


INTRODUCTION
Monogenean gyrodactylids are ubiquitous ectoparasites on the skin and gills of teleost fishes (Bakke et al. 2002), with the most recent compilation listing 409 species (Harris et al. 2004). Although most Gyrodactylus species are considered non-pathogenic, this is not true for Gyrodactylus salaris Malmberg, 1957. After the first observation of G. salaris in Norway (Johnsen, 1978), this parasite has seriously hampered the natural, juvenile production of wild Atlantic salmon (Salmo salar L.) and caused great harm both in ecological and economical terms (Johnsen et al. 1999; Mo et al. 2004). Thus, it is not surprising that G. salaris is the best studied of the Gyrodactylus species.
Recent studies indicate that the original species description of G. salaris is no longer satisfactory for a proper management of gyrodactylosis. Ziętara and Lumme (2002) showed that the majority of Gyrodactylus species can be discriminated based on sequencing of the internal transcribed spacer (ITS) regions (ITS-1 and ITS-2) of the nuclear ribosomal DNA (rDNA). However, G. salaris cannot be differentiated from G. thymalli Žitnaň, 1960, parasitizing grayling (Thymallus thymallus L.) based on the ITS-1 and ITS-2 sequences (Cunningham, 1997). In fact, parasites from Atlantic salmon, grayling, and rainbow trout (Oncorhynchus mykiss (Walbaum)) share identical ITS-1 and ITS-2 sequences over a wide geographical range (Ziętara and Lumme, 2002). However, G. salaris and G. thymalli can be grouped into several well-supported clades based on the sequences of the cytochrome c oxidase subunit 1 (cox1) gene of the mitochondrial DNA (mtDNA) (Hansen et al. 2003, 2006; Meinilä et al. 2004). However, there was no support for the monophyly of either of the 2 species. Thus, groups based on host specificity or pathogenicity to Atlantic salmon, are paraphyletic with reference to cox1 genealogy.
Despite G. salaris and G. thymalli being very closely related, their host-species preferences, as observed from laboratory infection experiments, are different (Soleng and Bakke, 2001; Bakke et al. 2002; Sterud et al. 2002). G. salaris is pathogenic to its host, the Atlantic salmon, whereas G. thymalli appears to be non-pathogenic to any known potential host (Soleng and Bakke, 2001; Bakke et al. 2002; Sterud et al. 2002).
The host preferences and specificity of G. salaris have been studied extensively (see Bakke et al. 2002) but, thus far, only little attention has been paid to possible variation in infectivity and reproductive potential among different populations of G. salaris. Nonetheless, Lindenstrøm et al. (2003) described a variant of G. salaris (Gx) exhibiting limited reproduction on Atlantic salmon. This variant, originating from a rainbow trout farm in Denmark, performed better on rainbow trout than on any of the experimentally infected Atlantic salmon stocks. A genetic characterization of this parasite based on the ITS region revealed 3 nucleotide substitutions (C<->T at position 276 in the ITS-1 and C<->T at position 911 and A<->T at position 1090 in the ITS-2) compared with the ‘standard’ G. salaris sequence (see Lindenstrøm et al. 2003).
Recently, Robertsen et al. (2006b) discovered a Gyrodactylus sp. parasitizing Arctic charr (Salvelinus alpinus) in Lake Pålsbufjorden, Buskerud County, southern Norway. This parasite was similar morphologically to G. salaris, and sequence data for the cox1 revealed a haplotype which was identical to that found in G. salaris populations parasitizing Atlantic salmon in the neighbouring Rivers Drammenselva and Lierelva (Buskerud County) and Lærdalselva (Sogn og Fjordane County), as well as the rainbow trout in the Swedish Lake Bullaren (sequences from Hansen et al. 2003). Despite a difference of 1 nucleotide in the ITS-2 (G<->A at position 288) between G. salaris of the same cox1 haplotype from Arctic charr and Atlantic salmon (Hansen et al. 2003), Robertsen et al. (2006b) concluded that this variant represented G. salaris.
The fish community in Lake Pålsbufjorden consists of Arctic charr, brown trout and minnow (Phoxinus phoxinus). Thus, this is the first observation of G. salaris maintaining a viable population in the absence of Atlantic salmon or other salmonids which are susceptible to G. salaris (see Jansen and Bakke, 1995; Bakke et al. 2002). Lake Pålsbufjorden drains into River Numedalslågen, in which the stretches holding anadromous Atlantic salmon end some 100 km downstream the outlet of the lake. Previously, observations of Arctic charr infected with G. salaris have been related only to river systems in which Arctic charr live in sympatry with Atlantic salmon which is also infected with G. salaris (see Mo, 1988; Knudsen et al. 2006, Kristoffersen et al. 2005). Arctic charr and rainbow trout (the latter species previously occurring in Lake Pålsbufjorden) are salmonid species susceptible to G. salaris from the River Lierelva (Bakke et al. 1990, 1991, 1996).
Gyrodactylosis of Atlantic salmon caused by G. salaris is a notifiable disease (OIE, 2004). Hence, a spread of the parasite downstream to salmon-populated, anadromous stretches of River Numedalslågen may have significant epidemiological and ecological consequences. Therefore, we studied the infectivity of G. salaris from Arctic charr from Lake Pålsbufjorden to Atlantic salmon and compared it with a known virulent G. salaris population from Atlantic salmon in the Lierelva/Drammenselva river complex (Bakke et al. 1990, 2002).
MATERIALS AND METHODS
Origin of the parasites
Adult Arctic charr infected with G. salaris were caught by overnight gillnetting in Lake Pålsbufjorden, Buskerud County and infected parr of Atlantic salmon were caught by electro-fishing in River Lierelva and the neighbouring River Drammenselva. The fish were transported alive to the laboratory in aerated plastic bags containing lake water. In the laboratory, the fish were kept in 200 l grey plastic tanks in charcoal-filtered tap-water (flow rate 2–4 l/min). Subsequently, the fish were anaesthetized in 0·04% chlorobutanol and killed by a blow to the head before the fins were cut off and examined for parasites in tap water under a stereomicroscope with fibre optic illumination. G. salaris from the adult Arctic charr were used directly, but G. salaris from the 2 stocks of Atlantic salmon parr were transferred to uninfected salmon parr and kept (in separate tanks) for >1 month prior to the start of the experiments to eliminate any accidental contamination with G. derjavini. As gyrodactylids are sensitive to host serum (see Buchmann, 1998; Harris et al. 1998), the water was changed several times between examinations for parasites and further experimental infection with the specimens. The Gyrodactylus specimens infecting Arctic charr and Atlantic salmon have been identified previously as G. salaris both by morphometry and sequencing of the ITS and intergenic spacer (IGS) regions of nuclear ribosomal DNA and the mitochondrial cox1 (Robertsen et al. 2006b).
Origin of the fish hosts
Experimental infections were performed on wild Atlantic salmon from River Numedalslågen, Buskerud County, southeastern Norway (1·4–8·2 g; 6–10 cm). The fish were caught by electro-fishing and brought to the aquarium in aerated plastic bags. All Atlantic salmon individuals from River Numedalslågen were tested genetically at the Norwegian Institute for Nature Research (NINA, Trondheim) for the possibility of brown trout (Salmo trutta L.) or Atlantic salmon×brown trout hybrids among them, which may interfere with the interpretation of results (see Bakke et al. 1999), but there was no evidence of these fishes. Due to low numbers and general problems acclimatizing wild fish to aquarium conditions, 1 experimental set-up included hatchery-reared Atlantic salmon (1·5–9·4 g; 6–11 cm) from River Drammenselva, Buskerud County, southeastern Norway. Since Arctic charr and rainbow trout have immigrated and been stocked, respectively, in Lake Pålsbufjorden, representatives of both species were included in the experiments. Hatchery-reared Arctic charr (2·3–9·2 g; 7–11·5 cm) originated from Tallvik settefiskanlegg, Alta, Finmark County, North Norway, and commercially reared rainbow trout (1·3–6·9 g; 5·5–9·5 cm) were supplied by Valdres ørretoppdrett, Røn, Buskerud County, southern Norway.
Infection experiments
The infection experiments were performed in the aquarium facilities in the Department of Zoology, Natural History Museum, University of Oslo. All fish were acclimatized for >1 month to the laboratory conditions prior to the commencement of the experiments. The fish were infected individually with 1–4 specimens (Experiment I, see below) or 1 specimen (Experiments II–V, see below) of G. salaris. For the initial infection with 1 parasite, the flukes were induced to attach to the tip of an insect pin before being moved to attach on to the bottom of a glass Petri dish. The parasites were then presented to the caudal fin of an anaesthetized fish, to which they frequently attached spontaneously. Extreme care was taken to prevent any contamination of parasites with host body fluids. For the initial infection with >1 G. salaris, the hosts were infected through the exposure to fins from infected, adult Arctic charr. After being infected, individual hosts were kept isolated in grey floating plastic boxes (20×10×10 cm) with wire-mesh bottoms to ensure a free flow of fresh water into the boxes. Infections were considered successful if 1 or more worms were present on the host 2 days post-infection (p.i.). The course of infection was monitored on a weekly basis (up to 11 weeks) by counting all of the parasites on the skin and fins of infected fish. For this examination, fish were anaesthetized in 0·04% chlorbutanol. The water temperature ranged from 7 to 12 °C during the experimental period. The fish hosts were under daily supervision, and dead fish were removed immediately.
Experimental protocol
Five different experiments were carried out.
Exp. I (Fig. 1A and B)
Wild Atlantic salmon from River Numedalslågen were each infected with G. salaris from Lake Pålsbufjorden. Originally, 21 fish were attempted to be infected with 1 G. salaris each. Due to the low number of fish available and low establishment of infection, hosts not successfully infected were attempted to be re-infected. Also, 8 fish were initially infected with [ges ]1 parasite, as described above (hosts were previously naïve). The experiment was conducted during 11 weeks.

Fig. 1. (A–C) The course of Gyrodactylus salaris infection on individually isolated wild salmon from River Numedalslågen. The data are based on weekly counts of the parasites on the skin and fins of successfully infected fish. (A) Exp. I: infection with G. salaris from Arctic charr in Lake Pålsbufjorden. Hosts initially infected with 1 parasite (n=9 hosts). (B) Exp. I: infection with G. salaris from Arctic charr in Lake Pålsbufjorden. Hosts initially infected with 1–4 parasites (n=6 hosts). (C) Exp. II: G. salaris from Atlantic salmon in River Lierelva (n=9 hosts). The salmonid fish in Exps I and II were kept in separate tanks. Note the different scales of the ordinate.
Exp. II (Fig. 1C)
Wild Atlantic salmon (n=13) from River Numedalslågen were each infected with 1 G. salaris from River Lierelva, as a control group for Exp. I. The G. salaris in River Lierelva is a known pathogen of Atlantic salmon (see Johnsen et al. 1999). This experiment was conducted for 10 weeks in a separate tank in parallel with Exp. I.
Exp. III (Fig. 1A–C)
Hatchery-reared Atlantic salmon from Drammenselva (duplicated, n=18 and 17), Arctic charr (n=18) from Alta and rainbow trout (n=18) from Røn were each infected with 1 G. salaris from Arctic charr from Lake Pålsbufjorden and kept in 3 separate tanks.
Exp. IV (Fig. 2D–F, Table 1)
Hatchery-reared Arctic charr (n=6), rainbow trout (n=6) and Atlantic salmon (n=6) were each infected with 1 G. salaris from Arctic charr from Lake Pålsbufjorden and kept individually in the same tank in small boxes.
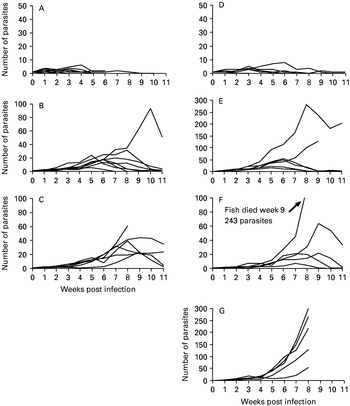
Fig. 2. (A–G) The course of Gyrodactylus salaris infection from Arctic charr in Lake Pålsbufjorden (A–F) and from Atlantic salmon in River Drammenselva (G) on individually isolated salmonids, in separate tanks (A–C, Exp. III and G, Exp. V) and in 1 common tank (D–F, Exp. IV), infected with 1 parasite each. The data are based on weekly counts of the parasites on the skin and fins of the successfully infected fish. (A) G. salaris on Atlantic salmon (n=13) of the Drammenselva stock. (B) G. salaris on Arctic charr (n=9) of the Tallvik stock. (C) G. salaris on rainbow trout (n=7) from Valdres commercial farm. (D) G. salaris on Atlantic salmon (n=6) of the Drammenselva stock. (E) G. salaris on Arctic charr (n=6) of the Tallvik stock, (F) G. salaris on rainbow trout (n=6) of the Valdres stock. (G) G. salaris on Atlantic salmon (n=6) of the Drammenselva stock. Note the different scales of the ordinate.
Table 1. Mean, median, maximum (Max) and minimum (Min) observations and number of infected fish (n) in weekly counts on Atlantic salmon, Arctic charr and rainbow trout successfully infected with Gyrodactylus salaris from Arctic charr in Lake Pålsbufjorden The fish were initially infected with 1 parasite and subsequently kept individually isolated in a common tank (Exp. IV). These data are based on weekly counts of the parasites on the skin and fins of the fish.

Exp. V (Fig. 2G)
Hatchery-reared Atlantic salmon (n=6) from the same stock as used previously (River Drammenselva) were included as a control group and each was infected with 1 G. salaris from River Drammenselva. The G. salaris in River Drammenselva is a known pathogen to Atlantic salmon (see Johnsen et al. 1999). This experiment was conducted simultaneously with Exp. III.
Statistical test
In an attempt to deduce the analytical power of the timing of the peak-points along the trajectories of infection intensities, we tested whether the peak points were locally statistically significant. A statistical method was employed for all host-parasite combinations, testing whether pairwise consecutive sampling points were equal or not. Tests for significant differences between pairwise similar sampling points (week) in Exps III and IV were conducted for each of the 3 host species, in order to reveal any effects of the experiments being carried out in separate tanks. Additionally, we tested for statistically significant differences along the trajectories of infection intensity between experimental groups of salmon and their respective controls (Exp. III vs. Exps V and IV vs. Exp.V). In all statistical tests, the method used was Kruskal-Wallis one-way analysis of variance, run in SYSTAT, ver. 10 (©SPSS Inc.).
RESULTS
The establishment success of G. salaris from Arctic charr in Lake Pålsbufjorden on Atlantic salmon from Rivers Numedalslågen and Drammenselva as well as on Arctic charr and rainbow trout was limited. Total average successful establishments from Exps III–IV, were 38·0, 48·5 and 30·5% on Atlantic salmon, Arctic charr and rainbow trout, respectively. Unexpectedly, the pathogenic G. salaris from Drammenselva established successfully on Atlantic salmon from River Drammenselva (Exp. V) in only 33·5% of all transfer attempts of the parasite. A number of fish died during the run of the experiments. The cause of death is unknown, but is assumed to not be related to Gyrodactylus infection, as numbers of parasites on dead fish were relatively low (maximum number of parasites registered on a dead host: 250).
Exp. I (Fig. 1A,B)
In total, 37 attempts were made to infect Atlantic salmon from River Numedalslågen with G. salaris from Lake Pålsbufjorden. Nine infections (24·3%) were successful. Of these 9 infections, 3 were successful on a first attempt, whereas 5 needed 2 and 1 needed 3 attempts. Reproduction was observed for 8 (88·9%) of the 9 successfully infected fish; this includes 2 host fish with parasite reproduction limited to 1 birth (Fig. 1A). The highest observed intensities from this experimental set-up were 109 and 82 parasites before apparently levelling off. The maximum intensity of infection on the other fish did not exceed 40 parasites. Four host fish were able to eliminate the infection during the experimental period. Attempts were made to infect Atlantic salmon from River Numedalslågen with >1 G. salaris from Lake Pålsbufjorden (once for each host). Six of 8 fish (75·0%) were successfully infected. Reproduction of G. salaris was observed on all successfully infected fish. However, the infrapopulations did not exceed 20 parasites, except on 1 fish with an intensity of 250 parasites at week 9 (Fig. 1B). None of these infections were eliminated during the course of the experiment.
Exp. II (Fig. 1C)
In the control group with wild Atlantic salmon from River Numedalslågen infected with G. salaris from River Lierelva, 9 of the 18 (50·0%) salmon were successfully infected. In this tank, reproduction was observed on all but 1 of the successfully infected salmon. All but this single salmon with a maximum of 6 parasites during the experimental period, were highly susceptible. The maximum number of parasites ranged from 70 to 742 by week 10 (Fig. 1C).
Exp. III (Fig. 2A–C)
Of the 18 hatchery-reared Atlantic salmon, 13 (72·2%) were each infected successfully with 1 G. salaris from Lake Pålsbufjorden (Fig. 2A). Reproduction was observed for 6 (46·2%) of these 13 fish, including 2 fish with reproduction limited to 1 birth. The parasite numbers increased for up to 4 weeks p.i. The highest intensity of infection observed was 6 parasites. All 13 G. salaris infrapopulations were eliminated by 8 weeks. One infected salmon died during the experiment. Of the 18 Arctic charr, 9 (50%) were successfully infected (Fig. 2B). Of these 9 fish, reproduction of G. salaris from Lake Pålsbufjorden was observed on 8 (88·9%). The peak intensity of infection was 93 parasites. The parasite infrapopulations increased in number from 5 to 10 weeks p.i. On 6 of the 9 (66·7%) successfully infected Arctic charr the parasite infrapopulations were eliminated during the experimental period of 11 weeks. Seven of the 18 (38·9%) rainbow trout were successfully infected with 1 G. salaris from Lake Pålsbufjorden (Fig. 2C). Reproduction was observed for 6 (85·7%) established infections. The peak intensity of infection was 61 parasites. An established infection was not eliminated during the course of the experiment. The parasite infrapopulations increased in number in 4 rainbow trout 7 weeks p.i. Two of the rainbow trout died during the experiment.
Exp. IV (Fig. 2D–F, Table 1)
The course of the infection of G. salaris from Lake Pålsbufjorden on individually isolated, hatchery-reared Atlantic salmon from Drammenelva (n=6), Arctic charr (n=6) and rainbow trout (n=6) in a common tank is shown in Fig. 2D–F. One salmon was considered to be innately resistant; for the others, the infection was eliminated by week 9. During the experimental period of 11 weeks, 2 Arctic charr and 1 rainbow trout eliminated their infections. The time of peak for the intensity of infection varied between host species: on salmon, 3–6 weeks p.i. (Fig. 2D), on Arctic charr and rainbow trout, 5–8 and 7–9 weeks p.i., respectively (Fig. 2E and F). The maximum numbers of parasites during the entire experimental period were 8, 282 and 243 for Atlantic salmon, Arctic charr and rainbow trout, respectively. One Arctic charr and 1 rainbow trout died during the experiment.
Exp. V (Fig. 2G)
In the control group with hatchery-reared Atlantic salmon from Drammenselva infected with G. salaris from Drammenselva, 6 of the 18 (33·3%) salmon were successfully infected. In this tank, reproduction was observed on all 6 infected salmon. The salmon were highly susceptible to the infection, and the size of the parasite infrapopulations increased continuously, albeit at different rates on individual fish (Fig. 2G). One salmon died during the experiment.
Substantial variation in susceptibility was recorded among the host individuals. The time lapse to an acquired response could not be specifically determined, as there were no statistically significant differences in mean parasite intensity between any 2 consecutive monitoring points for any of the infection trajectories in Exps I – V (Kruskal-Wallis one-way analysis of variance, P>0·05). The influence of a ‘tank effect’ might be present, as the parasite intensities of infection for Arctic charr were significantly different (P<0·05) in the weeks 1–6 between Exps III and IV. In similar tests for Atlantic salmon and rainbow trout, there were no significant differences (P>0·05). Infection intensity among salmon in Exp. I was significantly different (Kruskal-Wallis one-way analysis of variance, P<0·05) from the control in Exp. II from week 5 and throughout the experiment. Infection intensity among salmon in Exp. III was significantly different (Kruskal-Wallis one-way analysis of variance, P<0·05) from the control in Exp. V from week 2 and throughout the experiment. For Exp. IV, the infection trajectory was significantly different (P<0·05) from the control group in Exp. V from week 3.
DISCUSSION
The high pathogenicity of G. salaris from the Rivers Lierelva and Drammenselva on Atlantic salmon from the same rivers is well documented (see e.g. Johnsen et al. 1999). By contrast, G. salaris from Arctic charr in Lake Pålsbufjorden displayed (in general terms) a very restricted reproduction on hatchery reared Atlantic salmon parr from Drammenselva and also on wild Atlantic salmon parr from Numedalslågen, despite individual variation. Therefore, we consider G. salaris from Arctic charr in Lake Pålsbufjorden to be non-pathogenic to Atlantic salmon, at least from Drammenselva and Numedalslågen. The apparent differences in susceptibility between the 2 experimental groups, most likely reflect the fact that the former were hatchery-reared, whereas the latter from Numedalslågen represented a wild population, and thus would probably have been more stressed under laboratory conditions. This G. salaris population was not pathogenic to the Arctic charr or the rainbow trout stocks, although both appeared susceptible and suitable hosts. Based on these findings, the susceptibility of the 3 host species tested may be summarized as follows: Atlantic salmon was slightly susceptible, whereas Arctic charr and rainbow trout were moderately susceptible.
In recent years, a particular region of cox1 (~800 bp) has been sequenced for G. salaris and G. thymalli from >30 different geographical populations in Europe. A number of haplotypes have been identified (Hansen et al. 2003, 2006; Meinilä et al. 2004) which group into 3–6 distinct clades. The sequence data supported 3 independent introductions of G. salaris into Norway (Hansen et al. 2003). Robertsen et al. (2006b) reported that the cox1 sequence of G. salaris on Arctic charr in Lake Pålsbufjorden was the same as that of G. salaris recovered previously from Atlantic salmon from the Rivers Drammenselva, Lierelva and Lærdalselva in Norway as well as on rainbow trout from a hatchery in the Swedish Lake Bullaren (annotated clade III, haplotype F by Hansen et al. 2003). Additional sequence data for the ribosomal IGS and comparative morphometry of the haptoral hard parts of the parasites also identified this parasite as G. salaris (see Robertsen et al. 2006b).
Lindenstrøm et al. (2003) reported a G. salaris variant (Gx) from farmed Danish rainbow trout, which was non-pathogenic to Atlantic salmon in infection experiments. This observation may raise the question as to the relatedness of Gx and the G. salaris discovered in Lake Pålsbufjorden. However, the ITS sequence of the G. salaris variant described by Lindenstrøm et al. (2003) differs by 3 nucleotide substitutions from the G. salaris consensus sequence. Robertsen et al. (2006b) reported a single nucleotide substitution in the ITS-2 for the G. salaris parasites from Arctic charr in Lake Pålsbufjorden; this equates to a total of 4 nucleotide substitutions when the ITS sequences of Gx are compared with those of G. salaris from Lake Pålsbufjorden. More recently, another Danish G. salaris, which also had a low pathogenicity to salmon, has been discovered (Thomas Jørgensen and Kurt Buchmann, personal communication). It seems that G. salaris which are non-pathogenic to Atlantic salmon may occur more frequently than previously thought.
Halvorsen and Hartvigsen (1989) suggested a list of possible scenarios regarding G. salaris in Norway. One of these scenarios was that G. salaris is endemic in Norway. Currently, the introduction of different pathogenic G. salaris strains to Norway by several routes is unquestioned. Hansen et al. (2003) proposed at least 3 independent introductions on the basis of cox1 sequence data. However, the proposal of endemism of a benign G. salaris parasitizing an inland population of Arctic charr cannot be rejected, but historical events strongly support, also in this case, the hypothesis of a relatively recent introduction of the species. Originally, Arctic charr was introduced into the Numedalslågen river system from Lake Tinnsjøen (Aass, 1970), a lake in which Arctic charr infected with G. salaris has not yet been found (Robertsen et al. 2006a). However, rainbow trout has been introduced directly into several lakes connected to the Numedalslågen river system, including Lake Pålsbufjorden in the early 1960s. The rainbow trout originated from Denmark but came to Pålsbufjorden via various Norwegian hatcheries and farms (Per Aass, personal communication). As G. salaris has been shown previously to have a relatively wide host range among salmonids (Bakke et al. 2002) and host switches are considered common for species of Gyrodactylus (see Ziętara and Lumme, 2002; Huyse and Volckaert, 2002, 2005), it is reasonable to assume that G. salaris has been transferred to Arctic charr in this area through infected rainbow trout. Subsequently, the G. salaris may have adapted to the particular ecology of Arctic charr after rainbow trout disappeared from the lake (Robertsen et al. 2006b). G. salaris in the Rivers Drammenselva and Lierelva has also most likely been introduced via rainbow trout but, in this case, from Sweden (Mo, 1991; Hansen et al. 2003). Both G. salaris populations have the cox1 haplotype F which is commonly found on rainbow trout in farms throughout Fennoscandia (Meinilä et al. 2004).
Employing cox1 sequence data sets, several well supported clades of both G. salaris and G. thymalli (based on mitochondrial haplotypes) have been described, but there is no support for the monophyly of all haplotypes of either species (Hansen et al. 2003, 2006; Meinilä et al. 2004). However, host preference and host specificity vary significantly between at least some of the populations of G. salaris and G. thymalli (see Soleng and Bakke, 2001; Bakke et al. 2002; Sterud et al. 2002). Sterud et al. (2002) considered these biologically significant differences important when concluding that G. salaris and G. thymalli should remain as 2 distinct species. The present results indicate that the G. salaris population in Lake Pålsbufjorden is non-pathogenic to Atlantic salmon, and, importantly, Arctic charr and rainbow trout seem to be more suitable as hosts. The present results indicate that host specificity is no longer a striking argument for considering G. salaris and G. thymalli as 2 species. Previous experiments (unpublished data) indicate that G. thymalli from grayling, the preferred host of G. thymalli, can also reproduce on Arctic charr; the reciprocal experiment of testing G. salaris from Arctic charr on grayling needs to be conducted.
Gyrodactylosis is probably one of the most important threats to wild European Atlantic salmon populations, and G. salaris is listed by the ‘Office International des Épizooties’ (OIE) in the Aquatic Animal Code (OIE, 2004). Several countries have taken precautionary measures to avoid the introduction of the worm. Accordingly, the European Commission (EC) has restricted the import of salmonids that are susceptible to G. salaris to areas with an equivalent health status (Peeler et al. 2006). In Norway, comprehensive and expensive counter measures, such as, for example, rotetone treatment programs for entire river systems, have been launched to control epidemics and to eliminate G. salaris infection from wild salmon. The present observations of differing host preferences and reproduction potential between different populations of G. salaris to Atlantic salmon is important for the design and implementation of management strategies and international legislation. Of paramount importance is that the identification of G. salaris based on any presently used molecular markers or morphology alone is not sufficient for a diagnosis of gyrodactylosis. Another key aspect in a management context is the apparent ease with which species and strains closely related to G. salaris undergo host switching with subsequent reproduction. Frequent host switches could ultimately be an avenue for a significant expansion of the geographical range of pathogenic variants of G. salaris.
We are grateful to Henning Pavels and Bjørn R. Hansen for providing live parasites from Lake Pålsbufjorden and Øyvind Fladaas, Frode Løvik and Røn Endre for providing hatchery reared salmon, Arctic charr and rainbow trout, respectively. Kjetil Hindar, at the Norwegian Institute for Nature Research (NINA), was responsible for the genetic tests of the wild fish from Numedalslågen. Cathrine Vollelv helped in the lab at the Natural History Museum. Two anonymous referees provided valuable comments on the manuscript. This study was supported by grants from the National Centre for Biosystematics (NCB) (Project no. 146515/420), co-funded by the Norwegian Research Council (NRC) and the Natural History Museum, University of Oslo, the NRC ‘Wild Salmon Programme’ (Project no. 145861/720) and the Directorate for Nature Research (DN) (Contract no. 05040026).



