Introduction
As currently understood, vetulicolians are a group of exclusively marine, enigmatic, and extinct Cambrian animals represented by three families: the Vetulicolidae, the Didazoonidae, and the Banffidae (Aldridge et al., Reference Aldridge, Hou, Siveter, Siveter and Gabbott2007). They are characterized by a bipartite body. The anterior part is covered by a structure that superficially resembles the carapace of many arthropods. This structure, here referred to as the anterior section (Fig. 1), contains five lateral pouches, some possessing a lateral groove on each side, that have been interpreted by some as possible gill slits (Shu et al., Reference Shu, Conway Morris, Han, Chen, Zhang, Zhang, Liu, Li and Liu2001). The posterior section resembles the arthropod trunk, possessing a segmented and sometimes annulated morphology. Soft-part anatomy has been reported in some vetulicolians, including structures interpreted to represent muscle fibers (Aldridge et al., Reference Aldridge, Hou, Siveter, Siveter and Gabbott2007; Ou et al., Reference Ou, Conway Morris, Han, Zhang, Liu, Chen, Zhang and Shu2012; García-Bellido et al., Reference García-Bellido, Lee, Edgecombe, Jago, Gehling and Paterson2014). The vetulicolian body plan and the limited information about soft-bodied anatomical structures within the anterior section make the interpretation of these animals difficult. As a result, the phylogenetic position of vetulicolians is unclear, with proposed affinities much debated and varying from their interpretation as unusual arthropods (Hou, Reference Hou1987; Caron, Reference Caron2006), kinorhynchs (Aldridge et al., Reference Aldridge, Hou, Siveter, Siveter and Gabbott2007), or stem-group deuterostomes (Shu et al., Reference Shu, Conway Morris, Han, Chen, Zhang, Zhang, Liu, Li and Liu2001; Ou et al., Reference Ou, Conway Morris, Han, Zhang, Liu, Chen, Zhang and Shu2012) or chordates (Lacalli, Reference Lacalli2002; García-Bellido et al., Reference García-Bellido, Lee, Edgecombe, Jago, Gehling and Paterson2014).

Figure 1 A hypothetical vetulicolian, showing detailed morphological characters. Abbreviations are as follows (terms used by other authors are in parentheses). Ac=alimentary canal (gut); An=anus; Ao=anterior opening (oral opening, oral, mouth, head); Ar=articulation; As=anterior section (anterior body, carapace, head+trunk), presumed cover of the anterior part of the body; Dp=posterodorsal projection (fin-like structure, dorsal fin, fin); Ds=dorsal section; Fs=filamentous structures (gill filaments); Lg=lateral groove, a lateral midline that consists of a longitudinal groove associated with five openings at each side of the anterior section; Lp=lateral pouches/openings (gill pouches, serial lateral opening structure); Ls=lateral slit (gill slit); M=membrane; Ps=posterior section (posterior body, tail, abdomen), posterior part of the body; S=segment; S1–S7=first through seventh segment of posterior section; Vp=posteroventral projection of anterior section (fin-like structure, ventral fin); Vs=ventral section.
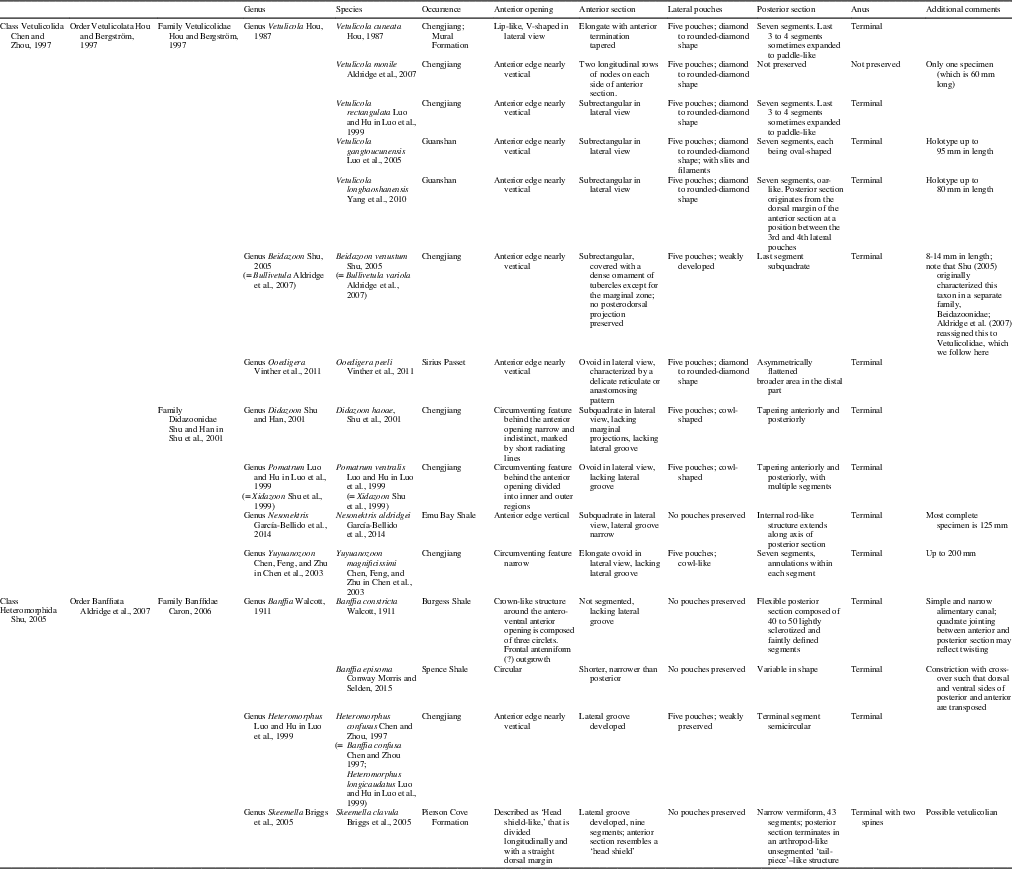
Vetulicolians, as currently understood, are represented by 15 species, which occur worldwide but only in Cambrian Lagerstätten, including the Chengjiang Biota (Hou, Reference Hou1987; Luo et al., Reference Luo, Hu, Chen, Zhang and Tao1999; Shu et al., Reference Shu, Conway Morris, Han, Chen, Zhang, Zhang, Liu, Li and Liu2001; Chen et al., Reference Chen, Feng, Zhu, Ma and Li2003; Shu, Reference Shu2005; Aldridge et al., Reference Aldridge, Hou, Siveter, Siveter and Gabbott2007), the Guanshan Biota (Luo et al., Reference Luo, Fu, Hu, Li, Chen, You and Liu2005; Yang et al., Reference Yang, Hou, Cong, Dong, Zhang and Luo2010; Li et al., Reference Li, Cong, Zhao and Hou2015), and the Shipai Formation of South China (Zhang and Hua, Reference Zhang and Hua2005); the Burgess Shale (Walcott, Reference Walcott1911) and the Mural Formation, both in Canada (Butterfield, Reference Butterfield2005); the Sirius Passet Biota of Greenland (Vinther et al., Reference Vinther, Smith and Harper2011); the Emu Bay Shale of Australia (García-Bellido et al., Reference García-Bellido, Lee, Edgecombe, Jago, Gehling and Paterson2014); and the Spence Shale in Utah, USA (Briggs et al., Reference Briggs, Lieberman, Halgedahl and Jarrard2005; Conway Morris et al., Reference Conway Morris, Halgedahl, Selden and Jarrard2015). They are known almost exclusively from their characteristic anterior section, which possesses lateral pouches, and segmented posterior section. Despite being widespread and common animals throughout Cambrian Lagerstätten, the affinity of vetulicolians remains controversial.
Yuyuanozoon magnificissimi Chen, Feng, and Zhu in Chen et al., Reference Chen, Feng, Zhu, Ma and Li2003 is exceptionally large, being up to 20 cm in length, compared with other vetulicolian species, which are generally 5–14 cm long. Thus far, it has been described from a single complete specimen, but here we show by reference to new material that some of the characteristics of this original specimen have been modified postmortem, and as such its current appearance does not accurately represent in vivo morphology. Here we describe new specimens, provide a new interpretation for the opening at the presumed anterior end of the animal, and reassign this taxon to the vetulicolian family Didazoonidae.
Geologic setting
The early Cambrian Chengjiang Lagerstätte (Cambrian Series 2, Stage 3, Eoredlichia-Wutingaspis trilobite Biozone of Eastern Yunnan, a biostratigraphical interval equivalent to part of the Ushbaspis trilobite Zone of South China) occurs through a 30–50 m thick succession of claystones that have been celebrated for yielding about 250 species of soft-bodied and weakly biomineralized organisms (Hou et al., Reference Hou, Siveter, Siveter, Aldridge, Cong, Gabbott, Ma, Purnell and Williams2017). The dominant component of the biota, both in numerical abundance and species diversity, is arthropods, but lobopodians, sponges, brachiopods, some enigmatic animals and chordates are also represented (Hou et al., Reference Hou, Siveter, Siveter, Aldridge, Cong, Gabbott, Ma, Purnell and Williams2017). Other significant animals are the vetulicolians, of which the Chengjiang Biota contains the greatest diversity and numbers from the Cambrian System worldwide.
The three new specimens of the vetulicolian Yuyuanozoon magnificissimi described here are from the Ercaicun section at Haikou, Kunming area, Yunnan Province, about 30 km south of Kunming city and about 50 km northwest of Maotianshan, the original locality from which Chengjiang fossils were discovered (see Hou et al., Reference Hou, Siveter, Siveter, Aldridge, Cong, Gabbott, Ma, Purnell and Williams2017 for the history of the Chengjiang Biota).
Materials and methods
The specimens were prepared using steel needles and examined with a Nikon SMZ–10A binocular microscope. We used a camera lucida attached to a Wild Heerbrug M10 microscope to elucidate and record the anatomy. The digital images of the specimens were captured with Nikon D3X and Canon 500D digital cameras and a Leica DFC5000 camera attached to a Leica M205C photomicroscope and were processed in Adobe Photoshop CC. All measurements were processed with ImageJ version 1.49.
Repository and institutional abbreviation
The three new specimens documented here are preserved with part and counterpart and are deposited in the Yunnan Key Laboratory for Palaeobiology, Yunnan University (abbreviated YKLP), Kunming, China, with numbers YKLP 13070, YKLP 13071, and YKLP 13072.
Morphology and terminology of the vetulicolian body
The anatomical terms applied to vetulicolians are complicated because there is no consensus on a common nomenclature in different publications. Vetulicolian taxonomy sometimes uses a range of standard morphological terms such as ‘carapace’ and ‘gill slits’ (see Table 1), though there is no certainty of the homology of such structures between taxa, or even whether these terms, some for example borrowed from arthropod terminology (e.g., for the genus Skeemella), are appropriate.
Table 1 Vetulicolian species and their major morphological characters. Vetulicola sp. from the Shipai Formation of South China (Zhang and Hua, Reference Zhang and Hua2005), Didazoon sp. from the Chengjiang Biota (Aldridge et al., Reference Aldridge, Hou, Siveter, Siveter and Gabbott2007), and Vetulicola sp. from the Guanshan Biota (Li et al., Reference Li, Cong, Zhao and Hou2015) are not included in the table because their full systematic relationships have not been discerned.

Here we use descriptive terms carefully, to avoid inference about evolutionary significance, thus avoiding the potential for circular reasoning regarding phylogenetic interpretation. For example, we do not use the term ‘carapace’ to denote the anterior section of the animal. Our morphological terms are summarized in Figures 1 and 2, which also denote how terms such as ‘ventral’ and ‘dorsal’ are applied to vetulicolians.

Figure 2 Additional morphological features for Yuyuanozoon. Abbreviations are as follows (terms used by other authors are in parentheses). Cv=circumventing feature behind the anterior opening (oral disc.); Mr=marginal region at the anterior opening (mouth plate); Ol=ordering lines (putative segments, anterior segments), lines circumventing the anterior section coalescing with lateral pouches and long axis perpendicular to the body axis.
Preservation and taphonomy
Specimen YKLP 13071 (Fig. 3) is, like most vetulicolians, laterally compressed and retains some three-dimensional (3-D) relief. The majority of the anterior section is preserved as a thin film that does not appear to have been biomineralized though the areas adjacent to the anterior opening and the posterior end show patches of dark color. The texture and composition of this material is identical to the iron oxide coatings typical in Chengjiang fossils (Gabbott et al., Reference Gabbott, Hou, Norry and Siveter2004; Forchielli et al., Reference Forchielli, Steiner, Kasbohm, Hu and Keupp2014) and is not consistent with in vivo mineralization. The anterior section appears to comprise six subdivisions that are demarcated by five lines that circumvent the entire structure and are coincident with the five lateral cowl-shaped pouches. At the midpoint between the boundaries of these subdivisions on the dorsal side a shorter parallel line occurs, which was termed second-order annulation by Aldridge et al. (Reference Aldridge, Hou, Siveter, Siveter and Gabbott2007). There are clear wrinkles along the ventral side of the anterior section, which appear to suggest an element of twisting and/or compression postmortem.
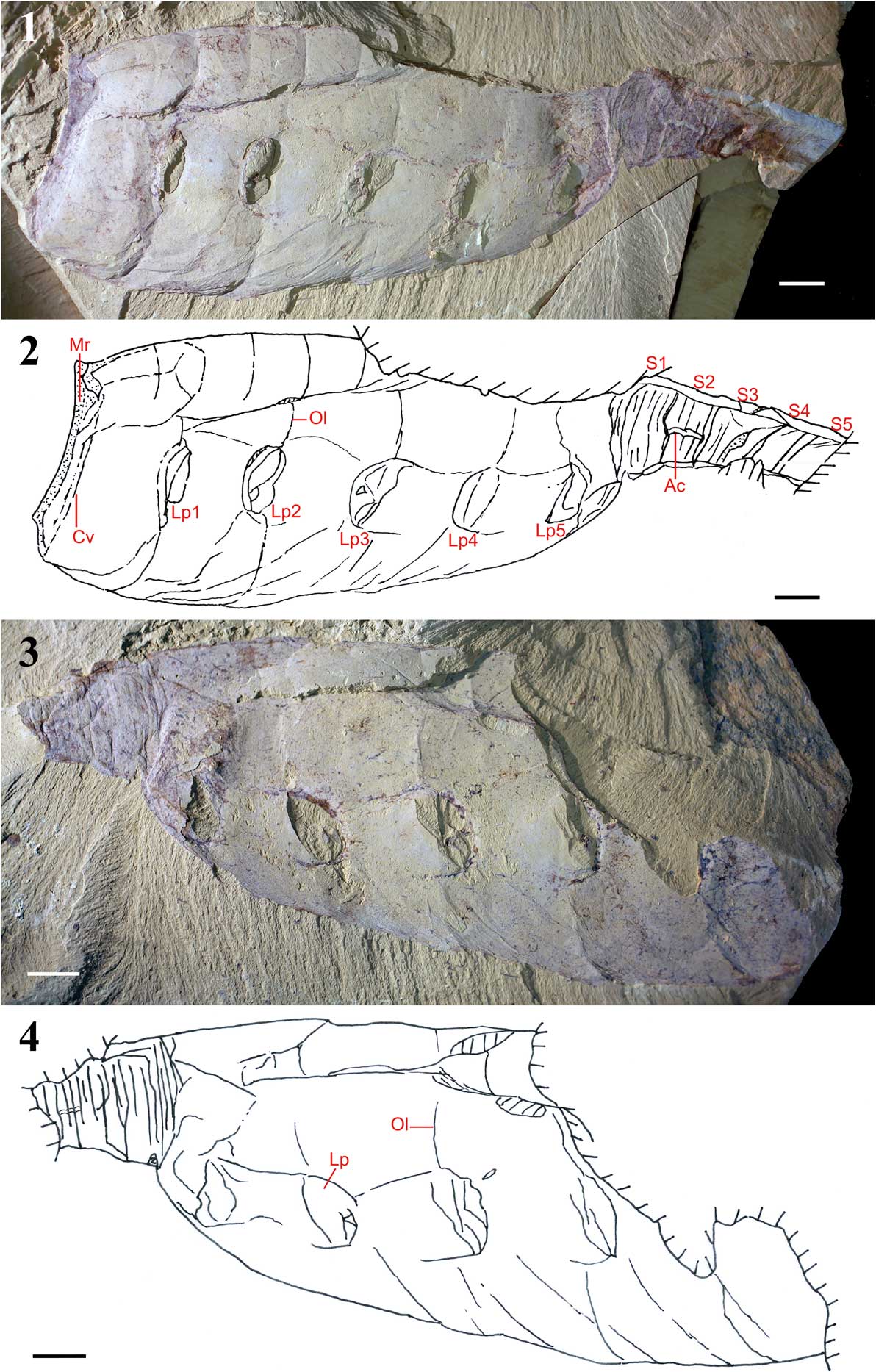
Figure 3 (1, 2) Lateral view of Yuyuanozoon magnificissimi YKLP 13071, anterior to left, dorsal to top. (1) Photograph of the fossil (largely the internal mold) with low-angle lighting; (2) camera lucida drawing of the same specimen. The six subdivisions of the anterior section are evident, as are the occurrence of secondary lines between these subdivisions. The ‘segmentation’ and ‘annulation’ of the posterior part of the body (which is incomplete posteriorly) are also evident. Ac=alimentary canal; Cv=circumventing feature behind the anterior opening; Lp1–Lp5=first through fifth lateral pouch; Mr=marginal region at the anterior opening; Ol=ordering lines; S1–S5=putative segments. (3, 4) Lateral view of anteriorly incomplete Yuyuanozoon magnificissimi YKLP 13071, anterior to right, dorsal to top. (3) Photograph of counterpart YKLP 13071 with low-angle lighting; (4) camera lucida drawing of counterpart. Lp=lateral pouches; Ol=the ordering lines. Scale bars=10 mm.
The boundary between anterior subdivisions five and six also coincides with a marked change in relief, with the sediment fill within the anterior section thicker anteriorly and the color darker posteriorly.
The posterior section of the holotype (CFM00059) possesses seven segments; within each segment there are five, or possibly six, less well-defined lines that appear to circumvent the whole structure and thus resemble annulation. Other specimens, which are incomplete, preserve five (YKLP 13071) or six (YKLP 13070) posterior segments.
Systematic paleontology
Class Vetulicolida Chen and Zhou, Reference Chen and Zhou1997 Order Vetulicolata Hou and Berström, 1997 Family Didazoonidae Shu and Han in Shu et al., Reference Shu, Conway Morris, Han, Chen, Zhang, Zhang, Liu, Li and Liu2001
Type Genus
Didazoon Shu and Han in Shu et al., Reference Shu, Conway Morris, Han, Chen, Zhang, Zhang, Liu, Li and Liu2001. Other genera.—Pomatrum Luo and Hu in Luo et al., Reference Luo, Hu, Chen, Zhang and Tao1999; Yuyuanozoon Chen, Feng, and Zhu in Chen et al., Reference Chen, Feng, Zhu, Ma and Li2003; Nesonektris García-Bellido et al. Reference García-Bellido, Lee, Edgecombe, Jago, Gehling and Paterson2014.
Diagnosis
(Modified from Shu et al., Reference Shu, Conway Morris, Han, Chen, Zhang, Zhang, Liu, Li and Liu2001, Aldridge et al., Reference Aldridge, Hou, Siveter, Siveter and Gabbott2007) Bilaterally symmetrical animal with clear subdivision of body into presumed anterior and posterior sections. Neither the subquadrate to ovoid anterior section nor the posterior section is biomineralized. The anterior section has a large, circular opening at the presumed anterior end of the animal, posterior of which are six subdivisions demarcated by five lines; five, presumed laterally positioned, oval openings on both sides of the anterior section coincide with the lines of subdivision. Segments of posterior section may bear up to six annulations.
Remarks
In addition to characters listed by Shu et al. (Reference Shu, Conway Morris, Han, Chen, Zhang, Zhang, Liu, Li and Liu2001), in well-preserved specimens of Yuyuanozoon (herein) and Didazoon (YJL’s personal observations), the posterior section displays annulation within each segment.
Morphological features, including the circumventing feature behind the anterior opening of the anterior section, the posterodorsal (‘fin-like’) prolongations on the anterior section, the shape of the lateral pouches, and the segmented posterior section with annulations, place Yuyuanozoon in the Didazoonidae rather than in the Vetulicolidae (see also Table 1).
Genus Yuyuanozoon Chen, Feng, and Zhu in Chen et al., Reference Chen, Feng, Zhu, Ma and Li2003
Type species
Yuyuanozoon magnificissimi Chen, Feng, and Zhu in Chen et al. Reference Chen, Feng, Zhu, Ma and Li2003, by monotypy.
Diagnosis
(Modified from Chen et al., Reference Chen, Feng, Zhu, Ma and Li2003, Aldridge et al., Reference Aldridge, Hou, Siveter, Siveter and Gabbott2007) Didazoonid with nonbiomineralized, elongate ovoid anterior section. A narrow, raised rim circumvents the entire perimeter of the anterior section, some 5 mm posterior of the anterior opening. Marginal projections and lateral groove absent from anterior section. Posterior section has seven segments, within each of which are five to six annulations; terminal segment longest.
Occurrence
Early Cambrian, Yu’anshan Member, Chiungchussu Formation, Eoredlichia-Wutingaspis Biozone, Yunnan Province, South China (Chen et al., Reference Chen, Feng, Zhu, Ma and Li2003).
Remarks
Analysis of the body shape of Didazoon, Pomatrum, and Yuyuanozoon distinguishes Yuyuanozoon as a separate genus based on its overall size and the proportions of the anterior and posterior sections, which were fully described by Chen et al. (Reference Chen, Feng, Zhu, Ma and Li2003). Yuyuanozoon is also clearly distinct from other vetulicolians, with the exception of Didazoon, in possessing annulations within each segment of its posterior section.
Yuyuanozoon magnificissimi Chen, Feng, and Zhu in Chen et al., Reference Chen, Feng, Zhu, Ma and Li2003

Figure 4 Detailed views of Yuyuanozoon magnificissimi. (1) Anterior end of Y. magnificissimi YKLP 13071; (2) anterior end of Y. magnificissimi YKLP 13072; (3) anterior end of Y. magnificissimi CFM00059. Arrows show wrinkles, providing a hint that the anterior opening is not observable due to distortion through compression. (4) Anterior section view of CFM00059. Arrows show the outlines of the pouches and possible underlying narrow grooves. (5) Anterior section view of YKLP 13071. Arrows show the outlines of the pouches. (6) Posterior section view of CFM00059. Arrows show the annulations within the first segment. (7) Posterior section view of YKLP 13071 (counterpart); (8) posterior section view of YKLP 13071 (part). Scale bars=10 mm.

Figure 5 Overall views of Yuyuanozoon magnificissimi. (1) YKLP 13072 (part) anterior to left; (2) YKLP 13072 (counterpart) anterior to right; (3) YKLP 13070, posterior section with numbered segments, S2 to S7. Lp=lateral pouch; Ol=ordering lines; S=segment. Scale bars=10 mm.

Figure 6 Composite camera lucida drawing of specimen YKLP 13071, superimposing part and counterpart, anterior to left, dorsal to top. Scale bar=10 mm.
2003 Yuyuanozoon magnificissimi Chen, Feng, and Zhu in Chen et al., p. 282, pl. 1, figs. a–d.
2004 Yuyuanozoon magnificissimi; Reference ChenChen, p. 318, fig. 509.
2007 Yuyuanozoon magnificissimi; Reference Aldridge, Hou, Siveter, Siveter and GabbottAldridge et al., p. 142, pl. 5, figs. 3, 5, text-fig. 6.
2012 Yuyuanozoon magnificissimi; Reference Ou, Conway Morris, Han, Zhang, Liu, Chen, Zhang and ShuOu et al., p. 81, fig. 5.
2017 Yuyuanozoon magnificissimi; Reference Hou, Siveter, Siveter, Aldridge, Cong, Gabbott, Ma, Purnell and WilliamsHou et al., p. 276, fig. 26.3.
Holotype
CFM00059, deposited in the Chengjiang Fauna Museum, Chengjiang National Geological Park, Yunnan. The holotype was found on the northern slope of Maotianshan, Chengjiang (Chen et al., Reference Chen, Feng, Zhu, Ma and Li2003).
Other material
Three additional specimens from Ercaicun (see Hou et al., Reference Hou, Siveter, Siveter, Aldridge, Cong, Gabbott, Ma, Purnell and Williams2017, fig. 4.3): YKLP 13071, a nearly complete laterally preserved specimen; YKLP 13072, preserving most of the anterior section and part of the posterior section; and YKLP 13070, preserving most of the posterior section.
Diagnosis
Monotypic, as for the genus.
Description
The anterior section forms an elongated ellipsoid in lateral view, with five lateral pouches on each side; the anterior opening is broad (Fig. 3). The posterior section is about one-third of the width of the anterior section, elongate, segmented and annulated throughout. Narrow gut situated medially (Fig. 4.6, 4.8).
Body
All specimens are laterally preserved. The most complete specimen YKLP 13071 (Fig. 3) has a length of at least 17.2 cm (the last two segments of the posterior section are not preserved). The anterior section is 11.8 cm long and up to 5.4 cm wide; the posterior section is more than 4.5 cm in length and has a maximum width of 2.0 cm. The dorsal surface of the anterior section from the anterior opening to the posterior section is coincident along the dorsal side but has a concave outline on the ventral side (Fig. 3). The length of the posterior section measures 11 mm for the first segment and then posteriorly 9 mm, 9 mm, 9 mm, and at least 7 mm for the incomplete fifth segment.
Anterior opening
YKLP 13071 possesses a wide anterior opening (Figs. 3.1, 4.1) with a circumventing rim situated 5 mm posterior of this. In YKLP 13072, the anterior section has a more quadrate shape than is typical, and the anterior opening is broad, but the circumventing feature is less well developed (Figs. 4.2, 5.1). No anterior opening has been detected in the holotype, but strongly developed wrinkles approximately parallel to the dorsal and ventral edge provide a hint that the anterior opening is not observable, owing to distortion through compression (Fig. 4.3).
Anterior circumventing feature
Raised rim, a narrow structure circumventing the entire perimeter of the anterior section, some 5 mm posterior of the anterior opening.
Ordering lines
Lines circumventing the anterior section and perpendicular to the body axis are displayed best in the anterodorsal area, and here they comprise two orders. The most prominent lines traverse the entire body. These delineate six subdivisions of the anterior section, and their boundaries are coincident with five lateral pouches on each side of the animal (Figs. 3, 5.1, 5.2). There are faint lines between these boundaries that are less continuous in the dorsal section. This pattern is similar to Vetulicola cuneata Hou, Reference Hou1987 (Chen and Zhou, Reference Chen and Zhou1997, fig. 135) and V. rectangulata Luo and Hu in Luo et al., Reference Luo, Hu, Chen, Zhang and Tao1999 (Aldridge et al., Reference Aldridge, Hou, Siveter, Siveter and Gabbott2007, pl.1, fig. 9; text-fig. 2) and is known in Pomatrum (Aldridge et al., Reference Aldridge, Hou, Siveter, Siveter and Gabbott2007, pl. 5, fig. 1, text-fig. 8).
Lateral pouches
The lateral pouches are one of the most prominent features of all vetulicolians and number five on each side of the anterior section. In the new specimens of Y. magnificissimi, the five pouches are cowl-shaped and form a series of lateral openings (Fig. 4.5). These pouches occur in the middle region of the dorsal and ventral sections of the anterior section and are similar to those in the holotype (Fig. 4.4). The lateral groove is absent in all specimens of Y. magnificissimi. The pouches show an orientation that may have allowed water to exit the anterior section in a posteriorly flowing direction. No filaments are preserved in the new material.
Posterior segmentation
Only five of up to seven segments in the posterior part are present in YKLP 13071, and within each of these segments are five to six annulations. Only a few annulations are apparent in the holotype of Y. magnificissimi possibly due to poorer preservation (Fig. 4.6). The posterior section of YKLP 13071 is twisted and deformed (Fig. 4.7, 4.8). Each segment of the posterior section has five to six annulations; junctions between segments are distinguished according to a slight change in relief, and these boundaries are one annulation in width. Both the holotype and the new specimens have annulations. Annulations are apparent in YKLP 13070 (Fig. 5.3) and YKLP 13072 (Fig. 5.2). In the holotype, annulations are clearly present on the first segment of the posterior section and the ventral area of the sixth (Fig. 4.6).
Possible alimentary tract
The alimentary tract is preserved in the medial part of the posterior section of YKLP 13071 (Fig. 4.7–4.8). This is irregular and curved, narrow, and apparently filled with fine sediment. This structure is incomplete and only preserved at the second segment of the posterior section.
Remarks
The last subdivision of the anterior section and first few segments of the posterior section are marked by a wrinkled zone with dark coloration and lower relief than the other part of the anterior section (Fig. 3). This may be a preservation feature of the split through the part and counterpart.
The holotype was described by Chen et al. (Reference Chen, Feng, Zhu, Ma and Li2003), in which the anterior end of the anterior section was considered smooth and round without any extensions or margin. However, observations on the holotype indicate that there is a visible fold that starts from the beginning of the arched anterior end, surrounded by narrow wrinkles. This provides evidence that the anterior end was compressed, and this morphology is an artifact of decay and/or preservation. That the walls of the anterior section in members of the family Didazoonidae are generally thinner than in Vetulicolidae lends support to this interpretation of a distorted anterior opening. The anatomical differences of the lateral filamentous pouches possessed by the holotype, and the absence of these structures in the new specimens, is most likely due to differences of preservation and to the level at which the rock has split through these structures in different specimens.
Yuyuanozoon differs from other vetulicolians in the morphology of its bulky, ovoid anterior section, the shape of its anterior opening, and possession of a posterior section with seven segments that are annulated. These features suggest that Yuyuanozoon is closest in general form to Pomatrum and Didazoon.
Contrary to the comment by Chen et al. (Reference Chen, Feng, Zhu, Ma and Li2003, p. 282) that Yuyuanozoon is distinct from other vetulicolians in the number of anterior and posterior segments, the possession of six anterior section subdivisions and seven posterior segments is characteristic of both Vetulicolidae and Didazoonidae. The anterior opening of Y. magnificissimi was described as ‘long-ellipsoidal’ by Chen et al. (Reference Chen, Feng, Zhu, Ma and Li2003). Our new material shows that the anterior end of Yuyuanozoon is widely open, with a circumventing feature. Reinvestigation of the holotype indicates that the anterior section of that specimen is distorted, as evidenced by a series of wrinkles, and the morphology of the anterior opening is obscured by compression.
In the holotype, the lateral pouches and associated wrinkles have been interpreted as gill sacs and external gill filaments (Chen et al., Reference Chen, Feng, Zhu, Ma and Li2003) or as a possible vascular system (Ou et al., Reference Ou, Conway Morris, Han, Zhang, Liu, Chen, Zhang and Shu2012). Filaments have not been recognized in the new specimens of Yuyuanozoon, and it is possible that the filaments described by Chen et al. (Reference Chen, Feng, Zhu, Ma and Li2003) and Ou et al. (Reference Ou, Conway Morris, Han, Zhang, Liu, Chen, Zhang and Shu2012) are a product of decay-induced collapse forming wrinkles or the split of the fine-layered sediment, or an expression of internal anatomy.
Inferences about mode of life
By analogy with other vetulicolids, the segmented posterior section of Yuyuanozoon might be interpreted as an adaptation for propulsion. However, Yuyuanozoon has a relatively small posterior section countered by a bulky anterior section without fin-like projections that seems ill adapted for protracted forward motion.
Vetulicolians were globally widespread in early and middle Cambrian seas and are known from strata that originated in various environmental settings. All share some common features such as a lateral collapse orientation retaining a degree of three-dimensional preservation, suggesting that the body was made of a material with a degree of robustness, even if it was nonbiomineralized. How Yuyuanozoon fed is a matter of conjecture, as currently no feeding apparatus has been preserved.
Conclusions
In this paper we have described new material of Yuyuanozoon magnificissimi. We have attempted to develop a unified terminology for the description of vetulicolians that avoids inferring animal relationships or functional morphology. Therefore, terms such as ‘carapace’ (suggesting an arthropod affinity) or ‘oral end’ (suggesting functionality for feeding) have been replaced with purely descriptive terms. We believe that much of the existing terminology applied to vetulicolians cannot be sustained. There is no convincing evidence to show the homology of the anterior part of vetulicolians with the carapace or head shield of arthropods. Furthermore, there is no evidence in Y. magnificissimi to sustain the notion that the anterior opening was ‘oral’ or even that it functioned in food collection. In deconstructing this taxonomically ‘loaded’ terminology we now propose a ‘ground-up’ approach to interpreting these animals that examines their full range of morphological characteristics, without preconception. New analyses should determine: (1) whether any features are homologous with extant groups of organisms, and (2) whether any features can be accorded a true function according to basic observations of their likely adaptation for different functions. From this deconstructed database we hope that a more realistic picture of the interspecific variation of vetulicolians can be assessed, which will lead to a greater understanding of their wider animal affinities.
Acknowledgments
This work was supported by funds from the National Natural Science Foundation of China (No. U1302232, 41372031, 41572015, 41662003) and from State Key Laboratory of Palaeobiology and Stratigraphy (Nanjing Institute of Geology and Palaeontology, CAS) (No. 143101). YJL thanks the China Scholarship Council (CSC) (No. [2015]3022) for fellowship support.