


Introduction
Atla Savić & Tibell is a genus of lichens that has recently been segregated from Polyblastia A. Massal. based on molecular data (Savić & Tibell Reference Savić and Tibell2008). Morphologically the genus may be confused with Sporodictyon A. Massal., a genus resurrected in a molecular study of Savić & Tibell (Reference Savić and Tibell2009). Both genera are characterized by large muriform spores (40–120 μm long), which may be pale or dark brown, large perithecia (0·3–1·2 mm) and a northern or alpine distribution. In Sporodictyon perithecia are usually thinly or thickly covered by thalline tissue, unlike those in most species of Atla. However, two species of Atla are characterized by perithecia with a thalline cover, which does not reach the ostiolum (Savić & Tibell Reference Savić and Tibell2009; Tibell & Tibell Reference Tibell and Tibell2015). In some species of Sporodictyon cephalodia are frequent, whereas cephalodia have not previously been reported from Atla.
Only six species of Atla are currently known. Two of them, A. wheldonii (Travis) Savić & Tibell and A. alpina Savić & Tibell, are fairly widely distributed in Europe, although they mainly occur in mountain areas (Savić & Tibell Reference Savić and Tibell2008; Hafellner Reference Hafellner2010). Two species (A. palicei Savić & Tibell and A. praetermissa Savić & Tibell) were described from Scandinavia by Savić & Tibell (Reference Savić and Tibell2008). Recently, Tibell & Tibell (Reference Tibell and Tibell2015) described the new species A. alaskana S. Tibell & Tibell from Alaska and A. recondita S. Tibell & Tibell from Sweden. All the known species of Atla occur in calcareous habitats: A. wheldonii is restricted to calcareous soil, while the remaining five species are found on calcareous or calciferous rocks.
During field studies of lichens of calcareous rocks in Finland, the first author found some specimens which, based on morphology, represented Atla or Sporodictyon, but which did not conform to any known species. In this paper we describe three new species of Atla based on morphology and ITS sequences: A. oulankaensis, A. tibelliorum and A. vitikainenii.
Material and Methods
Fieldwork
Specimens of Atla and Sporodictyon were searched for during a large-scale field study of lichens of calcareous rocks and lime quarries in Finland. The sampling was most intensive in southern Finland, but only one species belonging to Atla or Sporodictyon was found: A. wheldonii, new to Finland from two neighbouring localities (Pykälä Reference Pykälä2013). Field studies in the two largest areas of calcareous rocks in Finland, occurring in the northern part of the country, were more successful: Kilpisjärvi in the biogeographical province Enontekiön Lappi (EnL) and the Kuusamo-Salla area in the biogeographical province Koillismaa (Ks). Several dozen specimens of Atla and Sporodictyon were found, most of them belonging to Atla alpina and Sporodictyon schaererianum A. Massal. Five unidentified specimens were sequenced in this study. In addition, the status of one specimen representing A. palicei based on morphological characters (see Pykälä Reference Pykälä2013) was examined with molecular data.
Morphology
Perithecia and thalli were hand-sectioned with a razor blade. The sections were examined and measured in water. Asci and ascospores were also studied in squash preparations of perithecia mounted in water. Additionally, involucrellum characters and exciple colour and width were studied by cutting perithecia into two pieces and studying the pieces using a dissecting microscope.
Spore size is indicated as a range with the arithmetic mean in bold. Minimum and maximum values are given in parentheses. The size of the perithecia is given in surface view.
DNA extraction and sequencing
Total genomic DNA was extracted from perithecia (1–3) of two to four year-old herbarium specimens. All samples were placed in 96-well microplates and sent to the Canadian Centre for DNA Barcoding (CCDB). CCDB’s standard protocols (documentation available at http://ccdb.ca/resources.php) were used for extraction, PCR and sequencing. Primers ITS1-LM (Myllys et al. Reference Myllys, Lohtander, Källersjö and Tehler1999) and ITS4 (Gardes & Bruns Reference Gardes and Bruns1993) were used both for PCR and sequencing of the nuclear ribosomal ITS barcode region. The barcode sequences and their trace files along with all relevant collection data were uploaded to the Barcode of Life Data Systems (BOLD, http://www.boldsystems.org) database. The sequences are available in GenBank.
Phylogenetic analyses
We examined phylogenetic relationships of the new species using ITS sequence data. In our analysis we used our newly produced sequences, together with all Atla sequences and selected sequences of Sporodictyon, Henrica B. de Lesd., Verrucaria muralis Ach. and V. rupestris Schrad. obtained from GenBank. All these belong to the so-called “E clade” of the “Polyblastia group” as defined by Savić et al. (Reference Savić, Tibell, Gueidan and Lutzoni2008). Additionally we included sequences obtained from soil samples, which based on a BLAST search (Altschul et al. Reference Altschul, Gish, Miller, Myers and Lipman1990) were closely related to Atla (Timling et al. Reference Timling, Walker, Nusbaum, Lennon and Taylor2014). Thelidium umbilicatum Th. Fr., Polyblastia aurorae Savić & Tibell and P. borealis Savić & Tibell were used as outgroups, based on the studies of Savić et al. (Reference Savić, Tibell, Gueidan and Lutzoni2008) and Thüs et al. (Reference Thüs, Orange, Gueidan, Pykälä, Ruberti, Lo Schiavo and Nascimbene2015).
A total of 42 ITS sequences were aligned with MUSCLE v.3.8.31 (Edgar Reference Edgar2004) using EMBL-EBI’s freely available web service (http://www.ebi.ac.uk/Tools/msa/muscle/). The aligned data set was subjected to maximum parsimony analysis as implemented in TNT v.1.1 (Goloboff et al. Reference Goloboff, Farris and Nixon2008). The analysis was performed using Traditional search with random addition of sequences, with 100 replicates and TBR branch-swapping algorithm. Ten trees were saved for each replicate. Gaps were treated as missing data. Node support was estimated using the bootstrapping method (Felsenstein Reference Felsenstein1985) with 1000 replicates.
Results and Discussion
Phylogeny
Seven new nuITS sequences were obtained in this study representing four species of Atla and one of Sporodictyon schaererianum (Table 1). Our ITS phylogeny largely agrees with the previous two-gene-loci analyses of Savić et al. (Reference Savić, Tibell, Gueidan and Lutzoni2008) and Savić & Tibell (Reference Savić and Tibell2008), with one exception: in the strict consensus of 24 most parsimonious trees, the genus Atla is paraphyletic (but without support) (Fig. 1), while in the earlier analyses it appeared as a monophyletic sister group to Sporodictyon. An Atla-Sporodictyon clade is strongly supported, but the relationships between different Atla species remain unsupported.
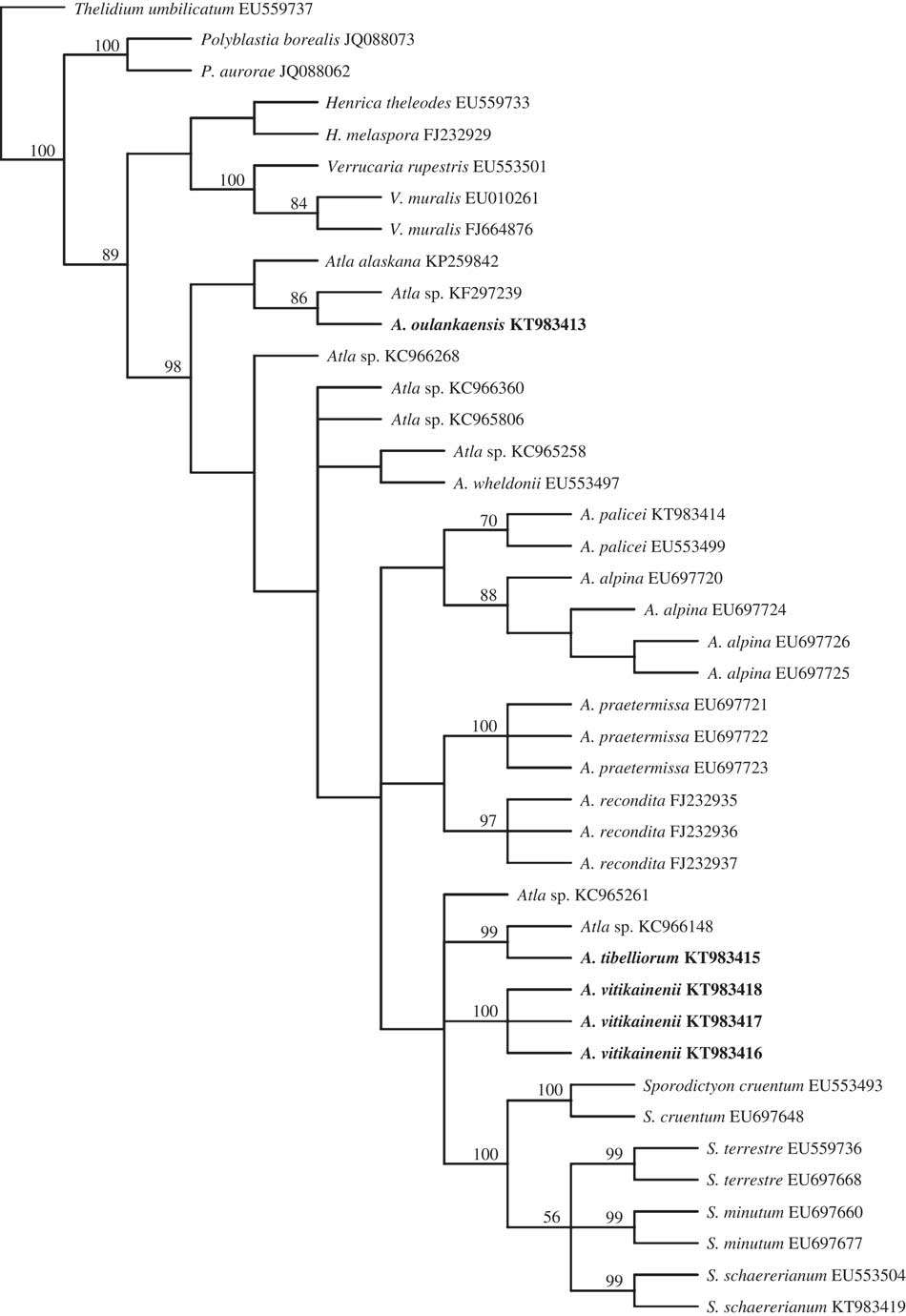
Fig. 1 Phylogeny of the genera Atla and Sporodictyon. Strict consensus based on ITS data set with bootstrap values (>50%) at nodes. New species described in this study are indicated in bold.
Table 1 Specimen information and GenBank accession numbers for taxa used in this study. New sequences are indicated by accession numbers in bold

One specimen representing a putative new species, Atla oulankaensis, groups with one unidentified ITS sequence collected from Banks Island, Canada (KF297239; Timling et al. Reference Timling, Walker, Nusbaum, Lennon and Taylor2014). The Canadian sequence differs from the Finnish one only by three gaps and most probably also represents A. oulankaensis. The sequence was obtained from a soil sample (probably spores dispersed from the nearby rocks or stones) for which no lichen voucher is available.
The ITS phylogeny suggests a close, though unsupported, relationship between A. oulankaensis and A. alaskana, and comparison of ITS sequences revealed 97·6% sequence similarity between the two species. However, A. oulankaensis and A. alaskana are morphologically clearly different (see taxonomy below).
Two of the new Atla species discovered in this study (A. vitikainenii, represented by three sequences, and A. tibelliorum) group with the members of Sporodictyon, but this relationship receives no support. One sequence obtained from the soil sample from the USA forms a strongly supported clade with A. tibelliorum and probably belongs to the same species. This suggests that the distribution of A. tibelliorum extends to North America.
In addition to the three new species, one specimen collected from Finland (and identified as A. palicei based on morphological characters; see Pykälä Reference Pykälä2013) may represent yet another undescribed species. It groups with one GenBank sequence of A. palicei but this relationship receives only 70% bootstrap support. The two sequences differ by 3%, a value which has traditionally been used as a threshold for delimiting fungal species (Begerow et al. Reference Begerow, Nilsson, Unterseher and Maier2010). However, we could not find any clear morphological difference between the specimen and the description of A. palicei by Savić & Tibell (Reference Savić and Tibell2008). The Finnish specimen has a slightly thinner involucrellum (70–90 μm thick) and a less developed non-areolate thallus. The specimen is for the time being included in A. palicei, pending more material.
Besides being distinguished from the other species of Atla by their ITS sequences, A. oulankaensis, A. tibelliorum and A. vitikainenii also show morphological differences with each other and with the previously known species of Atla and Sporodictyon. Thus, we are confident that they represent new species, although we have only one sequence available for each of A. oulankaensis and A. tibelliorum.
The main morphological differences between Atla and Sporodictyon have been the lack of cephalodia and usual absence of thalline cover of perithecia in Atla. Tibell & Tibell (Reference Tibell and Tibell2015) emphasized that thalline cover (if present) in Atla does not reach close to the ostiolum, unlike in Sporodictyon where it often (almost) reaches the ostiolum. However, A. oulankaensis and A. tibelliorum have cephalodia or cephalodia-like structures, and the perithecia of A. tibelliorum are thickly covered by the thallus up to the ostiolum. These results support the conclusion that Atla and Sporodictyon cannot always be separated by morphology alone.
Even though two of the new species, A. tibelliorum and A. vitikainenii, group with Sporodictyon in the ITS analysis, we have decided to include them in the genus Atla for the following reasons: members of Sporodictyon form a strongly supported group in the phylogeny and neither A. tibellii nor A. vitikainenii is included in this group. The strong support most probably stems from the characteristic ITS profile of all Sporodictyon sequences: these are c. 23–36 bases longer than ITS sequences of Atla due to indels. However, because Atla and Sporodictyon cannot be separated by morphology alone, we provide a key to all known species of Atla and Sporodictyon.
The new results underline the previously known fact that the genus Atla has a northern distribution. In Finland, all species of Atla are very rare and all of them (except A. wheldonii) occur only in northern Finland in areas with many other rare northern or even arctic species. Species described here as new were not found from Sweden or Norway in the study of Savić & Tibell (Reference Savić and Tibell2008), which suggests that they are probably rare or absent in the other Nordic countries.
The soil samples studied from the North American Arctic (Timling et al. Reference Timling, Walker, Nusbaum, Lennon and Taylor2014) were surprisingly well represented by species of Atla. Four North American specimens used in the analysis may represent undescribed species of Atla. One of these groups is with A. wheldonii but this relationship receives no support. The result suggests that the genus Atla may be more species-rich in North America than previously thought. More studies on lichens of arctic areas are needed to discover the real species richness of the genus. These kinds of studies are urgently needed as Atla may prove to be among the most sensitive groups of lichens to global warming because of its northern distribution.
The Species
Atla oulankaensis Pykälä & Myllys sp. nov.
MycoBank No.: MB815133
Perithecia smaller than in Atla alpina, involucrellum to the exciple base level, appressed to the exciple, in some perithecia enveloping the exciple, spores (20–)23–28(–32) μm wide.
Type: Finland, Koillismaa, Kuusamo, Juuma, Oulanka National Park, gorge Jäkälävuoma, calciferous (dolomitic) schistose rock outcrop, on NW-facing wall, 208 m, 66°15'N, 29°26'E, 16 August 2010, Pykälä 40105 (H—holotype); GenBank accession no. KT983413 (ITS).
(Fig. 2A)

Fig. 2 A, Atla oulankaensis (holotype); B, A. tibelliorum (holotype); C, A. vitikainenii (holotype). Scales=1 mm.In colour online.
Thallus grey to greenish grey, thin, c. 10–50 μm thick, continuous, rather frequently covered by cyanobacteria (Nostoc), some forming cephalodia or cephalodia-like structures, Nostoc cells 5–7 μm. Photobiont green, algal cells 5–8 μm.
Perithecia 0·30–0·38 mm diam., 1/4–immersed, young perithecia thinly covered by a thalline layer except at apex, thalline cover c. 8–15 μm thick, old perithecia without thalline cover; perithecial density c. 50–80 cm-2. Ostiole pale to dark, depressed, 30–100 μm wide. Involucrellum to the exciple base level or incurving under the exciple, or enveloping the exciple, 38–76 μm thick, appressed to the exciple. Exciple 0·25–0·37 mm diam., wall medium brown to dark brown, c. 30–35 μm thick. Periphysoids c. 30–63×1–2 μm, branching to anastomosing. Asci c. 147–177×63–72 μm, 8-spored. Ascospores dark brown (a few mature or even overmature spores pale), muriform, (42–)51–60–69(–75)×(20–)23–25–28(–32) μm (n=72), with (10–)12–16 transsepta reaching the periphery along one side of the spores in a median optical section, and with 4–6(–7) longisepta in the central part, spores often bent, old spores easily broken into two pieces.
Etymology. The species is named after the Oulanka National Park and surroundings, which is one of the lichenologically most valuable areas in Finland.
Habitat and distribution. The specimen is from a shady wall of a calciferous rock. The locality is a gorge famous for many rare and threatened lichens and plants. Atla oulankaensis is apparently a very rare and threatened species in Finland. The Canadian specimen from Banks Island was collected on bare soil with high pH (8·4) (Timling et al. Reference Timling, Walker, Nusbaum, Lennon and Taylor2014). It is possible that the main range of A. oulankaensis is in North America, reminiscent of the distribution pattern of Peltigera retifoveata Vitik., which occurs in the Oulanka area but is very rare elsewhere in Europe. Its main distribution lies in western North America (Goffinet Reference Goffinet1992). In fact, Jäkälävuoma, the only Finnish locality of A. oulankaensis, is also the type locality for P. retifoveata (Vitikainen Reference Vitikainen1985).
Notes. Based on ITS phylogeny, A. oulankaensis is closely related to A. alaskana but the two species clearly differ by their morphology and ecology. Atla alaskana has larger perithecia, pale spores and a thick thallus. It occurs on open calciferous rock (Tibell & Tibell Reference Tibell and Tibell2015), whereas A. oulankaensis occurs on the shady NW-facing wall of a calciferous rock. Morphologically, A. oulankaensis is rather similar to A. vitikainenii and more material is needed to determine whether the species can be unambiguously identified on the basis of morphological characters. In A. oulankaensis the involucrellum is appressed to the exciple and often incurving under or even enveloping the exciple, while in A. vitikainenii the involucrellum is often slightly diverging from the exciple and not incurving under it. Spores of A. oulankaensis are on average narrower, but there is much overlap in the spore size. Atla oulankaensis may also be confused with Sporodictyon cruentum (Körb.) Körb., which has larger perithecia, a brown thallus and different habitat (siliceous rocks by water). The spores of A. oulankaensis may sometimes turn brown rather late and thus it may also be confused with species having pale spores. Sporodictyon minutum Savić & Tibell (Savić & Tibell Reference Savić and Tibell2009) has smaller spores and a thinner involucrellum.
Atla tibelliorum Pykälä & Myllys sp. nov.
MycoBank No.: MB815134
Similar to Sporodictyon arcticum, but exciple wall dark brown and spores larger (63–73×40–46 μm).
Type: Finland, Enontekiön Lappi, Enontekiö, Porojärvet, Toskalharji, Toskaljärvi N, fell, calcareous alpine grassland, on dolomite pebbles, 730 m, 69°12'N, 21°26'E, 2 August 2011, Pykälä 43330 (H—holotype); GenBank accession no. KT983415 (ITS).
(Fig. 2B)
Thallus pale ochraceous, rimose to areolate, c. 200–280 μm thick, areoles 0·4–1·0 mm wide, algal cells 6–11 μm, cortex c. 5–25 μm thick, epinecral layer 8–13 μm thick; few cephalodia or cephalodia-like structures (containing Nostoc) present.
Perithecia immersed in thalline warts, only exposed surrounding the ostiole, exposed part 0·10–0·15 mm wide, thalline cover of perithecia c. 50–150 μm thick; perithecial density c. 30 cm−2 . Ostiole inconspicuous, tiny, dark, depressed, c. 30–60 μm wide. Involucrellum to the exciple base level, 50–70 μm thick, sometimes thickened towards the base to 60–110 μm thick, appressed to the exciple, sometimes somewhat diffusely pigmented at base. Exciple 0·25–0·48 mm diam., wall dark brown. Periphysoids c. 55–93×1·0–1·5 μm, branching. Asci c. 147–177×63–72 μm, 8-spored. Ascospores dark brown, muriform, (57–)63–68–73(–80)×(38–)40–43–46(–52) μm (n=30), with 10–15 transsepta reaching the periphery along one side of the spores in a median optical section, and with 5–7 longisepta in the central part.
Etymology. The species is named in honour of Sanja and Leif Tibell, the mother and father of the genus Atla, for their invaluable contribution to the taxonomy of the “polyblastioid” genera of lichens (Polyblastia s. lat.).
Habitat and distribution. The specimen is from a dolomite pebble on a fell in NW Finland. The species occurs on alpine grassland close to flat open dolomite rocks. Numerous rare lichens occur in the Toskalharji area, which is the largest area of calcareous rocks in Finland. Apparently A. tibelliorum is a very rare species in Finland. The North American soil sample including A. tibelliorum was collected from the dwarf shrub tundra in the Franklin Bluffs, Alaska (USA) from high pH soil (8·0) (Timling et al. Reference Timling, Walker, Nusbaum, Lennon and Taylor2014).
Notes. The species clearly differs morphologically from the other species of Atla by having the perithecia immersed in warts, and in the large brown spores. It is rather similar to Sporodictyon arcticum Savić & Tibell, which also has thickly thallus-covered perithecia, but that species has a pale exciple and smaller spores (see Savić & Tibell Reference Savić and Tibell2009). Based on morphology, A. tibelliorum and S. arcticum may be closely related. Unfortunately, no sequence of S. arcticum is available.
Atla vitikainenii Pykälä & Myllys sp. nov.
MycoBank No.: MB815132
Perithecia smaller than in Atla alpina, involucrellum mainly slightly diverging from the exciple, not incurving under the exciple, spores (25−)26–30(−33) μm wide.
Type: Finland, Koillismaa, Salla, Oulanka National Park, W of Savikoski, cliff, dolomite rock outcrop, NE-facing wall, on pebbles, with Verrucaria cf. inaspecta Servít, 185 m, 66°25'N, 29°10'E, 17 August 2010, Pykälä 40222 (H—holotype); GenBank accession no. KT983416 (ITS).
(Fig. 2C)
Thallus thin, continuous or fleck-like, occasionally slightly rimose, grey, ochraceous, pale green or dark brown, c. 20–100 μm thick. Photobiont green alga, algal cells c. 5–7 μm.
Perithecia 0·25–0·46 mm diam., 1/4–1/2(–3/4)-immersed, not leaving pits or leaving shallow pits, often thinly thallus-covered except for the apex; perithecial density c. 20–80 cm-2. Ostiole dark, plane or depressed, 20–80 μm wide. Involucrellum extending to the exciple base level, 50–100 μm thick, appressed to the exciple or slightly diverging from the exciple. Exciple 0·25–0·38 mm diam., wall pale to dark brown, 25–34 μm thick. Periphysoids c. 44–54×1·5–2·5 μm, branching. One mature ascus seen 150×63 μm, 8-spored. Ascospores dark brown, muriform, (40–)54–59–64(–70)×(25–)26–28–30(–33) μm (n=136), with (10–)12–16(–18) transsepta reaching the periphery along one side of the spores in a median optical section, and with (3–)4–6 longisepta in the central part, spores often bent, old spores easily broken into two pieces.
Etymology. The species is named in honour of Orvo Vitikainen, who has contributed in many ways to our knowledge of the taxonomy, ecology and biogeography of Finnish lichens. The first author is much indebted to Orvo, who has generously helped in identifying lichens over many years and has provided encouragement during the process of becoming a professional lichenologist. Furthermore, Orvo has previously described a new species from calcareous habitats of Finland (Peltigera retifoveata).
Habitat and distribution. Four specimens are known (three sequenced specimens), three of them collected from dolomite pebbles and one from a dolomite boulder. The species occurs both in the Kilpisjärvi and Oulanka areas, the only two areas in northern Finland with a cluster of larger calcareous rocks. Atla vitikainenii is a northern species, probably mainly occurring in arctic areas and in northern mountains above the alpine tree line. The occurrences in the Oulanka area may be relictual. The Oulanka area has many northern/ arctic-alpine lichens and plants considered to be relicts from colder climate periods.
Notes. Atla vitikainenii is fairly similar to A. oulankaensis (see A. oulankaensis for differences). Sporodictyon cruentum has larger perithecia, slightly smaller spores and a brown thallus. All three sequenced specimens of A. vitikainenii belong to the same haplotype, although they show considerable morphological differences in thallus colour and exciple wall colour.
Additional specimens examined. Finland: Enontekiön Lappi: Enontekiö, Porojärvet, Toskalharji, Toskaljärvi N, fell, dolomite scree, on dolomite pebbles, with Thelidium fontigenum A. Massal., 710 m, 69°11'N, 21°26'E, 2011, Pykälä 43378 (H); Enontekiö, Porojärvet, Toskalharji, 1·2 km NE of Toskaljärvi, fell, SW-slope, gentle E-slope, Dryas heath, on dolomite pebbles, 875 m, 69°12'N, 21°28'E, 2011, Pykälä 43614 (H). Koillismaa: Kuusamo, Oulanka National Park, Taivalköngäs, shore of Oulankajoki River, Picea abies- dominated herb-rich forest, NE-slope, dolomite boulder, on NE-facing wall, 171 m, 66°24'N, 29°11'E, 2011, Pykälä 44595 (H).

This study is part of the research project “Threatened lichens of calcareous rocks” (Grant number YTB059), which belonged to the research programme of deficiently known and threatened forest species (PUTTE) financed by the Ministry of the Environment. Kone Foundation and the Finnish Cultural Foundation are thanked for their financial support through the FinBOL project to the Finnish Museum of Natural History. Anne Palomäki took the photographs. Teuvo Ahti and two anonymous referees are thanked for constructive comments on the manuscript.



