


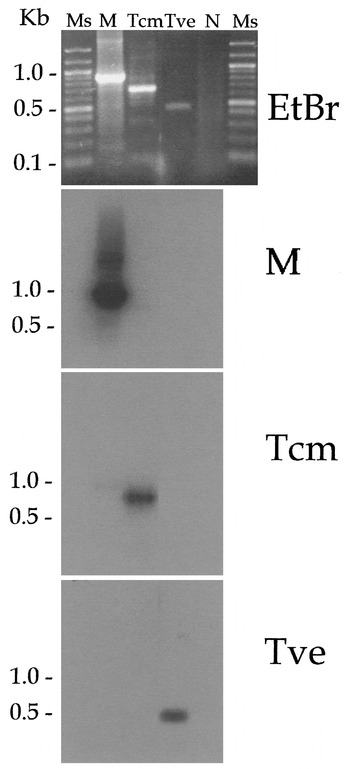

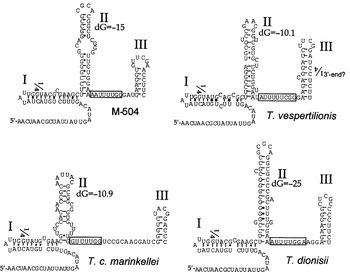
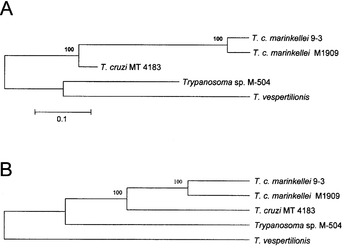
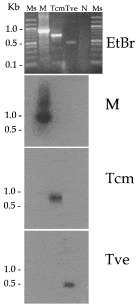


Crossref Citations
This article has been cited by the following publications. This list is generated based on data provided by Crossref.
WESTENBERGER, S. J.
STURM, N. R.
YANEGA, D.
PODLIPAEV, S. A.
ZELEDÓN, R.
CAMPBELL, D. A.
and
MASLOV, D. A.
2004.
Trypanosomatid biodiversity in Costa Rica: genotyping of parasites from Heteroptera using the spliced leader RNA gene.
Parasitology,
Vol. 129,
Issue. 5,
p.
537.
YURCHENKO, VYACHESLAV
LUKEŠ, JULIUS
XU, XING
and
MASLOV, DMITRI A.
2006.
An Integrated Morphological and Molecular Approach to a New Species Description in the Trypanosomatidae: the Case of Leptomonas podlipaevi n. sp., a Parasite of Boisea rubrolineata (Hemiptera: Rhopalidae).
Journal of Eukaryotic Microbiology,
Vol. 53,
Issue. 2,
p.
103.
Maia da Silva, F.
Marcili, A.
Lima, L.
Cavazzana, M.
Ortiz, P.A.
Campaner, M.
Takeda, G.F.
Paiva, F.
Nunes, V.L.B.
Camargo, E.P.
and
Teixeira, M.M.G.
2009.
Trypanosoma rangeli isolates of bats from Central Brazil: Genotyping and phylogenetic analysis enable description of a new lineage using spliced-leader gene sequences.
Acta Tropica,
Vol. 109,
Issue. 3,
p.
199.
COTTONTAIL, V. M.
WELLINGHAUSEN, N.
and
KALKO, E. K. V.
2009.
Habitat fragmentation and haemoparasites in the common fruit bat,Artibeus jamaicensis(Phyllostomidae) in a tropical lowland forest in Panamá.
Parasitology,
Vol. 136,
Issue. 10,
p.
1133.
Lima, Luciana
Silva, Flávia Maia da
Neves, Luis
Attias, Márcia
Takata, Carmen S.A.
Campaner, Marta
de Souza, Wanderley
Hamilton, Patrick B.
and
Teixeira, Marta M.G.
2012.
Evolutionary Insights from Bat Trypanosomes: Morphological, Developmental and Phylogenetic Evidence of a New Species, Trypanosoma (Schizotrypanum) erneyi sp. nov., in African Bats Closely Related to Trypanosoma (Schizotrypanum) cruzi and Allied Species.
Protist,
Vol. 163,
Issue. 6,
p.
856.
Hamilton, Patrick B.
Teixeira, Marta M.G.
and
Stevens, Jamie R.
2012.
The evolution of Trypanosoma cruzi: the ‘bat seeding’ hypothesis.
Trends in Parasitology,
Vol. 28,
Issue. 4,
p.
136.
Cámara, María de los Milagros
Bouvier, León A.
Canepa, Gaspar E.
Miranda, Mariana R.
Pereira, Claudio A.
and
Buscaglia, Carlos A.
2013.
Molecular and Functional Characterization of a Trypanosoma cruzi Nuclear Adenylate Kinase Isoform.
PLoS Neglected Tropical Diseases,
Vol. 7,
Issue. 2,
p.
e2044.
Votýpka, Jan
Suková, Eva
Kraeva, Natalya
Ishemgulova, Aygul
Duží, Ivana
Lukeš, Julius
and
Yurchenko, Vyacheslav
2013.
Diversity of Trypanosomatids (Kinetoplastea: Trypanosomatidae) Parasitizing Fleas (Insecta: Siphonaptera) and Description of a New Genus Blechomonas gen. n..
Protist,
Vol. 164,
Issue. 6,
p.
763.
Sun, Hongchao
Wang, Yiyin
Zhang, Yingguang
Ge, Wei
Zhang, Fuqiang
He, Biao
Li, Zuosheng
Fan, Quanshui
Wang, Wei
Tu, Changchun
Li, Jiping
and
Liu, Quan
2013.
Prevalence and Genetic Characterization of Toxoplasma gondii in Bats in Myanmar.
Applied and Environmental Microbiology,
Vol. 79,
Issue. 11,
p.
3526.
Lima, Luciana
Espinosa-Álvarez, Oneida
Hamilton, Patrick B
Neves, Luis
Takata, Carmen SA
Campaner, Marta
Attias, Márcia
de Souza, Wanderley
Camargo, Erney P
and
Teixeira, Marta MG
2013.
Trypanosoma livingstonei: a new species from African bats supports the bat seeding hypothesis for the Trypanosoma cruzi clade.
Parasites & Vectors,
Vol. 6,
Issue. 1,
Austen, Jill M.
O'Dea, Mark
Jackson, Bethany
and
Ryan, Una
2015.
High prevalence of Trypanosoma vegrandis in bats from Western Australia.
Veterinary Parasitology,
Vol. 214,
Issue. 3-4,
p.
342.
Barbosa, Amanda D.
Mackie, John T.
Stenner, Robyn
Gillett, Amber
Irwin, Peter
and
Ryan, Una
2016.
Trypanosoma teixeirae: A new species belonging to the T. cruzi clade causing trypanosomosis in an Australian little red flying fox (Pteropus scapulatus).
Veterinary Parasitology,
Vol. 223,
Issue. ,
p.
214.
da Costa, Andréa P.
Nunes, Pablo Henrique
Leite, Beatriz Helena Santos
Ferreira, Juliana Isabel G. da S.
Tonhosolo, Renata
da Rosa, Adriana Ruckert
da Rocha, Patricio Adriano
Aires, Caroline Cotrim
Gennari, Solange Maria
and
Marcili, Arlei
2016.
Diversity of bats trypanosomes in hydroeletric area of Belo Monte in Brazilian Amazonia.
Acta Tropica,
Vol. 164,
Issue. ,
p.
185.
Szpeiter, Bruno Bernal
Ferreira, Juliana Isabel Giuli da Silva
Assis, Francisco Flávio Vieira de
Stelmachtchuk, Felipe Nascimento
Peixoto Junior, Kleber da Cunha
Ajzenberg, Daniel
Minervino, Antonio Humberto Hammad
Gennari, Solange Maria
and
Marcili, Arlei
2017.
Bat trypanosomes from Tapajós-Arapiuns Extractive Reserve in Brazilian Amazon.
Revista Brasileira de Parasitologia Veterinária,
Vol. 26,
Issue. 2,
p.
152.
2017.
Bats and Human Health.
p.
285.
Bento, Elaine Cristina
Gómez-Hernández, César
Batista, Lara Rocha
Anversa, Laís
Pedrosa, André Luiz
Lages-Silva, Eliane
Ramírez, Juan David
and
Ramirez, Luis Eduardo
2018.
Identification of bat trypanosomes from Minas Gerais state, Brazil, based on 18S rDNA and Cathepsin-L-like targets.
Parasitology Research,
Vol. 117,
Issue. 3,
p.
737.
Barros, Juliana Helena da Silva
Roque, André Luiz Rodrigues
Xavier, Samanta Cristina das Chagas
Nascimento, Kátia Cristina Silva
Toma, Helena Keiko
and
Madeira, Maria de Fatima
2020.
Biological and Genetic Heterogeneity in Trypanosoma dionisii Isolates from Hematophagous and Insectivorous Bats.
Pathogens,
Vol. 9,
Issue. 9,
p.
736.
Austen, Jill M.
and
Barbosa, Amanda D.
2021.
Diversity and Epidemiology of Bat Trypanosomes: A One Health Perspective.
Pathogens,
Vol. 10,
Issue. 9,
p.
1148.
Sato, Hiroshi
and
Mafie, Eliakunda
2022.
“Visiting old, learn new”: taxonomical overview of chiropteran trypanosomes from the morphology to the genes.
Parasitology Research,
Vol. 121,
Issue. 3,
p.
805.
Torres, Jaire Marinho
de Oliveira, Carina Elisei
Santos, Filipe Martins
Sano, Nayara Yoshie
Martinez, Érica Verneque
Alves, Fernanda Moreira
Tavares, Luiz Eduardo Roland
Roque, André Luiz Rodrigues
Jansen, Ana Maria
and
Herrera, Heitor Miraglia
2024.
Trypanosomatid diversity in a bat community of an urban area in Campo Grande, Mato Grosso do Sul, Brazil.
Infection, Genetics and Evolution,
Vol. 118,
Issue. ,
p.
105563.