INTRODUCTION
The clam is among the 10 most noticeable bivalves produced by European aquaculture. This production mainly relies on 2 different species: the Japanese carpet shell clam Ruditapes philippinarum and the grooved carpet shell clam R. decussatus with an estimated production of 68 010 t and 7994 t respectively in 2009 if we include aquaculture and fisheries figures (FAO, 2009). In France, clam production, estimated at 1263 t (FAO, 2009), is the third most important bivalve production after Crassostrea gigas and mussels. This production is mainly concentrated in Arcachon Bay and South Brittany (D'Hardivillé et al. Reference D'Hardivillé, Bouché and Péronnet2010a, Reference D'Hardivillé, Bouché and Péronnetb; Sanchez et al. Reference Sanchez, Caill-Milly, Lissardy, de Casamajor and Morandeau2010). However, several natural beds are present along the French coasts including lagoons like Leucate Lagoon or semi-closed bays like Gulf of Morbihan and Bonne Anse in Charente Maritime (Fig. 1). The Japanese carpet shell clam is mainly present in North and West of France while the grooved carpet shell clam is dominant in lagoons from the Mediterranean Coast (Garcia et al. Reference Garcia, Arzul, Chollet, François, Goubet, Joly, Miossec and Robert2006). In spite of being fairly well organized and regulated, the French clam industry is still facing some problems with economic consequences for producers, including diseases such as the brown ring disease due to Vibrio tapetis (Paillard, Reference Paillard2004) or the Brown Muscle Disease (Dang et al. Reference Dang, de Montaudouin, Gonzalez, Mesmer-Dudons and Caill-Milly2008). In addition, clam population dynamics may be affected by the presence of parasites belonging to the genus Perkinsus (Lassalle et al. Reference Lassalle, de Montaudouin, Soudant and Paillard2007).

Fig. 1. Locations where clams Ruditapes decussatus or R. philippinarum were collected for the current study.
Microorganisms of this genus have frequently been associated with important mortality events in different mollusc species worldwide. The first described species in this genus was Perkinsus marinus in the 1940s in the USA during mass mortality of the American oyster Crassostrea virginica (Ray, Reference Ray1996). Perkinsus olseni was then reported and described in the abalone species Haliotis ruber and H. laevigata in Australia (Lester and Davis, Reference Lester and Davis1981; Goggin and Lester, Reference Goggin and Lester1995). A Perkinsus species initially named P. atlanticus was described in the grooved carpet shell clam R. decussatus following mortality events in Portugal (Azevedo, Reference Azevedo1989). Molecular investigations demonstrated that P. olseni was a senior synonym of P. atlanticus (Murrell et al. Reference Murrell, Kleeman, Barker and Lester2002). More recently other Perkinsus species have been characterized: P. chesapeaki, a parasite of the soft-shell clam Mya arenaria (McLaughin et al. Reference McLaughin, Tall, Shaheen, Elsayed and Faisal2000) which is the senior synonym of P. andrewsi detected in the Baltic clam Macoma balthica (Burreson et al. Reference Burreson, Reece and Dungan2005) present on the eastern coast of the USA; P. qugwadi associated with mortalities of Japanese scallop Patinopecten yessoensis in Canada (Bower et al. Reference Bower, Blackbourn and Meyer1998); Perkinsus mediterraneus described in flat oysters Ostrea edulis in Balearic Islands, Spain (Casas et al. Reference Casas, Grau, Reece, Apakupakul, Azevado and Villalba2004); P. honshuensis described in the Japanese carpet shell clam R. philippinarum in Japan (Dungan and Reece, Reference Dungan and Reece2006) and P. beihaiensis characterized from oyster species Crassostrea ariakensis and C. hongkongensis of Souhern China (Moss et al. Reference Moss, Xiao, Dungan and Reece2008). These characterizations are based on initial detection using non-specific assays like histology or culture in Ray's Fluid Thioglycolate Medium (RFTM), geographical distribution, host range, parasite morphology especially the description and comparison of different stages of several in vitro isolates and on phylogenic analysis. In Europe, Perkinsus sp. parasites have been reported in France, Spain, Portugal and Italy. However, some of these reports are only based on genus-specific assays. From the molecular characterization of the 18S and ITS rDNA sequences conclusions could be made regarding the presence of P. olseni in Ruditapes decussatus from Catalonia and Galicia in Spain and from Adriatic Sea in Italy (Elandaloussi et al. Reference Elandaloussi, Carrasco, Furones and Roque2009; Casas et al. Reference Casas, Villalba and Reece2002; Abollo et al. Reference Abollo, Casas, Ceschia and Villalba2006) and about the presence of P. mediterraneus in Ostrea edulis from Balearic Islands (Casas et al. Reference Casas, Grau, Reece, Apakupakul, Azevado and Villalba2004).
While high clam mortality associated with perkinsosis has been recorded in European affected areas (Ruano and Cachola, Reference Ruano and Cachola1986; Azevedo, Reference Azevedo1989; Figueras et al. Reference Figueras, Robledo and Novoa1992; Villalba et al. Reference Villalba, Lopez, Carballal, Cervino, Landin, Coo de, Guerra and Torre1993), the actual impact of perkinsosis on clam production is still under debate (Villalba, Reference Villalba2008) and seems to be positively related to environmental factors such as water temperature and salinity (Cigarria et al. Reference Cigarría, Rodríguez and Fernández1997; Casas et al. Reference Casas, Villalba and Reece2002; Villalba et al. Reference Villalba, Casas, Lopez and Carballal2005). Furthermore, Choi et al. (Reference Choi, Park, Lee and Matsuoka2002) reported that Perkinsus sp. infection levels relate significantly to sediment type: clams living on muddy flats tend to have higher levels of infection than those on sandy tidal flats. Considering the lack of data related to Perkinsus species in France and that infection with Perkinsus olseni is notifiable to the World Organization for Animal Health (OIE 2011), the study reported here was undertaken to specifically identify parasites of this genus occurring in different French clam-growing areas. For that purpose, clams collected from different areas and shown to be infected by RFTM assays were selected for PCR-RFLP tests and sequencing works. Perkinsus olseni was detected in most of the investigated areas and unexpected results were obtained in 2 locations: Leucate Lagoon in the south of France and Bonne Anse in the Charente Maritime on the middle West coast of France (Fig. 1). Additional sampling was carried out in Leucate Lagoon. Clams were treated in order to study intra- and inter-individual genetic variability of the parasite. Moreover, parasites were cultivated in order to describe in vitro stages. Taken together, results obtained during this study are indicative of the presence of 2 Perkinsus species P. olseni and P. chesapeaki in 2 French clam-production areas.
This is the first report of P. chesapeaki infections among clams outside of North America, the first report of P. chesapeaki infections among Ruditapes decussatus and R. philippinarum clams, and the first report of P. chesapeaki in vitro isolates from European clams.
MATERIALS AND METHODS
Sampling
Clams, including Ruditapes decussatus and R. philippinarum, were collected in October 2004 and 2005 during an epidemiological survey aiming at establishing the distribution, prevalence and infection rates of perkinsosis in main French clam production areas (Fig. 1). For some samples, gill tissues were processed for in vitro parasite culture (see section ‘culture’ below). Clams from the Golfe of Morbihan, Charente Maritime and Arcachon consisted of Japanese carpet shell clams R. philippinarum while clams from Leucate consisted of grooved carpet shell clams R. decussatus.
Additional samplings were carried out in Leucate Lagoon (60 clams R. decussatus) in October 2008 and in Bonne Anse, Charente Maritime, in September 2010 (30 clams R. philippinarum). Pieces of gills collected from these clams were ground and processed for in vitro parasite culture in order to describe parasite stages (see below section ‘culture’). Clams from Leucate were also processed in order to test them by in situ hybridization.
DNA extraction
Sixty-four samples of cryopreserved or fresh parasite cultures were used for DNA extraction. For cultures obtained from clams collected before 2008, DNA was extracted using the QIAamp DNA minikit (Qiagen) according to the manufacturer's instructions. DNA was eluted and resuspended in a final volume of 50 μl of sterile deionized water and then diluted at a final concentration of 100 ng μl−1. For cultures obtained from clams collected in 2008 or after, cells were concentrated by centrifugation (150 g for 5 min) and 100 μl of cell suspension (minimum concentration 105cells ml−1) were placed and stored on a QIAcard FTA spot (Qiagen) according to the manufacturer's instructions. QIAcard FTA spot was used after 2008 to simplify the handling and processing of DNA from parasite cultures.
PCR, PCR-RFLP, cloning and sequencing
Three PCR assays targeting sequences of rDNA internal transcribed spacer regions (ITS), large subunit rRNA genes (LSU), and actin-1 genes were used in order to detect and characterize parasites isolated during this study. All the PCR reactions were performed using the Go Taq Polymerase (Promega) in a final volume of 25 μl. DNA (50–100 ng μl−1 or QIAcard FTA disk) was added to 24 μl of the PCR mix presented in Table 1. Negative PCR controls consisting of water were included for every 10 tested samples in order to check potential contamination. A positive PCR control corresponding to DNA extracted from the culture of previously characterized P. olseni from Arcachon Bay was included in each PCR test.
Table 1. Conditions and primer sequences used for the PCR assays

First, parasite species was determined using the PCR-RFLP approach developed by Abollo et al. (Reference Abollo, Casas, Ceschia and Villalba2006). Analyses were performed by separate digestions for 2 h at 37 °C of 10 μl of PerkITS750-85 with 0·25 units of RsaI and HinfI (Promega). Enzymes were inactivated for 20 min at 65 °C and the resulting fragment patterns were analysed electrophoretically on 2% agarose gel.
Some PCR products were cloned using the original TOPO TA cloning kit (Invitrogen) according to the manufacturer's recommendations and positive clones were then selected for plasmid DNA purification by FastPlasmid® Mini (Eppendorf). Some plasmidic DNA suspensions were bidirectional sequenced using the Big Dye V3 sequencing kit (Applied Biosystem) and standard M13 forward and reverse primers. Obtained sequences were compared with those included in GenBank using BLAST algorithm (Atschul et al. Reference Altschul, Madden, Schaffer, Zhang, Zhang, Miller and Lipman1997).
Phylogenetic analyses
Available ITS, LSU and actin gene sequences from Perkinsus spp. and dinoflagellates were downloaded from GenBank and included in phylogenetic analyses (Table 2) with sequences obtained in the present study.
Table 2. GenBank Accession numbers of rRNA-ITS, rRNA-LSU and actin-1 gene sequences used in phylogenetic analyses

Alignments were performed using Clustal W (Thompson et al. Reference Thompson, Higgins and Gibson1994) including in MEGA 5 with open and extended gap penalties of 7 and 3, respectively. Parsimony analyses were also conducted using MEGA version 5 (Tamura et al. Reference Tamura, Peterson, Peterson, Stecher, Nei and Kumar2011) with the close neighbour interchange (CNI) heuristic option. Bootstrap values were calculated over 100 replicates and cut-off value for condensed tree was of 50%. Phylogenetic analysis of the translated actin gene sequences was performed using deduced amino acid sequences.
Genetic distance corresponding to the number of base substitutions per site from averaging over all sequence pairs was estimated using the Tajima-Nei model (Tajima and Nei, Reference Tajima and Nei1984) in MEGA5 (Tamura et al. Reference Tamura, Peterson, Peterson, Stecher, Nei and Kumar2011).
In vitro culture of Perkinsus spp. and cryopreservation
Gill tissues were rinsed 3 times in sterile seawater (SSW) and then decontaminated twice for 30 min in 30 ml of SSW supplemented with an antimicrobial suspension (penicillin G 61·3 mg L−1; streptomycin sulphate 131 mg L−1; nystatin 50 000 U L−1). Tissues were rinsed again 3 times in SSW and ground in 250 μl of the antimicrobial suspension using a disposable homogenizer. The suspension was mixed and complemented with 750 μl of the antimicrobial suspension before incubation for 15 min at room temperature. Homogenized tissues were centrifuged at 12 000 g for 5 min, supernatants were discarded and pellets were resuspended with 1 ml of SSW. Suspensions were then filtered at 100 μm and 50 μl of the filtrates were finally inoculated into 6 wells per sample, each containing 1 ml of antimicrobial and salt-supplemented Dulbecco's modified Eagle's/Ham's F12 (1:2) medium (Gauthier and Vasta, Reference Gauthier and Vasta1995; Ordas and Figueras, Reference Ordás and Figueras1998) in 24-well culture plates. Culture plates were incubated at 26 °C in a humidified air atmosphere chamber and regularly observed under an inverted microscope.
Some clonal cultures were established by limiting dilution of the above mentioned cultures according to Gauthier and Vasta (Reference Gauthier and Vasta1995). These clonal cultures were used to describe in vitro parasite stages.
Parasite cultures were cryopreserved in medium consisting of 10% DMSO, 50% fetal bovine serum and 40% DME: Ham's F12 (1:1) according to the method described by Gauthier and Vasta (Reference Gauthier and Vasta1995).
Cells from 4 clonal cultures from 2 clams characterized as Perkinsus olseni and cells from 3 clonal cultures from 1 clam characterized as P. chesapeaki were observed and measured after 16 days of culture. For that purpose, 20 μl of each clonal culture were collected twice and observed microscopically using a Malassez-cell haemocytometer. All the cells present in the Malassez-cell haemocytometer were observed in order to estimate proportions of trophozoïtes, schizonts and zoosporangia. Mean cell diameter of each cell type was determined by measuring 100–200 trophozoïtes and all the schizonts and zoosporangia using the Analysis software (Olympus). Finally, for each Perkinsus species, we estimated the mean proportion and cell diameter of each parasite stage by averaging data obtained from the different conspecific clonal cultures.
Histology and in situ hybridization (ISH)
After 48 h in Davidson's fixative, half clam soft tissues were maintained in 70% ethanol until they were dehydrated and embedded in paraffin for histology according to standard procedures. Paraffin blocks were cut in 2–3 μm sections and stained by haematoxylin and eosin.
The protocol followed for ISH was adapted from Moss et al. (Reference Moss, Burreson and Reece2006) and Reece et al. (Reference Reece, Dungan and Burreson2008). The digoxigenin-labelled probes consisted of a Perkinsus olseni-specific LSU-rRNA gene probe (Moss et al. Reference Moss, Burreson and Reece2006) PolsLSU-464DIG (5′-CTCACAAGTGCCAAACAACTG-3′) and Perkinsus chesapeaki-specific LSU-rRNA gene probe (Reece et al. Reference Reece, Dungan and Burreson2008) PchesLSU-485DIG (5′-CAG GAA ACA CCA CGC ACK AG-3′).
Five μm thick tissue sections on silane-prep™ slides (Sigma, France) were dewaxed, rehydrated, and treated with proteinase K (100 μg ml−1 in TE buffer [Tris 50 mM, EDTA 10 mM]) at 37 °C for 5 min. Slides were dehydrated by immersion in an ethanol series and air-dried. Sections were then incubated with 100 μl of hybridization buffer (50% formamide, 10% dextran sulfate, 4 × SSC [0·06 M Na3 citrate, 0·6 M NaCl, pH 7], 250 μg ml−1 yeast tRNA and 10% Denhardt's solution) containing 7 ng μl−1 of digoxigenin-labelled probes (Eurogentec). Target DNA and digoxigenin-labelled probe were denatured at 95 °C for 5 min and the hybridization was carried out overnight at 42 °C. Sections were washed in 2 × SSC at room temperature (RT) (2 × 5 min), in 0·4 × SSC at 42 °C (10 min) and in solution I (100 mM maleic acid, 0·15 M NaCl, pH 7·5) for 5 min. Tissues were then blocked for 30 min at room temperature with blocking reagent (Amersham Life Science) (1% w/v) in solution I. Specifically bound probe was detected using an alkaline phosphatase-conjugated mouse IgG antibody against digoxigenin diluted at 1·5 U ml−1 in solution I (1 h, RT). Excess of antibody was removed by 2 washes in solution I (1 min) and 1 wash in solution II (0·1 M Tris pH 8, 0·1 M NaCl, 0·05 M MgCl2, pH 9·5). Slides were incubated in NBT/BCIP, a chromogenic substrate for alkaline phosphatase, diluted in solution II (20 μl ml−1) in the dark until the parasitic cells are completely stained black-purple. The reaction was stopped with solution III (100 mM Tris, 1 mM EDTA, pH 8). Slides were counterstained for 1 min with Bismarck brown yellow (5 mg ml−1), dehydrated with ethanol and mounted in Eukitt resin. Negative controls included samples without digoxigenin-labelled probe in hybridization mixture or without antibodies during colour development. The positive control consisted of sections from Ruditapes philippinarum infected with Perkinsus olseni originating from Arcachon Bay (France) and Mya arenaria infected with P. chesapeaki (kindly provided by Dr R. Carnegie).
RESULTS
Characterization of parasites of the genus Perkinsus detected in clam producing areas in France
In total, 30 and 33 in vitro Perkinsus sp. isolate cultures were propagated from clams R. philippinarum and R. decussatus respectively (Table 3).
Table 3. Number of tested clams and cultures tested per clam for each locations included in the present study

Direct PCR-RFLP was performed on 5 cultures from Arcachon Bay, 8 cultures from Morbihan Gulf, 17 cultures from Bonne Anse, Charente Maritime, 33 cultures from Leucate Lagoon (Table 3). Restriction profiles after RsaI and HinfI digestion appeared similar to Perkinsus olseni profiles (around 413 bp, 193 bp, 74 bp after digestion with RsaI and around 363 bp, 160 bp, 150 bp after digestion with HinfI) for all the tested samples except for 7 cultures from 5 clams collected in Leucate and for 17 cultures from 5 clams from Bonne Anse, Charente Maritime (Table 3). Indeed, for these cultures, profiles similar to P. chesapeaki were obtained after RsaI digestion (around 248 bp, 195 bp, 157 bp, 74 bp) (Fig. 2).

Fig. 2. Restriction profiles obtained after RsaI digestion of Perk ITS750-85 PCR products from clams collected in 2005 in Leucate Lagoon. Lanes 2–7, 10 and 11 show Perkinsus olseni-P. mediterraneus restriction profiles (around 413 bp, 193 bp and 74 bp) while lanes 8 and 9 show P. chesapeaki restriction profiles (around 248 bp, 195 bp, 157 bp, and 74 bp). Lanes 1 and 12 show a 100-bp ladder (Smartladder, Eurogentec).
Generally, when several cultures were tested for 1 clam, RsaI digestion yielded similar restriction profiles. However, for 3 clams collected in 2005 in Leucate Lagoon, both P. olseni and P. chesapeaki RFLP profiles were obtained from replicate cultures propagated from individual clams (Table 4).
Table 4. Results obtained for six clams Ruditapes decussatus collected in Leucate Lagoon in 2005

Genetic variability of the parasite
Twenty-four Perk ITS 750–85 PCR products were cloned and up to 27 clones per sample were tested again using PCR-RFLP (Table 3). Generally, restriction profiles obtained on clones were concordant with restriction profiles observed after direct PCR-RFLP. However, in some cases, restriction profiles were different from the profiles described by Abollo et al. (Reference Abollo, Casas, Ceschia and Villalba2006) (noted as being ambiguous in Table 3) or corresponded to P. olseni restriction profiles whereas the other clones tested for this culture showed P. chesapeaki profiles.
These clones (which have given ambiguous or unexpected restriction profiles) were selected for sequencing as well as up to 3 clones pr culture which have yielded concordant PCR-RFLP results before and after cloning.
In total, 28 sequences were obtained for clones showing Perkinsus olseni restriction profiles as well as ambiguous ones and displayed between 99 and 100% identity with P. olseni (U07697). The genetic distance within these sequences was low (0·4%). These sequences showed between 1 and 3 point nucleotide modifications (mainly substitutions) and concerned 1 clone. However, 4 substitutions could be noticed in more than 1 clone. These substitutions were located in the ITS-2: A instead of G in positions 40 and 54, G instead of A in position 158 and C instead of T in position 225. Five types of sequence (POa; POb; POc; POd and POe) were finally obtained and deposited in GenBank under Accession numbers JQ669641–JQ669645. No correlation could be observed between sequences and individual or geographical origin.
In total, 21 sequences were obtained for clones showing Perkinsus chesapeaki restriction profiles and displayed 96–97% identity with P. chesapeaki (AF091541). The overall mean distance between these sequences was 1·6%. Some sequences showed point nucleotide substitutions. However, some modifications could be observed in more than one clone. One substitution (T instead of A) could be noticed in the ITS-1 (80 bp before the end of the fragment) in 3 clones obtained from clams collected in Charente Maritime (Bonne Anse). In positions 143–145 of the ITS-2, 13 sequences showed an insertion of ATA: 8 of those were obtained from clams collected in Charente Maritime, and 4 were from Leucate clams. Finally, 4 types of sequence (PCa; PCb; PCc and PCd) were obtained and deposited in GenBank under Accession numbers JQ669646–JQ669649. Similarly to P. olseni, no correlation could be observed between sequences and individual or geographical origin.
Phylogenetic analyses of Perkinsus olseni and P. chesapeaki strains isolated during this study
ITS region sequence analysis
To determine the taxonomic affiliation of Perkinsus species reported in the present study with other congeneric species, phylogenetic analysis was first performed on the rRNA-ITS region.
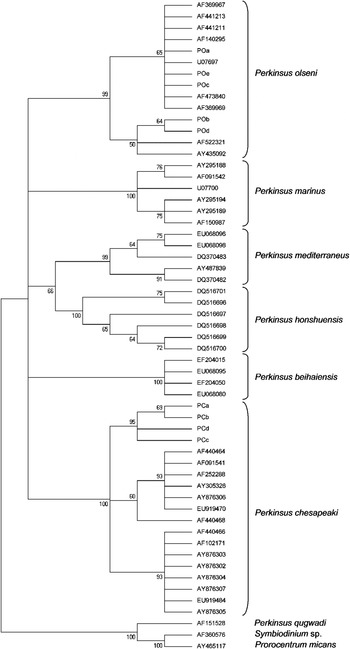
The different sequence types (9) obtained on the rRNA-ITS region were included in a maximum parsimony (MP) analysis (Fig. 3). As expected, the 5 sequences showing maximum identity with Perkinsus olseni were in the P. olseni clade and the 4 sequences showing maximum identity with P. chesapeaki were in the P. chesapeaki group. However, the MP analysis revealed with 95% bootstrap support the existence of a subclade including only French strains within the P. chesapeaki clade (Fig. 3).

Fig. 3. Maximum Parsimony analysis showing the taxonomic position of Perkinsus ITS sequences obtained in the present study. Numbers at branch nodes indicate bootstrap confidence values as a percentage. The analysis included 57 nucleotide sequences and was conducted using 841 aligned nucleotide positions. Sequences (Accession numbers) obtained in this study were : POa (JQ669641); POb (JQ669642); POc (JQ669643); POd (JQ669644); POe (JQ669645); PCa (JQ669646); PCb (JQ669647); PCc (JQ669648) and PCd (JQ669649).
LSU and actin-1 sequence analyses. In order to complete the data obtained on the rRNA-ITS region, 1 culture of the Perkinsus olseni clade and 1 culture from the P. chesapeaki clade were selected for amplification using primers targeting the actin-1 gene region and the LSU region.
Both cultures were successfully amplified using PerkActin1-439R and PerkActin1130F. After cloning, three 330 pb-PCR products were sequenced. Two of them were identical and showed maximum identity (98%) with P. chesapeaki actin-1 (AY876361). The GenBank Accession number for this sequence is JQ669650. The last PCR product showed 100% identity with P. olseni (AY876352).
Both cultures were amplified using LSU A and LSU B primers. PCR products were cloned and five 970 bp-clones were sequenced. Three clones showed maximum identity (99%) with Perkinsus olseni LSU (AY876332) whereas the 2 others showed maximum identity (99%) with P. chesapeaki (AY876347). These 5 sequences were deposited in GenBank under the following Accession numbers: JQ669651–JQ669655.
Phylogenetic analyses performed on the LSU and actin-1 genes supported results obtained on the ITS region: some cultures showing maximum identity on the ITS region with P. olseni and P. chesapeaki grouped with the same conspecific strains (Figs 4 and 5).

Fig. 4. Maximum Parsimony analysis showing the taxonomic position of Perkinsus Actin 1 gene sequences obtained in the present study. Numbers at branch nodes indicate bootstrap confidence values as a percentage. The analysis included 28 amino acid sequences and was conducted using 88 aligned amino acid positions. (*) Sequences obtained in this study.

Fig. 5. Maximum Parsimony analysis showing the taxonomic position of Perkinsus large subunit (LSU) rRNA gene sequences obtained in the present study. Numbers at branch nodes indicate bootstrap confidence values as a percentage. The analysis included 32 nucleotide sequences and was conducted using 984 aligned nucleotide positions. (*) Sequences obtained in this study.
Description of in vitro stages of Perkinsus olseni and P. chesapeaki strains isolated from Leucate Lagoon
Cells from 4 clonal cultures from 2 clams characterized as Perkinsus olseni and cells from 3 clonal cultures from 1 clam characterized as P. chesapeaki were observed and measured after 16 days of cultures.
Cultures of both species were propagated in the DME/Ham's F-12 culture medium, where they proliferated predominantly by schizogony, and to a lesser extent by zoosporulation. Trophozoïtes from both species showed a typical signet ring morphology with a large vacuole and eccentric nucleus with a prominent nucleolus (Fig. 6A and B) and presented comparable mean cell diameter (Table 5). However, P. chesapeaki trophozoites could enlarge up to 41 μm while those of P. olseni showed a maximum cell diameter of 27 μm.

Fig. 6. (A–C) In vitro Perkinsus olseni (A) and P. chesapeaki (B–C) cultures obtained from Ruditapes decussatus clams (Leucate Lagoon). (A) Trophozoïte (T) showing a typical signet-ring morphology with a large vacuole and eccentric nucleus, schizont (S) containing numerous daughter cells, cluster (C) of sibling daughter trophozoites. (B) Trophozoïte (T) showing a typical signet-ring morphology with a large vacuole and eccentric nucleus, zoosporangium with probable discharge tube (*). (C) Zoosporangium with extended discharge tube (*) enclosing hundreds of zoospores. Scale bars = 50 μm.
Table 5. Mean diameters, diameter ranges and proportional abundances of the different parasite stages observed for the in vitro culture of Perkinsus olseni and P. chesapeaki

Perkinsus olseni showed more but smaller schizonts than P. chesapeaki (Table 5, Fig. 6A). Finally, only 3 zoosporangia (0·2%) could be observed for P. olseni while at the same time, P. chesapeaki presented about 18% of zoosporangia (Table 5, Fig. 6B and C).
Histology and in situ hybridization
Sixty Ruditapes decussatus clams collected during 2008 from Leucate Lagoon were analysed histologically and by ISH assays with the Perkinsus chesapeaki probe. Twenty of the same clams were also analysed by ISH assays with the P. olseni probe.
Fifty-one clams were found by histology to be infected and showed mature trophozoïtes in connective tissue of different organs including gills, mantle, digestive gland, gonad, muscle, heart, kidney and palps. Mature trophozoïtes were characterized by an eccentric vacuole and a signet ring (Fig. 7). Several trophozoïtes were observed inside haemocytes or encapsulated in an eosinophylic acellular matrix (Figs 7 and 8) and haemocyte infiltration could be observed closed to the trophozoïtes (Fig. 9).

Fig. 7. H&E-stained tissue section showing Perkinsus trophozoïtes in tissues of Ruditapes decussatus from Leucate Lagoon. Cluster of mature trophozoïtes inside an eosinophilic acellular matrix (*) showing an eccentric vacuole and a signet ring located in the connective tissue of the mantle.

Fig. 8. H&E-stained tissue section showing Perkinsus trophozoïtes in tissues of Ruditapes decussatus from Leucate Lagoon. Isolated or grouped trophozoïtes located in the connective tissue of the mantle. Parasite clusters are associated with eosinophilic acellular matrix (*).

Fig. 9. H&E-stained tissue section showing Perkinsus trophozoïtes in tissues of Ruditapes decussatus from Leucate Lagoon. Isolated and grouped trophozoïtes associated with an important infiltration of haemocytes in the connective tissue of the digestive gland.
Among 60 tested clams, 83% (50/60) showed specific labelling of parasite cells with the P. chesapeaki probe. All (20/20 = 100%) tested clams showed labelling of parasite cells by the P. olseni probe. Ten clams (10/20 = 50%) that showed dual infections through labelling of parasite cells by one of both probes, were selected for comparisons of in vivo tissue tropisms, relative abundances, and morphologies among cells of the co-infecting pathogen species.
Connective tissues of all the organs appeared infected with both probes. Gills, mantle and then digestive glands were more often found to be infected than kidney palps and heart. Gonad and muscle appeared positive for both assays in one clam. Generally in clams with dual infections, P. olseni cells were more abundant and widely distributed than P. chesapeaki cells. Indeed, P. olseni was observed in more organs than P. chesapeaki. Moreover, 1·3 up to 17 times more P. olseni were counted than P. chesapeaki in co-infected clams except in 1 clam for which this last was twice more abundant than P. olseni.
Both Perkinsus species could appear as isolated trophozoïtes or in clusters (Figs 10A and B and 11A and B). Haemocytic infiltration was observed more frequently associated with P. olseni cells (Fig. 10A) than with P. chesapeaki cells. In histological material, no consistent size differences were detected between cells of the two parasite species (9·1 ± 2·8 μm, n=161 for P. olseni labelled cells; 9·8 ± 2·9 μm, n = 58 for P. chesapeaki labelled cells).

Fig. 10. (A and B) In situ hybridization assay using Perkinsus olseni probe on a clam Ruditapes decussatus from Leucate Lagoon. (A) Positive isolated and grouped trophozoïtes in the connective tissue of gills. The presence of the parasites is associated with a strong haemocytic infiltration. (B) Positive isolated and grouped trophozoïtes in the connective tissue of the digestive gland.

Fig. 11. (A and B) In situ hybridization assay using Perkinsus chesapeaki probe on a clam Ruditapes decussatus from Leucate Lagoon. (A) Encapsulated positive trophozoïtes in the connective tissue of the digestive gland. (B) Positive isolated and grouped trophozoïtes in the gonadal connective tissue.
DISCUSSION
Parasites of the genus Perkinsus have been associated with mortalities of molluscs around the world, including oysters, clams, abalones and scallops (Perkins, Reference Perkins1996; Villalba et al. 2004). Among these parasites, Perkinsus olseni affects the clams Ruditapes decussatus and R. philippinarum (Casas et al. Reference Casas, Villalba and Reece2002), 2 bivalves with important commercial value in Europe where mortalities associated with this parasite have been reported (Figueras et al. Reference Figueras, Robledo and Novoa1992, Reference Figueras, Robledo and Novoa1996). Although parasites of the genus Perkinsus are known to infect clams in France (Goggin, Reference Goggin1992; Garcia et al. Reference Garcia, Arzul, Chollet, François, Goubet, Joly, Miossec and Robert2006; Lassalle et al. Reference Lassalle, de Montaudouin, Soudant and Paillard2007; Dang et al. Reference Dang, de Montaudouin, Caill-Milly and Željka2010), no molecular characterization was previously performed on these parasites.
The main objective of the current study was to characterize at the species level, Perkinsus sp. parasites that were detected by genus-specific histological or RFTM assays. For that purpose, parasite cultures obtained from infected clams collected in various French locations were screened by PCR-RFLP according to Abollo et al. (Reference Abollo, Casas, Ceschia and Villalba2006). In order to be able to detect potential co-infection (infection with several species or strains), PCR products were cloned and several clones were screened again by PCR-RFLP. This work obtained P. olseni-like restriction profiles in all the investigated areas and also P. chesapeaki like restriction profiles in 2 locations: Leucate Lagoon in southern France and Bonne Anse in Charente Maritime on the middle Atlantic coast of France. In these locations, replicate cultures from individual clams presented different PCR-RFLP profiles; indicating co-infections by multiple pathogen species. Some clones obtained from clams collected in different places and displaying P. olseni and P. chesapeaki or ambiguous restriction profiles were selected for sequencing. Phylogenetical analyses confirmed RFLP results and showed that parasites of the genus Perkinsus present in France belong either to P. olseni or P. chesapeaki clades. However, based on the ITS region parsimony analysis, French P. chesapeaki strains appeared slightly different from other conspecific strains and grouped together (95% bootstrap support). Genetic distances within our isolate strains were low: 0·6% and 1·6% for P. olseni and P. chesapeaki sequences respectively. These values are in the range of the intraspecific variations observed within the currently accepted Perkinsus species (Moss et al. Reference Moss, Xiao, Dungan and Reece2008). During the present study, 5 and 4 different ITS sequences were obtained for P. olseni and P. chesapeaki respectively. In a previous study, Reece et al. (Reference Reece, Bushek, Hudson and Graves2001) showed that allelic and genotypic frequencies differed significantly among Perkinsus marinus strains isolated from 3 regions of the USA and those genotypic differences could be related to differences in virulence. Based on the analysis of the NTS domain of P. marinus, sequence type frequencies varied according to the geographical origin of the samples (Robledo et al. Reference Robledo, Wright, Marsh and Vasta1999). In a study on P. olseni cultures, different levels of microsatellite polymorphism varied with the geographical origin of the parasites (Vilas et al. Reference Vilas, Cao, Pardo, Fernandez, Villalba and Martinez2011). More specifically, parasites from Japan and New Zealand appeared much more variable than those from Spain (Vilas et al. Reference Vilas, Cao, Pardo, Fernandez, Villalba and Martinez2011). In our study, we could not observe any relationship between sequence type and geographical origin. This lack of correlation could partly be explained by the low number of sequences to support regional comparison.
The detection of several sequence types (up to 3 different ones) in the same clam can reflect the presence of different P. olseni and P. chesapeaki strains in the same individual clam as has been reported for P. marinus (Reece et al. Reference Reece, Bushek and Graves1997, Reference Reece, Bushek, Hudson and Graves2001) and for P. olseni (Vilas et al. Reference Vilas, Cao, Pardo, Fernandez, Villalba and Martinez2011). It can also reflect variability in the rRNA locus or between the rRNA repeats in a same Perkinsus sp. strain. Indeed, previous studies suggested that in vitro P. marinus cells are diploids (Reece et al. Reference Reece, Bushek and Graves1997) and that distinct rRNA units exist in P. andrewsi – newly called P. chesapeaki (Pecher et al. Reference Pecher, Robledo and Vasta2004).
In order to confirm and support results obtained on the ITS region, 1 Perkinsus olseni and 1 P. chesapeaki culture were selected for further molecular characterization on the LSU and actin 1 genes. Maximum Parsimony analyses performed on both regions confirmed our previous results and allowed us to conclude that P. olseni and P. chesapeaki are present in France.
The propagation of monoclonal cultures of Perkinsus olseni and P. chesapeaki produced all parasite stages previously described for these parasite species. Mature trophozoïtes of both isolates showed vacuolated, signet-ring morphology with eccentric nuclei bearing prominent nucleoli. These trophozoïtes presented simultaneous schizogonic and zoosporogonic proliferative cycles. However, in our conditions, zoosporulation was less frequent in P. olseni compared to P. chesapeaki. Schizonts and zoosporangia of P. chesapeaki appeared bigger than those of P. olseni. Such differences can also be deduced from data available in the literature (Casas et al. Reference Casas, Villalba and Reece2002; Burreson et al. Reference Burreson, Reece and Dungan2005; Dungan and Reece, Reference Dungan and Reece2006). Compared to these previous observations, P. olseni isolated during this study looks very similar in size to other P. olseni isolates (Casas et al. Reference Casas, Villalba and Reece2002; Dungan and Reece, Reference Dungan and Reece2006); while the P. chesapeaki isolates of the current investigation showed larger schizonts and smaller zoosporangia than those reported for North American isolates (Coss et al. Reference Coss, Robledo and Vasta2001a; Dungan et al. Reference Dungan, Hamilton, Hudson, McCollough and Reece2002; Burreson et al. Reference Burreson, Reece and Dungan2005).
Histological examination of infected R. decussatus clams from Leucate Lagoon revealed the presence of spherical trophozoïtes in the connective tissue of many different organs, occasionally isolated but more often in clusters. Haemocytic infiltration was sometimes observed around infected zones. Parasites could be observed inside haemocytes or encapsulated. ISH allowed distinction of P. chesapeaki and P. olseni in the same infected clams. Generally P. olseni appeared more spread out and more abundant. These observations were supported by molecular works. Indeed, direct PCR-RFLP allowed detection of P. olseni more frequently than P. chesapeaki in Leucate Lagoon.
Infections of different mollusc hosts at the same site by different Perkinsus species and strains have been previously reported (Reece et al. Reference Reece, Bushek and Graves1997; Dungan and Reece, Reference Dungan and Reece2006; Reece et al. Reference Reece, Dungan and Burreson2008). More specifically, a molecular epizootiology study revealed the presence of P. marinus and, to a lesser extent, P. chesapeaki in oyster samples from several sites located in Chesapeake Bay (Reece et al. Reference Reece, Dungan and Burreson2008). In our study, P. olseni and P. chesapeaki appeared sympatric in 2 distinct locations: in the South of France along the Mediterranean sea (Leucate) and in Charente Maritime on the middle West coast of France (Bonne Anse). In the first case P. olseni appeared to be more abundant while in Bonne Anse, PCR-RFLP results suggested that P. chesapeaki was more abundant than P. olseni. These apparent abundance differences could be due to the difference of host species. Indeed, during this study, clams collected from Leucate were Ruditapes decussatus whereas clams collected from Charente Maritime (Bonne Anse) were R. philippinarum. The apparent abundance difference could also reflect competition between both parasite species. However, the distribution of these parasite species among the organs appeared identical which does not support the hypothesis of interspecific competition.
The respective impact of both these parasite species on clams is difficult to assess from our results. No mortality and no population decline were reported in the investigated locations during this study and through the French mollusc health surveillance network (REPAMO). However, considering that mortality has already been reported in association with both parasite species, their prevalence and infection intensity in French clam populations should be monitored.
Although the presence of Perkinsus olseni was suspected in France because of the presence of this parasite in other closed European country (Casas et al. Reference Casas, Villalba and Reece2002; Abollo et al. Reference Abollo, Casas, Ceschia and Villalba2006; Elandaloussi et al. Reference Elandaloussi, Carrasco, Furones and Roque2009), the detection of P. chesapeaki was more surprising. Indeed, until now this parasite has only been reported in North America and more especially in the soft-shell clam Mya arenaria in Chesapeake Bay (McLaughlin and Faisal, Reference McLaughlin and Faisal2000; McLaughlin et al. Reference McLaughin, Tall, Shaheen, Elsayed and Faisal2000; Dungan et al. Reference Dungan, Hamilton, Hudson, McCollough and Reece2002) and the razor clam Tagelus plebeius in Chesapeake and Delaware bays (Dungan et al. Reference Dungan, Hamilton, Hudson, McCollough and Reece2002; Bushek et al. Reference Bushek, Landau and Scarpa2008). The synonymization by Burreson et al. (Reference Burreson, Reece and Dungan2005) of P. chesapeaki and P. andrewsi previously characterized in Macoma balthica from Rhode River, Maryland (Coss et al. Reference Coss, Robledo, Ruiz and Vasta2001b) has consequently enlarged the geographical and host ranges of this parasite. Other clam hosts for P. chesapeaki include: Macoma mitchelli, Mercenaria mercenaria, Mulinia lateralis, Rangia cuneata and Cyrtopleura costata (Burreson et al. Reference Burreson, Reece and Dungan2005; Reece et al. Reference Reece, Dungan and Burreson2008). This is the first description of P. chesapeaki in Ruditapes decussatus and R. philippinarum in Europe. Its introduction might have occurred through introduction of susceptible species including Mya arenaria or Mercenaria mercenaria from North America. Mercenaria mercenaria was first introduced to the Atlantic coast of France in 1861 and then to the Mediterranean coast in 1965 (Lambert, Reference Lambert1947–1949; Ruckebusch, Reference Ruckebusch1947–1949; Bascheri, Reference Bascheri1965; Goulletquer et al. Reference Goulletquer, Bachelet, Sauriau, Noel, Leppäkkosski, Gollasch and Olenin2002). In Europe Mya arenaria occurs widely; but it is believed to have been extinct in Pleistocene times and re-introduced by man in historical times (Petersen et al. Reference Petersen, Rasmussen, Heinemeier and Rud1992).
The work presented herein would require testing the presence of P. chesapeaki in other bivalve species including Mya arenaria or Mercenaria mercenaria and in other European locations, in particular areas where clams were introduced from North America. Apparent co-habitation of P. olseni and P. chesapeaki in 2 distinct locations in France also raises interesting questions regarding relationships between these two parasite species as well as parasite-host interactions.
ACKNOWLEDGEMENTS
We gratefully acknowledge Dr K. Reece for her advice before starting this investigation. Our esteemed colleague Dr R. Carnegie generously provided positive control for in situ hybridization (Mya arenaria infected with Perkinsus chesapeaki). Our special thanks to referees and Dr A. Travers for their critical review of this manuscript. We would also like to thank Y. Pichot, P. Le Gall, A. Langlade, F. D'Amico, G. Trut, J.-C. Piquet very much for their participation or their help in facilitating clam collecting.