Introduction
Cowpea is a crop grown commonly in the dry savanna regions of sub-Saharan Africa (SSA). The crop is produced under rain-fed conditions in areas lying mainly between the northern limit of 15°N and 10°S. Compared with many other crops, cowpea thrives in places considered too dry for their production. Despite its drought tolerance, the productivity of cowpea could be adversely affected by erratic rainfall which occurs frequently in the dry savannas of SSA. In recent times, the pattern of rainfall in the sub-region, which either comes late, erratic at the beginning of the season or stops earlier than usual, requires that efforts be made to enhance the level of drought tolerance in currently available improved crop varieties. This is more so because crops are grown predominantly under rain-fed conditions in SSA. According to the World Development Report (2008), changes in temperature and rainfall will have dramatic effects on agricultural production in Africa. Only about 12.6 million hectares, i.e. 7% of the total agricultural land in Africa, are irrigated with 40% of this in North Africa.
The availability to and adoption by farmers of cowpea varieties with higher levels of drought tolerance should contribute positively to ensuring food security and improved human nutrition in the SSA region. Efforts made in the past to develop drought-tolerant cowpea varieties have met with limited success. This may have to do with the complexity of the factors at play in determining drought tolerance in the crop. Plant breeders need traits that can be readily used to discriminate between resistant/tolerant and susceptible plants in order for selection to be effective. The basis for drought tolerance in cowpea has not been well established, and this may have contributed to the limited progress so far recorded in increasing drought tolerance in cowpea varieties. In a detailed study on water-saving traits in cowpea, Belko et al. (Reference Belko, Zaman-Allah, Cisse, Diop, Zombre, Ehlers and Vadez2012) observed that lower early vigour, lower transpiration rate under well-watered conditions during the vegetative stage, lower leaf area development, sustained transpiration until the soil was relatively drier and lower canopy conductance under high vapour pressure deficit conditions appeared to be the main features discriminating drought tolerant from susceptible genotypes.
The present levels of drought tolerance in commonly grown cowpea varieties could be further enhanced through genetic improvement. It is conceivable that genes that could contribute to these higher levels of drought tolerance exist in some of the unexploited cowpea germplasm lines maintained in the genetic resources unit at the International Institute of Tropical Agriculture (IITA). In order to identify such lines that could be sources of genes for higher levels of drought tolerance, 1288 cowpea germplasm accessions were evaluated along with three breeding lines for tolerance to drought during the flowering stage in the field, and those selected were further tested in pots placed in the screenhouse to determine their tolerance to vegetative-stage drought. Six lines, of which all four plants recovered in the screenhouse evaluation, were chosen as parents for use in developing drought-tolerant improved breeding lines.
Materials and methods
One thousand two hundred and eighty eight cowpea germplasm lines (which had a sufficient number of seeds) obtained from the genetic resources centre at the IITA and three improved breeding lines were grown in the experimental field at the IITA, Ibadan during the dry season of 2007–2008. The three improved breeding lines were IT99K-494-6, IT99K-1122 and IT98K-506-1. The plots were ridged 1.0 m apart and each line was planted in single rows 1.0 m long. Spacing within a row was 20 cm and 1.0 m between the rows with three replications for each in the two water regimes. The cowpea lines were sown on December 4, 2007 in two blocks, with one block irrigated from planting to plant maturity while the second block was irrigated from planting for 5 weeks after which irrigation was stopped. There was no rainfall from mid November 2007 to February 2008, i.e. during the period when the study was conducted at Ibadan. Irrigation was carried out twice in a week using the sprinkler system. The plants were protected against insects by spraying ‘Rocket’, a broad-spectrum insecticide, with chlorpyrifos (Emulsifiable concentrate (EC) 20%) active ingredient at the rate of 2.0 litres/ha 3 and 6 weeks after sowing. Weed control was carried out manually, when necessary, using hoes. Data were collected on the number of days to the first flower opening, pod yield per plant, seed yield per plant and the number of days to 50% maturity (Supplementary Table S1, available online only at http://journals.cambridge.org). At plant maturity, all pods were harvested from the three middle plants in each row and weighed after drying. The pods were threshed and the seeds collected were weighed. Excel Microsoft software was used to summarize the data collected.
The 142 lines (Supplementary Table S2, available online only at http://journals.cambridge.org) identified as having enhanced drought tolerance in the field study were further evaluated in pots placed in the screenhouse. These were tested along with two early maturing lines (‘Sanzi’ and IT88DM-345), a drought-susceptible line (TVu 7778), two lines previously identified as drought tolerant (Danila and TVu 14 676) as well as an improved breeding line (IT99K-494-6) with good performance under drought. Each line was planted in four pots each filled with 5.0 kg of well-mixed top soil with one plant per pot. Each pot represented an experimental unit and was placed randomly on the screenhouse floor. The pots were watered for 14 d from seedling emergence, i.e. 4 d after sowing, and watering was then suspended for 28 d to impose drought stress. Watering was resumed on the 29th day after drought was imposed when most of the plants showed yellowing and browning of leaves (Supplementary Fig. S1, available online only at http://journals.cambridge.org). By the time watering was resumed, some plants recovered while many did not.
Results
Drooping of leaves, especially from around noon, which is the evidence of water stress on plants, was first observed among some cowpea lines sown in the water-stressed plots from about 20 d after suspension of irrigation. The number of lines showing wilting in the afternoon, however, increased with time. About 5 weeks after suspension of irrigation, only a few of the cowpea lines in the water-stressed plots showed no wilting. In some of the lines more sensitive to water stress, yellowing of lower leaves started within 4 weeks of exposure to drought. We also observed that in most of the lines in the water-stressed plots, the leaves became paraheliotropic, i.e. orientated in a position that was parallel to the sun's rays.
Anthesis occurred from about 37 d after sowing in the early maturing lines (Fig. 1). This early flowering habit is expected at this time of year which is characterized by short day lengths. However, when all of the germplasm lines tested are considered, the mean number of days to first flowering was about 12 d earlier when they were water stressed as compared with when not stressed. The induction of early flowering by water stress was more pronounced in the medium and late flowering lines compared with the early flowering ones (Fig. 2). Early flowering refers to those lines in which flowering occurred between 37 and 45 d, while the medium are those lines in which flowering occurred between 46 and 55 d after sowing. The lines that flowered in more than 55 d after sowing are considered as late in this study. When the plants were fully irrigated, flowering occurred in the latest line at 78 d after sowing. This same cowpea line flowered in 55 d after sowing in the water-stressed plot. Flowering occurred 59 d after sowing in the cowpea line that flowered late under water stress. Some of the cowpea lines tested including TVu294, TVu801, TVu1452, TVu5415, TVu10441, TVu14346, TVu14434 and TVu15913 flowered late and produced no pods in the water-stressed plot. These plants, however, remained green and retained some leaves until the study was terminated in March 2008. They exhibited a delayed-leaf-senescence (DLS) characteristic. When the raining season started from March, these plants flowered but the pods were not harvested since the plants were no longer protected against insect pests and no weeding was carried out due to the termination of the field study.

Fig. 1 Frequency distribution of the number of days to flower by germplasm lines under drought stress and non-stressed conditions. (A colour version of this figure can be found online at http://www.journals.cambridge.org/pgr)

Fig. 2 Relationship between the number of days to flower in cowpea germplasm lines under stress and non-stress conditions. (A colour version of this figure can be found online at http://www.journals.cambridge.org/pgr)
The number of days to 50% maturity ranged from about 60 to 100 d after sowing in the non-stressed plot (Fig. 3). The number of days to 50% maturity for the germplasm lines sown in the non-stressed plot followed a fairly normal distribution with the mean ranging between 75 and 83 d after sowing. Drought stress resulted in many lines attaining early maturity compared with those grown in the non-stressed plot. The number of days to 50% maturity of the test lines ranged from about 60 to 83 d after sowing in the drought-stressed plot, with over 90% of them maturing within 70 d after sowing.

Fig. 3 Frequency distribution of the number of days to 50% maturity in cowpea germplasm lines when drought stressed or not stressed. (A colour version of this figure can be found online at http://www.journals.cambridge.org/pgr)
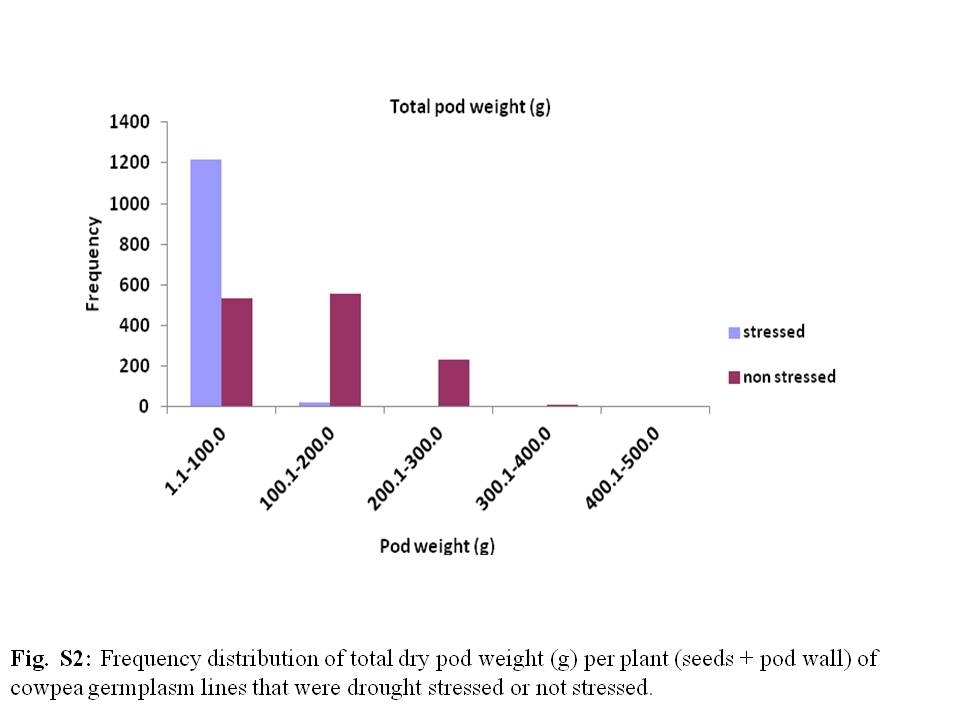
The grain yield per plant among the germplasm lines used in this study ranged from 0.0 g (in those that did not produce the pods under water stress or in which the pods shattered and the seeds dispersed before the harvest) to 15.4 g in line TVu9693 in the water-stressed plot and from 1.5 g in line TVu6644 to 58.4 g in line TVu441 in the non-stressed plot (Fig. 4). On average, water-stressed plants had a lower grain yield per plant than when non-stressed. The average grain yields per plant were 5.6 and 17.2 g when the plants were water stressed and non-stressed, respectively. Among the cowpea lines, the extent of grain yield reduction per plant due to water stress ranged from 0.0% in line TVu14195 to 100% in lines with a mean of 67.28%. TVu1436, TVu9693, TVu12115, TVu14632 and TVu15055 are the five germplasm lines with the highest grain yield per plant under the water-stressed condition. Among the early flowering group, lines TVu1059, TVu3562, TVu4607 and TVu7061 produced the highest grain yield while among the late flowering lines, the highest grain yield was exhibited by TVu522, TVu975, TVu3452, TVu3898 and TVu12568. Of the lines mentioned above, only TVu1436, TVu12568, TVu14632 and TVu15055 ranked among the 100 highest grain-yielding lines under the non-stressed condition. Grain yield reductions of 100% were recorded in the lines that produced no pods as well as in many of those whose seeds were dispersed following shattering of pods that was exacerbated by drought and low relative humidity. The total weight of pods (seeds+pod wall) per plant followed the same trends as for seed weight per plant (Supplementary Fig. S2, available online only at http://journals.cambridge.org). Very strong relationships were observed between seed weight and pod weight per plant when stressed (R 2 = 0.9198) and not stressed (R 2 = 0.9539).

Fig. 4 Frequency distribution of seed weight (g) per plant in cowpea germplasm lines when drought stressed and not stressed. (A colour version of this figure can be found online at http://www.journals.cambridge.org/pgr)
Based on the grain yield per plant under drought, the relative grain yield reduction due to drought and the stay-green characteristic, 142 lines (Supplementary Table S2, available online only at http://journals.cambridge.org) were selected for further evaluation in the screenhouse. The screenhouse evaluation discriminated the susceptible checks that were included in the study. No plant survived when watering was resumed among the susceptible checks and some of the 142 tested lines. The two early maturing lines ‘Sanzi’ and IT88D-345, which did not recover when watering was resumed, had flowered and produced one pod each with few seeds per pod before drying up (Supplementary Figs. S3 and S4, available online only at http://journals.cambridge.org). All the four plants of the six lines (Danila, TVu557, TVu1438, TVu4574, TVu6443 and TVu11982) recovered when watering was resumed 28 d after the imposition of drought. These six lines with vegetative-stage and flowering-stage drought tolerance have been included among those used as parents in crosses to generate segregating populations from which lines having enhanced drought tolerance and farmers' preferred traits will be selected.
Discussion
To develop new cowpea varieties with better performance under drought conditions requires the identification of new sources of genes with effects on this trait. When these genes are added to those presently available in existing improved varieties, the latter's levels of drought tolerance would be further enhanced. An evaluation of germplasm lines for their drought tolerance should make it possible to identify those with the desirable genes. The present study revealed a variation in drought tolerance among the 1291 cowpea germplasm and breeding lines tested. In earlier studies on drought tolerance in cowpea, Watanabe et al. (Reference Watanabe, Hakoyama, Terao and Singh1997) found some germplasm lines with better drought tolerance than many of the improved breeding lines and varieties. Some of these lines have been used to generate improved cowpea breeding lines. In the present study, three improved breeding lines, namely IT99K-494-6, IT99K-1122 and IT98K-506-1, with good performance under drought were included. The results have identified germplasm lines with a higher level of drought tolerance than these improved lines.
In order for a rapid progress to be made in the development of more drought-tolerant cowpea varieties, it would be necessary to identify easily recognized characteristics that are associated with this trait for which selection could be applied. While Agbicodo et al. (Reference Agbicodo, Fatokun, Muranaka, van der Linden and Visser2008a, Reference Agbicodo, Fatokun, Muranaka, van der Linden and Visserb) reported that stomatal conductance was associated with drought tolerance in cowpea, Hall et al. (Reference Hall, Thiaw, Ismael and Ehlers1997) could not find any positive relationship between water-use efficiency and drought tolerance in the crop. Belko et al. (Reference Belko, Zaman-Allah, Cisse, Diop, Zombre, Ehlers and Vadez2012) have also reported that lower early plant vigour, lower transpiration rate under non-moisture stress conditions during the vegetative stage of growth, lower leaf area development and lower canopy conductance under high vapour pressure deficit helped in distinguishing drought tolerant from sensitive cowpea lines. The authors have also reported a significant and close relationship between transpiration rate and transpiration efficiency. A relatively higher grain yield under drought could also be used for selecting cowpea lines with enhanced levels of drought tolerance.
The use of wooden boxes or pots for screening for drought tolerance in the screenhouse could help to quickly identify cowpea plants that would show drought tolerance especially in the vegetative stage. However, Ewansiha and Singh (Reference Ewansiha and Singh2006), Watanabe et al. (Reference Watanabe, Hakoyama, Terao and Singh1997) and Singh et al. (Reference Singh, Mai-Kodomi and Terao1999) have reported positive and significant correlations between drought tolerance in the seedling stage as observed using wooden boxes placed in the greenhouse and drought tolerance in the field. They therefore have concluded that cowpea lines found to be drought tolerant at the seedling stage as found when using wooden trays placed in the greenhouse should also perform well under drought in the field. While the box screening method may be rapid and help to identify lines with vegetative-stage drought tolerance, those with escape mechanisms, reproductive-stage drought tolerance and stay-green characteristic could be missed, hence the need for the evaluation of germplasm lines in the field. In the present study, the field evaluation helped to identify lines with reproductive-stage drought tolerance and stay-green characteristic, both of which could enhance better performance of cowpea under drought (Turk et al., Reference Turk, Hall and Asbell1980; Ziska et al., Reference Ziska, Hall and Hoover1985; Hall, Reference Hall2004).
We observed that leaves on most cowpea lines changed their orientation by assuming a paraheliotropic position especially when the plants were drought stressed. Schackel and Hall (Reference Schackel and Hall1979) made a similar observation and reported that leaves on cowpea plants under water stress assumed this position in order to reduce evapotranspiration. The leaves are said to be generally cooler when they assume the paraheliotropic position. This probably is part of the mechanisms adopted by cowpea plants for conservation of moisture and survival when exposed to drought conditions.
A number of the cowpea germplasm lines retained their green leaves in the field even under drought stress for up to 6 weeks. The ability of some cowpea plants to retain their leaves in the green state when water stressed for such a length of time has previously been reported by Hall et al. (Reference Hall, Thiaw, Ismael and Ehlers1997) and Gwathmey and Hall (Reference Gwathmey and Hall1992). This trait is commonly referred to as stay green or DLS. Their studies have revealed that cowpea plants which exhibit this trait have the capacity to survive the mid-season drought caused by an intermittent rainfall and are able to come into flowering and pod production should rainwater later become available. Flowering and pod formation continued for a longer time in the DLS cowpea lines when rainfall became stable and this should translate to more grain yield when compared with lines lacking the trait. In sorghum, a positive association has been reported between stay green and grain yield when planted in drought conditions (Borrell and Douglas, Reference Borrell and Douglas1996). In some Lolium perenne populations, Thorogood et al. (Reference Thorogood, Humphreys, Turner and Laroche1999) reported the detection of six quantitative trait loci with the effects on leaf senescence characteristic.
Crossing plants with the ability to flower early, as exhibited by a number of the cowpea lines evaluated in this study, with those with DLS characteristic, as shown by some other lines, should produce progeny that combine both attributes. Hall et al. (Reference Hall, Cisse, Thiaw, Elawad, Ehlers, Ismail, Fery, Roberts, Kitch, Murdock, Boukar, Phillips and McWatters2003) reported that in Senegal, an early erect DLS cowpea cultivar started flowering in about 35 d and produced 2000 kg/ha grain by 60 d followed by a second flush of pods that gave additional 1000 kg/ha grain in 100 d from sowing. In the dry savanna agroecologies of SSA, it has become more frequent for crops in farmers' fields to experience irregular rainfall during the cropping season. Drought could occur very early in the season in which rainfall ceases for several days shortly after planting. It may also occur during the mid-season when plants are yet to flower or during the reproductive phase. Cowpea varieties that flower early and still have stay green characteristics should be able to give farmers some grain yield even with irregular rainfalls. Hall (Reference Hall2004) concluded that such varieties would enable farmers to obtain good harvests in areas with tendency for mid-season drought occurrence. In addition, more stable yield should be expected from plants that combine drought tolerance at the seedling stage, reproductive stage and DLS under drought conditions. If cowpea varieties that combine the above attributes become available, they will enable farmers to obtain a better grain yield in those years when rainfall is irregular. While early flowering is a drought escape mechanism, drought tolerance at the seedling stage and DLS should enhance the plants' ability to survive drought during early and mid season and at pod filling. The prolonged life span that DLS confers on plants would also add to the plants' ability to tolerate terminal drought better. In sorghum, genotypes with DLS (stay green) remain physiologically active during the late stages of grain filling, and this enhances the plants' stress tolerance by increasing assimilate supply for grain filling and maintaining root function or moisture uptake, or even both (van Oosterom et al., Reference van Oosterom, Jayachandran and Bidinger1996). However, Ziska et al. (Reference Ziska, Hall and Hoover1985) reported that in cowpea, grain yield is strongly dependent upon the water available to the plant during the reproductive stage and with very little influence when drought occurred during the vegetative stage of growth. Rosenow et al. (Reference Rosenow, Quisenberry, Wendt and Clark1983) have suggested DLS as an indirect selection criterion in breeding for post-flowering drought tolerance in crops. The sufficiently high heritability for DLS and substantially low genotype by environment (g × e) interactions for this trait in cowpea (Hall et al., Reference Hall, Thiaw, Ismael and Ehlers1997) would enable successful incorporation of this trait in improved varieties.
Conclusion
The variation observed among the cowpea germplasm lines for drought tolerance in this study following evaluations in the field and greenhouse is an indication that progress should be expected in efforts aimed at enhancing the level of drought tolerance in presently available cowpea varieties. Combining drought tolerance at the seedling stage, early flowering and DLS in new cowpea varieties should lead to the enhancement of their drought tolerance.
Acknowledgements
The authors would like to thank the Tropical Legumes II project for providing funds through the International Crops Research Institute for the Semi-Arid Tropics and the IITA for all the support received.