



INTRODUCTION
Chalinidae Gray, Reference Gray1867 is the largest family within the Haplosclerida Topsent, Reference Topsent1928. It is also the most complicated taxonomically due to the paucity, simplicity and high variability of its taxonomic characters (De Weerdt, Reference De Weerdt1989, Reference De Weerdt2000). Haliclona is the richest genus in the family and an unusually species-rich genus in the Porifera, with more than 400 species listed in the World Porifera Database (Van Soest et al., Reference Van Soest, Boury-Esnault, Hooper, Rützler, de Voogd, Alvarez de Glasby, Hajdu, Pisera, Manconi, Schoenberg, Janussen, Tabachnick, Klautau, Picton, Kelly, Vacelet, Dohrmann and Díaz2013). Many of these species were described for other genera, later synonymized with Haliclona. In spite of the six subgenera currently used for taxonomic housekeeping in Haliclona, over 200 species remain unassigned to a subgenus, which reflects the exceedingly poor knowledge on the morphology of these sponges, and calls for urgent redescription of historical materials from all around the world. Ideally, these reassessments will be conducted side by side with taxonomic revisions of regional faunas, undertaken on the basis of large series of freshly collected materials. This is the rationale underpinning our efforts for an improved knowledge about Brazilian Haliclona spp. In this paper, we describe two new species of Haliclona, collected at Pernambuco, Alagoas and Bahia States, between 08°46′S and 13°56′S.
MATERIALS AND METHODS
Specimens were collected by snorkelling or SCUBA diving in several localities indicated in Figure 1. The preparation of spicules for light and electron microscopy (SEM), and thick sections were made following the method of Hajdu et al. (Reference Hajdu, Peixinho and Fernandez2011). The SEM microscope used was a JEOL 6390 LV of the Departamento de Invertebrados of Museu Nacional/UFRJ. Spicule dimensions are based on measurements of 30 fully grown spicules for each specimen, and presented as minimum(mi.)–mean(me.)–maximum(ma.) length/mi.–me.–ma. width. To avoid confusion of Brazilian geopolitical regions (north-eastern) and marine ecoregions (Eastern and North-eastern), whenever referring to the latter, the word ecoregion is employed.

Fig. 1. Collection sites of the two new species. ■, Ponta dos Carneiros, Tamandaré, PE; ![]() , Ponta do Prego, Maceió, AL; ♦, Praia do Francês, Marechal Deodoro, AL; ★, Ponta de Montserrat, Salvador, BA; •, Taipús de Fora, Maraú, BA. Abbreviations of the States are as follow: AL, Alagoas; BA, Bahia; PE, Pernambuco.
, Ponta do Prego, Maceió, AL; ♦, Praia do Francês, Marechal Deodoro, AL; ★, Ponta de Montserrat, Salvador, BA; •, Taipús de Fora, Maraú, BA. Abbreviations of the States are as follow: AL, Alagoas; BA, Bahia; PE, Pernambuco.
Holotype and paratypes of Haliclona (Reniera) chlorilla sp. nov. are deposited in the Museu Nacional, Universidade Federal do Rio de Janeiro Porifera collection (MNRJ). Holotype and two paratypes of Haliclona (Soestella) peixinhoae sp. nov. are deposited in the MNRJ Porifera collection. The three remaining paratypes of this species are deposited in the Porifera collection of Universidade Federal da Bahia (UFBA-POR). Fragments of types originating from Alagoas State are deposited in the Porifera collection of Universidade Federal de Alagoas (UFALPOR).
Classification follows the Chalinidae chapter in the Systema Porifera (De Weerdt, Reference De Weerdt, Hooper and Van Soest2002)
SYSTEMATICS
Class DEMOSPONGIAE Sollas, Reference Sollas1885
Order HAPLOSCLERIDA Topsent, Reference Topsent1928
Family CHALINIDAE Gray, Reference Gray1867
Genus Haliclona Grant, Reference Grant and Todd1836
Subgenus Reniera Schmidt, Reference Schmidt1862
Haliclona (Reniera) chlorilla sp. nov.
(Figures 2, 3; Tables 1, 2)
Fig. 2. Morphological variability of Haliclona (Reniera) chlorilla sp. nov. in situ: (A) black specimen growing on calcareous algae (MNRJ 17026, holotype); (B) dark green specimen growing together with macroalgae and covered by fine sediment; (C) black specimen with tendency to form erect branches in a reef crevice; (D) dark green specimen growing together with macroalgae in close up (MNRJ 17027, paratype). Scales bars: A–C, 5 cm; D, 2 cm.
Fig. 3. Haliclona (Reniera) chlorilla sp. nov.: (A) ectosomal skeleton; (B) choanosomal skeleton; (C) detail of the choanosomal skeleton; (D) oxeas. Scale bars: A, C, 400 µm; B, 500 µm; D, 20 µm.
Table 1. Spicule dimensions of Haliclona (Reniera) chlorilla sp. nov. Values are as follow: minimum–mean–maximum length × mi.–me.–ma. width. N = 30.
Table 2. Comparative morphological and distributional data for the shallow-water species of Haliclona known from the tropical western Atlantic. Halicl., Haliclona; Re., Reniera; S., Soestella; Halich., Halichoclona; Rh., Rhizoniera. Scale bars = 50 µm.
TYPE MATERIAL
Holotype: MNRJ 17026—Praia do Francês (sandstone reefs, ~09°45′58.35″S 35°50′10.62″W), Marechal Deodoro (AL, Brazil), ~1 m depth, coll. M.D. Correia, 16 June 2011. Fragment deposited under UFALPOR 0718.
Paratypes: MNRJ 1528—Praia de Ponta dos Carneiros (seagrass, ~08°41′46.14″S 35°04′42.59″W), Tamandaré (PE, Brazil), ~1 m depth, coll. E. Hajdu & G. Muricy, 1998; MNRJ 17027—Ponta do Prego (tide pool on coral reef, ~09°31′48″S 35°35′30″W), Maceió (AL, Brazil), 1 m depth, coll. A. Bispo, 11 January 2012. Fragment deposited under UFALPOR 0773; MNRJ 17610—Ponta Verde (tide pool on coral reef, ~09°39′58″S 35°41′38″W), Maceió (AL, Brazil), ~0.5 m depth, coll. M.D. Correia, 6 February 2008. Fragment deposited under UFALPOR 0454.
DIAGNOSIS
Haliclona (Re.) chlorilla sp. nov. is set apart from other tropical western Atlantic (TWA) Haliclona spp. by the combination of delicate, anastomosing branches, dark green or black live colour, and absence of any microscleres.
DESCRIPTION
The holotype was growing on calcareous macroalgae (Jania adhaerens) and has a more compact morphology than observed in other specimens (Figure 2A). The species is most frequently repent (Figure 2A–D), consisting of slender and delicate, cylindrical, anastomosing branches, 2–5 mm in diameter, with a smooth surface. These branches could grow erect in specimens found in more sheltered habitats, such as crevices in sandstone reefs (Figure 2C). Oscula are numerous, circular, flush with the surface or just slightly elevated, usually aligned in rows along the branches, 0.4–1 mm in diameter. Consistency is very delicate, soft and fragile. Colour alive is dark green or black, becoming beige in ethanol.
Skeleton
The ectosomal skeleton is a tangential, isotropic and unispicular reticulation forming three-, four-sided or polygonal meshes (Figure 3A). The choanosomal skeleton is an isotropic and unispicular reticulation, forming three-, four-sided or polygonal meshes, around an off-central lumen running longitudinally inside the branches (Figure 3B, C). Spongin is scarce, present at the nodes of the reticulation, as usual in the subgenus.
ECOLOGY
Found in very shallow waters (~0.1–2 m) and in tide pools, growing together with macroalgae or seagrass, mostly in well lit microhabitats.
DISTRIBUTION
The type locality is Praia do Francês (Marechal Deodoro, AL, Brazil), at the southern limit of the species' known geographical distribution, which extends northwards to the southern coast of Pernambuco State. This far the species is not yet known outside the North-eastern Brazil ecoregion.
ETYMOLOGY
The specific epithet is used as a noun in apposition. It is derived from the juxtaposition of two Greek words, chloros (green) and illas (rope), and conveys the dark-green, slender ramose habit of the new species.
REMARKS
Table 2 compiles literature data on Haliclona spp. used in comparison with the new species. Haliclona (Reniera) chlorilla sp. nov. is distinguished within the subgenus by the combination of its repent delicate cylindrical branches of dark green or black colour, and absence of microscleres of any sort. In the TWA, such slender cylindrical branches are mostly seen as off-shoots from the main sponge body, as reported by De Weerdt (Reference De Weerdt2000) for H. (Re.) manglaris and H. (Re.) tubifera. The exception is H. (Re.) ruetzleri De Weerdt, Reference De Weerdt2000, reported to be entirely composed of slender, delicate, anastomosing branches, 1–3 mm in diameter. This species, albeit its rather similar habit, has light brown live colour, larger oxeas, as well as sigmas and toxas as microscleres, which renders it clearly distinct from the new species.
The only additional TWA Haliclona (Reniera) with a dark live colour (dark brown to black) is H. (Re.) strongylophora Lehnert & Van Soest, Reference Lehnert and Van Soest1996. This Jamaican species is cushion-shaped and has strongyles as megascleres, two markedly distinct features given what is seen in the new species. Furthermore, its single record was obtained at 77 m depth, while the new species is this far known only from 0.1–2 m. Other rather dark TWA Haliclona spp. fall within H. (Soestella), being notably distinct from the new species in terms of skeletal architecture. These are H. (S.) luciencis De Weerdt, Reference De Weerdt2000; H. (S.) melana Muricy & Ribeiro, Reference Muricy and Ribeiro1999 and H. (S.) walentinae Díaz et al., Reference Díaz, Thacker, Rützler, Piantoni, Custódio, Lôbo-Hajdu, Hajdu and Muricy2007.
Subgenus Soestella De Weerdt, Reference De Weerdt2000
Haliclona (Soestella) peixinhoae sp. nov.
(Figures 4, 5; Tables 2, 3)
Fig. 4. Morphological variability of Haliclona (Soestella) peixinhoae sp. nov. in situ.: (A) yellowish beige specimen with full-growth tubes and thorn-like projections (uncollected specimen from Ponta de Montserrat, Salvador, BA); (B) yellowish specimen with juxtaposed tubes growing with macroalgae (Taipús de Fora, Maraú, BA); (C, D) beige specimen with a repent-tubular shape (MNRJ 13299, holotype). Scale bars: A, 2 cm; B, 5 cm; C, 2 cm; D, 1 cm.
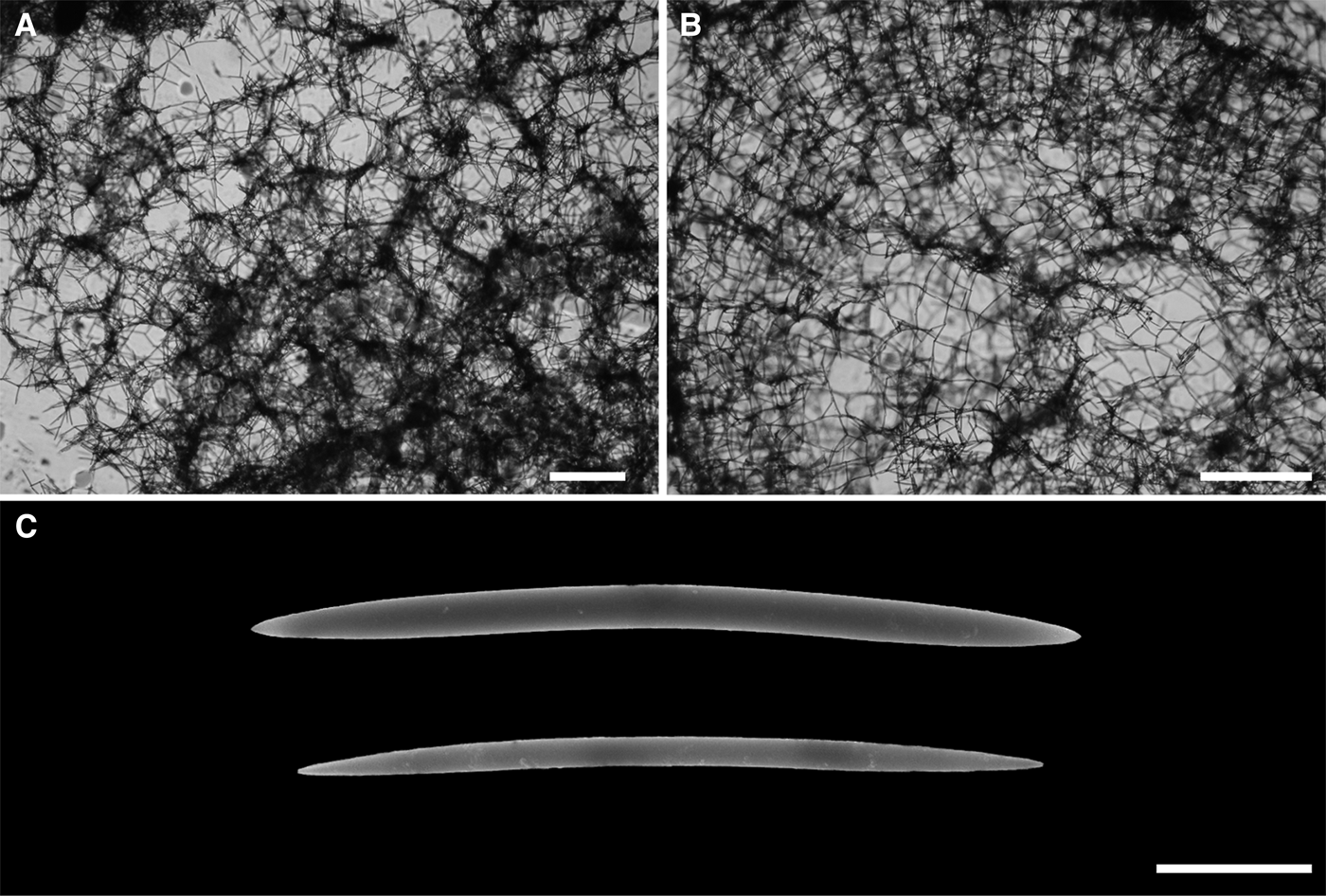
Fig. 5. Haliclona (Soestella) peixinhoae sp. nov.: (A) ectosomal skeleton; (B) choanosomal skeleton; (C) oxeas. Scale bars: A, B, 500 um; C, 20 um.
Table 3. Spicule dimensions of Haliclona (Soestella) peixinhoae sp. nov. Values are as follow: minimum–mean–maximum length × mi.–me.–ma. width. N = 30.

Haliclona sp.: Hajdu et al., Reference Hajdu, Peixinho and Fernandez2011: 188–190.
MATERIAL EXAMINED
Holotype: MNRJ 13299—Taipús de Fora (algal/coral reef, ~13°56′03.64″S 38°55′32.55″W), Maraú (BA, Brazil), 1 m depth, coll. E. Hajdu & G. Lôbo-Hajdu, 26 July 2009.
Paratypes: MNRJ 2472, 2563—Ponta de Montserrat (rocky coast, ~12°55′37″S 38°31′11″W), Salvador (BA, Brazil), 3–5 m depth, coll. E. Hajdu et al., 31 July 1999 and 5 August 1999, respectively; UFBA 1596–POR, UFBA 1602–POR, UFBA 1616–POR—Ponta de Montserrat (~12°55′37″S–38°31′11″W), Salvador (BA, Brazil), 4.5 m depth, coll. E. Hajdu, 14 January 1997.
DIAGNOSIS
Haliclona (Soestella) peixinhoae sp. nov. is set apart from other TWA Haliclona spp. by a combination of its habit comprising a dense aggregation of erect tubes, mostly bearing large thorns, albeit very soft, and a conspicuous subsuperficial reticulation visible to the naked eye on live specimens.
DESCRIPTION
Sponge forming dense aggregations of tubes up to 5 cm high (Figure 4), sometimes recalling a pan flute (partially fused tubes, Figure 4A–B). Specimens usually reach up to 15–25 cm in diameter. Tubes stand mostly erect, and appear to originate from a reptant tube (holotype). Tubes usually bear slender thorn-like projections up to 1 cm long (Figure 4A, C, D). Oscula are abundant, large (1–15 mm in diameter), usually circular and located apically on tubes. Surface smooth, but sometimes irregularly outlined due to development of early stage tubes or thorn-like projections. A conspicuous, irregular sub-superficial meandering reticulation is visible to the naked eye (Figure 4D), but disappears in ethanol. Consistency is very soft and fragile, easily torn. Colour alive is light brown, beige or yellowish beige, fading in alcohol.
Skeleton
The ectosome is a tangential, uni- to paucispicular reticulation with a tendency to form ill- or well-defined rounded meshes (Figure 5A). The choanosome consists of ill-defined, uni- to paucispicular primary lines, irregularly connected by uni- to paucispicular secondary ones, with a slight tendency to form rounded meshes (Figure 5B). A few of the secondary lines, if not artefactual, are two or three spicules long, forming an open and discontinuous reticulation with wide and irregular meshes.
ECOLOGY
A rare species, found in 1–5 m depth, in well-lit areas with low hydrodynamics. It frequently grows intermingled to bushy algae.
DISTRIBUTION
The type locality is Taipús de Fora (Maraú, BA, Brazil). This far the species is endemic from the upper half of the state of Bahia (Eastern Brazil ecoregion, north-eastern Brazil geopolitical region), known from Salvador and Maraú only.
ETYMOLOGY
The new species is named after the late Professor Dr Solange Peixinho, a dear friend, and an important advocate of the importance of taxonomy, and more specifically, taxonomy of Bahian sponges.
REMARKS
Hajdu et al. (Reference Hajdu, Peixinho and Fernandez2011) refered to the presence of rare toxas in Haliclona (Soestella) peixinhoae sp. nov. (as Haliclona sp.), but we carefully re-examined all the material available of this species and no toxa was found. The illustration given by those authors surely represents a contamination.
Haliclona (Soestella) peixinhoae sp. nov. is well distinguished among other Haliclona spp. in the TWA by its habit, comprising a dense aggregation of erect tubes with large apical oscula, mostly bearing large thorns, albeit very soft, and by a conspicuous subsuperficial reticulation visible to the naked eye on live specimens.
Tropical western Atlantic Haliclona (Soestella) spp. includes only H. (S.) melana and Haliclona (S.) smithae De Weerdt, Reference De Weerdt2000 as tubular sponges (Table 2). Both are rather distinct from the new species in that they do not form dense aggregates of erect tubes, do not possess the conspicuous sub-superficial reticulation, have black or yellow-green (respectively) live-colour, and possess microscleres (respectively, toxas and raphides). Other subgenera comprise a few additional tubular species in this area, even if optional. These are H. (Halichoclona) magnifica De Weerdt et al., Reference De Weerdt, Rützler and Smith1991; H. (Re.) mucifibrosa De Weerdt et al., Reference De Weerdt, Rützler and Smith1991; Haliclona (Re.) tubifera De Weerdt, Reference De Weerdt2000; and H. (Rhizoniera) curacaoensis (Van Soest, Reference Van Soest1980). Besides their skeletal architectures, these species differ from the new species in several traits. The first of these has a crispy consistency and much larger oxeas, reaching up to 220 µm, in contrast to the rather soft consistency and oxeas only up 115 µm. H. (Re.) mucifibrosa produces copious amounts of mucus when torn apart, a feature not observed in the new species, and also has much larger oxeas, up to 250 µm. Haliclona (Re.) tubifera, albeit its name, has only optional tubes. Specimens are frequently thin, ramose. Live colour is generally pink or pink/violet, and oxeas can reach over 170 µm. Finally, H. (Rh.) curacaoensis has oxeas only a bit larger (up to 146 µm) than those of the new species, but its bluish purple live colour adds to the skeletal architecture in setting it quite far apart from the new species.
DISCUSSION
Recent taxonomic effort dedicated to sponges of reef ecosystems in the north-east Brazilian geopolitical region has been rewarding in terms of new records and species found (Cedro et al., Reference Cedro, Hajdu, Sovierzosky, Correia, Custódio, Lôbo-Hajdu, Hajdu and Muricy2007, Reference Cedro, Hajdu and Correia2011, Reference Cedro, Hajdu and Correia2013; Peixinho et al., Reference Peixinho, Fernandez, Oliveira, Caires, Hajdu, Custódio, Lôbo-Hajdu, Hajdu and Muricy2007; Hajdu et al., Reference Hajdu, Peixinho and Fernandez2011; Barros et al., Reference Barros, Santos and Pinheiro2013; Sandes & Pinheiro, Reference Sandes and Pinheiro2013). The two new Haliclona spp. reported upon here highlight the fact that discoveries need not always rely on new fieldwork, as both were already present in scientific collections by the 1980s and 1990s.
Notwithstanding the 400+ extant species of Haliclona, only eleven have been registered for the Brazilian coast so far (Hajdu et al., Reference Hajdu, Peixinho and Fernandez2011; Muricy et al., Reference Muricy, Lopes, Hajdu, Carvalho, Moraes, Klautau, Menegola and Pinheiro2011; this study). Six are provisional endemics from various sectors of this coastline. Haliclona (Halicl.) catarinensis Mothes & Lerner, Reference Mothes and Lerner1994; H. (Halicl.) lilaceus Mothes & Lerner, Reference Mothes and Lerner1994; and H. (Rh.) mammillaris Mothes & Lerner, Reference Mothes and Lerner1994 are known only from their type localities in the South-eastern Brazil ecoregion (Bombinhas, ca 27°S). Haliclona (Halich.) lernerae Campos et al., Reference Campos, Mothes, Eckert and Van Soest2005 is known from a single record for the Amazonia ecoregion (off São Luís, ~0°S), and both new species described herein stem each from a distinct ecoregion, namely North-eastern Brazil with H. (Re.) chlorilla sp.nov., and Eastern Brazil with H. (S.) peixinhoae sp. nov. (cf. above).
We have adopted De Weerdt's (Reference De Weerdt2000) subgeneric assignment for H. (Rh.) mammillaris, as she had access to the holotype. Muricy et al.'s (Reference Muricy, Lopes, Hajdu, Carvalho, Moraes, Klautau, Menegola and Pinheiro2011) assignment of this species to Haliclona (Halicl.) is thus considered mistaken. On the other hand, these latter authors also assigned H. (Halicl.) catarinensis and H. (Halicl.) lilaceus to the nominotypical subgenus, which is followed here, despite types not being re-examined so far. From Mothes & Lerner's (Reference Mothes and Lerner1994) descriptions, all three Haliclona spp. would appear to have nearly the same architecture.
The remaining five Haliclona spp. known from Brazil also occur in the Tropical North-western Atlantic (Caribbean region): H. (Soestella) caerulea (Hechtel, Reference Hechtel1965); H. (Reniera) implexiformis (Hechtel, Reference Hechtel1965); H. (Re.) manglaris Alcolado, Reference Alcolado1984; H. (S.) melana Muricy & Ribeiro, Reference Muricy and Ribeiro1999; and H. (Re.) tubifera (George & Wilson, Reference George and Wilson1919) (sensu Lerner (Reference Lerner1996), = H. (Halich.) sp. sensu De Weerdt (Reference De Weerdt2000)). The record of H. (Rh.) curacaoensis by Cedro et al. (Reference Cedro, Hajdu, Sovierzosky, Correia, Custódio, Lôbo-Hajdu, Hajdu and Muricy2007; MNRJ 10280, UFALPOR 0241) has been briefly revised here and found to be best assigned to H. (Re.) manglaris. Its oxeas were found to reach only up to 85 µm in length, and its architecture, to be more on the Reniera side of the spectrum.
ACKNOWLEDGEMENTS
A.B. is thankful to Victor Cedro for help in fieldwork and discussions on the taxonomy of sponges, to the colleagues of the Laboratório de Taxonomia de Poríferos (TAXPO) of Museu Nacional/UFRJ for assistance during visits to the laboratory; to Professor Dr Hilda H. Sovierzoski and all other colleagues from Setor de Comunidades Bentônicas of the Universidade Federal de Alagoas (UFAL) for help in field trips. Professor Dr Cláudio Sampaio (UFAL at Penedo) is thanked for kindly allowing use of Figure 4B. MSc Júlio Cesar Cruz Fernandez is thanked for kindly providing Figure 5A–B. Two anonymous referees are thanked for providing valuable comments that improved the manuscript.
FINANCIAL SUPPORT
A.B., M.D.C. and E.H. are grateful to Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), and Programa de Pós-graduação em Diversidade Biológica e Conservação nos Trópicos (PPGDiBiCT/UFAL) for grants and/or scholarships.







