the base of the Guzhangian Stage (Peng et al. Reference Peng, Babcock, Zuo, Lin, Zhu, Yang, Robison, Qi, Bagnoli and Chen2009). In Scandinavia, the upper L. laevigata Zone corresponds to the Proagnostus bulbus Zone of South China and Siberia (Axheimer et al. Reference Axheimer, Eriksson, Ahlberg and Bengtsson2006). In Siberia, P. difformis has been recorded from the Anomocarioides limbataeformis Zone (Mayan Stage), which directly underlies the L. laevigata Zone (Kouchinsky et al. Reference Kouchinsky, Bengtson, Gallet, Korovnikov, Pavlov, Runnegar, Shields, Veizer, Young and Ziegler2008).
?Family Ptychopariidae Matthew, Reference Matthew1887
Genus Holasaphus Matthew, Reference Matthew1895
Type species. H. centropyge Matthew, Reference Matthew1895 from the Cambrian Series 3 of Nova Scotia, Canada, by monotypy.
Holasaphus cf. centropyge Matthew, Reference Matthew1895
Figure 5g–i
Material. Three cranidia (MPZ 2012/52–54) and three pygidia (MPZ 2012/55–57) preserved as internal and external moulds, respectively.
Description. Cranidium smooth, subtrapezoidal in outline, with rather convex glabella and fixigenae; cranidial length (sag.) about 70% of cranidial width across posterior border furrow; all furrows slightly impressed, except posterior border furrows and occipital furrow; glabella tapering forwards and broadly rounded anteriorly, about 80–84% of cranidial length, including occipital ring, and 50–54 % of cranidial width across palpebral lobes; glabellar furrows indistinct; preglabellar field flat, separated from anterior border by broad and shallow anterior border furrow; anterior border slightly convex, about twice as long (sag.) as preglabellar field; occipital and posterior border furrows wide and deep, the latter curved slightly forwards and disappearing abaxially; occipital ring about 20% of glabellar length, broadest (sag.) at sagittal line and with evenly curved posterior margin; posterior border widens abaxially; palpebral lobes short (exsagittal length about 10% of cranidial length), narrow, slightly curved, located opposite mid-length of glabella; eye ridges nearly effaced and curved gently towards anterolateral part of glabella; anterior branch of facial suture short, slightly divergent but becoming strongly convergent at anterior border furrow; posterior branch of facial suture up to twice as long as anterior branch, distinctly curved and diverging backwards, shaping uniform arc.
Pygidium smooth, slightly convex, widest (tr.) anteriorly, subtriangular in outline, and wider than long (excluding posterior spine); axis (excluding articulating half-ring) occupying 75% of total pygidial length (excluding spine); nearly parallel-sided to slightly tapered posteriorly; length about 70% of maximum pygidial width (tr.); segmentation of axis poorly defined, but comprising at least three faintly visible rings and well-rounded terminal axial piece; axial furrows faint, marked by change in slope; posterior border extending backwards into long and slender spine, with length (sag.) exceeding 50% of remainder of pygidium; pleural field flat to slightly down-sloping; border indistinct, marked by smooth strip lying marginally to faded ends of pleural furrows; pleural field made up of four fused pleurae, separated by faint interpleural furrows.
Comparison. The Iberian cranidia are very close to those of H. centropyge as described by Dean (Reference Dean1972) from Nova Scotia (Avalonia). However, the pygidia differ from H. centropyge in the nearly effaced interpleural furrows (see latex cast in Fig. 5g), although this character can be taphonomically induced; another difference is the lack of small punctae and granules displayed by the best-preserved specimens from Nova Scotia. The cranidium of H. centropyge differs from H. mesopotamicus Dean, Reference Dean1972 from the Cambrian Series 3 Sosink Formation, Turkey, in the narrower glabella with a more sharply curved front. The palpebral lobes are situated opposite the glabellar centre in the cranidia from Nova Scotia, whereas the posterior tip of the palpebral lobes is placed opposite the glabellar centre in the Turkish material, and hence the posterior part of the fixigenae is slightly smaller in the specimens from Nova Scotia. The pygidial spine of H. mesopotamicus Dean, Reference Dean1972 appears to be shorter and stouter than in pygidia of H. centropyge from Nova Scotia and Spain.
Stratigraphical distribution. The age of H. mesopotamicus is not well constrained, although it occurs at least 220 m above the Solenopleuropsis-bearing shales (upper Caesaraugustan–lower Languedocian) of the Sosink Formation (Dean, Reference Dean1972). In Nova Scotia, H. centropyge is associated with Paradoxides abenacus Matthew, Reference Matthew1886, which suggests a broad correlation with the lower part of the Paradoxides hicksii Zone in Newfoundland and the P. paradoxissimus Superzone of Scandinavia (see discussion in Dean, Reference Dean1972; cf. Fletcher, Reference Fletcher2007).
Family Paradoxididae Hawle & Corda, Reference Hawle and Corda1847
gen. et sp. indet.
Figure 6a–f
Material. Three incomplete cranidia (MPZ 2012/58–60) and two pygidia (MPZ 2012/61–62) preserved as internal and external moulds.

Figure 6. Paradoxidid fragments from the Guzhangian siliceous concretions of the Valtorres Formation at Purujosa. (a–c) Incomplete cranidia, internal moulds, MPZ 2012/58–60. (d–f) Incomplete pygidia, internal mould and latex cast of external mould, dorsal, posterior and left lateral views, respectively, MPZ 2012/61–62. All scales equal 5 mm.
Description. The cranidia exhibit the anterior part of the glabella and the adaxial part of the fixigenae. Glabella convex, widening forwards almost evenly, broadly rounded in front, and with four glabellar furrows; S1 and S2 transglabellar, subparallel and curved backwards medially; S3 and S4 short, shallow and not transglabellar; S3 almost transverse or curved slightly backwards medially, disconnected from axial furrows; S4 directed inwards and forwards adaxially. Frontal lobe of glabella rounded, almost semicircular; preglabellar field absent; anterior border much narrower in front of glabella than abaxially; anteriormost part of glabella and border covered with distinct terrace lines.
Pygidium smooth, subtrapezoidal in outline, widest posterior to the end of the axis, strongly convex transversely; pygidial length (sag.) about 85–90% of maximum width; axis gently convex (tr.), tapering gradually posteriorly and occupying 75% of sagittal length; axis laterally and posteriorly bounded by shallow, almost obsolete axial furrows; anteriormost axial ring separated from unfurrowed terminal piece by well-impressed, straight furrow; anterior width of axis 25% of maximum pygidial width; axis not reaching posterior border, but separated from it by short (20–25% of pygidial length) depression; pleural fields unfurrowed, in posterior view sloping rapidly downwards; border furrow absent; posterior margin indented by a prominent and wide posterior notch.
Discussion. The material is fragmentary, but the glabellar furrows, the medially narrow (sag.) anterior border and the outline of the glabella and the pygidium are suggestive of a species of Paradoxides. The material is, however, too incomplete for species and generic identification.
Family Conocoryphidae Angelin, Reference Angelin1854 (emended by Cotton, Reference Cotton2001)
Discussion. Fortey (Reference Fortey1990) and Geyer (Reference Geyer1998), among others, have suggested that the traditional concept of the Conocoryphidae (e.g. Poulsen, Reference Poulsen and Moore1959) may be an artificial grouping of blind taxa bearing marginal or submarginal sutures. Cotton (Reference Cotton2001) demonstrated that the family is polyphyletic and comprises four distantly related clades. He also showed that a monophyletic Conocoryphidae group should be restricted to comprise Bailiaspis Resser, Reference Resser1936, Bailiella Matthew, Reference Matthew1885, Cainatops Matthew, Reference Matthew1899 (= Cornucoryphe Sdzuy & Liñán, Reference Sdzuy and Liñán1996), Conocoryphe Hawle & Corda, Reference Hawle and Corda1847, Ctenocephalus Hawle & Corda, Reference Hawle and Corda1847, Elyx Angelin, Reference Angelin1854, Hartella Matthew, Reference Matthew1885, Parabailiella Thoral, Reference Thoral1946 and Tchaiaspis Korobov, Reference Korobov1966. This emended concept of the family has been followed by Kim, Westrop & Landing (Reference Kim, Westrop and Landing2002) and is also applied herein, along with Cotton's (Reference Cotton2001) revised diagnosis. Following Cotton's concept of the family, Occatharia Álvaro, Reference Álvaro2007 was added to this group based on the presence of fixigenal pseudoeyes (for a discussion of the genus see Álvaro & Vizcaïno, Reference Álvaro and Vizcaïno2003 and Álvaro, Reference Álvaro2007).
Another family close to the Conocoryphidae, the Holocephalidae Hupé, Reference Hupé1953 (emended by Cotton, Reference Cotton2001), is considered in this section and includes the following genera: Dasometopus Resser, Reference Resser1936, Holocephalina Salter, Reference Salter1864, Holocephalites Zhou in Zhou, Li & Qu, Reference Zhou, Li and Qu1982, Meneviella Stubblefield, Reference Stubblefield1951 (= Errinys, Menevia, Salteria) and Sdzuyella Hajrullina in Repina, Petrunina & Hajrullina, Reference Repina, Petrunina, Hajrullina, Repina, Yaskovich, Aksarina, Petrunina, Poniklenko, Rubanov, Bolgova, Golikov, Hajrullina and Posokhova1975. Holocephalina (type species: Holocephalina primordialis Salter, Reference Salter1864 from the Paradoxides davidis Zone at Porth-y-rhaw, Wales, UK, OD) is diagnosed as follows (modified after Fletcher et al. Reference Fletcher, Theokritoff, Lord and Zeoli2005): cephalon semielliptical, length exceeding half width, evenly convex, with very faint axial furrow; glabella slightly raised above genae; slightly more than half as long as cephalon and one-third as wide, narrowing forwards to a slightly rounded anterior, with three pairs of barely differentiated lateral furrows; occipital furrow well impressed; facial suture marginal; genal spines extending backwards to about fifth segment; thorax of 11–17 segments.
In his revision, Lake (1938) included in the genus H. primordialis (= H. teres Grönwall, Reference Grönwall1902), H. incerta Illing, Reference Illing1916 from the P. hicksii Zone Passage Beds, Hartshill, England, and H. meneviensis Hicks, Reference Hicks1872 from the Menevian Porth-y-rhaw, St David's, Wales. A specimen from the P. davidis Zone in Newfoundland was referred to H. americana by Resser (Reference Resser1937). H. incerta was referred to the new genus Holocephalites Zhou in Zhou, Li & Qu, (Reference Zhou, Li and Qu1982). The species described by Egorova et al. (Reference Egorova, Shabanov, Pegel, Savitsky, Suchov and Chernysheva1982) as H. aff. incerta and H. sp. also belong to Holocephalina (Cotton, Reference Cotton2001). H. americana and H. meneviensis are subjective junior synonyms of H. primordialis (M. Lewis, unpub. Ph.D. thesis, Univ. Wales, Cardiff, 1988). The concept of H. teres Grönwall, Reference Grönwall1902 is based on a drawing of a single cranidium (Grönwall, Reference Grönwall1902, pl. 2, fig. 2; redrawn in Harrington et al. Reference Harrington, Henningsmoen, Howell, Jaanusson, Lochman-Balk, Moore, Poulsen, Rasetti, Richter, Richter, Schmidt, Sdzuy, Struve, Tripp, Weller, Whittington and Moore1959, fig. 181.5) and is in need of revision. The occurrences of H. sp. nov. aff. H. levis from the Kiskinella Zone of Massachusetts and Newfoundland (Fletcher et al. Reference Fletcher, Theokritoff, Lord and Zeoli2005) and H. levis (leve) Gozalo & Liñán, Reference Gozalo and Liñán1996, from the Paradoxides asturianus Zone of the Iberian Chains, Spain are, in chronological order, apparently the oldest for the genus; these species are characterized by a (agrauloid) submarginal opisthoparian suture that displays incipient palpebral lobes on the fixigenae. The species discussed below is H. agrauloides Sdzuy, Reference Sdzuy1966 from the uppermost middle Cambrian (Guzhangian) Bergleshof Formation in the Franconian Forest, Germany. It most likely represents juveniles of Bailiaspis? glabrata.
Genus Bailiaspis Resser, Reference Resser1936
Type species. Conocephalites elegans Hartt in Dawson, Reference Dawson1868, from the Cambrian Series 3 of New Brunswick, Canada, OD.
Discussion. The distinction between Bailiella and Bailiaspis has been a matter of discussion. Westergård (Reference Westergård1950) noted that the medial thickening of the anterior border, a character used by Resser (Reference Resser1936, Reference Resser1937) to diagnose Bailiaspis, could be produced in two ways: ‘in some species, including the type species, the border “expands backwards” (i.e., has a true plectrum), and the anterior border furrow is bowed sharply posteriorly. In other species, such as Conocoryphe emarginata Linnarsson (Westergård, Reference Westergård1950, pl. 5, figs. 1–4; assigned to Bailiaspis by Resser, Reference Resser1936, p. 19), medial thickening reflects the course of the submarginal suture, and the anterior border furrow is not deflected backwards (i.e., a plectrum is absent)’ (Kim, Westrop & Landing, Reference Kim, Westrop and Landing2002, p. 830). A thickening of the anterior border has also been reported in other conocoryphid genera, including Conocoryphe (Dean, Reference Dean1982). Lake (1940, p. 283) suggested that Bailiaspis differs from Bailiella in a ‘depression of the preglabellar area so that the fixed cheeks are separated from each other instead of continuous’. As noted by Kim, Westrop & Landing (Reference Kim, Westrop and Landing2002), the depression is a result of inflated fixigenae in Bailiaspis, and according to these authors the genus comprises species with moderately to strongly inflated fixigenae.
Kim, Westrop & Landing (Reference Kim, Westrop and Landing2002) also discussed the discrimination between Bailiella and Bailiaspis, and noted that the former lacks a plectrum and has a border similar to other conocoryphid genera, including Conocoryphe and Elyx. The weakly inflated fixigenae in Bailiella may be a plesiomorphic character (Kim, Westrop & Landing, Reference Kim, Westrop and Landing2002). The course of the marginal suture and the morphology of the cephalic border and border furrow in Bailiella and Bailiaspis were briefly discussed by Kim, Westrop & Landing (Reference Kim, Westrop and Landing2002), who concluded that these characters cannot be used for discrimination between the two genera. They also emphasized that some species assigned to Bailiella (see Zhang & Jell, Reference Zhang and Jell1987) have lateral borders that extend backwards to join the posterior border, and that there is no compelling evidence for monophyly of Bailiella, nor probably of Bailiaspis either.
According to Kim, Westrop & Landing (Reference Kim, Westrop and Landing2002), it is unclear whether Bailiaspis is monophyletic because a plectrum is also present in the poorly known genus Tchaiaspis. The plectrum of its type species, T. sdzuyi Korobov, Reference Korobov1966 (see Cotton, Reference Cotton2001), ‘is unusually long and narrow, but is shorter and similar to the condition in Bailiaspis in the only other known species, T. sp. nov. St John & Babcock (Reference St John, Babcock, Dumoulin and Gray1997, fig. 6R–U)’ (Kim, Westrop & Landing, Reference Kim, Westrop and Landing2002, p. 830). Tchaiaspis is, however, a distinctive genus in having a well-defined paradoublural band at the inner edge of the lateral border furrow (Kim, Westrop & Landing, Reference Kim, Westrop and Landing2002). Cotton (Reference Cotton2001) did not resolve the relationship between Tchaiaspis and Bailiaspis, but some of his cladograms suggest that Tchaiaspis is best treated as a junior synonym of the latter.
Kim, Westrop & Landing (Reference Kim, Westrop and Landing2002) transferred B. howelli Hutchinson, Reference Hutchinson1962, B. cf. howelli of Hutchinson (Reference Hutchinson1962) and B. meridiana Sdzuy, Reference Sdzuy1958 (see also Sdzuy, Reference Sdzuy1961) to Bailiella. Following Kim, Westrop & Landing (Reference Kim, Westrop and Landing2002), Bailiaspis includes, at least, B. elegans (Hartt in Dawson, Reference Dawson1868), B. dalmani (Angelin, Reference Angelin1854), B. venusta Resser, Reference Resser1937 (= B. prominens Resser, Reference Resser1937), B. inflata Lake, 1940, B. tuberculata Lake, 1940, B. latigenae Hutchinson, Reference Hutchinson1962, B. menneri Korobov, Reference Korobov1973, B. bobrovi Korobov, Reference Korobov1973, B. jakutensis Korobov, Reference Korobov1973, B. botomensis Korobov, Reference Korobov1973, B. picta Korobov, Reference Korobov1973, B. curta Korobov, Reference Korobov1973 and B. senonta Korobov, Reference Korobov1973. Although Cotton (Reference Cotton2001) provided some evidence for assigning Conocoryphe? glabrata Angelin, Reference Angelin1854 to Bailiella, we follow Westergård (Reference Westergård1950) and Kim, Westrop & Landing (Reference Kim, Westrop and Landing2002) in tentatively treating it as a species of Bailiaspis. Bailiaspis nicholasi Resser in Lake (1940) seems to represent a species of Bailiaspis, but is based on poorly preserved and distorted material; Young et al. (Reference Young, Martin, Dean and Rushton1994) and Álvaro et al. (Reference Álvaro, González-Gómez and Vizcaïno2003) suggested that it is a junior synonym of B.? glabrata.
Bailiaspis? glabrata (Angelin, Reference Angelin1854)
Figure 7a–w
1940 Bailiaspis nicholasi Resser; Lake, pp. 286, 345, pl. 41, figs 1–4.
1950 Bailiaspis glabrata (Angelin, Reference Angelin1854); Westergård, pp. 30–1, pl. 6, figs 6–8 (with previous synonymies).
1966 Bailiella? glabrata (Angelin, Reference Angelin1854); Sdzuy, pp. 74–5, pl. 9, figs 16–24 (with previous synonymies).
1966 Holocephalina agrauloides n. sp.; Sdzuy, pp. 75–6, pl. 9, figs 9–15, pl. 10, fig. 4.
1973 Holocephalina cf. agrauloides Sdzuy, Reference Sdzuy1966; Courtessole, p. 215, pl. 26, figs 1, 2.
1988 Bailiaspis cf. glabrata; Pillet in Courtessole, Pillet & Vizcaïno, pp. 24, 29, 33, 45–6, figs 13, 18, table 2.
1988 Holocephalina cf. agrauloides Sdzuy, Reference Sdzuy1966; Courtessole, Pillet & Vizcaïno, pp. 24, 29, 33, 44, figs 13, 18, table 2.
1994 Bailiaspis glabrata (Angelin, Reference Angelin1854); Young et al., p. 348, 351, fig. 9g, k, m?, n.
1998 Bailiaspis cf. glabrata; Álvaro & Vizcaïno, fig. 4.
1998 Holocephalina cf. agrauloides; Álvaro & Vizcaïno, pp. 237–8, figs 3, 4.
2001 Bailiella glabrata; Álvaro et al., p. 208, fig. 1.
2001 Holocephalina cf. agrauloides; Álvaro et al., p. 208, fig. 1.
2001 Bailiaspis glabrata (Angelin, Reference Angelin1854); Cotton, p. 193, text-figs 2–5, table 2.
2002 Bailiaspis glabrata (Angelin, Reference Angelin1878); Kim, Westrop & Landing, p. 830.
2010 Bailiella? glabrata (Angelin, Reference Angelin1854); Geyer, p. 85, figs 4, 7.
2010 Holocephalina agrauloides Sdzuy, Reference Sdzuy1966; Geyer, p. 85, fig. 8.

Figure 7. Juvenile and adult specimens of Bailiaspis? glabrata (Angelin, Reference Angelin1854) from the Guzhangian siliceous concretions of the Valtorres Formation at Purujosa. (a–e) Juvenile cranidia, internal moulds, dorsal (a–d) and lateral (e) views. Scale of (a) equals 1 mm; scales of (b–e) equal 2 mm, MPZ 2012/63-66. (f–l) Adult cranidia, internal moulds (f, h, l) and latex casts of external moulds (g, i–k). All scales equal 5 mm, except for (h) which is 10 mm, MPZ 2012/67-70. (m–o) Enrolled and outstreched specimens. Scale of (m) equals 1 mm; scale of (n) equals 2 mm; scale of (o) equals 5 mm, MPZ 2012/57, 59–60. (p–w) Pygidia, internal moulds except (t) and (v) (latex casts of external moulds). Scale of (p) equals 2 mm; scales of (q–w) equal 5 mm, MPZ 2012/34–40.
Material. Seventy-one cranidia (MPZ 2012/63–133), 23 pygidia (MPZ 2012/134–156) and 20 complete or nearly complete specimens from which two are enrolled juveniles (MPZ 2012/157–176), preserved as internal and external moulds.
Diagnosis. Probable species of Bailiaspis with a true plectrum (a character controlled by size) and a facial suture that runs slightly outside instead of slightly inside the lateral border furrow.
Description of juvenile specimens (referred to below as morphotype H. agrauloides Sdzuy, Reference Sdzuy1966; Figure 7a–e, m). Cephalon semielliptical in outline, almost evenly curved to angulate anteriorly, with imperceptible anterior and lateral border furrows; glabella raised above genae, slightly wider (tr.) than fixigenae; frontal area about 75% glabellar length (sag.); glabellar width (tr.), at level of occipital furrow, about 120% of fixigenal width (tr.); occipital furrow curved slightly backwards medially; occipital ring long and subtriangular in outline.
Thorax with at least 11 thoracic segments, moderately decreasing in width posteriorly; axial furrows well defined and deep; width of axial ring (tr.) about 55% of pleural width (tr.) on each thoracic segment, strongly convex (tr.); axis slightly tapering backwards; articulating half-ring relatively long (sag.), length nearly twice that of axial ring; articulating furrow well marked and wide (sag. and exsag.); thoracic pleurae projecting horizontally to fulcrum, then downwards and posterolaterally to distal end; anterior pleural band narrowing abaxially to pleural tip, whereas posterior pleural band expanding; pleural furrow moderately deep and uniform in length (exsag.), not reaching the pleural tip.
Comparison of juvenile specimens. The Iberian specimens of the morphotype H. agrauloides differ from the type material from the Bergleshof Formation of Germany and H. cf. agrauloides from the upper Languedocian part of the La Gardie Formation in the Montagne Noire (Courtessole, Reference Courtessole1973) only in the relative length (sag.) of the preglabellar field (50 and 40% of total cephalic length in the German and French material, respectively) and the relative width of the glabella at the level of the occipital furrow (70 and 110% of fixigenal width, respectively); both the German and French material is preserved in shales, and some morphological differences and variability are associated with compaction and deformation. H. agrauloides differs from H. primordialis in the lack of a distinct anterior border furrow, and from H. levis in lacking dorsal sutures.
Discussion of juvenile specimens. It is worth noting that some characters vary with the size of the cranidia. Cephala of H. agrauloides are less than 9 mm wide in Spain, less than 7.5 mm wide in Germany and less than 7 mm wide in France. Cephala of H. primordialis generally do not exceed 10.6 mm in width (Cotton, Reference Cotton2001), and those of H. levis (Gozalo & Liñán, Reference Gozalo and Liñán1996) and H. aff. levis (Fletcher et al. Reference Fletcher, Theokritoff, Lord and Zeoli2005) are less than 21 and 13 mm, respectively. It is also worth noting that H. agrauloides co-occurs with specimens of Bailiaspis? glabrata (Angelin, Reference Angelin1854) in Germany, Spain and probably also France.
Description of adult specimens (Fig. 7f–l, n–w). Cranidium smooth, semielliptical in outline, with width greater than length (ratio 1.6–1.7); glabella subtrapezoidal, tapering forwards, bluntly rounded in front; glabella (including occipital ring) occupying about 70–75% of cranidial length; axial and preglabellar furrows well incised; S1 and S2 lateral furrows oblique and nearly effaced; occipital furrow relatively shallow but distinct and subtransverse medially, curved slightly forwards distally; occipital ring long (sag.) with evenly curved posterior margin; preglabellar field flat to slightly depressed, slightly longer than or subequal in length (sag.) to anterior border; border moderately convex and widest medially, narrowing laterally and posterolaterally; anterior border furrow shallow, subtransverse medially, then curving outwards and slightly backwards; fixigenae broad, but slightly narrower (tr.) than glabella; posterior border furrow broad and shallow, defining narrow posterior border; facial suture truncating lateral border furrow at the rear.
Thorax with at least 12 thoracic segments, moderately decreasing in width posteriorly; axial furrows deep and well defined; width of axial ring (tr.) approximately 55% width of pleura on each thoracic segment; axial rings strongly convex (tr.), straight medially, slightly tapering backwards, and slightly wider (exsag.) at their flanks; articulating half-ring relatively short (sag.), length slightly smaller than that of axial ring, posterior margin straight; articulating furrow distinct and wide (sag. and exsag.); thoracic pleurae projecting horizontally to fulcrum, then downwards and posterolaterally to distal end; anterior pleural band expanding abaxially to pleural end; posterior pleural band narrowing abaxially and becoming considerably narrower (exsag.) than anterior pleural band; pleural furrow moderately deep, wide (exsag.), narrowing abaxially and not reaching rounded pleural tip.
Pygidium fairly smooth, subelliptical in outline, about twice as wide as long, with anterior, horizontally orientated flanges; convex axis long, occupying about 90% of pygidial length, not reaching posterior margin; upstanding axis parallel-sided to backwardly tapered, comprising three (rarely two) axial rings and a rounded terminal piece; deep articulating furrow defining short articulating half-ring; axial and axial ring furrows prominent; three pairs of wide, distinct pleural furrows, anteriormost deeper than others; pleural furrows reaching axial furrow; border absent; posterior margin evenly curved.
Remarks on adult specimens. The librigenae and thorax are unknown from the Swedish material, but they have been recognized in southern Germany. As stated by Thoral (Reference Thoral1946) and Westergård (Reference Westergård1950), the course of the facial suture can be used for discrimination between Bailiella and Bailiaspis. A true plectrum is not clearly present in all specimens from Spain. According to the cladistic analysis of Cotton (Reference Cotton2001), Bailiaspis? glabrata should be assigned to Bailiella, as tentatively suggested by Sdzuy (Reference Sdzuy1966), but following Kim, Westrop & Landing (Reference Kim, Westrop and Landing2002) this species is here with caution assigned to Bailiaspis. Some poorly preserved specimens assigned to Bailiaspis cf. glabrata have been reported from the upper Languedocian strata of the Coulouma-Refescals sector of the southern Montagne Noire (Courtessole, Pillet & Vizcaïno, Reference Courtessole, Pillet and Vizcaïno1988; Álvaro & Vizcaïno, Reference Álvaro and Vizcaïno1998; Álvaro et al. Reference Álvaro, Lefebvre, Shergold and Vizcaïno2001), but further material is necessary to properly describe this assemblage.
In addition to the course of the facial suture, B.? glabrata differs from B. venusta (= B. prominens), from the Chamberlain's Brook Formation of southeastern Newfoundland, in the longer (sag.) preglabellar field and the lack of a cranidial sculpture (B. venusta has closely spaced granules); from B. elegans of New Brunswick in the lack of an occipital spine, less pronounced glabellar furrows and the lack of a cranidial surface sculpture; from the Swedish B. dalmani and the Siberian B. jakutensis, B. bobrovi, B. botomensis, B. picta, B. curta and B. senonta in the shorter (sag.) preglabellar field and more inflated fixigenae; from B. inflata of England in the shorter preglabellar field and the glabellar shape; from B. latigenae of the Manuels River Formation in Newfoundland in the shorter and less inflated fixigenae; from B. tuberculata of England in the lack of a surface sculpture; and from the Siberian B. menneri in the presence of a preglabellar field (Korobov, Reference Korobov1973; Kim, Westrop & Landing, Reference Kim, Westrop and Landing2002).
Ontogeny of Bailiaspis? glabrata. Owing to the abundance of B.? glabrata at Purujosa, an ontogenetic study is described below linking the problematic species H. agrauloides and Bailiaspis? glabrata. Digital photographs in dorsal view of the juvenile morphotype H. agrauloides and Bailiaspis? glabrata were acquired using an image analyser linked to a binocular microscope. Cranidia (n = 21) and pygidia (n = 14) were defined by 15 and 13 landmarks, respectively. All landmarks were projected in a single graph (Fig. 8). A standard orientation is given by a reference axis (sagittal axis). At the final stage, the graph was balanced by a symmetrical horizontal projection. This plotting of landmarks discloses growth tendencies and intraspecific variation during ontogeny, but also deformation.

Figure 8. Plotting of landmarks and related distances illustrating the cranidia and pygidia of Bailiaspis? glabrata for graphic and statistical analyses. Abbreviations of cranidial and pygidial linear dimensions measured are, for the cranidia: Lc – cranidial length, Lab – length anterior border, Lpf – length preglabellar field, Lg – preoccipital glabellar length, Lo – length occipital ring, Wc – cranidial width, Wg – glabellar width; for the pygidia: Lp – pygidial length, La – length axis, Lt – length terminal axial piece + posteriormost axial ring + Le, Le – length (sag.) posteroaxial area, Wp – pygidial width, Wa – length axis.
To quantify the ontogenetic trends, a set of bivariate scatter plots were established for the cranidia and pygidia (Fig. 9). The morphological variation in each of the linear characters was assessed with reference to a standard measure for size, in this case the width (tr.) of the cranidium. Data from linear characters were analysed using the Pearson square correlation coefficient (r2) (Pearson, Reference Pearson1900) to determine the extent to which two variables covary during growth. The coefficient varies between zero and one, with zero indicating no correlation and one indicating a perfect linear correlation.

Figure 9. Bivariate relationships of cranidia and pygidia of Bailiaspis? glabrata. Each linear character is plotted against a standard measure for size, in this case, the cranidial and pygidial width; see Figure 8 for abbreviations.
A growth series for meraspid and holaspid stages is shown. Meraspides and juvenile holaspides with up to 11 thoracic segments were previously assigned to H. agrauloides. The absence of sclerites from considerably larger specimens suggests that a complete holaspid ontogeny of B.? glabrata is documented. An uninterrupted growth series ranging from 2.6 to 39.4 mm in cranidial width (Wc) is represented. Half of the specimens occur as partly articulated sclerites.
The morphometric analyses show no discontinuous populations clustered into instar groupings. This is probably owing to intraspecific variation in size within the growth stages and the resulting overlap in size between adjacent instars. There is no evidence for drastic morphological changes during the ontogeny, and the co-occurrence of all the stages, commonly preserved as partly articulated specimens, indicates that they were once living in the same environment and growth may not have involved any significant transformation in the mode of life.
The ontogenetic trends can be summarized as follows:
(1) The growth series of the cranidium does not show remarkable changes in growth isometry. With growth, the convexity of the glabella and cheeks increased; the axial, glabellar and occipital furrows deepened and became enlarged; the cranidium became relatively wider in adult stages due to the lateral extension of the glabella and fixigenae (see Lc/Wc, Wg/Lg and Wg/Wc ratios in Fig. 9); the anterior margin of the cranidium is clearly pointed in small specimens and becomes more evenly curved with growth. The glabella is subtrapezoidal in outline and becomes more tapered forwards during growth; there is a gradual decrease in length (sag.) of the occipital ring, partly related to a change in the shape of the occipital furrow during ontogeny, which changes from relatively straight in small cranidia to become curved backwards medially in larger cranidia.
Once the cranidial series reached 9.2 mm in width, a pair of faint lateral fossulae appeared at the anterolateral margin of the cranidium. These subsequently migrated adaxially during the next holaspid stages, forming a distinct anterior border furrow that progressively widened and flattened. The last growth stages (Wc > 18.5 mm) display a distinct fold of the furrow, leading to the appearance of a plectrum.
(2) The growth series of the pygidium shows no allometries but displays a high degree of morphological plasticity. Independently of the growth stage, the terminal axial piece can exhibit an obsolete furrow, which is reflected in the pleural field by an additional incipient pleural rib. The posterior margin of the pygidium ranges from nearly straight to slightly convex.
Thus, the ontogenetic reconstruction shows that species previously referred to as Holocephalina agrauloides (Wc < 9.2 mm), Bailiella? glabrata (sensu Sdzuy, Reference Sdzuy1966) (9.2 < Wc < 18.5 mm) and Bailiaspis? glabrata (Wc > 18.5 mm) most likely represent three growth stages of a single species. It also strongly indicates that H. agrauloides should be considered as a junior synonym of B.? glabrata and that a direct phylogenetic link should exist between the conocoryphids and the holocephalids.
Stratigraphical range and age. The species occurs in the lower Guzhangian Andrarum Limestone Bed of the Alum Shale Formation (currently assigned to the lower L. laevigata Zone; Axheimer et al. Reference Axheimer, Eriksson, Ahlberg and Bengtsson2006) of Sweden, in the Bergleshof Formation of the Franconian Forest, Germany (Sdzuy, Reference Sdzuy1966) and in the ‘calcareous grit’ of the Maentwrog Formation of St Tudwal's, Wales. B. cf. glabrata (Pillet in Courtessole, Pillet & Vizcaïno, Reference Courtessole, Pillet and Vizcaïno1988) occurs in the upper Languedocian part of the La Gardie Formation, Montagne Noire.
5. Mid–late Languedocian benthic community replacements
The trilobites described above from the basal part of the Valtorres Formation are important for correlation with areas outside Spain: Peronopsis cf. insignis is close to the species described from the Lejopyge laevigata Zone of Scandinavia; Oidalagnostus trispinifer is geographically widespread and has been recorded from the Guzhangian Stage in Sweden, western Newfoundland, Siberia, China, North Greenland and Australia; Proampyx difformis and Bailiaspis? glabrata (= H. agrauloides) occur in the Solenopleura? (or Erratojincella) brachymetopa Zone of the Andrarum Limestone of southern Scandinavia; Bailiaspis? glabrata has also been recorded from the uppermost Cambrian Series 3 Bergleshof Formation of Germany and probably from upper Languedocian (Eccaparadoxides macrocercus Zone) strata of the Montagne Noire; and specimens here identified as Holasaphus cf. centropyge are close to topotype and other material of the species from the upper Cambrian Series 3 of Nova Scotia, Canada (see Dean, Reference Dean1972). The Solenopleura? brachymetopa Zone of Scandinavia reflects a facies change, and because Lejopyge laevigata appears even below the base of this zone (Axheimer et al. Reference Axheimer, Eriksson, Ahlberg and Bengtsson2006; Ahlberg et al. Reference Ahlberg, Axheimer, Babcock, Eriksson, Schmitz and Terfelt2009), it is currently included in the global Lejopyge laevigata Zone that marks the base of the global Guzhangian Stage; the upper part of the Scandinavian Lejopyge laevigata Zone can be broadly correlated with the Proagnostus bulbus Zone of South China (Axheimer et al. Reference Axheimer, Eriksson, Ahlberg and Bengtsson2006), which directly overlies the global L. laevigata Zone. The trilobite assemblage described herein indicates that the lowermost part of the Valtorres Formation belongs to part of the upper Languedocian Substage in the Mediterranean chronostratigraphical scheme and the global Guzhangian Stage.
The trilobites described herein show that the conocoryphid–paradoxidid–solenopleurid assemblage, characteristic of the Leonian–early Languedocian benthic communities throughout the Acado-Baltic Province sensu Sdzuy (Reference Sdzuy1972), persisted into upper Languedocian clayey substrates. However, the palaeobiogeographic patterns of the late Languedocian trilobite immigration were different in mixed (East Gondwana invaders in the Montagne Noire, such as Abharella, Chelidonocephalus, Derikaspis and Dorypyge; Courtessole, Pillet & Vizcaïno, Reference Courtessole, Pillet and Vizcaïno1988; Álvaro et al. Reference Álvaro, González-Gómez and Vizcaïno2003, Reference Álvaro, Ferretti, González-Gómez, Serpagli, Tortello, Vecoli and Vizcaïno2007) and clayey substrates (Baltic invaders in the Iberian Chains). The late Languedocian represents the beginning of a broad transgression that persisted until mid Tremadocian times in the Montagne Noire (Álvaro et al. Reference Álvaro, Ferretti, González-Gómez, Serpagli, Tortello, Vecoli and Vizcaïno2007). This transgression could be a eustatic rise in sea level in early Guzhangian times. It has been identified in South China and elsewhere by Peng et al. (Reference Peng, Babcock, Zuo, Lin, Zhu, Yang, Robison, Qi, Bagnoli and Chen2006, Reference Peng, Babcock, Zuo, Lin, Zhu, Yang, Robison, Qi, Bagnoli and Chen2009). In North Wales, the Guzhangian transgression appears to be represented by the Maentwrog Formation, which directly overlies a distinct stratigraphic gap spanning the lower Guzhangian (Harlech Dome) or the St David's/Merioneth (British) Series transition (St Tudwal's Peninsula; Young et al. Reference Young, Martin, Dean and Rushton1994).
The appearance of late Languedocian trilobite assemblages in southwestern Europe represents a stepwise immigration event related to transgressive pulses: clayey offshore substrates migrated towards the inner platform, accompanied by facies-controlled off-platform trilobites. The inner-platform settings were recolonized from an oceanic stock, where the conocoryphid–paradoxidid–solenopleurid association was still present. Babcock (Reference Babcock1994aReference Babcockb) pointed out that this assemblage of trilobites was widely distributed in cool marine waters of various latitudes, including deep-water substrates surrounding the tropical palaeocontinent of Laurentia.
A further character of the late Languedocian trilobite assemblage found in Purujosa is the abundance of Bailiaspis? glabrata, which represents about 76% of the total number of specimens in the association (the remaining percentages are: Proampyx and Peronopsis, 7.2%; Holasaphus, 3.9%; Oidalagnostus and paradoxidids, 3.2%, respectively). The relative abundance of blind trilobites in benthic communities living on offshore, clayey substrates during Cambrian Epoch 3, led Álvaro & Vizcaïno (Reference Álvaro, González-Gómez and Vizcaïno2003; see also Cocks & Fortey, Reference Cocks, Fortey and Bassett2009) to recognize the conocoryphid biofacies. This biofacies is characterized by autochthonous epibenthic assemblages of normal-eyed and blind trilobites, displaying similar patterns of disarticulation and fragmentation (Álvaro & Vizcaïno, Reference Álvaro, González-Gómez and Vizcaïno2003). In bottom conditions of increasing sedimentation rates and turbidity, muds lacking cohesion are easily resuspended to generate high-turbidity conditions. As a result, mud suspended in water may greatly reduce the amount of light reaching the seafloor, and dark to dimly lit substrate conditions can be formed. For microphagous feeders, the lack or loss of eyes would not have affected their feeding habits, but predation against them might well have been greater. However, if oxygenated, deep and/or dark waters prevailed, predators would certainly have been at a disadvantage, and the selection pressure against blind and eye-reduced trilobites, such as Bailiaspis, would have been sharply reduced. Blind species were viable in shallow waters as microphagous feeders, only becoming dominant in particular ecological conditions of high turbidity (Álvaro & Vizcaïno, Reference Álvaro and Vizcaïno2003). The abundance of blind trilobites may reflect a dark or dim environment where normal-eyed trilobites with predatory habits (such as the agraulids and paradoxidids) were, at least episodically, at a disadvantage.
6. Conclusions
In mixed (carbonate-siliciclastic) substrates, such as those preserved in the Montagne Noire (France), the aftermath of the mid Languedocian regression is characterized by a late Languedocian major turnover of trilobite families, followed by a renewed Furongian–early Tremadocian radiation related to the stepwise immigration of trilobite invaders from East Gondwana under persistently transgressive conditions.
In clayey substrates, such as those preserved in palaeogeographically distal parts of the Iberian Chains (Spain), biogenic silica, formed through dissolution of hexactinellid spicules, has locally segregated into fossiliferous concretions. These have yielded the trilobites Peronopsis cf. insignis, Oidalagnostus trispinifer, Proampyx difformis (= Proampyx aculeatus), Bailiaspis? glabrata (= Holocephalina agrauloides, by ontogeny), Holasaphus cf. centropyge and paradoxidid gen. et sp. indet. The identification of this preservational window in clayey offshore substrates allows the recognition of a distinct biogeographical link with Baltica, and a correlation of the Mediterranean upper Languedocian Substage with the global Guzhangian Stage and the Baltic Solenopleura? brachymetopa Zone (lower Lejopyge laevigata Zone in the global agnostoid zonation). Taphonomic and sampling biases are major factors that control the knowledge of biodiversity patterns in Cambrian Epoch 3 siliciclastic and mixed platforms of West Gondwana and must be taken into account when describing ecostratigraphical patterns.
The evolution of the Cambrian Epoch 3 trilobite faunas on the Mediterranean margin of West Gondwana was disturbed by a major biotic crisis, related to the onset of the mid Languedocian regression. Despite immigration and adaptive radiations, or a combination of both, after the regression, the trilobite diversity was considerably lower during mid and late Languedocian times than during the preceding Caesaraugustan–early Languedocian biodiversity peak.
Acknowledgements
The authors thank Anna Żylińska and an anonymous referee for their kind and constructive comments, Fernando Gracia for help with field sampling, and Isabel Pérez (MEC-FSE) for assistance with photography. SZ has received a postdoctoral fellowship (EX2009-0815) from the Spanish MICINN. This paper is a contribution to projects CGL2006-12975 and CGL2010-19491 from the Spanish MICINN and FEDER.









