Introduction
In the alpine and pre-alpine regions of Austria and its neighboring countries, ruminant production plays a vital role in organic agriculture, mostly due to the high proportion of total farm land that is permanent grassland. In Austria, 51.2% of the utilized agricultural area is permanent grassland 1 . Owing to ruminants’ ability to digest fibrous materials, dairy production systems are well suited for utilizing grassland for the production of foods. As a consequence, organic milk production is commonly based on fresh and conserved forages from permanent grassland and perennial clover–grass leys. The latter are an essential component of crop rotation, which is of particular importance in organic farming systems. In organic crop rotations, nitrogen (N)-demanding and N-fixing crops are balanced in order to optimize yields. On an organic dairy farm, sufficient forage for cows has to be produced while still maintaining soil fertilityReference Weller and Bowling 2 . Clover–grass provides both N for the next crop in the rotation as well as forage for the dairy cows. Ley farming also prevents soil erosion and provides carbon-rich crop residues as feed for soil organisms. Apart from feeding forages, most Austrian farmers supplement with concentrates to improve milk yield. Common home-grown concentrates are barley and triticale, whereas in commercial concentrates, industrial by-products (e.g., bran, press cake) are used as well. Where local growing conditions allow maize production, maize silage is valued as a high-energy forage. Including it in the diet increases the supply of ruminally fermentable carbohydrates from forage and thereby improves the capture of dietary N, a precious and scarce resource in organic agriculture. Studies in which perennial ryegrass silage is replaced with maize silage have shown that increasing inclusion of maize silage positively affects dry matter (DM) intake, milk yield, and milk protein content, while milk fat concentration is either not affected or decreasesReference Burke, Murphy, O'Donovan, O'Mara, Kavanagh and Mulligan 3 – Reference Kliem, Morgan, Humphries, Shingfield and Givens 5 .
Italian ryegrass (Lolium multiflorum Lam.) is a fodder grass that has been grown in northern Italy since the 12th centuryReference George 6 , making it the oldest fodder grass cultivated by humans. Today, Italian ryegrass can be found in temperate regions all around the world. In Austria, it can be found in natural grassland, but is rarely cultivated due to its low frost resistance, although using it for annual (or bi-annual, depending on the winter losses) leys would avoid this problem.
Among the fodder grasses, Italian ryegrass has one of the highest energy contents, and due to its favorable composition of structural carbohydrates, it is easily digestibleReference Tamburini, Rapetti, Crovetto and Succi 7 . Reports about its positive effects on the forage intake of dairy cows are frequentReference Baldinger, Baumung, Zollitsch and Knaus 8 , Reference Bernard, West and Trammell 9 , and some researchers even report better feed efficiency than when feeding maize silageReference Cooke, Bernard and West 10 .
In Austrian areas where the cultivation of both maize and Italian ryegrass is possible, Italian ryegrass may be an alternative to forage maize, resulting in longer rotation breaks between maize and therefore a more diverse crop rotation. In areas where conditions are not optimal for maize production, Italian ryegrass might be a valuable high-energy forage in addition to the usual clover–grass. Italian ryegrass and maize differ in their cultivation demands and characteristics: whereas maize only requires an annual precipitation of 500–800 mm 11 , the annual water demand of Italian ryegrass is much higher, about 900–1200 mmReference Dietl and Lehmann 12 . Both forage crops are sensitive to low temperatures: for successful cultivation of Italian ryegrass, an average annual temperature of about 8.5 °C is neededReference Dietl and Lehmann 12 . Maize for silage production requires an average temperature between 10 and 15 °C between May and September 11 . Italian ryegrass is either sown in fall or in spring, establishes quickly, and covers the ground effectively. Under Austrian conditions it rarely survives more than one winter. Maize is sown in April/May and harvested for silage in September/October, and especially under organic cultivation, establishment is slow. Owing to its high nutrient demand, maize is commonly planted after clover–grass or other legumesReference Freyer 13 . Italian ryegrass has no special demands with regard to the preceding crop, although it thrives when planted after legumes. Possible succeeding crops after Italian ryegrass are grain legumes or summer cerealsReference Freyer 13 . Special attention must be paid to the N supply of succeeding crops, because the crop residues left behind by Italian ryegrass are rich in carbon but poor in N. Italian ryegrass itself requires regular, moderate fertilization with manure in order to realize its potential. Thus, both maize and Italian ryegrass can be considered intensive forage crops under the conditions of organic agriculture.
The objective of this study was to compare Italian ryegrass silage and maize silage as forage for dairy cows under the conditions of Austrian organic agriculture. Therefore, a feeding trial was conducted in which Italian ryegrass silage and maize silage were included in grass-silage-based mixed basal diets for dairy cows during the winter feeding period. The main questions were how intake of DM, milk yield, and quality and production efficiency, especially gross N efficiency, would differ depending on the forages used. The milk fatty acid profile was chosen as an additional parameter to describe milk quality.
Materials and Methods
Italian ryegrass harvest
Italian ryegrass (L. multiflorum Lam. var. italicum, cultivar: TIGRIS) was grown in a field belonging to the Secondary Agriculture and Forestry College at Ursprung (HLFS Ursprung) in the province of Salzburg, Austria (570 m NN, 1250 mm precipitation, 8.5 °C average annual temperature). The preceding crop was clover–grass. After seeding on April 16, 2010, the first cut on May 11 was a cleaning cut due to weeds, and the second (June 24) and third cuts (July 19) were ensiled without additives as round bales. After the second and third cut, liquid manure was applied at a rate of 40 kg N (10 kg of which were NH4-N) per hectare. The two ensiled cuts combined yielded 2.9 t DM ha−1.
Experimental design and animals
The feeding trial was conducted with the organically managed HLFS Ursprung dairy herd, which is housed in a cubicle housing system equipped with Calan gates for individual feeding. Milking takes place in a 2×3 herringbone milking parlor twice a day, at 6:00 and 16:30. A total of 22 lactating Holstein cows were included in the feeding trial between November 2010 and February 2011, but only 19 cows were included for the whole trial. The remaining three cows were included belatedly because of delayed calving or were eliminated prematurely because they were dried off. At the beginning of the experiment, the cows were divided into two treatment groups of 11 cows each, based on number of lactation (2.5±1.7 versus 3.0±1.5), days in milk (110±65 versus 141±109), milk yield (27.4±9.5 versus 23.9±5.1 kg), and live weight (654±62 versus 665±47 kg). After an initial 3-week adaptation period to the Calan gates and the feeding regimen, data were collected over 12 weeks. Treatments were switched after week 6 to ensure that every cow received every treatment. The feeding trial was conducted according to the EU Directive 2010/63/EU for animal experiments 14 and the Austrian Act on Animal Experiments 15 . No invasive procedures were performed on the cows.
Feeding regimen
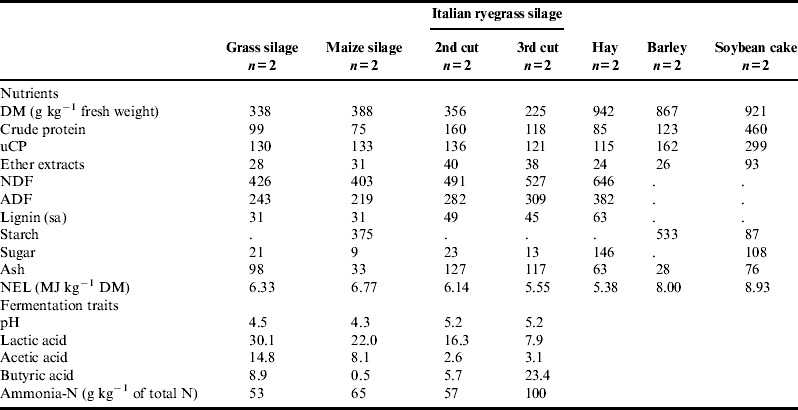
Two mixed basal diets based on grass silage and hay and supplemented with additional concentrates were compared. The nutrient content and fermentation traits of the feeds included in mixed basal diets are summarized in Table 1. Grass silage consisted of a mixture of a first cut from permanent grassland and a third cut from a perennial clover–grass ley in its third year; therefore, clover proportion was already low. Hay was produced from a first cut of permanent grassland, and residual drying was performed in the barn by a ventilation system. Owing to a crop failure at HLFS Ursprung, maize silage produced according to the regulations of Austrian organic agriculture was purchased from a farmer in the Austrian province Lower Austria. The DM content of the silages ranged between 338 and 388 g kg−1 fresh matter, except for the third cut of Italian ryegrass, which was poorly fermented and numerically lower in energy and DM (225 g kg−1 fresh weight) than the other silages. The crude protein content of the silages varied greatly, from as low as 75 g kg−1 DM in the maize silage to 99 g kg−1 DM in the grass silage, and up to 160 g kg−1 DM in the ensiled second cut of Italian ryegrass.
Table 1. Nutrient content and fermentation traits of silages, g kg−1 DM unless stated otherwise.

DM, dry matter; uCP, utilizable crude protein at the duodenum; NDF, neutral detergent fiber;
ADF, acid detergent fiber; and NEL, net energy lactation.
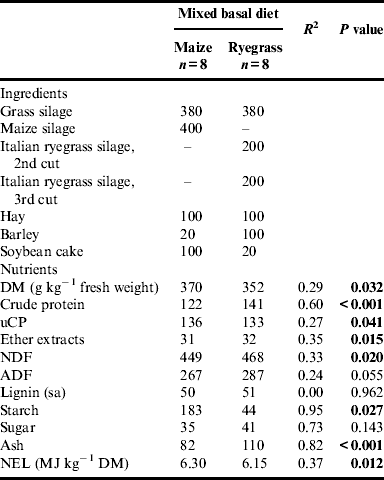
Table 2 shows the ingredients and nutrient content of the mixed basal diets. Considerable amounts of maize silage and Italian ryegrass were included in diet M (maize diet) and diet R (ryegrass diet), respectively, to make the effects of and differences between the two forages clearly visible. In order to avoid the negative effects of protein deficiency in the maize diet, differing amounts of barley and soybean cake were added to the forages. The resulting mixed basal diets were meant to be isonitrogenous with a calculated crude protein content of 142 g kg−1 DM, but not isoenergetic. However, in fact, the crude protein content of mixed basal diet R turned out to be significantly higher than in mixed basal diet M. Regarding the content of utilizable crude protein at the duodenum (uCP), which is influenced by the energy content of the feed, the reverse was true. Regarding carbohydrate composition, only neutral detergent fiber (NDF) content was significantly higher in mixed basal diet R, whereas starch content was considerably and significantly higher in mixed basal diet M. Owing to the third cut of Italian ryegrass silage being considerably wetter than the other silages, mixed basal diet R had a significantly lower content of DM than mixed basal diet M.
Table 2. Ingredients and nutrient content of mixed basal diets, g kg−1 DM unless stated otherwise.

DM, dry matter; uCP, utilizable crude protein at the duodenum; NDF, neutral detergent fiber;
ADF, acid detergent fiber; NEL, net energy lactation.
The mixed basal diets were prepared daily and offered twice a day in amounts to ensure feed refusals of 5–10%. The herd included cows in different stages of lactation and therefore a wide range of milk yields. In order to enable cows to maintain their production level, additional concentrates were supplemented according to milk yield: Cows exceeding a daily milk yield of 15 kg received a commercial concentrate mixture (166 g crude protein, 7.79 MJ net energy for lactation (NEL) kg−1 DM; n=2) at a rate of 0.5 kg DM for each additional kg of milk yield, up to a maximum of 6 kg DM concentrates. Concentrates were fed via an automatic feeding station, and quantities were adjusted weekly. Furthermore, all cows received 50 g of a commercial mineral and vitamin mixture per day.
Data collection and analytical procedures
Throughout the experiment, milk yield and intake of additional concentrates were documented continuously, while individual intake of mixed basal diets was recorded manually only during four 6-day recording periods in weeks 3, 6, 9 and 12. For feed analysis, the following samples (each pooled over two consecutive days) were taken for each recording period: one sample of each dietary component (only during weeks 3 and 9), two samples of both freshly mixed basal diets, and one sample of each treatment's feed refusals. The feed samples were analyzed according to the German Handbook of Agricultural Experimental and Analytical Methods 16 and method numbers are given. The DM content of feed offered and feed refusals was determined by oven-drying at 105 °C (3.1). Ash, ether extracts, and sugar and starch content were analyzed using methods 8.1, 5.1.1, 7.1.1, and 7.2.1, respectively. Content of crude protein was assayed with Dumas combustion (4.1.2). NDF (6.5.1) in grass and ryegrass silages was determined without a heat stable amylase, while for analysis in maize silage and mixed basal diets alpha-amylase was added. Both NDF and acid detergent fiber (ADF, 6.5.2) are expressed inclusive of residual ash. Lignin was determined by solubilization of cellulose with sulfuric acid (6.5.3). Content of NEL and utilizable crude protein at the duodenum (uCPReference Lebzien and Voigt 17 ) were calculated according to GfE 18 .
A total of 16 milk samples (each pooled over two consecutive milkings) per cow were taken and conserved with Bronopol® until analysis for fat, protein, lactose and urea concentrations were conducted. An additional set of individual milk samples from the 19 cows that were included in the whole trial was taken for analysis of milk fatty acid profile during weeks 6 and 12. These samples were stored at −20 °C until gas chromatographic analysis of fatty acid composition was performed at the laboratory and quality service center Muva, Kempten, Germany. Once a week, cows’ live weights were determined by weighing them twice immediately after milking and calculating the mean. Nutrient balances and efficiency calculations were computed according to GfE 18 .
Statistical analysis
For statistical analysis, the program package SAS 19 was used. The nutrient content of the mixed basal diets was analyzed using proc GLM, with a model including only the fixed effect of treatment. All other data were analyzed using proc MIXED, using the following model:
 $$\eqalign{Y_{{\rm klmnopqr}}\! = \tab {\rm \mu} + {\rm treatment}_{\rm k} + {\rm day}_{\rm l} + {\rm order}_{\rm m} + {\rm LN}_{\rm n} \cr & + {\rm LS}_{\rm o} + {\rm ECM}_{\rm p} + {\rm LW}_{\rm q} {\rm} + {\rm treatment} \times {\rm LN} \cr & + {\rm treatment} \times {\rm LS} + {\rm cow (order)}_{{\rm rm}} + {\rm \varepsilon} _{{\rm klmnopqrs}}} $$
$$\eqalign{Y_{{\rm klmnopqr}}\! = \tab {\rm \mu} + {\rm treatment}_{\rm k} + {\rm day}_{\rm l} + {\rm order}_{\rm m} + {\rm LN}_{\rm n} \cr & + {\rm LS}_{\rm o} + {\rm ECM}_{\rm p} + {\rm LW}_{\rm q} {\rm} + {\rm treatment} \times {\rm LN} \cr & + {\rm treatment} \times {\rm LS} + {\rm cow (order)}_{{\rm rm}} + {\rm \varepsilon} _{{\rm klmnopqrs}}} $$
where Y=variable studied; μ=overall mean; treatment=fixed effect of treatment k (M, R); day=fixed effect of day l in the feeding trial (1, 2, … , 83); order=fixed effect of order m (1, 2); LN=fixed effect of number of lactation n (1,2, …, 6); LS=fixed effect of stage of lactation o, expressed as thirds (1st and 2nd 100 days, >200 days in milk); ECM=continuous effect of daily ECM yield p of cow r (only for parameters of feed intake); LW=continuous effect of live weight q of cow r (only for parameters of feed intake and milk fat, protein and lactose content); treatment×LN=interaction between treatment k and lactation number n; treatment×LS=interaction between treatment k and stage of lactation o; cow(order)=random effect of cow r within order m; ε=random error.
Except for the main effects of treatment and day, fixed effects and interactions which were not statistically significant were excluded from the individual models. This did not result in any changes in the interpretation of results. Tables 3 and 4 include information on significant interactions, which have a marked influence on the interpretation of results. Seven covariance structures possibly suitable for the data at hand were tested: unstructured (UN), antedependence (ANTE[1]), compound symmetry (CS), heterogeneous compound symmetry (CSH), spatial power law (SP[POW]), spatial Gaussian (SP[GAU]), and spatial spherical (SP[SPH]). The covariance structure with the Bayesian information criterion value closest to zero, which indicates the best fit of the model to the data, was selectedReference Wang and Goonewardene 20 . For analysis of milk fatty acids, only the covariance structure CS was used. Statistical differences were considered to be significant when P<0.05. Table 1 shows the arithmetic means of the nutrients content and fermentation traits of the feeds, Table 2 shows the least-squares means of the nutrient content of mixed basal diets, the pertaining probability values (P value) and the coefficients of determination (R 2). Tables 3–5 show least-squares means for treatment, the residual standard deviation (SD), the P values, and the covariance structure used (CovS).
Table 3. Daily intake of DM, nutrients and energy of organically managed dairy cows fed diets containing either maize silage or Italian ryegrass silage.
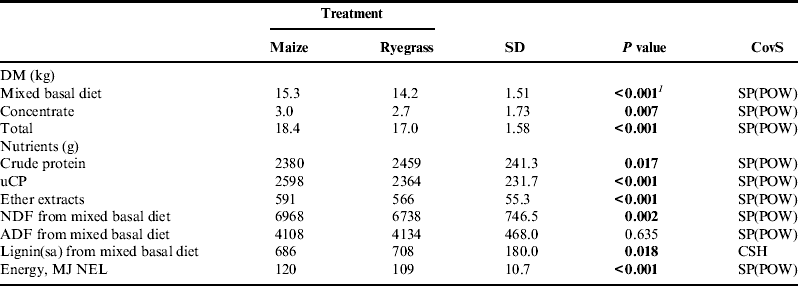
uCP, utilizable crude protein at the duodenum; NDF, neutral detergent fiber;
ADF, acid detergent fiber; NEL, net energy lactation.
CovS, covariance structure: CSH, heterogeneous compound symmetry; SP(POW), spatial power law.
1 P value of interaction treatment×number of lactations=0.008.
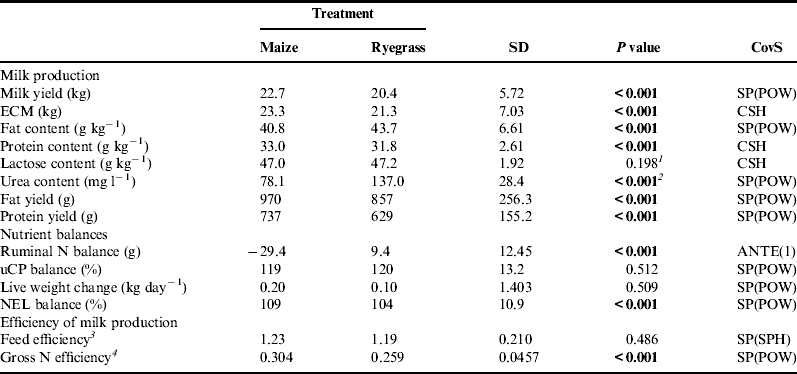
Table 4. Milk production traits, nutrient balances and efficiency of milk production of organically managed dairy cows fed diets containing either maize silage or Italian ryegrass silage.

ECM, energy corrected milk; uCP, utilizable crude protein at the duodenum;
NEL, net energy lactation; N, nitrogen;
CovS, covariance structure: ANTE(1), first-order antedependence structure;
CSH, heterogenous compound symmetry; SP(POW), spatial power law; SP(SPH), spatial spherical.
1 P value of interaction treatment×stage of lactation=0.004.
2 P value of interaction treatment×stage of lactation=0.002 and treatment×number of lactation=0.005.
3 g ECM kg−1 total intake of DM.
4 g N milk kg−1 N consumed.
Results
Feed and nutrient intake
Feeding the maize diet resulted in a significantly higher intake of mixed basal diet (+1.1 kg DM) and also of total energy (+11 MJ NEL, see Table 3). Even though a significant interaction between treatment and number of lactation was observed, treatment M led to an increase in every lactation. Feeding the ryegrass diet significantly increased crude protein intake by 3.3%, but due to the significantly higher energy content of mixed basal diet M, intake of uCP was 10% higher when feeding the maize diet. Although the mixed basal diet R contained significantly more NDF than the mixed basal diet M, intake of NDF was significantly higher (+3.4%) in treatment M due to the resulting higher intake of DM. Lignin determined by solubilization of cellulose with sulfuric acid [Lignin (sa)] intake was 3.2% higher in cows fed the ryegrass diet, whereas intake of ether extracts was significantly higher (+4.4%) in treatment M.
Milk production, nutrient balances, and feed efficiency
Feeding the maize diet significantly increased milk yield (+2.3 kg) as well as milk protein content (+1.2 g kg−1), whereas milk fat content was significantly lower (2.9 g kg−1, see Table 4). Despite lower milk fat content, both milk fat yield (+13.2%) and milk protein yield (+17.2%) were significantly higher in treatment M. Milk urea content was significantly lower when the maize diet was fed. Even though significant interactions between treatment and number of lactation and treatment and stage of lactation were observed, feeding the maize diet always decreased milk urea content. On average, cows fed the maize diet produced milk with a urea content of 78 mg l−1, whereas cows fed the ryegrass diet had a milk urea content of 137 mg l−1.
Nutrient balances and calculated parameters of production efficiency are given in Table 4. The uCP and NEL balances were positive in both treatments, but only NEL balance was significantly higher (+5 percentage points) in treatment M. Live weight change was slightly positive as well and did not differ. Ruminal N balance was significantly lower when the maize diet was fed. Feeding the ryegrass diet resulted in a gross N efficiency of 0.259, whereas feeding the maize diet resulted in a significantly higher gross N efficiency of 0.304.
Milk fatty acid profile
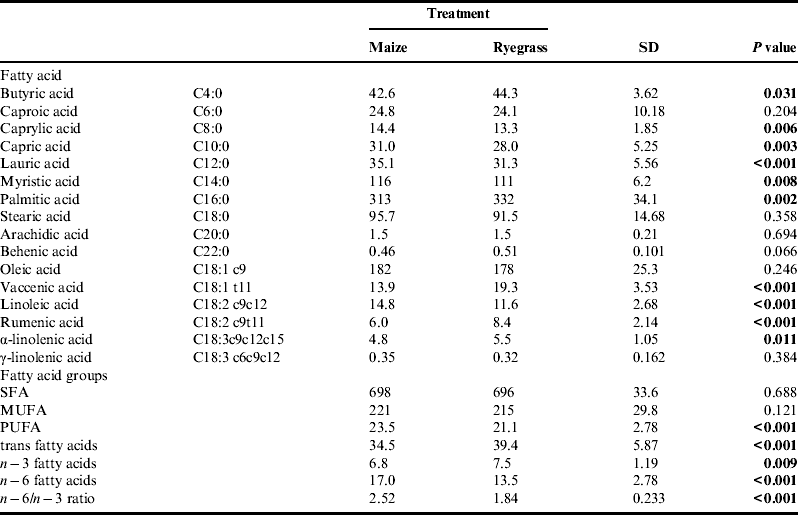
The effect of dietary treatment on milk fatty acid profile is summarized in Table 5. Of the main milk fatty acids palmitic, stearic, oleic and myristic acids, which together comprised approximately 700 g kg−1 total fat, only palmitic and myristic acids were affected by treatment. While myristic acid was significantly higher when the maize diet was fed, palmitic acid was significantly lower. Feeding the maize diet also resulted in a higher proportion of the de novo fatty acids C8:0-C14:0, as well as a higher proportion of linoleic acid, n−6 fatty acids and consequently a higher n−6/n−3 ratio. Proportions of vaccenic and rumenic acid and the sum of trans fatty acids were higher when the ryegrass diet was fed. Despite these differences in individual milk fatty acids, neither saturated fatty acids (SFA) nor mono-unsaturated fatty acid (MUFA) were influenced by treatment, only the poly-unsaturated fatty acids (PUFA) content was significantly increased when the maize diet was fed.
Table 5. Fatty acid profile and groups of fatty acids of milk fat (g kg−1 total fat) of organically managed dairy cows fed diets containing either maize silage or Italian ryegrass silage.
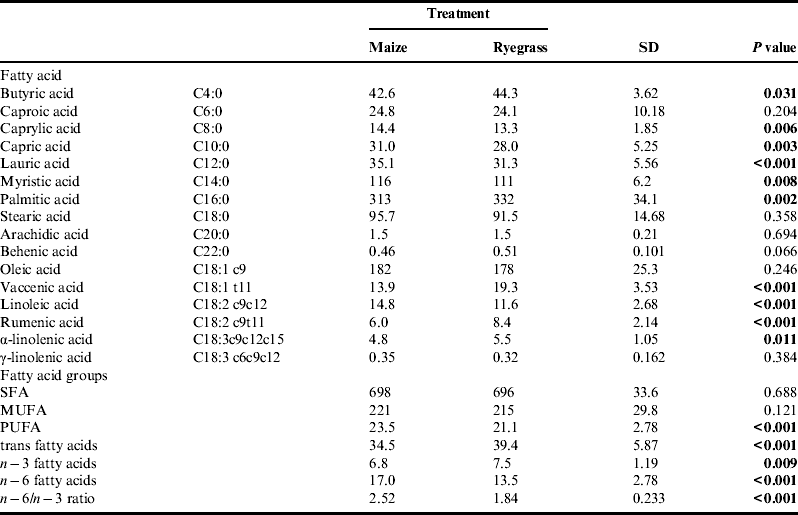
SFA, saturated fatty acids; MUFA, mono-unsaturated fatty acids; PUFA, poly-unsaturated fatty acids.
Discussion
This study was conducted to compare maize silage and Italian ryegrass silage as forage components in mixed basal diets for dairy cows under the conditions of Austrian organic agriculture. Despite the addition of adjusted amounts of barley and soybean cake to achieve isonitrogenous mixed basal diets, crude protein content was significantly higher in mixed basal diet R (including Italian ryegrass silage), while contents of uCP were slightly, but significantly higher in mixed basal diet M (including maize silage).
Feed composition, feed and nutrient intake
The DM content, fermentation quality and energy values of the silages ranged between good and ideal, except for the third cut of Italian ryegrass (see Table 1). Ash content of all samples of both ensiled cuts of Italian ryegrass (raw data not shown) was higher than 100 g kg−1 DM, indicating some contamination with soil particles during harvest. However, only in the third cut did this lead to higher levels of butyric acid and degradation of protein to ammonia-N, probably due to the fact that the butyric-acid-producing clostridia prefer wet conditionsReference Wilkinson 21 . The unusually low crude protein content of grass silage probably resulted from low amounts of clover in the grass-clover leys that provided part of the substrate.
The general level of intake of DM (17.0–18.4 kg, see Table 3) is in accordance with the average of 18.5 kg (13.0 kg forage plus 5.5 kg concentrates) observed by Gruber et al.Reference Gruber, Schwarz, Erdin, Fischer, Spiekers, Steingaß, Meyer, Chassot, Jilg, Obermaier and Guggenberger 22 based on the analysis of an extensive database of 2151 lactating dairy cows. Important factors influencing forage intake are DM and energy content, and fermentation quality of the forageReference Huhtanen, Khalili, Nousiainen, Rinne, Jaakkola, Heikkilä and Nousiainen 23 , Reference Krizsan, Westad, Ådnøy, Odden, Aakre and Randby 24 . The ryegrass diet had the disadvantage of containing a wet, suboptimally fermented and low-energy third cut of Italian ryegrass, and consequently almost all parameters of feed intake were improved when the maize diet was fed. However, even the energy content of the well-preserved second cut of Italian ryegrass was numerically lower than the maize silage, therefore, the ryegrass diet would have had a lower energy content than the maize diet even without the added disadvantage of the suboptimally fermented third cut. When substituting parts of a maize-silage-based diet (312 g DM kg−1) with Italian ryegrass silage (264 g DM kg−1), Cooke et al.Reference Cooke, Bernard and West 25 found a significant positive relationship between the proportion of maize silage and intake of DM, which is in accordance with the findings of the present study. However, Bernard et al.Reference Bernard, West and Trammell 9 report quite the reverse. In this study, cows showed numerically but not significantly, higher intakes of DM when Italian ryegrass was the sole forage as compared to maize silage (44.5% concentrates of total feed DM).
Milk yield and composition
There are frequent reports of a positive correlation between starch intake and milk yield, while milk fat content decreasesReference Murphy and O´Mara 26 – Reference Walker, Dunshea and Doyle 28 . Since the maize diet contained significantly more starch than the ryegrass diet, the difference in starch intake explains the higher milk yield and lower milk fat content when the maize diet was fed. Feeding trials comparing maize silage and Italian ryegrass silage as forage for high-yielding dairy cows (44–54% concentrates of total feed DM) do not give a consistent picture of the effect on milk yield. Neither Calabro et al.Reference Calabro, Caligiuri, Piccolo and Infascelli 29 nor Cooke et al.Reference Cooke, Bernard and West 10 , Reference Cooke, Bernard and West 25 report an effect when comparing Italian ryegrass silage as the sole forage to a mixture with maize silage, and when comparing total mixed rations with maize silage or Italian ryegrass silage, respectively. Bernard et al.Reference Bernard, West and Trammell 9 , however, report higher intake of DM as well as higher milk yield when feeding Italian ryegrass silage as the sole forage. Apart from severe undersupply with energy, milk lactose content is hardly ever influenced by feedingReference Chalupa and Sniffen 30 ; therefore, the lack of treatment effect on milk lactose content was to be expected.
Some of the most important dietary factors influencing milk fatty acid composition are fat intake, energy balance and proportion of starch in the diet.
Intake of ether extracts was significantly higher in treatment M. Although not measured in this experiment and although variation in the content of individual fatty acids is considerable, maize oil is generally higher in linoleic acid and lower in alpha-linolenic acid than grass silageReference Khan, Cone, Fievez and Hendriks 31 . As a result, cows fed the maize diet produced milk with significantly higher content of linoleic acid, n−6 fatty acids in general, and consequently a higher n−6/ n−3 ratio. On the other hand, milk from cows fed the ryegrass diet contained significantly higher proportions of trans fatty acids in general and of vaccenic and rumenic acid specifically. Vaccenic and rumenic acid are intermediates of the biohydrogenation of dietary alpha-linolenic and linoleic acids in the rumen, and as the major conjugated linoleic acid isomers in milk fat they seem to be associated with positive effects on human healthReference Bauman, Lock, Corl, Ip, Salter, Parodi, Sejrsen, Hvelplund and Nielsen 32 . The n−6/ n−3 ratio of 1.84 observed in treatment R is in accordance with levels reported by Fall and EmanuelsonReference Fall and Emanuelson 33 for organically managed dairy cows during the indoor season, whereas the ratio of 2.52 observed in treatment M is considerably higher. In general, a lower ratio of n−6/ n−3 fatty acids is more desirable for human consumption, but still the milk produced from all cows was well below the recommendations (5:1) given by DACH 34 .
According to Walker et al.Reference Walker, Dunshea and Doyle 28 , the higher proportion of de novo fatty acids observed when the maize diet was fed can be explained by the fact that both energy intake and energy balance were significantly higher in treatment M. When substituting increasing parts of a grass silage based diet with maize silage, Kliem et al.Reference Kliem, Morgan, Humphries, Shingfield and Givens 5 found an increasing proportion of de novo fatty acids as well. Usually, increasing starch intake at the expense of fiber is known to result in higher ruminal propionate and less acetate production. However, the slowly degradable starch in maize has also been shown to increase ruminal butyrate productionReference Chilliard, Ferlay, Mansbridge and Doreau 35 , which, together with acetate, is a precursor of the de novo synthesized milk fatty acids. Therefore, the higher starch intake when the maize diet was fed had a positive effect on the proportion of the short and middle-chain fatty acids C8:0–C14:0. Also, both diets were based on grass silage and hay and therefore fiber was not a limiting factor. A higher proportion of myristic acid, but lower proportion of palmitic acid in milk fat when feeding maize silage has also been found by Nielsen et al.Reference Nielsen, Straarup, Vestergaard and Sejrsen 27 who compared maize silage and grass silage as the sole forage for dairy cows. To summarize, even though the milk fatty acid profile showed some beneficial characteristics when the ryegrass diet was fed, the overall differences were small and are not expected to substantially influence human health.
Nutrient balances and efficiency of milk production
Positive nutrient balances and live weight change indicate an adequate supply of uCP and energy for all cows (see Table 4). Cows fed the maize diet had higher uCP intakes but also higher milk protein yields, therefore no difference in uCP balance was observed. Unlike uCP balance both ruminal N balance and milk urea content were significantly lower in treatment M; while the ruminal N balance observed in treatment R (9.4 g) indicates an adequate protein supply, the negative value of −29.4 g found in treatment M can be interpreted as a protein undersupply to ruminal microbes, according to the recommendations of GfE 18 . Milk urea content is used as a tool to monitor protein nutrition of dairy cows, because it reflects the N surplus for microbial synthesis in the rumenReference Hof, Vervoorn, Lenaers and Tamminga 36 . Nousiainen et al.Reference Nousiainen, Shingfield and Huhtanen 37 even report an adequate protein supply to microbes only at 251 mg l−1 milk urea content, without regarding N-recycling via the ruminohepatic circle. Compared to these results, the values measured in the experiment in question indicate that the protein supply to ruminal microbes was scarce when the ryegrass diet was fed (average milk urea content 137 mg l−1), and very scarce when the maize diet was fed (average milk urea content 78 mg l−1). When dairy cows are not adequately supplied with protein, at least part of the protein deficit can be mobilized from body tissue, before milk yield eventually starts to decline 38 , Reference Van Soest 39 . However, the fact that the protein-scarce maize diet resulted in significantly higher milk yields and did not cause negative live weight changes allows for the conclusion that scarce protein supply to the ruminal microbes did not (yet) harm the cows and their performance.
The observed feed efficiencies of 1.19–1.23 kg ECM kg−1 intake of DM lie within the range of previous reports about organically managed dairy cows, ranging from 1.11 kg milk kg−1 intake of DM when only 1.3 kg DM of concentrates were supplementedReference Velik, Baumung, Zollitsch and Knaus 40 (forage base consisted of grass–clover silage and maize silage) to 1.35–1.37 kg ECM kg−1 intake of DM when 10 kg DM of concentrates were supplementedReference Steinshamn and Thuen 41 (forage base grass–clover silage). The gross N efficiency of 0.259 observed when feeding the ryegrass diet is similar to the level of 0.255 observed by Velik et al.Reference Velik, Baumung and Knaus 42 (forage-base grass–clover silage, supplemented with 3 kg DM of concentrates), but at the lower end of the range of 0.25–0.30 that according to ChaseReference Chase 43 can be considered as average, while the level of 0.304 achieved with the maize diet is already slightly above average. Although the biological potential of cows to convert feed N into milk N is limited, N efficiency can be improved by reducing feed N intake and thereby optimizing the use of microbial protein synthesis in the rumenReference Powell, Gourley, Rotz and Weaver 44 . Accordingly, the positive effect of the maize diet can be attributed to the fact that cows fed the maize diet had a lower intake of crude protein but a higher milk protein yield than cows fed the ryegrass diet. A positive effect of including maize silage in grass-silage-based diets on N efficiency has also been found by Givens and RulquinReference Givens and Rulquin 45 , who reviewed numerous feeding trials. They attributed the effect to a better energy supply to the ruminal microbes and therefore enhanced synthesis of microbial protein, whereas in the experiment in question the effect of lower crude protein intake might have been more important.
Conclusions
Including maize silage at a rate of 40% (of DM) in a grass-silage-based diet for dairy cows and the conditions of Austrian organic agriculture was found to result in a significantly higher intake of DM and milk yield than including Italian ryegrass silage. The significantly lower intake of crude protein when feeding the maize diet as compared to the ryegrass diet, in combination with a higher milk protein yield, enabled an efficiency of gross N utilization as high as 0.304 when the maize diet was fed. This was significantly and considerably higher than the level of 0.259 observed when the ryegrass diet was fed. Therefore, maize silage upholds its reputation as an ideal energy-rich component in grass-silage-based dairy cow diets, and as long as it can be easily incorporated into crop rotation, its cultivation is recommended.
Acknowledgements
Funding of this research project was provided by the Austrian Federal Ministry of Agriculture, Forestry, Environment and Water Management, Bio Austria, the Salzburger Landesregierung and Raiffeisenverband Salzburg. Furthermore, the authors want to thank the Secondary Agriculture and Forestry College at Ursprung for housing the experiment, the farm staff for their help with the experimental work, the laboratory and quality service center Muva Kempten, Germany, for analysis of milk fatty acids and Kathleen Knaus for editing assistance.