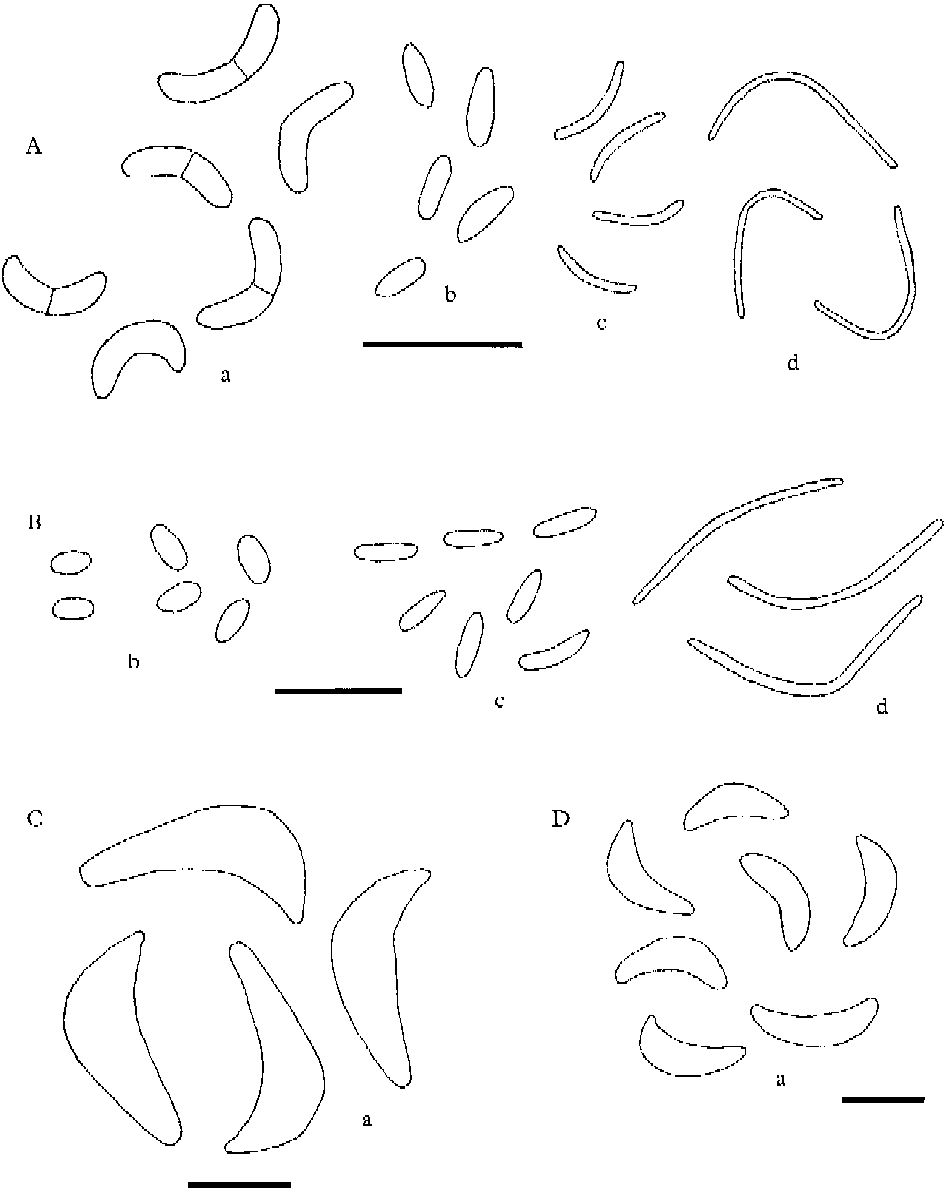Introduction
Lecanora species most closely related to L. saligna form a well-defined group. According to Printzen (Reference Printzen2001) it is characterized by a corticate, lecanorine apothecial margin, bacilliform and slightly bent conidia, isousnic acid, instead of or in addition to usnic acid, in the thallus and apothecia (with a few exceptions) and occurrence in habitats at elevations above 1200 m (this is the case for SW North American species). Observations on conidia are presented only for L. mughicola and L. saligna. However, in Printzen's opinion there is no reason for a generic distinction.
Conidial and pycnidial characters are of great help with the identification of the species in the L. saligna group. We have studied intensively the occurrence and the nature of pycnidia and conidia found in all species treated in the group and are now able to add some new information about conidia for several species. Four kinds of conidia are recognized. Our concept of the distinguishing features of three types, microconidia, mesoconidia and macroconidia, are analogous to those used by Coppins (Reference Coppins1983: 67) in Micarea. We introduce here a fourth conidium type, termed leptoconidia, which differ from macroconidia as defined in Coppins (Reference Coppins1983: 67). Leptoconidia are filiform, often curved and relatively narrow, non-septate, 0·6–1·0 μm wide and vary in length between 10 and 27 μm. They are known from many Lecanora species, even from some outside the L. saligna group. Macroconidia as treated here are slightly to strongly curved, 0–3-septate and 1·7–3 μm wide. Macroconidia have been found in 6, microconidia in 12, and mesoconidia in 7 of the taxa treated here. There is much confusion regarding measurements of conidia in the literature, which can be explained by the numerous specimens within the Lecanora saligna group that have been misidentified. Consequently conidial chararacters reported in the literature are compared with those observed in our study.
Some additional chemical compounds are also reported in this study for some species of the L. saligna group; most of them recorded for the first time in the group. These are brialmontin 1, neousnic acid, 7-O-methylnorascomatic acid, paraensic acid C and D, pseudoplacodiolic acid, placodiolic acid and squamatic acid. Although the chemistry is often useful for separation of the species, some compounds are encountered in several species. Lecanora sarcopidoides is unique in having pseudoplacodiolic acid as the major compound and the chemistry of L. hypoptoides is easily recognized by the presence of paraensic C and D acids. In our study, L. subintricata is unique in having usnic acid as its major compound, although it has been found as a rare major compound in North American specimens of L. albellula Printzen (Reference Printzen2001).
A detailed study of Lecanora specimens belonging to the L. saligna group, has led to the discovery of four new species and a new variety: L. calabrica, L. coppinsii, L. pseudosarcopidoides and L. subsaligna and L. albellula var. macroconidiata, all from western or central Europe. A few 19th century specimens and one recent collection of L. subravida have been studied and this species, described by Nylander (Reference Nylander1872), appears to belong to the L. saligna group. The present study includes twelve species and one variety of the L. saligna group or closely related species, all of them corticolous or lignicolous.
Material and Methods
This study is based on material of the Lecanora saligna group, mainly from the herbaria of both authors, with additional collections from B, BR, E, L, M, GZU, LG, LUX, MAF, H-Nyl and the private collections of P. Diederich and J. Etayo. We have attempted to observe all the species treated in their natural habitats, so both authors made many fieldtrips throughout most countries of Europe with an emphasis on western countries of continental Europe. A total of about 400 specimens has been examined, some of them are indeterminable because of the lack of pycnidiomata, often in combination with their scanty nature. We have studied specimens from central and western Europe and some specimens from elsewhere.
Hand-cut sections and squash preparations were usually mounted in water and 10% KOH. Measurements of internal features were made at a magnification of × 1000 or rarely at × 2000 using a Leitz microscope fitted with a drawing tube. For the examination of conidiogenesis and conidia or ascospores only material mounted in water was employed.
The granulation of tissues was observed in polarized light. Granules, bright in polarized light, have been found in all the species treated here except Lecanora hypoptoides. These granules are yellowish to pale brownish and soluble in 25% KOH (K) and not soluble in 35% HNO3 (N). These two reagent tests have been carried out on material of all species. The epihymenium in all treated species is K− and N± reddish. Amyloid reactions were tested using Lugol's iodine solution (K/I). Many specimens were checked by TLC (nearly always in TDA) and some by HPLC. The identity of chemical compounds was checked partly by the second author and partly by Prof. J. Elix, according to the methods of Culberson & Ammann (Reference Culberson and Ammann1979), Culberson & Johnson (Reference Culberson and Johnson1982), or Elix & Ernst-Russell (Reference Elix and Ernst-Russell1993).
Key to the species of the Lecanora saligna group
1 Ascospores 10–14 μm long and 4·4–6·2 μm wide 2
Ascospores usually shorter and/or narrower 3
2(1) Ascospores 10–14 × 4·4–5·2; apothecia plane, appressed; margin somewhat shiny; disc yellowish brown to often bluish grey to black L. mughicola
Ascospores 10·5–11 × 5–6(−6·2) μm; apothecia with a relatively thick yellowish margin, becoming folded or knobbly; habitus resembles L. varia L. subravida
3(1) Ascospores relatively small, c. 6–8·4 × 2·5–3·6 μm; disc yellowish beige to grey, becoming dark olive-brown or bluish pruinose 4
Ascospores usually >8 μm long and >3·5 μm wide; disc different 5
4(3) Ascospores 6·7–8·4(−10) × 3·1–3·6 μm; thallus and apothecia margin yellow (usnic acid as major substance); disc yellowish beige or grey, becoming dark olive brown, ± slightly white or bluish pruinose L. subintricata
Ascospores 6–8(−8·5) × 2·5–3·5 μm; thallus and apothecia margin greyish (isousnic acid as a major substance); apothecia becoming biatorine; disc bluish pruinose L. coppinsii
5(3) Ascospores 8·4–9·6 × 5–5·7 μm; granules of isousnic present in epihymenium; habitus resembles L. hagenii L. calabrica
Ascospores <5 μm wide; granules of isousnic acid ± present in epihymenium; habitus not resembling L. hagenii 6
6(5) Thallus KC+ yellow (pseudoplacodiolic acid); apothecia mostly plane, whitish pruinose, except in (over)mature apothecia L. sarcopidoides
Thallus KC−; apothecia often becoming convex; disc not or usually slightly pruinose 7
7(6) Thallus warted, thick, to 0·35 mm; apothecia without yellowish tinge; disc reddish brown to dark brown, becoming blackish, epruinose; ascospores simple to rarely 1-septate; thallus and apothecia with minute oil droplets (paraensic C and D acids) L. hypoptoides
Thallus thinner; apothecia with a pale yellowish margin; disc pale yellowish, yellowish brown to medium brown or a mixture of pale yellowish and more darker, ± pruinose 8
8(7) Macroconidia abundantly present, non-septate, curved >6 μm long and >1·5 μm wide; margin of apothecia often persistent; epihymenium yellow-brown to olive brownish, N− 9
Macroconidia different, usually present but not easily found, or mesoconidia present; margin of apothecia often excluded; epihymenium brownish, N+ reddish violet 10
9(8) Macroconidia 6·0–8·1 × 2·0–2·4 μm; lignicolous or corticolous L. saligna
Macroconidia 8·5–9·5 × 2·7–3·0 μm; corticolous or on decorticated trunks L. quercicola
10(8) Apothecia finally with a disc-coloured margin, pale to dark brown, becoming convex, ± biatorine; ascospores 8·6–11·3 × 3·4–4·7 μm; macroconidia not known; mesoconidia not rare, straight or very slightly curved, 4·4–6·3 × 1·4–2·2 μm L. pseudosarcopidoides
Apothecia with a thallus-coloured margin; ascospores usually less wide; macroconidiaseptate, strongly curved; mesoconidia absent or 2·9–5·1 × 1·6–2·1 μm 11
11(10) Macroconidia 1-septate, rarely a few 3-septate, (19–)22–27(−30) μm long, tips attenuated L. subsaligna
Macroconidia mainly 1–3-septate, 7–14 μm long, tips obtuse 12
12(11) Macroconidia 12·4–14 × 1·8–2·3 μm, 1–3-septate; lignicolous and on Pinus or Populus bark L. albellula var. macroconidiata
Macroconidia 7–10·5 × 1·8–2·1 μm, 1(−3)-septate; on bark of trunks and branches of various trees, decorticated trunks or rarely on wood of fence posts L. albellula
The New Taxa
Lecanora albellula Nyl. var. macroconidiata M. Brand & van den Boom var. nov
Varietas lignicola, Lecanorae albellulae similis sed thallo verrucoso et verrucis ad 0·15(−0·2) mm crassis, ascosporis 9–11·5(−13) × 3·4–3·7(−4) μm, macropycnidiis frequentibus; macroconidia 12·4–14·0 × 1·8–2·3 μm, valde curvata (c. 45–90°), paulo attenuata vel apicibus obtusatis, 1–3-septata; microconidia rara, 9·6–12·3 × 1·0–1·1 μm, valde curvata; leptoconidia rara, c. 14 × 0·8 μm; thallus acidum isousnicum et ± 7-O-methylnorascomaticum continens.
Typus: France, Jura, 10 km S of Champagnole, 1 km NE of Chaux Crotenay on fence post along meadow, 680 m, 25 July 1986, M. Brand 14651 (LG—holotypus; hb. Brand—isotypus).
(Figs 1C & 2)
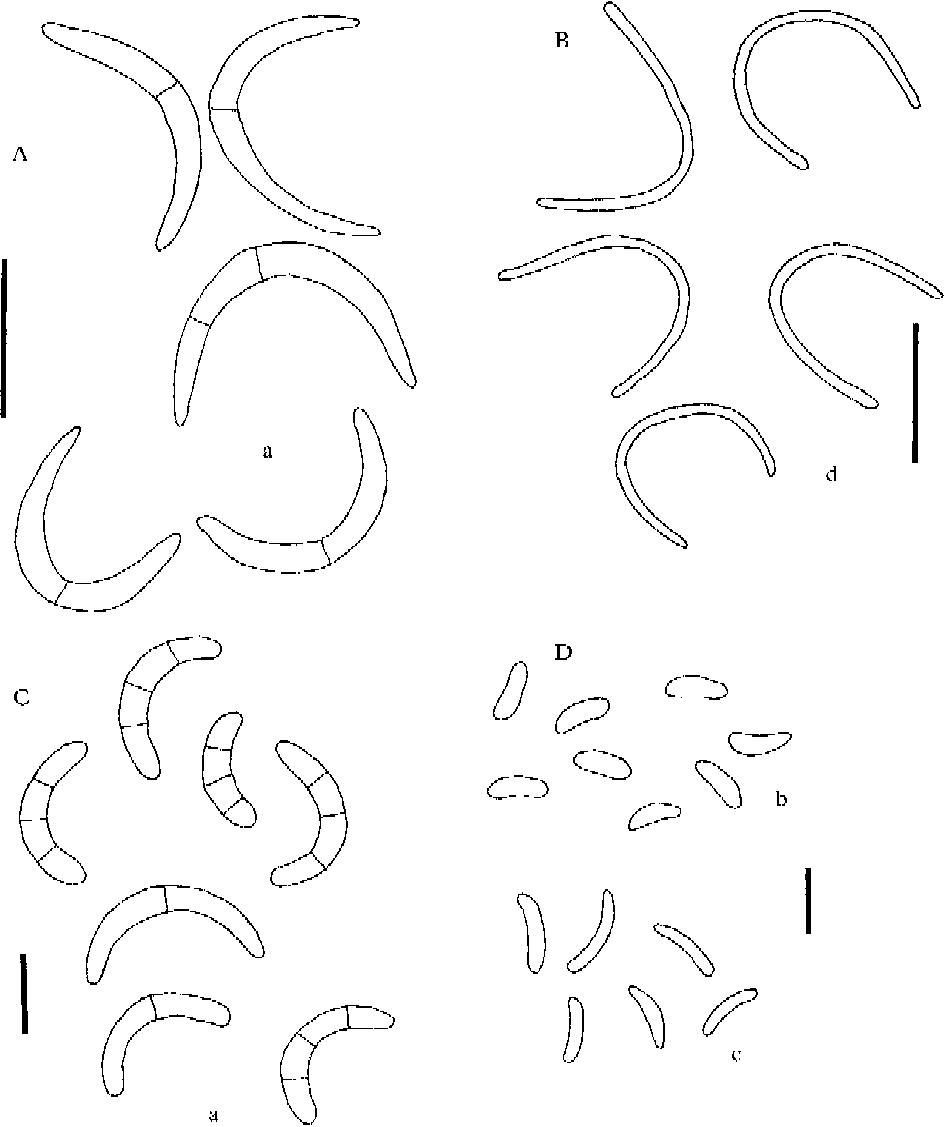
Fig. 1. Lecanora saligna group, conidia. A, L. subsaligna; B, L. calabrica; C, L. albellula var. macroconidiata; D, L. pseudosarcopidoides. a=macroconidia; b=mesoconidia; c=microconidia; d=leptoconidia. Scales: A & B=10 μm; C & D=5 μm.

Fig. 2. Lecanora albellula var. macroconidiata, habitus (holotype). Scale: 0·5 mm.
Thallus corticolous or lignicolous, effuse, up to 2 cm wide, often aggregating in patches, consisting of warted to sublobulate particles, warts subglobose and 0·15–0·25 mm diam., scattered to mostly crowded, to 0·15(−0·2) μm thick; without cortex, with a hyaline epinecral layer, 5–15 μm tall; in section with minute granules scattered all over the thallus, without medulla, algal layer not separated; upper surface pale brown or yellowish brown to medium brown, shiny. Photobiont cells ± globose, 6–15 μm diam. Prothallus not observed.
Apothecia abundant, scattered to crowded and overlapping, sessile, rounded, 0·2–0·6 mm diam.; margin prominent, initially raised, especially in young apothecia, becoming level with the disc and excluded in mature apothecia, entire, concolorous with thallus, 0·03–0·07 mm wide; disc plane, becoming moderately or strongly convex, pale or reddish brown, dark greyish brown to blackish, slightly pruinose or epruinose, often with a mixture of pale with darker brownish discs. Thalline excipulum with cortex, chondroid, 20–35 μm wide at base, sometimes reduced. Parathecium hyaline at inner part, brownish at outer rim and to 35 μm wide. Epihymenium hyaline to spotted greyish brown, with minute granules, N+ reddish violet. Hymenium hyaline, with a few orange granules, 35–65 μm. Hypothecium hyaline, to 120 μm high. Paraphyses mostly branched to anastomosing, septate, sometimes slightly constricted at septa, 1·5–2 (−2·5) μm wide in the middle, top sometimes slightly widened up to 3·5(−5) μm wide, hyaline to greyish olive-brown in a weakly developed gel matrix. Ascospores ellipsoid, simple, overmature spores sometimes 1-septate, 9–11·5(−13) × 3·4–3·7(−4) μm.
Macropycnidia often present, sessile or somewhat stalked, pale to dark brown in the upper part, 150–190 × 100–140 μm. Macroconidia curved, with obtuse, not or slightly attenuate apices (allantoid), (0–)1–3-septate, 12·4–14·0 × 1·8–2·3 μm. Micropycnidia rarely found, immersed, c. 50 μm diam. Microconidia rather strongly curved, 9·6–12·3 × 1·0–1·1 μm. Leptopycnidia rarely found, immersed, c. 50 μm diam. Leptoconidia c. 14 × 0·8 μm.
Chemistry. K−, C−, KC−, Pd−. Isousnic acid (major), 7-O-methylnorascomatic acid (major). This latter compound was detected in most specimens recorded here (TLC & HPLC).
Ecology and distribution. Lecanora albellula var. macroconidiata is known from bark of Pinus and Populus tremula, and on wooden fence posts. The holotype, collected from a wooden fence post, is accompanied only by fragments of Parmelia cf. sulcata. The other specimens are accompanied by Lecanora symmicta and also sometimes by some poorly developed lichen material, belonging in Buellia and Rinodina. The new variety is known so far only from western Europe, where it seems to be rare and confined to lowland areas, apart from the most southern record from 1600 m.
Notes. This variety is easily mistaken for Lecanora albellula s. str., but differs by its more warted, thicker and shiny, sometimes sublobulate thallus with more convex (subglobose) apothecia and with macropycnidia, and even macroconidia, which are larger and more often 3-septate. In addition, the microconidia are longer than those in L. albellula. The chemistry also differs, as it contains 7-O-methylnorascomatic acid (major), a chemical compound which has been recently recognized only in the Lecanora saligna group (van den Boom Reference van den Boom2007). It has also been detected in L. subsaligna and L. albellula s. str., but only in small amounts. The mixture of yellow and brown apothecial discs, representing all states of development, is typical of this new variety. This same characteristic has sometimes been recorded, probably from misidentified material, for L. albellula. Comparison of the two varieties suggests that L. albellula var. macroconidiata should perhaps have the status of species. However, of the few collections available, only two are well developed. More knowledge of its variability and ecology, as well as molecular data, are needed to determine its taxonomic status.
Additional specimens examined. The Netherlands: Friesland: Ameland, Lange Duinen, binnenduin, 5°39.3′E, 53°27.4′N, Populus tremula woodland in dune area, 1989, M. Brand 21079 (hb. Brand). Gelderland: Nijkerkerveen, 5°30.1′E, 52°11.6′N, dead Pinus trunk in heathland, 1974, M. Brand 4070 (hb. Brand).—Sweden: Norrbotten: 95 km NNE of Umeå, 50 m, on Salix, 1988, P. van den Boom 8025 (hb. v.d. Boom).—France: Yonne: N of Pontigny, N of Auxerre, 3°41′E, 47°55′N, 200 m, on wood of pole of fence, 1976, M. Brand 6602 (hb. Brand).—Spain: Teruel: Javalambre, 7 km NNE of top, 1°0.8′W, 40°9.85′N, 1600 m, Pinus halepensis, open Pinus wood, wood of dead branches, 2005, M. Brand 52019 (hb. Brand).
Exsiccata examined: Crombie, Lich. Brit. exs. 160 (B). [as Lecanora piniperda: ad ligna vetusta fabrefacta Mill Hill in Angliae Middlesex].
Lecanora calabrica M. Brand & van den Boom sp. nov
Species lignicola vel corticola, habitu Lecanorae hageni similis, sed thallo laeve, nonnumquam rimuloso, ad 120 μm crasso, ascosporae late ellipsoideae, 8·4–9·6 × 5·0–5·7 μm, simples; leptopycnidia frequentes; leptoconidia filiformia, curvata, 22–27 × 0·6–0·9 μm; thallus et apothecium acidum isousnicum continentes.
Typus: Italy, Calabria, Sila Grande, Lago Arvo, 1 km NE of Barachella, 16°29′E, 39°14′N, 1200 m, on Robinia along road near bridge over a small stream, 14 May 2001, M. Brand 42986 (LG—holotypus; hb. Brand—isotypus).
(Figs 1B & 3)

Fig. 3. Lecanora calabrica, habitus (holotype). Scale: 0·5 mm.
Thallus in rather small patches, up to c. 1·5 cm, sometimes weakly rimose, up to 120 μm thick, composed of loose hyphae, without a clear cortex, sometimes containing granules; upper surface rather smooth, whitish. Photobiont chlorococcoid, cells c. 11–14 μm diam., rather thick walled, with only a few haustoria. Prothallus often present, bluish grey.
Apothecia broadly appressed, c. 0·5–0·6 μm wide; margin thin (0·05–0·09 mm wide), persistent, pale greyish; medulla filled with algae, with rather large granules at the outer edge; disc plane to slightly convex, beige or pale to dark brown. Thalline excipulum cortex clearly present, 25–55 μm thick at the base, hyaline, chondroid, consisting of anticlinal or branched hyphae to 6 μm thick, lumina 1 μm wide; granules present at the top of the excipulum and more or less in epipsamma, stout, for example 2·5–3 μm wide. Parathecium hyaline at inner part, dark brown outer rim and to 35 μm wide. Epihymenium hyaline to slightly brownish, N+ reddish violet. Hymenium hyaline, c. 45–70 μm. Hypothecium hyaline, to 120 μm high. Paraphyses simple, conglutinated, to 2·7 μm wide, top not or slightly widened, often pale brownish. Asci c. 35 × 12 μm; tholus amyloid with a broad axial mass. Ascospores broadly ellipsoid, simple, wall rather thick, 0·4–0·5 μm, 8·4–9·6 × 5·0–5·7 μm.
Leptopycnidia often present, immersed, to c. 150 μm diam., apices pale brown or hyaline: leptoconidia thin and relatively long, curved, 22–27 × 0·6–0·9 μm.
Chemistry. K−, C−, KC−, P−. Isousnic acid (TLC).
Ecology and distribution. Lecanora calabrica occurs mostly on smooth bark of Abies, Castanea, Corylus, Fagus, Juglans, Populus, Robinia and Salix in mountain areas in southern Italy (Abruzzi and Vesuvius to Calabria) where it appears not to be rare. It occurs, for example, with Caloplaca cerina, Lecanora carpinea, Lecidella elaeochroma, Rinodina capensis, etc. Elsewhere it is known only from Croatia (Velebit).
Notes. In the field, Lecanora calabrica is very easily confused with L. hagenii, but differs in its chemistry and by its wider, sometimes nearly globose ascospores. Both species have a similar ecology. Other species in the L. saligna group which also possess isousnic acid, have usually paler apothecia, a more yellowish appearence and smaller ascospores, except L. subravida, which has larger ascospores, 10·5–11 × 5–6(−6·2) μm, and also a more prominent apothecial margin.
Additional specimens examined. Italy: Lazio: pr. Frosinone, W of Roccasecca, NE of Colle S.Magno, 13°42.7′E, 41°33.2′N, 750 m, Juglans in meadow in mountain valley, thinner branches, 2001, M. Brand 42636 (hb. Brand). Abruzzo: shore of Fiume Sangro, SW of Viletta Barrea, 13°54.9′E, 41°46.3′N, 1000 m, Corylus in wood, 2001, M. Brand 43553 (hb. Brand). Calabria: Sila Grande, Botte Donata, c. 0·5 km SW of top, 16°26.7′E, 39°16.7′N, 1880 m, on Fagus in wood on flat top of mountain, 2001, M. Brand 42947 (hb. Brand); Sila Greca, M. Altare (col in road Longobucco–Lago di Cecita), 16°33.3′E, 39°25.2′N, 1500 m, Fagus in wood on top of mountain ridge, 2001, M. Brand 42931 (hb. Brand); 18 km ENE of Catanzaro, along road from Cropani to Botricello, 200 m, on Juglans along meadow and road, 1986, P. van den Boom 4974 (hb. v.d. Boom); Aspromonte, 2 km S of S. Eufemia, road to Villagio de Leo, edge of plateau, 15°51.4′E, 38°14.8′N, 950 m, old Castanea, in open field, 2001, M. Brand 43105 (hb. Brand); Aspromonte, 7.5 km SE Gambari, Diga Menta, valley below new constructed dam, 15° 53.9′E, 38°7.4′N, 1600 m, on Abies in forest in cleft, 2001, M. Brand 43262 (hb. Brand); Aspromonte, c. 3 km S of Gambari, 15°51′E, 38°8′N, 1600 m, Salix shrub near brook in valley with Abieto-Fagetum, 2001, M. Brand 43146 (hb. Brand). Campania: Napoli, Vesuvio, 0·6 km N of top, 14°25.5′E, 40°49.6′N, 1100 m, Populus, isolated, small trees on N slope, on bare slope of lapilli, 2001, M. Brand 42711 (hb. Brand).—Croatia: Velebit : 21km S of Senj, 1 km N of Vucjak, 14°58.5′E, 44°49.1′N, 1450 m, on Fagus in Fagus forest, 1987, M. Brand 16660 (hb. Brand).
Lecanora coppinsii M. Brand & van den Boom sp. nov
Species lignicola, Lecanorae abellulae similis, sed differt thallo endoxylico ad epixylico, paulo rimoso areolato, interdum laeve, ad 100 μm crasso, ascosporis 6·0–8·0 (−8·5) × 2·5–3·5 μm, ellipsoideis, simplicibus; mesoconidia 2·8–3·3 × 1·1–1·2 μm parva, ellipsoidea vel bacilliformia, vel ± curvata; microconidia simplicia, 4·5–5·5 × 0·8–0·9 μm, bacilliformia vel curvata; thallus acidum isousnicum continens.
Typus: Great Britain, Scotland, V.C. 82 East Lothian, Lammermuir Hills, Monynut Forest, 36/689.671, on lignum of conifer branch by forest path, alt. 340 m, 31 October 1999, B.J. Coppins 18567 (E—holotypus; hb. v.d. Boom—isotypus).
(Figs 4 & 5)

Fig. 4. Lecanora coppinsii, habitus (holotype). Scale: 0·5 mm.

Fig. 5. Lecanora coppinsii, conidia. a=macroconidia; b=microconidia. Scales: a=1 μm; b=2 μm.
Thallus endoxylic to epixylic, very thin, sometimes slightly rimose areolate, uneven to sometimes contiguous, patches up to 4 cm wide, up to 100 μm thick, but mostly found in the substratum, without cortex, granules <3 μm wide, present and scattered throughout the thallus, algal cells scattered over the thallus; upper surface smooth, grey, yellowish grey, or pale to dark brownish grey. Photobiont chlorococcoid, cells c. 9–18 (−20) μm.
Apothecia c. 0·4–0·8 mm wide, closely appressed to broadly sessile and slightly constricted at the base, sometimes innate in the substrata, irregular in appearance, rounded to oval; margin at first prominent, 0·02–0·05 mm wide, initially raised, becoming level with the disc, concolorous with the thallus or paler, becoming darker and concolorous with the disc, mainly crenulate, evanescent and apothecia finally biatorine; disc at first flat, later convex to sometimes nearly subglobose, pale pinkish brown to pale orange, cream, yellowish brown, pale to intensively grey-brown to bluish brown or greyish, grey to bluish pruinose. Thalline excipulum with a weakly developed cortex, first thin but mostly becoming thicker (20–50 μm), finally evanescent, algae present from the outermost part to inner part reaching the hypothecium. Parathecium not developed. Hymenium with polarized, orange-brown scattered to clustered granules, 25–50 μm high. Epihymenium yellowish to greyish brown with olive parts (spotted), filled with many coarse, yellowish granules, N−. Hypothecium hyaline to pale yellowish, 40–60 μm high. Paraphyses immersed within a gelatinous matrix of the hymenium, simple to more often strongly branched, septa difficult to observe, 1–1·5(−2) μm diam., top not widened, hyaline to sometimes weakly more or less brown to olive greyish pigmented. Asci clavate, Lecanora- type, 25–30 × (7–)8–10(−11) μm. Ascospores c. 6·0–8·0(−8·5) × 2·5–3·5 μm, ellipsoid, simple.
Micropycnidia immersed to semi-immersed, often on apothecia (margin or hymenium), 60 μm high × 50–70 μm wide, ostiole moderately to dark brown with greenish tinge; microconidia bacilliform to slightly curved, 4·5–5·5 × 0·8–0·9 μm. Mesopycnidia (not rarely present) immersed, 65–100 μm × 50–100 μm, top brownish pigmented, or bluish green to blackish, mesoconidia short bacilliform to weakly curved, 2·8–3·3 × 1·1–1·2 μm.
Chemistry. K± yellow, C−, KC−, P−. Isousnic acid (major) (TLC).
Ecology and distribution. Lecanora coppinsii has been found on wood, usually on rotting trunks of conifers, at altitudes of 100–380 m, in poorly developed lichen communities. In all the collections studied, Lecanora conizaeoides has been found, sometimes accompanied by Hypocenomyce scalaris, Hypogymnia physodes, Mycoblastus fucatus and Parmelia sulcata. All this material is in poor condition.
Notes. The known distribution of L. coppinsii is limited to a rather small area in southern Scotland, but it is probably an overlooked species. We found no reference to this species, either in the literature or among herbarium specimens. All the specimens studied were labelled as Lecanora piniperda (=L. albellula). Although the latter species is rather similar (it can have a similar chemistry with isousnic acid as major substance), it has longer and wider ascospores, a better developed apothecial margin, larger mesoconidia (see Table 1), is not found exclusively on wood and is distributed from lowland to alpine areas. Lecanora coppinsii can be confused with a Lecanora saligna in poor condition which is also found in lowland areas. Lecanora saligna, however, has larger ascospores, apothecia with a more prominent margin and a dark red-brown, rarely pale white yellowish pruinose disc (bluish pruinose in L. coppinsii). Macropycnidia are abundant in L. saligna and unknown in L. coppinsii. Even L. pseudosarcopidoides may be confused with L. coppinsii, but in the former, the apothecia are paler and become subglobose, the mesoconidia and ascospores are larger and it has a preference for more rotten wood. The only two species of the L. saligna group with similar small ascospores are L. subintricata and L. latens. However, these two species possess usnic acid as a major compound and the ascospores in L. subintricata are in general somewhat wider. Lecanora latens is also further distinguished by a shallower hymenium and is so far known only from California. Most of the collections of L. coppinsii are infected by an unknown hyphomycete and one specimen with an unknown Psammina sp. with non palmate and non-septate branches.
Table 1. Comparison of chemistry, ascospores and conidia of the species studied in the Lecanora saligna group

* a-atranorin; b-brialmontin 1; i-isousnic acid; m-7-O-methylnorascomatic acid; n-neousnic; p-pseudoplacodiolic acid; pa-paraensic; pc-placodiolic acid; s-squamatic acid; u-usnic acid; z-zeorin. All measurements in μm.
Additional specimens examined. Great Britain: Scotland: V.C. 78, Peebles: Glentress Forest, 36/28.41, on old conifer stumps among replanted plantation, 300 m, 1993, B. J. Coppins 16378 (E). V.C. 85, Fife: East Lomond, 37/20, on hillside on rotting log, 16 v 1970, U. Duncan s.n. (E); ibid., on fir fence post on moor, 37/239059, 380 m, 16 v 1970, P. Topham s.n. (E). V.C. 90, Angus: Glenprosen, Balnabooth, West Burn, 37/3167, on rotting conifer log by stream, 1 xii 1967, U. Duncan s.n. (E); Sidlaws, near summit of Auchterhouse Hill, on rotting conifer log, iii 1975, U. Duncan s.n. (E). V.C. 104, North Ebudes: Eigg, Sandavore, on wooden post of old sheep pens, 100 m, 2000, B. J. & A. M. Coppins 19146 (E).
Lecanora pseudosarcopidoides M. Brand & van den Boom sp. nov
Species lignicola, Lecanorae salignae similis, differt thallo endoxylico ad epixylico, tenuissimo, nonnumquam paulo rimoso areolato, laeve vel irregulariter verrucoso, ad 130 μm crasso, ascosporis 8·6–11·3 × 3·4–4·7 μm, ellipsoideis, simplicibus, rariter 1-septatis; conidiarum typi tres, frequentiores; mesoconidia 4·4–6·3 × 1·4–2·2 μm parva, ellipsoidea vel bacilliformia vel curvata; microconidia simplicia ad nonnumquam 1-septata, 5·3–7·4 × 1·0–1·3 μm, semper paulo curvata; leptoconidia curvata, 10·5–16·8 × 0·7–0·9 μm; thallus acidum isousnicum continens.
Typus: Italy, Trentino-Alto Adige, Bolzano, 54 km WSW of Merano, NP Stilfser Joch, road Prad-Stilfserjoch, SW of Trafoi, SE slope with Picea forest and outcrops, 1850 m, on Pinus stump, 14 August 1990, P. van den Boom 10793 (B—holotypus; hb. v.d. Boom—isotypus).
(Figs 1D & 6)

Fig. 6. Lecanora pseudosarcopidoides, habitus (holotype). Scale: 0·5 mm.
Thallus endoxylic to epixylic, very thin, sometimes slightly rimose areolate, uneven to sometimes slightly warted, warts often scattered, sometimes contiguous, patches up to 4 cm wide, up to 130 μm thick; without cortex, granules present only in the upper layer of the thallus; upper surface grey, yellowish grey, or pale to brownish grey, smooth. Photobiont chlorococcoid, algal cells scatteredover the thallus. Prothallus not observed.
Apothecia appressed, broadly sessile, becoming subglobose, c. 0·4–0·9 mm diam.; margin at first strongly prominent, 0·04–0·07 mm wide, concolorous with thallus or paler, becoming darker and concolorous with the disc, smooth, evanescent, apothecia finally biatorine; disc at first flat, later convex, pale pinkish to pale orange, pale to intensively red-brown to dark brown or greyish, epruinose to slightly white or bluish pruinose. Thalline excipulum with cortex, variable in development, at first thin but becoming thick (20–35 μm), chondroid, algae present in outermost parts, finally algae only present at base of excipulum and up to c. 13 × 16 μm diam.; I ± blue to violet. Parathecium slightly widened at the outer part, closely connected to the cortex, to 30 μm wide at the outer rim. Epihymenium yellowish, yellowish brown, to medium greyish brown, with many coarse, yellowish granules, N+ reddish violet. Hymenium hyaline, with orange-brown granules, 40–50 μm high. Hypothecium hyaline to pale yellowish, 60–90 μm high. Paraphyses simple, branched or more rarely anastomosing, ± septate, rarely constricted at the septum, 1·5–2 μm diam., apices not or slightly widened to 3·5 μm, hyaline to sometimes more or less brown to olive greyish pigmented. Ascospores ellipsoid, simple, rarely 1-septate, c. 8·6–11·3 × 3·4–4·7 μm.
Micropycnidia not rare, immersed, greenish brown at upper part, hyaline below, c. 40–50 μm diam.; microconidia ± allantoid, simple to sometimes 1-septate, always weakly curved, 5·3–7·4 × 1·0–1·3 μm diam. Mesopycnidia not rare, immersed, upper part pale to brown or dark blackish olive, (80–) 100–180 × 90–150 μm; mesoconidia bacilliform to weakly curved, 4·4–6·3 × 1·4–2·2 μm. Leptopycnidia rather rare, immersed to semi-immersed, often on the margin or hymenium of apothecia, upper part hyaline, brown or greenish blue-black, 80–100 × 80 μm; leptoconidia curved, c. 10–17 × 0·7–0·9 μm diam.
Chemistry. K−, C−, KC−, P−. Isousnic acid (major) (TLC).
Ecology and distribution. Lecanora pseudosarcopidoides has been found on wood, mostly on the rotting trunks of conifers (Larix, Pinus cembra) mainly in the subalpine zone, at (800–)1400–2200 m, in poorly developed lichen communities, but nearly always associated with Parmeliopsis ambigua.
Notes. This new species is not rare in central Europe and has most probably been much overlooked because we did not find any reference to it in the literature or on herbarium specimens. It can easily be confused with L. saligna in poor condition, but L. saligna has different conidia (see Table 1), darker, reddish brown apothecia, with a more prominent margin and is distributed mainly in lowland areas. It is also very similar to poorly developed L. sarcopidoides, but the latter has a different chemistry, shorter and smaller ascospores, a better developed apothecial margin, smaller mesoconidia and is distributed in both lowland and montane areas. Lecanora pseudosarcopidoides is easily mistaken for a poorly developed L. albellula which has similar apothecia and pruina colour, but the thallus in the new species is much thinner and mostly endoxylic and never corticolous as are most specimens of L. albellula. Furthermore, macroconidia have never been found in L. pseudosarcopidoides whereas in L. albellula they are conspicuous, 1-septate and curved; the mesoconidia are larger in the former. The new species differs from L. mughicola by its paler apothecia with a much thinner pale yellowish margin becoming subglobose and subbiatorine, the presence of mesoconidia, a less well-developed thallus, shorter ascospores and a preference for more rotten wood. The I+ blue reaction of the excipulum has been found in one specimen only (P. v. d. Boom 13464). Several specimens of this species have been found labelled as L. subintricata. However L. subintricata has usnic acid as a major compound and also has much smaller ascospores. Two collections, M. Brand 24827 and P. v.d. Boom 9847, are strongly infected by Lichenodiplis lecanorae (Vouaux) Dyko & D. Hawksw.
Selected specimens examined. France: Puy de Dome: Mts du Forez, E of Col de Chamboite, 3°48′E, 45°40′N, 1480 m, branches of Fagus, ‘Krumholz’, rel.1208, 1976, M. Brand 6510 (hb. Brand). Savoie: 3·5 km NE of Col du Galibier, Valloirette S of Plan Lachat, 6°26.4′E, 45°5.1′N, 2000 m, Rhododendron ferrugineum, near brook in open valley, 2002, M. Brand 45506 (hb. Brand). Alpes Maritimes: 1·6 km ESE of Col de Cayolle, 6°45.9′E, 44°15.0′N, 2000 m, Larix in open wood near tree line, 2002, M. Brand 45332 (hb. Brand). Alpes-de-Haute-Province: 3·7 km NNW of Col de la Bonette, near Torr. de Clapouse, 6°47.1′E, 44°21.3′N, 2100 m, sandstone boulders at shore of brook, 2002, M. Brand 45069 (hb. Brand). Savoie: c. 66 km S of Mt Blanc, 1·5 km SW of Bessans, 6°59′E, 45°18′N, 1760 m, on Larix in open wood at foot of rock face, 1984, M. Brand 11726 (hb. Brand). Hautes Alpes: c. 27 km SE of Briancon, near Fontgillarde, 6°52′E, 44°43′N, 1970 m, on decorticated wood of Larix in open wood on N-slope, 1984, M. Brand 11674 (hb. Brand). Pyrénées Atlantiques: 46km SW of Pau, 4·2 km ENE of St. Engrâce, Col de St. Gracie, Serre de Benou, gently sloping Fagus forest along road, on decorticated rotting trunk (1 m tall), 3°26.7′W, 47°47.2′N, 1350 m, 1992, P. van den Boom 13464 (hb. v.d. Boom).—Switzerland: Valais: val d'Hérens, 2·6 km SSW of Haudères, 7°29.7′E, 46°3.8′N, 1700 m, wood of tree trunk washed ashore along mountain stream in narrow valley, 1990, M. Brand 24918 (hb. Brand); 0·4 km SW of Evolène, 7°29.5′E, 46°6.6′N, 1380 m, wood of hay-shed in meadow, 1990, M. Brand 24566 (hb. Brand); 1·2 km S of Evolène, 7°29.5′E, 46°6.7′N, 1680 m, in forest, wood of trunk of Larix, 1990, M. Brand 24827 (hb. Brand); 1·4 km S of Evolène, 7°29.5′E, 46°6.0′N, 1760 m, dead trunk in Larix forest, 1993, M. Brand 29705b (hb. Brand); 0·7 km N of Arolla, 7°27.3′E, 46°1.7′N, 2120 m, decayed wood of dead log in open Larix-Pinus cembra wood on E slope, 1990, M. Brand 24876b (hb. Brand); E of Evolène, E facing exposed meadow with calcareous schistose outcrops and barns, on vertical W-exposed wood, 1400 m, 1990, P. van den Boom 9847 (hb. v.d. Boom).—Austria: Tirol: SE of Innsbruck, Patscherkofel, between Bosschede and Isshütte, 1900 m, open forest on N slope, on dead wood of Pinus cembra, 1981, M. Brand 31811 (hb. Brand). Karnten: Gailtaler Alpen, 12 NW of Hermagor, near Weiss-briach, on wood of bird-observation post, 820 m, 1994, P. van den Boom 15728 (hb. v.d. Boom).—Spain: Lérida: Pireneos, Aigües Tortes national park, S-side of lake S. Mauricio, 1°2′E, 42°34.7′N, 1940 m, on old, free standing Abies on N-slope, 1983, M. Brand 32762 (hb. Brand).—Italy: Trentino-Alto Adige: Bolzano, 54 km WSW of Merano, NP Stilfser Joch, road Prad-Stilfserjoch, SW of Trafoi, SE slope with Picea forest and outcrops, on stump, 1850 m, 1990, P. van den Boom 10824 (hb. v.d. Boom); ibid., Pinus forest with stumps, on Pinus stump, P. van den Boom 10790 (hb. v.d. Boom). Alpi Dolomitiche: c. 1 km S of Marmolada, SW of barrage, 11°51′E, 46°27′N, c. 2100 m, wood of dead trunk of Pinus cembra on open N-slope with limestone and shrubs, 1994, M. Brand 31632, 31633 (hb. Brand).
Lecanora subsaligna M. Brand & van den Boom sp. nov
Species lignicola vel corticola, Lecanorae salignae similis, sed thallo minore, inconspicuo, ad 3 cm lato, 20–150 (−200) μm crasso, ascosporae 0(−1)-septatae, 8·4–11·5 × 3·3–4·0 μm; macroconidia (19–)22–27(−30) × 1·7–2·0 μm, valde curvata (c. 45–90°), apicibus attenuatis (falcata), 1-septata, nonnumquam ad 3-septata, non rara; microconidia c. 8–10 × 0·9–1·0, paulo curvata; leptoconidia tenues, longitudine variabiliore, c. 13–22 × 0·8–0·9 μm, curvata.
Typus: Belgium, Namur, SSW of Beauraing, WSW of Gedinne, 300 m, E of Croix Scaille, open area with many big stumps (K5.26), on Vaccinium, 500 m, 2 May 2000, P. & B. van den Boom 24408 (LG—holotypus; hb. van den Boom, hb. Brand—isotypus).
(Figs 1A & 7)

Fig. 7. Lecanora subsaligna, habitus (holotype). Scale: 0·5 mm.
Thallus corticolous or lignicolous, often endophloeodal, relatively small, patches up to c. 3 cm wide, mostly much smaller, thin, c. 20–150(−200) μm high, uneven to slightly verrucose, continuous, inconspicuous; without true cortex, with epinecral layer, often abundantly filled with granules at the upper surface, also with epipsamma on upper surface and at the inside of the ‘cortex’ and then spotted by anticlinal streaks, granules ± finely to coarsely granular, 0·2–3 μm wide, sometimes granules only present near upper surface around the apothecia and pycnidia; algal layer not differentiated, algal cells filling the whole thallus; upper surface grey or withish to pale grey, yellowish grey, or pale grey-orange, smooth, matt to somewhat shiny, some specimens being overgrown by algae. Photobiont chlorococcoid, cells 5–15 μm diam. Prothallus not observed.
Apothecia appressed to sessile, rounded to rarely irregularly angular in outline, slightly constricted at the base, (0·2–)0·3–0·5 mm wide, scattered to densely crowded; margin distinct, rather small, initially thick and clearly prominent, finally thin and level with the disc, 0·03–0·07 mm wide, sometimes becoming excluded, concolorous with the thallus, entire or sometimes irregularly incised; disc pale beige, pale orange to orange-brown, pale brown or pale reddish brown, rarely darkened, flat to slightly or sometimes moderately convex, epruinose. Thalline excipulum only corticated in lower part, 20–30 μm thick, at least partly, chondroid, cells with thicker, conglutinated margins and inconspicuous lumina of c. 2 × 4 μm, filled with many yellow-brown to orange-brown, fine granules, algae 15–18 × 11–14 μm, with haustoria. Parathecium not or weakly developed, up to 10 μm wide in lower part, up to 25 μm wide at outer rim, strongly conglutinated with elongate cells, containing yellowish brown to orange-brown, fine granules. Epihymenium yellowish, yellowish brown or medium to dark brown, abundantly filled with fine granules, N+ reddish violet. Hymenium hyaline, with yellow-brown to orange-brown granules often present butinconspicuous and scattered, 35–50(−55) μmhigh. Hypothecium hyaline, of unoriented hyphae, sometimes with pseudoparaplectenchymatous cells, especially in lower parts, up to 80 μm high. Paraphyses ± conglutinated, simple to often branched, non-septate, 1–2 μm wide, sometimes slightly widened at the tips and rarely up to 5 μm wide, tips hyaline to rarely brown pigmented. Asci clavate to pyriform, Lecanora-type, 10–15 × 20–35 μm. Ascospores ellipsoid to oblong, 0(−1)-septate, c. 8·4–11·5 × 3·3–4·0 μm.
Macropycnidia not common, immersed, or top somewhat exposed, upper part hyaline to brown-pigmented, lower part hyaline, c. 70–110 (high) × 50–100 μm; macroconidia strongly curved (c. 45–90°), attenuate at both apices (falcate), 1(−3)-septate, (19–) 22–27(−30) × 1·7–2·0 μm. Micropycnidia rare, c. 50 μm diam.; microconidia slightly curved, c. 8–10 × 0·9–1·0. Leptopycnidia not rare, c. 70–80 × 50–60 μm; leptoconidia thin, rather variable in length, curved, c. 13–22 × 0·8–0·9 μm.
Chemistry. K−, C−, KC−, Pd−. Isousnic acid (major), 7-O-methylnorascomatic acid (traces and very rare); many specimens tested by TLC or HPLC.
Ecology and distribution. Lecanora subsaligna is weakly acidophilous and clearly a pioneer species. It is known from a wide range of substrata, such as bark of trunks and branches of Sambucus, Salix in woodlands, on Quercus in a wide range of habitats, on Sarothamnus in dune areas, on branches of shrubs (Vaccinium), and also on decorticated trunks, wooden constructions and fence posts. The holotype on Vaccinium is accompanied only by Scoliciosporum umbrinum, which has just a few apothecia. It has been found mainly in western Europe, with the most eastern collection from Italy. In the Benelux countries this species has sometimes been identified as Lecanora piniperda (= L. albellula) and was also recorded under this name in Diederich & Sérusiaux (Reference Diederich and Sérusiaux2000) and Sérusiaux et al. (Reference Sérusiaux, Diederich, Ertz and Boom2003). However, all these specimens were misidentified.
Notes. Lecanora subsaligna occurs in similar habits as L. albellula, but differs in its conidia (for measurements see Table 1), somewhat smaller ascospores, thinner margin of the apothecia, less warted thallus and an epruinose apothecial disc. It has been found more often in lowland areas than L. albellula. It is easily mistaken for L. hagenii, small and not well-developed L. saligna, or a Lecania species. The only specimen from the USA is rather small, so that it is difficult to confirm its identity, but we found a small amount of isousnic acid and the characteristic, falcate macroconidia. Several specimens, mainly from southern Europe, have a somewhat similar habitus, but even after extensive study we could find only leptoconidia, so that we are not sure of their status and have excluded them here from L. subsaligna.
Selected specimens examined. Norway: Sogn-Fjordane: Askvoll, Fureneset, near Planteskole, 5°2.79′E, 61°17.58′N, large trees in garden, near coast, on Acer, 2003, M. Brand 48107 (hb. Brand).—Denmark: Mandoe: village, N part, 8°32.2′E, 55°16.6′N, on wood of poles on dune, 1977, M. Brand 7126b (hb. Brand).—The Netherlands: Utrecht: Darthuizen, grid ref. 39–13; in Pini sylvestri, s.d. (19th century), H. C. van Hall H254 (NBV). Friesland: Vlieland, Kooislid, 5°2.4′E, 53°17.6′N, on dead Salix repens in dune area, 1977, M. Brand 6374 (hb. Brand); 6 km WNN van Winschoten, Midwolda, 6°59.3′E, 53°11.8′N, on apple tree in garden, 1991, M. Brand 25865 (hb. Brand). Groningen: 26 km NE of Groningen, Oosteinde, near churchyard, on Acer, 1993, P. van den Boom 15191 (hb. v.d. Boom). Drenthe: Spier, (Beilen), 6°28.0′E, 52°49.1′N, on Tilia along road in front of hotel, 1976, M. Brand 3331 (hb. Brand). Flevoland: Oostelijk Flevoland, Roggebotzand, 5°48.6′E, 52°34.1′N, on Salix (20 years old), 1981, M. Brand 9562b (hb. Brand); Dronten, Abbert (W side), 5°49.7′E, 52°29.6′N, on old Salix alba in open woodland with dense low shrubs of a.o. Rubus, 1991, M. Brand 26665b (hb. Brand); Zuidelijk Flevoland, N van Kitsweg, 1·2 km WSW van Knardijk, 5°24.3′E, 52°27.1′N, Salix woodland with Salix alba, S. triandra and S. alba, 1990, M. Brand 22529 (hb. Brand); woodland near pumping engine “Block van Kuffeler”, 5°14.1′E, 52°25.1′N, on young Salix in swamp, 1985, M. Brand 12636 (hb. Brand).—Belgium: Liège: Hoge Venen, S side of Soor valley, on decaying Picea stump in young reafforestation, 500 m, 1978, H. Sipman 10270 (B). Luxembourg: 2 km SE of Bertrix, 5°17′E, 49°50′N, 400 m, at side of road, open, among fields, on Tilia, 1989, M. Brand 22272 (hb. Brand).—Luxembourg: Oesling: 2 km WNW of Troisvierges, Bois de Biwisch, Picea forest (J8.22), 475 m, on young Acer, 1992, P. van den Boom 12574 (hb. v.d. Boom); 11 km WSW of Esch sur Sure, Moulin de Bigonville, SW side of Sure, 5°48′E, 49°52′N, 325 m, branches of Sarothamnus on S-exposed rocky ridge, 1992, M. Brand 27193 (hb. Brand); Hoscheid, Banzenhuuscht, road to Goebelsmuhle, roadside trees, on Tilia, 480 m, 1995, P. van den Boom 16813 (hb. v.d. Boom); W Hoffelt, Kalebur (J8.31), on wood, 1987, P. Diederich 7778 (hb. Diederich). Gutland: Mamer, Tossebierg (M8.14), on Sambucus, 1997, P. Diederich 12876 (hb. Diederich). Lorrain: W Steinfort, berge de l'Eisch (L8.51), 1984, Diederich 5165, (hb. Diederich).—Germany: Niedersachsen: Waddensea island Juist (E-side), near airfield, 7°2.8′E, 53°41.2′N, Sambucus on S-slope in dune area, 1977, M. Brand 7006 (hb. Brand); Norderney, near Postbake, 7°16.6′E, 53°42.9′N, Sambucus in dune area, 1977, M. Brand 6943b (hb. Brand). Brandenburg: W of Berlin, Döberitzer Heide (nature reserve), open grassy heathland, on horizontal surface of wood of fence post, 50 m, 2004, P. van den Boom 32077, 32079, 32080 & H. Sipman 51568, 51573 (B, hb. v.d. Boom).—Czech Republic: Ústecký Kraj: N of Prague, Lužicke Hory, NE of Diěčín, Národní park, České Švícarsko, E of Hřensko, N of Vysoká Lípa, unpaved road along field and open places in woodland, with sandstone outcrops, on branch of Quercus, 350 m, 2005, P. van den Boom 34750 (hb. v.d. Boom).—France: Pyrénées Occidentales: St. Engrâce, Arpideko Ibarra, near Sc. d'Harricagna, 0°47.6′W, 42°59.4′N, 820 m, Fagus trees in open wood on N-slope, on rotting wood on soil, 1992, M. Brand 28033 (hb. Brand). Cantal: W of Murat, 3 km NW of Puy Mary, Abies Fagus forest, on Abies, 1140 m, 1998, P. van den Boom 21065 (hb. v.d. Boom).—Italy: Liguria: c. 5 km NW of Monte Penna, at road to Rezzaglio, 9°26′E, 44°30′N, c. 1000 m, Quercus, scattered in hay fields, 1993, M. Brand 30095b (hb. Brand).—Portugal: Tras-os-Montes: SW of Mogadoura, road (315) from Meirnhos to Estiais, near Quinta das Quebradas, Olea orchard, on wood of Olea trunk, 500 m, 1999, P. van den Boom 22810 (hb. v.d. Boom); Serra do Gerês, NE of Cabril, 2·5 km N of Zertelo, near Lapela, E slope with a few mature Quercus robur in the field, on exposed root of Quercus, 650 m, 1999, P. van den Boom 23288 (hb. v.d. Boom). Minho: ENE of Braga, Povoa de Lanhoso, monumental castle with mixed trees in the surrounding, on Olea europaea, 380 m, 2005, P. van den Boom 35055 (hb. v.d. Boom). Beira Alta: NW of Viseu, S of Sao Pedro do Sul, Termas de S. Pedro do Sul, centre of village, Querus and Corylus along parking lot, on Quercus, 195 m, 2005, P. van den Boom 35441 (hb. v.d. Boom); Serra da Estrela, W of Manteigas, south route to Penhas Dousadas, small road with slope and mixed trees, on young Castanea tree, 1100 m, 1997, P. van den Boom 19016 (hb. v.d. Boom). Alentejo: Serra de Mamede, Natural Park, SE of Marvão, N of Pitaranha, near border of Spain, E slope with exceptionally big granite outcrops, on Rubus, 540 m, 2003, P. van den Boom 31389 (hb. v.d. Boom); NE of Cercal, Barragem de Campillhas, shaded slope with schistose outcrops, near bridge, on Quercus suber, 125 m, 1997, P. van den Boom 19176 (hb. v.d. Boom).—Spain: Lérida: Pireneos, Aigües Tortes natural park, S-side of lake S. Mauricio, 1°0.2′E, 42°34.7′N, 1940 m, on old, free standing Abies on N-slope, 1983, M. Brand 32761 (hb.Brand). Extremadura: 41 km ENE of Placencia, N of Jarandilla, c. 2 km N of Guijo de Santa Bárbara, trail along river ‘Jaranda’, on Sarothamnus, 1400 m, 1989, P. van den Boom 8873 (hb. v.d. Boom). Mallorca: ENE of Soller, Sierra Torrellas, E of Son Torella, meadow with calcareous outcrops and mature Pinus trees, 975 m, 1997, P. van den Boom 18591 (hb. v.d. Boom).—USA: California: San Luis Obispo, S of Morro Bay, State Park road, N of Museum of Natural History, among mature Cupressus macrocarpa trees beside beach, on stump, on sloping sawn surface, 2 m, 2002, P. & B. van den Boom 29447 (hb v.d. Boom).
Descriptions of some related species
Lecanora albellula Nyl. var. albellula
Not. Sällsk. Fauna Fl. Fenn. Förh. 8: 147 (1866); type: Russia, Lapponia Inandrae, prope Kantalaks, 1863, Fellman 129 (H-Nyl.26472—lectotype!).
Lecanora piniperda Körb., Parerg. Lich. 81 (1859), nom illeg.
(Fig. 8A)
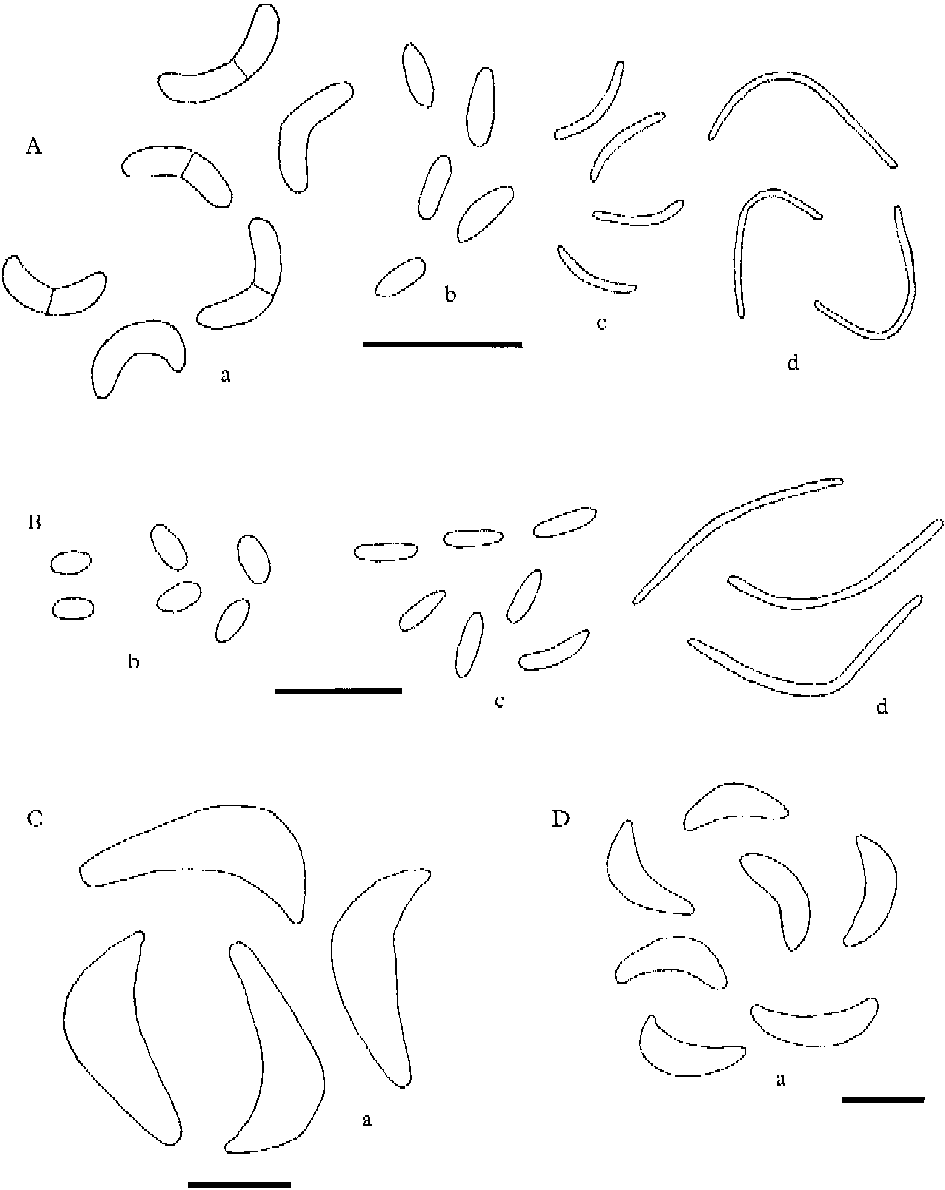
Fig. 8. Lecanora saligna group, conidia. A, L. albellula; B, L. sarcopidoides; C, L. subravida; D, L. saligna. a=macroconidia; b=mesoconidia; c=microconidia; d=leptoconidia. Scales: A & B=10 μm; C & D=5 μm.
Thallus small to effuse, up to 4 cm wide, mostly smaller, often forming continuous, warted patches, warts 0·15—0·2 mm wide, rounded to sublobate, convex to semiglobose, sometimes very thin to endoxylic, 50–600 μm thick; cortex not clearly developed, thallus filled with granules, mainly at the upper layer and in patches or only near apothecia and pycnidia; upper surface whitish grey, pale grey to brownish grey, matt to more or less shiny. Photobiont chlorococcoid, algal cells 8–20 μm diam., scattered throughout the thallus. Prothallus not detected.
Apothecia situated in the centre of the thallus or dispersed, sessile and often constricted at the base, c. 0·25–0·55°–0·7) mm diam.; margin relatively thick, initially prominent above the disc, becoming thin and level with the disc, concolorous with the thallus, entire to crenulate, to 0·08 mm wide; disc plane to convex, becoming subglobose, then with excluded margin, pale beige to dark grey, pale to dark greyish brown or pale to reddish brown, slightly to densely whitish to pale yellowish pruinose. Thalline excipulum with a cortex which is best developed in the lower part; chondroid, c. 10–30 μm thick, with thick, conglutinated cell margins, lumina not clear, often reduced, algae c. 14–18 × 10–12 μm, with haustoria, with granules at the outer surface of the cortex and in anticlinal stripes. Parathecium sometimes visible, small, to 30 μm wide and brownish pigmented at the outer rim. Epihymenium yellowish, pale brownish to greyish brown, interspersed with fine granules of c. 3 μm, N+ reddish violet. Hymenium hyaline, with granules mainly in the upper part, c. 35–55 μm high. Hypothecium hyaline to pale yellowish, to 90 μm high. Paraphyses simple, branched to sometimes anastomosing, ± septate, rarely constricted at the septum, in a gelatinous matrix, 1·5–2 μm wide, top often slightly widened to 4(−5) μm, mainly hyaline, or with a brownish, bluish grey or greenish blue outer tip. Ascospores ellipsoid to broadly ellipsoid, simple, rarely 1-septate, 8·5–11·5 (−12·5) × 3·1–4·2 μm.
Macropycnidia often present but not abundant, immersed or semi-immersed in the thallus or on the apothecium (epihymenium as well as margin), top brown or hyaline, c. 70–120 × 60–100 μm; macroconidia slightly curved (− 45°) with roundish (obtuse) tips, 1-septate, rarely to 3-septate, c. 7–10·5 ×1·8–2·2 μm. Micropycnidia not rare, immersed, c. 50 μm; microconidia slightly to strongly curved, simple, 5·6–9·8 × (0·8–) 0·9–1·3 μm. Mesopycnidia rare, small, mostly with a brown pigmented top; mesoconidia ellipsoid-bacilliform, straight with rounded apices, 2·9–5·1 × 1·6–2·1. Leptopycnidia rare, immersed; leptoconidia rather variable in length, curved, 10–15 × 0·7–1 μm.
Chemistry. K−, C−, KC−, P−. Isousnic acid (± major), ± usnic acid (trace); rarely 7-O-methylnorascomatic acid (minor) and only in apothecia (TLC & HPLC).
Ecology and distribution. On the bark of trunks and branches of a wide range of phorophytes such as Acer, Corylus, Fagus, Larix, Picea, Pinus, Salix and Sambucus, as well as on decorticated trunks and more rarely on wood and timber. This species is widely distributed in western Europe, from northern Scandinavia (Santesson et al. Reference Santesson, Moberg, Nordin, Tønsberg and Vitikainen2004) to northern Spain. We have seen several exsiccate specimens from central Europe.
Notes. Lecanora albellula s. str. is closely related to L. albellula var. macroconidiata but they differ principally in the size of their macroconidia; 1(–3)-septate and 7–10·5 × 1·8–2·1 μm in the former and 1–3-septate, 12·4–14 × 1·8–2·3 μm in the latter. Further differences are discussed above under L. albellula var. macroconidiata.
Lecanora albellula s. str. can be distinguished from L. saligna by its more dispersed thallus warts and apothecia which are often aggregated in the centre of the thallus; its macroconidia, microconidia and chemistry are also different. Although both species contain isousnic acid as a major substance, usnic acid and 7-O-methylnorascomatic acid (rare in L. albellula s. str.) have never been found in L. saligna. L. albellula s. str. differs from L. subsaligna by its warted thallus, strongly convex apothecia and smaller macroconidia (much longer and falcate in L. subsaligna). The dark greyish brown, often slightly pruinose, young flat, as well as strongly convex old, apothecia are also typical. We have studied the very scanty (lecto)type specimen and, of the four known types of conidia, have found only macroconidia and mesoconidia. Their measurements agree well with other specimens studied in the literature. While in the most recent and extensive description (Printzen Reference Printzen2001) pycnidia are not mentioned, they are recorded as 7–14 ×1–2 μm (Clauzade & Roux Reference Clauzade and Roux1985); as macroconidia, 7–10 × 1–2 μm, and microconidia, 3–5 × 1 μm (Hawksworth & Dalby Reference Hawksworth, Dalby, Purvis, Coppins, Hawksworth, James and Moore1992); as macroconidia, 7–14 × 1–2 μm (curved), or microconidia 3–5 × 1 μm (straight) (Wirth Reference Wirth1995); and 7–11 × 1 μm (Hinteregger Reference Hinteregger1994), all without remarks about septation. This variation, most probably, can be explained by the many misidentifications we have found during our study. We have studied various collections of ‘L. piniperda’ and found several to belong to L. saligna, L. subsaligna, L. subintricata or another genus, including those recorded from Luxemburg and Belgium (Sérusiaux et al. Reference Sérusiaux, Diederich, Ertz and Boom2003). Thus in our opinion, the occurrence of L. albellula in Belgium is not confirmed.
In an investigation of more than ten specimens treated by Hinteregger (Reference Hinteregger1994: 221) as Lecanora sp. 1 (aff. saligna), in which isousnic- and usnic acid were present, the same three types of conidia as described from L. albellula s. str. were found, so that we are convinced that this material belongs in this species. This material is mainly from bark of Rhododendron ferrugineum or more rarely from R. hirsutum.
Selected specimens examined. Great Britain: Scotland: V.C. 105, West Ross: Torridon, Rubha, na h-Airde Glaise, coast to north, 18/80.56, on lignum of branch, of Salix, 1994, B. J. Coppins 16454, A. M. Fryday & A. M. O'Dare (E). England: V.C. 57, Derbyshire: Chatsworth Old Deer Park, 43/26.68, on Quercus stump, 1991, B. J. Coppins 15669 & O. L. Gilbert (E).—The Netherlands: Friesland: Terschelling, Oosterend, Kopjesplak, Betula forest, SO of ‘Jan Thijssenduin’, 5°25.1′E, 53°24.6′N, on mature Betula in open woodland, 1997, M. Brand 35987 (hb. Brand); Vlieland, Kooislid, 5°2.4′E, 53°17.6′N, dead Salix repens in dune area, 1977, M. Brand 6374, 6375 (hb. Brand). Flevoland, Noordoostpolder, Kuinderbos (N side), on wood of barrier, 1979, M. Brand 7943 (hb. Brand); Oostelijk Flevoland, Roggebotzand, 5°48.6′E, 52°34.1′N, on Salix (20 years old), 1981, M. Brand 9562 (hb. Brand); Zuidelijk Flevoland, N of Kitsweg, c. 1·2 km WSW of Knerdijk, on old Salix triandra in woodland, 1990, M. Brand 22540, 22552 (hb. Brand).—Germany: Niedersachsen: Norderney, NE of Grohde, on Sambucus in dune area, 1977, M. Brand 6945 (hb. Brand); Hohes Venn, W of Monschau near Belgian border, 6°10.8′E, 50°33.3′N, 600 m, wood of Picea trunks, in felled woodland, 2004, M. Brand 50489 (hb. Brand).—France: Cantal: SW of Murat, Albepierre, W side of village, campsite, trees along stream, and meadow, on wood of fence post, 1050 m, 1998, P. van den Boom 20626 (hb. v. d. Boom).—Switzerland: Valais: S of Evolène, SE of Les Hauderes, along La Borgne, N-facing slope with Acer in meadow and well-lit schist, on trunk of Acer, 1540 m, 1990, P. van den Boom 10071 (hb.v. d. Boom); Val d'Hérens, 0·7 km N of Arolla, 7°27.3′E, 46°1.7′N, 2120 m, twigs of Larix in open Larix-Pinus cembra wood on E slope, 1990, M. Brand 24880 (hb. Brand); SW of Brig, SSE of Visp, Visperterminen, 1 km E of village, path W of small lake, to Simplonpass and Gibidumpass, 2050 m, on shrub, on vertical bank of path, 1996, P. van den Boom 17803 (hb. v. d. Boom); Graubünden, Engadin, Val Susauna, 9°59.5′E, 46°38.9′N, 1690 m, isolated Larix near stream in open valley, 1977, M. Brand 6687, 6690 (hb. Brand).—Austria: Tirol: SW von Mittelberg im Pitztal, Taschach-alpe, Kraftwerk am Bach, oberhalb der Strasse Richting Taschachferner, NW-Hang, MTB 9031/1, on Rhododendron ferrugineum, c. 1810 m, 8 viii 1986, E. Hinteregger s.n. (GZU); Ötztaler Alpen, Platzal, im Zirben wald S von Obergurgl, N-Hang, MTB 9132/1, on R. ferrugineum, 2000 m, 1 vii 1986, E. Hinteregger s.n. (GZU). Steiermark: Schladminger, Tauern, Kleinsölktal-Obertal, Lemperkar über der Putzentalalm, on R. ferrugineum, 1830–1880 m, 17 vii 1986, H. & M. Mayrhofer & Ch. Scheuer s.n. (GZU). Kärnten: Karnische Alpen, 4 km S of Hermagor, 1 km WNW of Egger Alm, forest on SW slope, on wood of dead branch of Acer, 13°22.1′E, 46°35.2′N, 1350 m, M. Brand 31534 (hb. Brand).—Spain: Lérida: Pireneos, Aigües Tortes National Park, S-side of lake S. Mauricio, 1°2′E, 42°34.7′N, 1940 m, on old, free standing Abies on N-slope, 1983, M. Brand 32761 (hb. Brand).—Italy: Parma: NE of Passo del Pelizzone, 9°42′E, 44°40′N, 1050 m, Corylus in scrub on limestone near mountain ridge, 2001, M. Brand 42348 (hb. Brand). Calabria: Sila, N of Lago Arvo, Silvana Mansio, 16°32′E, 39°18′N, 1475 m, on Pinus, 1986, M. Brand 52926 (hb. Brand).—Portugal: Alentejo: SE of Sines, NE of Cercal, Barragem de Campilhas, near bridge, field with Cistus ladanifer and schistose outcrops and Quercus suber, on Q. suber, 125 m, 1997, P. van den Boom 19176 (hb. v. d. Boom). Algarve: NE of Albufeira, road Benafim Grande to Salir, 3 km W of Salir, Olea orchard along road, on decorticated trunk of Olea, 400 m, 1993, P. van den Boom 14360 (hb. v. d. Boom).—USA: New Hampshire: Coos County, White mts, nat. forest, Presidential Dry River Wilderness Area, one mile SW of Mt. Crawford along Davis path, at edge of ridge with balsam fir, birch and black spruce, 2600 ft., 1988, C. Wetmore 61973 (GZU). Colorado: Boulder county, one mile west of Ward, along Lefthand Canyon, east of red rock Lake, Spruce-fir forest, 9800 ft, 1967, C. Wetmore 16022 (GZU). California: Tulare, Sequoia National Park, ridge S of Little Badly, on E facing slope of ridge in fir and pine forest with some incense cedar, on conifer log, 6800 ft, 1984, C. Wetmore 51091 (GZU).
Exsiccata examined: Arnold, Lich. exs. 665 (L, GZU); Arnold, Lich. Mon. exs. 235, 380, 472, 515 (L); Körber, Lich. Sel. Germ. 215 (BR); Magnusson, Lich. sel. scan. exs. 403 (B); Malme, Lich. suec. exs. 435 (B); Rabenhorst, Lich. Europ. 640 (BR); Vězda, Lich. sel. exs. 2189 (B, LG); [all as Lecanora piniperda]; Rabenhorst, Lich. Europ. 784 (BR) [as Lecanora varia var. glaucella].
Lecanora hypoptoides (Nyl.) Nyl
Flora 55: 249 (1872);—Lecidea hypoptoides Nyl., Flora 50: 371 (1867); type: Finland, Tavastia, 1866, J. P. Norrlin 507 (H-Nyl. 26529—holotype!).
(Fig. 9)

Fig. 9. Lecanora hypoptoides, habitus °Equipo 1878 (MAF). Scale: 0·5 mm.
Thallus corticolous or lignicolous, epi- to endosubstratal, effuse, in patches of up to 2 cm wide, sometimes aggregating, sometimes diffusely covering larger surfaces, warted, warts ± subglobose and to 0·25 mm diam., sometimes disintegrated and nearly sorediate, c.100–350 μm thick; cortex poorly developed, not clearly differentiated, filled with minute oil droplets (<0·4 μm diam.), without granules; epinecral layer poorly developed, to 10 μm high; algal layer or medulla not differentiated; algal cells covered by relatively thick hyphae of 2·5–5 μm and thick walled, surface of hyphae covered with a fine granular substance not or weakly visible in polarized light and soluble in KOH and acetone; upper surface pale grey to dark grey, pale yellowish brown to moderately brown, matt to slightly shiny. Photobiont chlorococcoid, cells 8–20 μm and thick walled. Prothallus not developed.
Apothecia appressed to sessile, sometimes constricted at the base, 0·4–0·6 mm diam.; margin level with the disc or slightly raised, entire to irregular or crenulate, often becoming excluded, concolorous with the thallus, 0·04–0·12 mm wide; disc plane to slightly convex, rarely subglobose, pale or reddish brown, dark greyish brown to dark brown, epruinose. Thalline excipulum initially raised and conspicuous, becoming excluded; in section cortex mainly visible at the lower part of the apothecia, to 20–35 μm wide, with minute granules; algal layer clearly differentiated, to 90 μm high; ± I+ bluish. Parathecium poorly to well developed, to 50 μm wide at the outer rim, pseudoparenchymatous cells sometimes visible. Epihymenium pale to medium reddish brown or medium to dark brown or brownish-spotted, sometimes slightly olivaceous, without granules, mostly with minute oil droplets (<0·4 μm diam.), N+ reddishviolet. Hymenium hyaline, with a few orange granules, 35–60 μm. Hypothecium hyaline, 45–55 μm high. Paraphyses mostly branched and anastomosing, ± septate, 2–2·5 μm widein the middle, the apices sometimes widened up to 4(−5) μm wide, hyaline to greyish dark brown. Asci Lecanora-type, broadly ellipsoid, 25–35 × 12–17. Ascospores ellipsoid, simple or rarely 1-septate, 9·6–13·3 × 3·1–4·0 μm.
Micropycnidia rather rare, inconspicuous, immersed, c. 50–80 μm diam.; microconidia bacilliform to slightly curved, 4·8–5·7 × 0·9–1·1 μm. Mesopycnidia sometimes abundant, immersed, globose, c. 150 μm diam., dark brown to bluish green in upper part, hyaline below; mesoconidia tear drop-shaped, 2·7–3·9 × 1·4–1·8 μm.
Chemistry. K−, C−, KC−, Pd−. Paraensic acid D (major), paraensic acid C (minor) (TLC and HPLC).
Ecology and distribution. Lecanora hypoptoides occurs on the bark of Castanea, Larix, Pinus and also on decorticated wood, often with Hypocenomyce scalaris and Pseudevernia furfuracea.
It is known from mountain areas in southern Europe (400–1800 m) and from boreal Europe (Santesson et al. Reference Santesson, Moberg, Nordin, Tønsberg and Vitikainen2004). According to Wirth (Reference Wirth1995) it is known from SW Germany and Switzerland (Haute Vienne).
Notes. Lecanora hypoptoides is easily mistaken for a member of the L. saligna group, such as L. albellula var. macroconidiata. However, this variety has a more warted, thinner and shiny, more yellowish brown thallus, which is sometimes sublobulate with more strongly convex (subglobose) warts, macropycnidia with relatively large macroconidia which are mostly 3-septate, and it contains 7-O-methylnorascomatic acid. The last two characters are lacking in L. hypoptoides. Lecanora hypoptoides can also be mistaken for L. albellula s. str. or L. sarcopidoides, but both these species often have (slightly) pruinose apothecia. Lecanora hypoptoides is somewhat similar in appearance to L. hypoptella (Nyl.) Grummann, but the latter species has smaller, darker brown apothecia and a thinner thallus. Although the mesoconidia in L. hypoptoides and L. hypoptella are somewhat similar, the microconidia, rather rare in both species, are clearly shorter in the latter. Lecanora hypoptella is a poorly understood species and is currently being revised (Z. Palice pers. comm.).
The chemistry and the rather uniformly coloured apothecial discs at all states of development are typical for L. hypoptoides. The holotype is in very poor condition, but the chemistry agrees with other specimens in good condition. Several exsiccate specimens of Lecanora ochrostoma and L. piniperda var. ochrostoma are referred to this species, but the type of L. piniperda var. ochrostoma belongs to Lecania fuscella (Printzen Reference Printzen2001). The collection by Equipo (MAF 1878), listed below, was originally identified as Lecanora sarcopidoides and is infected by a most probably undescribed Phoma sp., with pycnidia, 150–190 × 100–140 μm, and tear drop-shaped conidia, 3–4 × 1·6–2 μm.
Additional specimens examined. Norway: Oppland: Sel: Horgesetrene, W above Rosten, towards Mt. Horgensoi, 61°52′N, 9°22′E, c. 1000 m, Betula nana-Cladina-heath, on hill ridge just above tree limit, on isolated Betula tree trunk, 1999, H. Sipman 44164 (B).—Finland: Lapponia inarensis: parish Utsjoki: valley of Kevojoki river, 1–2 km before outlet into Lake Kevojärvi, W of Kevo Scientific Station, light mossy Betula forest in valley, on E-facing slope, on dead branchlets of Pinus tree, 100 m, 1988, H. Sipman 23155 (B).—Germany: Niedersachsen: Harz mts., along the Rehberger Graben, N of the Goetheplatz, Staatsfort Andreasberg, N of St. Andreasberg, MTB1. 4229/3, Picea forest with scattered Acer pseudoplatanus, on steep E-facing slope, on wood of Picea stump, 750 m, 1984, H. Sipman 17523 (B).—Italy: Calabria: Sila Greca, M. Altare (col in road Longobucco-Lago di Cecita), 16°33.3′E, 39°25.2′N, 1500 m, Fagus in wood on top of mountain ridge, 2001, M. Brand 42937 (hb. Brand).—Spain: Navarra: Pireneos, 3·5 km ENE of Isaba, Baranco de Belabarce, on Pinus, 1000 m, 1992, M. Brand 28538 (hb. Brand). Madrid: Serra Guadarrama, Pto. Cotos, on trunk of Pinus sylvestris, 1850 m, 1981, Equipo 1878 (MAF).
Exsiccata examined: Arnold, Lich. Mon. exs. 395, 434, 495 (B,L,M) [as L. piniperda var. ochrostoma or L. ochrostoma]; Hepp, Flecht. Eur. 387 (BR, M) [as L. ochrostoma]; Malme, Lich. suec. exs. 212 (B).
Lecanora mughicola Nyl
Flora 55: 248 (1872); type: Italy, ad nudos exsiccatorum Mughorum ramos in alpibus Rhaeticis. Anzi, Lich. Lang. 376 (H-Nyl. 26738—holotype; M—isotype).
Thallus epixylic to partly endoxylic, sometimes effuse, to 5 cm wide, or in smaller, ± delimited patches, with scattered warts to areolate, up to 0·3(−0·5) mm thick; areoles angular, to 2 mm wide, plane to convex; cortex up to 80 μm high, often poorly developed, with granules in upper part of the thallus, epinecral layer sometimes present, to 25 μm high; upper surface pale to dark grey or yellowish grey, smooth and somewhat shiny with a waxy appearance. Photobiont chlorococcoid, cells 8–17 μm, often forming a clearly differentiated layer. Prothallus not observed.
Apothecia abundantly present, broadly appressed, c. 0·7–1·1 mm diam.; margin conspicuous, persistent, 0·06–0·1 mm wide, concolorous with the thallus, smooth or warted, flexuose or sometimes crenulate, matt or slightly shiny; disc flat, initially pale yellowish brown, reddish brown or mostly dark blue-grey to intensively blackened, epruinose to rarely slightly greyish or pale bluish pruinose. Thalline excipulum corticate, with 15–35 μm wide cortex, rather equal in thickness or reduced at the top, chondroid or with many granules and then with an indistinct structure, algal layer clearly differentiated from the cortex; algae present to the upper outer rim, algae c. 16 ×11 μm. Parathecium slightly widened, at the outer rim to 40 μm, hyaline, pseudoplectenchymatous cells c. 2–4 μm. Epihymenium yellowish brown to olive-brown, with many, relatively large, yellowish granules, N+ reddish violet. Hymenium hyaline, with orange-brown granules, 50–70 μm high. Hypothecium, hyaline to pale yellowish, 60–100 μm high. Paraphyses branched and anastomosing, (1–)1·5–2 μm, abundantly septate, ± widened at the top, up to 6 μm diam., more or less brown or green to bluish-grey pigmented in upper cell. Ascospores ellipsoid, simple, 10–13·8 × 4·4–5·2 μm.
Micropycnidia inconspicuous, immersed, rare; microconidia bacilliform to slightly curved, 6–9 × 0·8–1·1 μm. Leptopycnidia usually present, immersed, brown or green blue-black above, hyaline below, 100–130 μm high × 60–110 μm; leptoconidia mostly strongly curved, not septate, 13–16 × 0·8–1 μm.
Chemistry. K−, C−, KC−, P−. Isousnic acid (major) (TLC).
Ecology and distribution. Lecanora mughicola is known from wood of conifers such as Larix and Pinus uncinata, from dry, non-rotten decorticated wood of trees, dead branches and worked timber. It occurs mostly in the subalpine zone between 1700 and 2400 m, but we have also seen a collection from c. 800 m. We have studied specimens from western, central as well as eastern Europe.
Notes. Lecanora mughicola is a conspicuous species because of its often blackish apothecia with a pale yellow margin. It differs from L. saligna by its smaller microconidia and by the absence of macroconidia, which are abundant in L. saligna. The apothecia are darker pigmented in L. mughicola and the ascospores are larger than in L. saligna. We have never found these two species growing together possibly because the distribution of L. saligna is restricted to lowland areas. In Printzen (Reference Printzen2001), conidia are described as 4–5·5 × 1–1·5 μm, which is somewhat different from our observations, and probably relates to mesoconidia.
Selected specimens examined. Norway: Oppland: Leirdalen, Ytterdalsseter, 8°11.3′E, 61°39.6′N, 1000 m, on scattered Betula trees near tree line, 1975, M. Brand 5719 (hb. Brand).—Great Britain: Scotland: V.C. 105, West Ross: Coulin Forest SSSI, high ground between Allt na Luib and An Leth-allt, on lignum of decorticated Pinus branch, c. 230 m, NG(18)993.555, 2004, A. M. & B. J. Coppins 21658 (E); Loch Maree, Eilean Subhainn, 18/91(−2).72, on exposed, dry lignum of Pinus branch, 1999, B. J. Coppins 18517 & V. J. Giavarini (E); Kinlochhewe, Beinn Eighe NNR, Coille na Glas-leitir, valley of Allt a'Chuirn, S side, 28/00.60, on decorticate branch of mature Pinus, 170 m, A. M. & B. J. Coppins 19821 (E). V.C. 96, Easterness: Rothiemurchus Forest, N to NE of Airgoid-meall, 28/96(−7).07, on exposed lignum of Pinus branches, 430–500 m, 1989, B. J. Coppins 13210 & T. Duke (E).—Switzerland: Valais: val d'Hérens, 0·7 km N of Arolla, 7°27.3′E, 46°1.7′N, 2120 m, decayed wood of dead log in open Larix-Pinus cembra wood on E slope, 1990, M. Brand 24875, 24876 (hb. Brand).—Austria: Steiermark: Schladminger Tauern, Stubneralm, Kleinsölktal, Kleinsölk-Obertal, steile felsige S-Abhänge des Säulecks, on Rhododendron ferrugineum, 1800 m, 1975, H. Mayrhofer & P. Remler (GZU); Alpes orientales, Gurktaler Alpen, Estiria, Frauenalm sur de Murau, bosque subalpino de Picea y Larix pasado refugio Murauer Hütte, on wood, 1700 m, 1981, A.Crespo & L. Sancho (MAF). Tirol: SE of Innsbruck, Patscherkofel, between Bosschede and Isshütte, 11°27′E, 47°12′N, 1900 m, dead branch of Pinus cembra, 1981, M. Brand 31809, (hb. Brand). Kärnten: Kreuzeckgruppe, Hochtristen, SW of Sattleggerhütte, 13°9.3′E, 46°46.2′N, 1700 m, on wood of roof of disused hay shed, 1994, M. Brand 31104 (hb. Brand); Gailtaler Alpen, Weissbriach, road at S-side of Schwarzenbach, 0·4 km from main road, 13°14.9′E, 46°40.9′N, 800 m, wood of railing of camping place, 1994, M. Brand 31423 (hb. Brand); Karnische Alpen, c. 2·3 km W of Nassfeldpass, W of Madritschen, 13°14.8′E, 46°33.5′N, isolated Larix, 1994, M. Brand 31335b (hb. Brand); SW of Hermagor, 1·2 km W of Nassfeld, path 403, Madritschen, Larix forest with Kreuzeckgruppe, 3 km NW of Greifenburg, 0·5 km S of Emberger Alm, SE slope with log-cabin in meadow, on wood of roof, 1725 m, 1994, P. van den Boom 15856 (hb. v.d. Boom).—Slovakia: Vysoke Tatry: 1·5 km S of Skalnate Pleso along Tratranske Magistrale, 20°13.6′E, 49°10.4′N, 1630 m, Pinus mugho in scrub on SE slope, decorticated wood, 2006, M. Brand 53596 (hb. Brand).—France: Savoie: Briancon, 0·9 km NNE of Col du Galibier, 6°25′E, 45°4′N, 2480 m, on exposed calcareous rock on gentle N-slope, 1984, M. Brand 11575 (hb. Brand). Vercors: Plateau Veymont, de la Maison forestière de la Cloche vers la Grande Cabane, Pâturages avec sapins disséminés, 1400–1500 m, viii 1986, E. Sérusiaux s.n. (LG). Pyrénées Atlantiques: E of Col de la Pierre St. Martin, 0°45.8′W, 42°57.9′N, 1800 m, stunted Pinus uncinatus at tree line, on wood of decorticated trunk, 1992, M. Brand, 28288 (hb. Brand); 14 km W of St. Lary-Soulan, Lac d'Aumar, crète d'Estibère, 0°9′E, 42°50′N, 2350 m, on old Pinus uncinatus, on N-slope, 1985, M. Brand 12814 (hb. Brand); 49 km SW of Pau, 4·6 km SE of St Engrâce, along trail to Pic d'Arlas, on decorticated wood of Pinus trunk, 1760 m, 1992, P. van den Boom 13157, 13129 (hb. v. d. Boom). Pyrenees Orientales: Pic Carlit, W of Lac de Bouillouses, 1°59.5′E, 42°33.8′N, 2100 m, on trunk of old, thick Pinus, 1983, M. Brand 32665 (hb. Brand).—Spain: Aragón: Huesca, Benasque, Ibón de Paderna (bajo) desde La Renclusa, on Pinus uncinata, 2240 m, 2004, J Etayo 21732 (hb. Etayo). Teruel: Javalambre, 6 km NNE of top, 0°59.32′W, 40°8.94′N, 1690 m, wood of dead Juniperus, 2005, M. Brand 52041 (hb. Brand). Navarra: Col de Arette-Pierre-St-Martin, 1550 m, on wood of Pinus, 1989, P. James, Rose, Etayo & Sérusiaux 10598 (LG, hb. v. d. Boom). Lérida: Pireneos, Aigües Tortes National Park, E-side of Val de Subenuix, 0°59.8′E, 42°34.5′N, 2150 m, on dead wood of Pinus cf. uncinata in open valley, 1983, M. Brand 32769 (hb. Brand). Soria: Puerto de Piqueras, 2°32.2′W, 42°3.2′N, 1700 m, wood of snow fence, 2003, M. Brand 50335 (hb. Brand). Madrid: Serra Guadarrama, Pt Cotos, on Pinus sylvestris, 1850 m, 1981, Equipo 1888 (MAF).—Italy: La Spezia: E of Passo di Cento Croci, 9°38.1′E, 44°24.7′N, 1100 m, wood of posts of fence on mountain ridge, 2001, M. Brand 42385 (hb. Brand). Lazio: pr. Frosinone, road Pontecorvo-Castrocielo, S of railway, 13°40′E, 41°30′N, 100 m, old Quercus along main road in flat fields, 2001, M. Brand 42621 (hb. Brand).—Greece: Kentriki Makedonia: W of Litochoro, Mt. Olympus, near refuge A, on decorticated trunk of mature Pinus, 2100 m, 1984, P. van den Boom 1992 (hb. v. d. Boom).
Lecanora quercicola Coppins & P. James
Lichenologist 11: 145–149 (1979); type: Anglia, V.C. 11, Hampshire: New Forest, Beaulieu, Stubbs Wood, Hawk Hill, 41/35.03, ad apricam faciem trunci Quercie margine sylvae, 2 xii 1975, P. W. James (BM—holotypus).
Thallus forming small patches of c. 1 cm, or effuse and up to 3 cm wide, uneven, slightly verrucose or granular-areolate to rimose-areolate, thin to relatively thick, up to c. 250 μm, sometimes endophloeodal; cortex poorly developed, algal cells scattered over the thallus, with granules at the upper surface; upper surface grey to yellowish grey or pale brownish grey, matt to slightly shiny. Photobiont chlorococcoid, cells 6–18 μm diam. Prothallus not observed.
Apothecia, broadly appressed, slightly constricted at base, 0·2–0·5 mm diam.; margin elevated, persistent, 0·06–0·09 mm wide, somewhat paler than the thallus, often uneven; disc flat or slightly convex, pale to greyish brown, or reddish brown, ± slightly pruinose.Thalline excipulum without cortex, or with pseudocortex, outer rim c. 20 μm wide, filled with granules, algae in the margin c. 9–12 μm diam. Parathecium often present, yellowish to yellow brown and c. 30–70 μm at outer rim, cells of inner part pseudoplectenchymatous. Epihymenium yellowish to pale brown, greyish brown to sometimes dark brown, filled with yellowish granules, N−. Hymenium hyaline, sometimes with orange-brown granules, 40–50 (−60) μm high. Hypothecium hyaline to yellowish, somewhat gelatinized in the lower part, 60–100 μm. Paraphyses simple to branched, sometimes constricted at the septa, mostly covered by a gelatinous sheath, (1·5–)2–2·5 μm diam., rarely slightly thickened towards the top and 3 μm wide, not pigmented, or wider, up to 3·5 μm and brown pigmented. Asci c.45 × 14 μm; tholus amyloid, without or with a small ocular chamber, with small axial body and clear, strongly amyloid cylinder around axial body. Ascospores ellipsoid, simple, 8·6–10 × 4·3–4·8 μm.
Macropycnidia always present, immersed, upper part not pigmented or brown, ostiole sometimes gaping and pigmented dark red-brown inside, up to 170 μm high × 100 μm; macroconidia weakly curved, non-septate, 8·5–9·3 × (2·5–)2·7–3·0(−3·2) μm. Micropycnidia sometimes present, only a few and hard to find, c. 90 μm wide, upper part pale brown; microconidia c. 10–11 × 1·1 μm.
Chemistry. K−, C−, KC−, P−. Isousnic acid (major), ±neousnic acid (minor) (TLC).
Ecology and distribution. Lecanora quercicola is known from old trunks of Quercus in open places in forests. In Bentheim (Germany) it has been found associated with Lecanora sublivescens, Caloplaca lucifuga, Pyrrhospora quernea, Calicium salicinum, Ochrolechia subviridis and Lepraria incana; in Brittany (France) with Opegrapha viridis, Rinodina roboris, Fuscopannaria cf. mediterranea and Frullania tamarisci. In southern Portugal and Spain (including Mallorca) well-developed material has been collected. Here the species has also been found growing on the bark or decorticated wood of Olea europaea trunks, accompanied by a few lichens, such as Lecanora aff. horiza, L. subsaligna, Lecidella achristotera and an unidentified sorediate crust.
Notes. Lecanora quercicola is distinguished from L. saligna by a less granular thallus and larger macroconidia (only 6·0–8·1 × 2·0–2·4 μm in L. saligna). Furthermore, the ecology of L. quercicola is characteristic, being found mainly on the sides of vertical to leaning very old Quercus or Olea trees in rich lichen communities. In Coppins & James (Reference Coppins and James1979) the measurements for macroconida are given as 8–12 × 3–4 μm, but they are only 8·8–9·5 × 2·6–2·9 μm in their drawing, which fits well with our measurements. Lecanora quercicola as well as L. saligna can both have slightly pruinose apothecia, but L. saligna has more irregular conidiogenous cells, 5·5–8 × 2–3 μm, whereas, according to the drawing in Coppins & James (Reference Coppins and James1979), the conidiogenous cells of L. quercicola are more cylindrical and 8·5–13 × 2·4–3·3 μm. A further type of conidium (8–13 × 0·4 μm) is also recorded by Coppins & James (Reference Coppins and James1979), which was not observed by us.
Selected specimens examined. Germany: Niedersachsen: Bentheim, Bad, 7°10′E, 52°19′N, on old mature Quercus at S-side of building in park, 1976, M. Brand 6032 (hb. Brand).—France: Morbihan: 27 km ENE of Lorient, Forêt de Camors, 1 km E of Lambel, 3°1.0′W, 47°50.0′N, 100 m, one old Quercus (circumf. 3 m), on road cross in rather young woodland, 1988, M. Brand 18457 (hb. Brand); 27 km ENE of Lorient, Forét de Camors, 1 km E of Lambel, on mature Quercus, 110 m, 1988, M. Brand 18457 (hb. Brand).—Spain: Mallorca: N of Palma, along road C-710, 3 km W of Deia, small orchard with Olea trees, on Olea, 200 m, 1997, P. van den Boom 18422 (hb. v. d. Boom); W of Palma, N of Andratx, 5 km N of Coll de Sa Gremola, W side of road C-710, Olea orchard, on Olea, 300 m, 1997, P. van den Boom 18298 (hb. v. d. Boom). Galicia: Pontevedra, 4km SE of Lalín, 8°5.23′W, 42°38.01′N, 650 m, on old Quercus in open forest, 2003, M. Brand 49275 (hb. Brand).—Portugal: Minho: Serra Do Geres, W of Cabril, along road to Pincães, scattered mature Quercus robur, on trunk of Quercus, 450 m, 1999, P. van den Boom 23197 (hb. v. d. Boom). Estremadura : Serra dos Candeeiros, S of Leiria, SE of Porto de Mós, Zambujal de Alcaria near crossing to Alvados, Olea orchard, on Olea, 1995, P. van den Boom 17194, 17215 (hb. v. d.Boom). Alentejo: Éuvora SE of Alvito, on Olea, P. van denBoom 17035 (hb v. d. Boom); Serra de S. Mamede Natural Park, E of Portalegre, S of Pico de Serra de Mamede, near Ribeira de Arronches, mature Castanea trees in garden, on trunk of Castanea sativa, 680 m, 2003, P. & B. van den Boom 31436 (hb. v. d. Boom).
Lecanora saligna (Schrad.) Zahlbr
Cat. Lich. Univ. 5: 536 (1928).—Lichen salignus Schrad. Spic. Fl. Germ. 1: 84. (1794). (?LE—holotypus, not seen) [material requested but not received].
Lecanoropsis saligna (Schrad.) M. Choisy, Bull. Mens. Soc. Linn. Lyon 18: 143 (1949).
(Fig. 8D)
Thallus often effuse, in patches to 8 cm, uneven, slightly to strongly warted, granular to warted-areolate, warts sometimes aggregated, if on wood sometimes partly endoxylic, up to c. 300 μm thick, without cortex, with granules mainly on the upper surface, algal layer and medulla not differentiated; upper surface grey, whitish grey, yellowish grey, to pale brownish grey, smooth. Photo-biont chlorococcoid, cells 7–20 μm diam. Prothallus not observed.
Apothecia broadly appressed, scattered to crowded and deformed, 0·4–0·7(−1·1) mm diam.; margin persistent, 0·04–0·1 mm wide, concolorous with the thallus or paler, often uneven, persistent, sometimes flexuose; disc flat to very weakly convex, pale to deep red-brown, epruinose to sometimes slightly pale yellowish pruinose. Thalline excipulum abundantly filled with granules, cortex clearly developed, ± chondroid, algae abundantly present throughout the thalline excipulum, often reaching the outer rim. Parathecium often present, rather narrow, up to 20 μm wide at outer rim and pigmented medium brown. Epihymenium yellowish brown, reddish brown or medium brown, with yellowish granules, N−. Hymenium hyaline or pale yellowish with pale orange-brown granules, 40–60 μm high. Hypothecium hyaline or rarely weakly yellowish pigmented, to 100 μm high. Paraphyses simple, branched or anastomosing, 1·5–2(−2·5) μm wide, mostly covered by a gelatinous sheath, abundantly septate, often slightly constricted at septa to more rarely nearly moniliform, apically only scarcely widened up to 3·5 μm, rarely brownish pigmented in upper cell. Ascospores simple, ellipsoid, c. 8–10 × 4·1–4·8 μm.
Macropycnidia always abundant, immersed, upper part hyaline, reddish brown, brown or bluish black, 90–150 μm high × 60–110 μm; macroconidia always weakly curved, not septate, 6·0–8·1 × 2·0–2·4 μm. Micropycnidia scarce and hard to find, moderately brown in upper part, hyaline below, c. 50 × 40 μm; microconidia always slightly curved, 8·4–10·5 × 0·9–1·2 μm. Mesopycnidia rare and hard to find; mesoconidia bacilliform, 3–5 × 1·1–1·5 μm.
Chemistry. K−, C−, KC−, P−. Isousnic acid (major), neousnic acid (found in c. 50% of the specimens), ± atranorin (minor) and ±zeorin (minor), the latter two have been found only very rarely (TLC & HPLC).
Ecology and distribution. Lecanora saligna is a very common species in lowland areas throughout Europe. It grows on not too strongly acidic, dead wood (e.g. impregnated by dust or shifting sand), on fence posts, fallen branches and stumps. We have found it also on bark of trees, such as Betula, Malus, Salix, Populus, Quercus, Robinia, often in Xanthorion communities with Amandinea punctata, Lecanora expallens, Micarea denigrata, Placynthiella dasaea, Strangospora pinicola and Xanthoria parietina. Buellia schaereri has rarely been found as an accompanying species. Although L. saligna is essentially a lowland species, we have studied specimens from altitudes of 1150 m in France and 1400–1600 m in Russia.
Notes. This is the most common of the species in western Europe treated here. It is characterized by brownish apothecia with a yellowish, persistent margin, and by the typical, weakly curved macroconidia measuring 6·0–8·1 × 2·0–2·4 μm, found in the usually abundant reddish to dark-brown macropycnidia. While Hawksworth & Dalby (Reference Hawksworth, Dalby, Purvis, Coppins, Hawksworth, James and Moore1992) record that this species grows on “wood, worked timber and more rarely on dead bark”, the records in Printzen (Reference Printzen2001) are mainly from wood and the bark of conifers. However, we have collected it from the less acidic bark of Populus, Salix and Sambucus so that its ecological amplitude is even wider. Populations on living bark have been found mostly on bases of tree trunks, only more rarely on higher parts of the trunks. The species can appear in large populations up to several dm2.
Most characters of this species are rather constant, for example, the macroconidia, which we measured from many specimens were 6·0–8·1 × 2·0–2·4 μm. However, all more or less recent publications report variable measurements: 6–11 × 1–3 μm (Clauzade & Roux Reference Clauzade and Roux1985), 7–8 × 2–2·5 μm (Diederich Reference Diederich1989), 7–8(−9) × 1·5–2·5 (−3) μm (Hawksworth & Dalby Reference Hawksworth, Dalby, Purvis, Coppins, Hawksworth, James and Moore1992), 4–7·5 × 1·0–1·5 μm (Printzen Reference Printzen2001), and 10–12 × 0·7–1·2 or 6–8(−11) × 1·5–3 μm (Wirth Reference Wirth1995). Some of these measurements refer to microconidia which are scarce, or they include two kinds of conidia. In well-developed specimens macroconidia are always present; only in a few (scanty) specimens, mostly with a reduced thallus are pycnidia lacking. The conidia in the specimen Sérusiaux 6495, are 10–12 × 0·5 μm, as annotated by B. J. Coppins, but we were unable to find such conidia.
The lichenicolous fungus Lichenodiplis lecanorae growing on apothecia and thalli was present in many collections. Infections by Pseudodiplodia lichenis Vouaux, Unguiculariopsis lesdainii (Vouaux Etayo & Diederich) or Taeniolella delicata M.S. Christ. & D. Hawksw. are rare.
Selected specimens examined. Great Britain: England: V.C. 14, East Sussex: S of London, Lewes, on timber, s.d. Crombie s.n. (B). V.C. 36, Hereford: Kentchurch Deerpark, in parkland, on tree bole of Populus, 100 m, 1974, Sipman 6599 (B).—The Netherlands: Friesland: Vlieland, Kroonspolders, 4°°57.9′E, 53°15.3′N, piece of wood near the beach, 1973, M. Brand 3015 (hb. Brand); Ameland, Hollum, Lange Duinen, small woodland with Sambucus and Hippophae, on Sambucus, 1995, P. van den Boom 17518 (hb. v. d. Boom). Drenthe: W of Emmen, Sleen, brink, 6°48.2′E, 52°46.5′N, on mature Quercus tree, 1976, M. Brand 6318 (hb. Brand); ENE of Mantinge, N of Balingerbosch, meadow with Alnus trees, on Alnus, 1999, P. van den Boom 23645 (hb. v. d. Boom). Noord-Holland: Bergen aan zee (N), 4°38′E, 52°40′N, on dead fallen trunk of tree in dune area, 1975, M. Brand 5221 (hb. Brand). Flevoland: Noordoostpolder, Urk, along ‘Staart’ reserve, 5°37′E, 52°40′N, Salix alba along road, 1979, M. Brand 7904 (hb. Brand). Overijssel: Twente, De Lutte, Bloemenbeek, 7°2′E, 52°19.1′N, wood of fence post along meadow, 1976, M. Brand 6033 (hb. Brand). Zuid-Holland: Voorne, Oostvoorne, v. Itersonbos, 4°4.0′E, 51°54.6′N, on Populus in dune area, open woodland, 1971, M. Brand 1738 (hb. Brand). Gelderland: Herwijnen, decoy at Detsteeg, 5°6.7′E, 51°51.2′N, dead wood of Salix, 1975, M. Brand 5201 (hb. Brand); Winterswijk (NW), farm ‘Kleuverskamp’, S of Korenburg, 6°40.2′E, 51°58.7′N, Quercus along unpaved road, 1992, M. Brand 29097 (hb. Brand). Zeeland: NW of Sas van Gent, 3°48.7′E, 51°14.0′N; on base of pollard-willow, along ditch in meadow, 1972, M. Brand 2373 (hb. Brand). Noord Brabant: Biesbos, W van Noorderplaat, Biezengors beoosten 130, S bank of gully, 4°47.4′E, 51°44.8′N, Salix dasyclados in neglected, low willow-ground, 1990, M. Brand 22600 (hb. Brand); SSW of Leende, Groote Heide, S of unpaved road, Calluna heathland, on solitary Sambucus shrub, 2005, P. van den Boom 35694 (hb. v. d. Boom); ibid., on large Pinus stump, 2005, P. van den Boom 35684 (hb. v. d. Boom). Limburg: Nieuw Bergen, E of Meeuwenven, at picnic place, on wood of plaything, 6°4.3′E, 51°36.4′N, 1992, M. Brand 29060 (hb. Brand); SW of Reuver, Beesel, ‘Het Broek’, damp Populus woodland, on Populus, 1992, M. Brand 29027 (hb. Brand).—Belgium: Antwerpen: NW of Kalmthout, Kalmthoutse Heide, Calluna heathland, on rotting wood, 2000, P. van den Boom 24012 (hb. v. d. Boom). Limburg: N of Lommel, Blekerheide, former heathland with Pinus forest, along maize field, on Salix, 1999, P. van den Boom 22706 (hb. v. d. Boom). Liège: Angleur, Parc de ‘La Vieille Montagne’ (Umicore), rive gauche de l'Ourthe, on Aesculus, iv 2005, E. Sérusiaux s.n. (LG). Luxembourg: Straimont, Martilly, bord du chemin de Gribomont, à la sortie du village, sur tronc de Fraxinus, 1965, J. Lambinon, 65/B/929 (LG); Rochehaut, Route descendant vers la Semois par le Moulin du Bochet, on Acer, 1984, E. Sérusiaux 6908 (LG); 2 km WSW of Durbuy, 1·5 km SSE of Septon, W above Fond du Bois, shaded calcareous outcrops in Quercus fagus forest, on fence post, 200 m, 1989, P. van den Boom 8214 (hb. v. d. Boom); 2·5 km N of Virton, 0·2 km N of Croix St. Jacques, 5°31.9′E, 49°35.4′N, 290 m, on old Quercus at crossroads in wood, 1991, M. Brand 25447 (hb. v.d. Brand). Lorrain, SE of Florenville, S of Pin, Forét d'Orval, path along stream ‘Frere Simon’, mixed forest on N exposed slope (L6.58), on wood of fence post, 250 m, 1998, P. van den Boom 20053 (hb. v. d. Boom).—Germany: Hamburg: Am Elbufer bei Kittenbergen, on Sambucus, 25 ix 1932, C. Erichsen (GZU). Brandenburg: SE of Berlin, 8 km S of Storkow, E of Bugk, W side of small lake, Pinus forest, on stump, 40 m, 2002, P. van den Boom 21065, 21203 & H. Sipman (hb. v. d. Boom). Nordrhein-Westfalen, near Münster, Welbergen, on wood of an old barrier, viii 1862, Lahm? s.n. (B). Rheinland-Pfalz: Hamm, bei dem Punkt ‘Schöne Aussicht, on Malus, 18 ii 1984, V. John s.n. (hb. John); Saarland, Entre Lautenbach et Dörrenbach, on Malus, 1984, P. Diederich 5776 & V. John (hb. Diederich).—Luxembourg: Gutland: route Berdorf vers le Mullerthal au Sud, Pretigstull, on Quercus, 1984, P. Diederich & E. Sérusiaux 6495 (LG). Ardenne: SWW of Boulaide (K7.47), on wood in a forest, 1987, P. Diederich 8494 (hb. Diederich); 0·5 km SSE of Hoscheid, 6°5.0′E, 49°56.5′N, 465 m, old Fraxinus at side of former high road on plateau with arable land, 1989, M. Brand 20203 (hb. Brand). Lorrain: NNW of Contern, Weierboesch (M8.27), on Quercus, 1987, P. Diederich 8938 (hb. Diederich); E of Emerange, berge de la Gander (M8.58), on Salix, 1987, P. Diederich 8614 (hb. Diederich); SW of Ettelbruck (K8.54), on wood, 1987, P. Diederich 7811 (hb. Diederich); Berschbach (L8.35), on Salix, 1893, Feltgen 15 (LUX); ibid., SW of Septfontaines, W of Sauerwisen (L8.42), on Pyrus, 1979, P. Diederich 1957 (hb. Diederich).—Austria: Salzburg: N of Radstadt, road from Ebe to Filzmoos, near Ploimühle, Picea forest with Alnus incana, on Picea, 860 m, 2002, P. van den Boom 28420 (hb. v. d. Boom).—Finland: N Karelia: Koli, 0·5 km SE of Kotaniemi, 29°52.5′E, 63°2.8′N, 100 m, wood of fence along meadow, 1980, M. Brand 9708 (hb. Brand).—Sweden: Götland: Öland, Mörbylanga Kirka, 16°23.5′E, 56°30.8′N, old Fraxinus in churchyard, 1989, M. Brand 21333 (hb. Brand).—Denmark: Jutland: Rømø, Toftum, near museum, 8°33.4′E, 55°9.9′N, on old Malus, 1977, M. Brand 7079 (hb. Brand).—Hungary: Bacs Kiskun: 35km SE of Budapest, Csévharaszt, 19°2′E, 47°56′N, 100 m, dunes of lime rich sand, on rabbit pellets, 1981, M. Brand 10541 (hb. Brand).—France: Vosges: 10 km WSW of Munster, 0·1 km W of Kastelbergwasen, 7°0.4′E, 48°7′N, 1150 m, on free standing old Fagus in open meadow, 1984, M. Brand 11776 (hb. Brand); SE of Kantelberg, S of Honeck, on old freestanding Fagus trunk, 1200 m, 1984, H. Sipman 17586. Jura: S of Besançon, SSE of Champagnole, Cornu, 800 m, on base of Picea and on fence post, 1986, P. van den Boom 3833 (hb. v. d. Boom).—Austria: Steiermark: Seetaler Alpen, Neumarker Sattel, 3 km NW von Neumarkt, Strasse von Hoferdorf, Richting Moar am Berg, 940 m, Pfosten einer Holzbrucke, 1989, Obermayer 2609 (GZU).—Italy: Toscana: 6 km WNW of Cortona, NW of S. Caterina, 11°54′E, 43°17′N, 300 m, Cupressus along road through fields, flat land, 2001, M. Brand 43725 (hb. Brand). Lazio : Frosinone, c. 1 km S of Strangolagalli, 13°29′E, 41°35′N, 100 m, wood of poles supporting vines, blue by Cu-compounds, 2001, M. Brand 42617 (hb. Brand).—Russia: Caucasus distr.: Ochamchira, vicinitas latior pagi mons Bardunakravar, 1400–1600 m, 14 vi 1981, V. Vašák s.n. (B).—Spain: Galicia: 34 km ESE of Ourense, E of Paredes, 7°27.0′W, 42°16.1′N, 1070 m, on rotting wood of fence post in meadow, 2003, M. Brand 49308 (hb. Brand).—Portugal: Tras-os-Montes: N of Vila Real de Santo Antonio, Reserva Natural do Sapalde Castro Marim, W of Leziria, unpaved road among orchard with Olea europaea and Ceratonia siliqua, 110 m, on wood of trunk of mature Olea, 2006, P. & B. van den Boom 36086 (hb. v. d. Boom).
Exsiccata examined: Arnold, Lich. exs. 472 [as L. piniperda] (B); Arnold, Lich. Mon. exs. 159 [as L. piniperda] (B); Hillman, Lich. exs. 2568 (B); Plant. Graec. exs. 149 [as L. subintricata](B); Plant. Graec. exs. 404 (B); Poelt & Hafellner, Lich. exs. 317 (B); Schade, Stolle & Riehmer, Lich. exs. 547 [as L. metaboloides](B); Tobolewski, Lich Polonica Fasc. IX, X (B).
Lecanora sarcopidoides (A. Massal.)A. L. Sm
Bihang till Kgl. Svensk Vetensk. Akad. Handl. 18: 47 (1892).—Biatora sarcopidoides A. Massal. Ricerch. Auton. Lich. (1852); type: Italy, prov. Veronensis, et Vicentina, ad nudos, et marcescentes castanearum truncos, ?1851, leg. A. Massal. Anzi, Lichenes rariores Veneti exs. 61 (M—lectotype, selected here).
Lecanora sarcopidoides (A. Massal.) Hedl. var. hypnophaga Poelt, Mitteil. der Bot. Staatssammlung München 2(17–18): 386–399 (1957); type: Germany, Bayern, Isarwinkelgebirge, Gergwald zw. Benediktbeuren und der Hausstattalpe, on Hypnum cupressiforme filiforme, on Picea, c. 900 m, ?1956, J. Poelt s.n. (M—holotype!).
(Figs 8B & 10)

Fig. 10. Lecanora sarcopidoides, habitus (M. Brand 6224). Scale: 0·5 mm.
Thallus effuse, to 4 cm wide, endosubstratal to episubstratal, mostly thin, 0·05–0·3 mm high; areolate, areoles irregularly warted; wartsc. 0·5 mm wide, sometimes aggregated; cortex not well developed, mainly at the lower part of the apothecium, below the excipulum; epinecral layer not developed, algal layer to 80 μm, granules present in all parts of the thallus, more often in the upper layer of up to 60 μm high; upper surface grey, greyish brown, pale yellowish brown, matt.Photobiont chlorococcoid, cells 8–17 μm diam. Prothallus not observed.
Apothecia broadly appressed to sessile, slightly constricted at the base, 0·25–0·5 (−0·7) mm diam.; margin thin, 0·05–0·07 mm wide laterally, initially clearly raised, becoming level with the disc, mostly persistent, concolorous with the thallus to concolorous with the disc; disc initially plane to slightly convex becoming strongly convex to subglobose; pale yellowish brown to sometimes dark greyish brown to bluish grey and becoming black, often slightly to densely whitish pruinose, in older apothecia often epruinose. Thalline excipulum corticated, chondroid, up to 70–130 μm high, upper part diffuse brown, sometimes reduced, algal layer reaching the upper surface, granules in anticlinal rows in apothecial cortex and partly in the algal layer. Parathecium reddish brown or sometimes pigmented bluish green in outer part and up to 20 μm wide, pseudo-parenchymatous. Epihymenium yellowish brown, olive brown to more rarely dark brown, N+ reddish violet, with a mix of smaller and larger (1–5 μm) hyaline granules reaching the parathecium and the outer rim of the excipular cortex. Hymenium upper part ± diffuse reddish brown, sometimes spotted and changing to blue grey, 40–45 μm high. Hypothecium hyaline, to 100 μm high. Paraphyses simple to branched, sometimes irregularly septate, rarely constricted at septa, 1–2 μm wide, not or rarely widened above, apices hyaline, rarely olive-brown pigmented, ± conglutinated. Asci clavate, Lecanora-type, 20–27 × 7–12 μm. Ascospores ellipsoid, simple, rarely 1-septate, 8–10 × 3·3–3·7 μm.
Micropycnidia not often found, mostly immersed, ostiole brown or greenish black, 70 μm high × 70–80 μm; microconidia bacilliform, 4·0–5·7 × 1·0–1·2 μm. Mesopycnidia often present, immersed, ostiole pale brown, c. 110 × 80 μm; mesoconidia ellipsoid, 2·7– 3·7(−4·1) × (1·1–)1·3–1·7 μm. Leptopycnidia very rare, immersed, ostiole greenish black, c. 150 × 110 μm; leptoconidia slightly to strongly curved,c. 19 × 0·7 μm.
Chemistry. Pseudoplacodiolic acid (major),placodiolic acid (trace), usnic acid (trace); K−, C−, KC− yellow, P± pale yellow (TLC & HPLC).
Ecology and distribution. Lecanora sarcopidoides is known from lowland and upland areas up to 2000 m, on dead, rotting wood or the bark of conifers (Abies, Picea, Pinus), and also on Quercus in the montane zone (500–1500 m), often accompanied by calicioid lichens such as Calicium glaucinum, C. salicinum and Chaenotheca brunneola. Lecanora conizaeoides and L. varia have also been found growing nearby.
Notes. Although L. sarcopidoides is a very characteristic species, it is one of the most easily confused species in the L. saligna group. However, the combination of pseudoplacodiolic acid, placodiolic acid and usnic acid is unique in the group and distinguishes it from all known related species. The first compound is not known from any of the other species treated here. Lecidea leprarioides Tønsb. has pseudoplacodiolic acid, a sorediate thallus, black lecideine apothecia and unknown pycnidia (Tønsberg Reference Tønsberg1992), but it is not related to the Lecanora saligna group. Lecidea turgidula Fr. also has placodiolic acid, but differs in having black lecideine apothecia, paraphyses with dark brownish apices up to 5 μm wide, and an epithecium with a bluish green pigment (several specimens in the herbaria of the authors have been studied). Young and well-developed specimens of L. sarcopidoides have pruinose apothecia, but in (over)mature apothecia pruina is lacking and then the apothecia are darker. Well-developed specimens can have a thickish, areolate thallus, for example Brand 6224. This last specimen contains three types of pycnidia of which macropycnidia are the most common and the associated species are L. conizaeoides, L. varia and Chaenotheca brunneola.
The closest relative is L. subintricata, which has microconidia of somewhat similar size, however it has smaller ascospores, a more yellowish thallus and apothecia and the thallus is thinner, less warted and with smaller areoles. In addition, the chemistry is different, with usnic acid being the major compound. Conidial sizes of 6–11 × 1–3 μm (Clauzade & Roux Reference Clauzade and Roux1985), 3–7 × 1–1·5 μm (Wirth Reference Wirth1995) and 3–7 × 1–1·5 μm (Hawks-worth & Dalby Reference Hawksworth, Dalby, Purvis, Coppins, Hawksworth, James and Moore1992) are reported in the literature so these measurements apparently refer to microconidia. Lecanora sarcopidoides is easily confused with L. pseudosarcopidoides, especially specimens with pruinose apothecia, and in these cases conidial and chemical characters are necessary for identification. Many collections of L. sarcopidoides have been found in herbaria under the name Lecanora piniperda.
Lecanora sarcopidoides seems to be a rather rare species. According to Hawksworth & Dalby (Reference Hawksworth, Dalby, Purvis, Coppins, Hawksworth, James and Moore1992) it is a poorly understood species in the British Ises. We have studied the type of L. sarcopidoides var. hypnophaga, described by Poelt (Reference Poelt1957), and, although it is the only specimen we have seen growing on mosses, we did not find any relevant differences in either morphology, or in chemistry, so we regard this variety as belonging to L. sarcopidoides s.str.
Additional specimens examined. Sweden: Värmland, 12 km NW of Sunne, Gettjärn, 12°56.9′E59°53.5′N, 150 m, 2003, M. Brand 47210 (hb. Brand).—Great Britain: Scotland, V.C. 89, East Perthshire: Enochdhu, Kindrogan Field Centre, 37/05.62, on lignum of standing decorticated conifer trunk, c. 250 m, 2001, B. J. Coppins 20208 (E).—Ireland: V.C. H33, Fermanagh: Crom, Crom Castle Deer Park, 23/36.24 on exposed decorticated stump, 60 m, 1993, A. M. & B. J. Coppins 15898 & A. M. O'Dare (E).—The Netherlands: Noord Brabant: between Nieuwkuik and Helvoirt, on Pinus, ?1904, J. H. Wakker s.n. (L).—Belgium: Liège: Hautes Fagnes, Sourbrodt, Nesselo, edge of bog, wood of dead tree, 6°8.6′E, 50°29.8′N, 565 m, 1976, M. Brand 6224 (hb. Brand). Luxembourg : W of Bouillon, W of Corbion, Le Tabourin, on wood of fence post along Picea woodland, 360 m, 1999, P. van den Boom 21870 (hb. v. d. Boom); 3·5 km SE of Herbeumont, Fotêt d'Herbeumont, SE of Pavillon de chasse, along Semois, at E steep acidic outcrop (L6.26), on dead standing Quercus trunk, 300 m, 2006, P. & B. van den Boom 37126 (hb. v. d. Boom).—Germany: Schleswig-Holstein: Eichenpfähle im Prökelmoor bei Wellingsbüttel, 20 iii 1904, F. Erichsen s.n. (B); Flensburg, on stump of Quercus, 10 xi 1929, W. Saxen s.n. (B).—Austria: Steiermark: Salzburger Alpen, Ennstal, 5 km NW of Irdning, valley with limestone outcrops, on wood of Picea trunk, 14°2′E, 47°31′N, 700 m, 1981, M. Brand 31775 (hb. Brand).—France: Pyrénées atlantiques: St. Engrâce, 2·5 km NNE of Senta, S slope of Pic d'Issarbe, 0°48.0′W, 43°.4′N, 1500 m, wood of standing, dead Abies in Abies-Fagus wood on SW slope, 1992, M. Brand 28699 (hb. Brand).—Czech Republic: Western Bohemia: SSE of Klatovy, Národni park, E of Srni, NW of Turnerova Chata, E side of Vydra, well-lit relict pine forest, SW exposed scree, with stumps and Betula trees at edge of forest, on vertical side of stump, 750 m, 2003, P. van den Boom 31701 & Z. Palice (hb. v. d. Boom).—Italy: Trentino-Alto Adige : prov. Bolzano, 54 km WSW of Merano, NP Stilfser Joch, SW of Trafoi, S slope with Pinus trees and stumps, on Pinus stump, 1960 m, 1990, P. van den Boom 10793 (hb. v. d. Boom).
Exsiccata examined: Anzi, Lich. merid. rar. 175 (M) [as L. varia var. saepincola]; Arnold, Lich. exs. 708 (B, GZU, H-Nyl, L) [as L. metaboloides]; Hepp, Flecht. Europ.69 (L) [as L. piniperda]; H. Olivier, Lich. exs. 230 (LUX) [as L. piniperda ]; Rehm, Lich. exs. 34 (L) [as L. piniperda]; Zwack, Lich. exs. 116 (L) [as L. varia var. saepincola].
Lecanora subintricata (Nyl.) Th. Fr
Lichenogr. Scand. 1: 265 (1871).—Lecanora varia var. subintricata Nyl., Flora 51: 478 (1868); type: Russia, Kola, super corticem Betulae, 1863, N. I. Fellman [Fellman, Lichenes artici 133] (H-Nyl. 26661—holotype).
Lecanoropsis subintricata (Nyl.) Choisy, Bull. Mens. Soc. Linn. Lyon 18: 143 (1949).
Thallus mostly endosubstratal, sometimes episubstratal, up to several cm wide, of dispersed, c. 0·15 mm wide warts, without cortex and epinecral layer, with granules in the upper part of the thallus, algae dispersed over the thallus, reaching the upper side; upper surface yellow grey, yellow green to pale yellowish brown, matt or sometimes shiny. Photobiont chlorococcoid, cells to 25 μm wide. Prothallus not observed.
Apothecia appressed to sessile, juvenile apothecia erumpent, at first ± gyalectoid, 0·4–0·6 mm diam.; margin initially prominent, concolorous with the thallus, turning inward and irregular, becoming ± excluded, more or less biatorine and concolorous with the disc, 0·03–0·06 mm wide; disc plane to convex, yellowish beige or grey, moderately brown, pale yellowish brown, to olive-brown, sometimes slightly white or bluish pruinose. Thalline excipulum corticate, cortex 20–30 μm thick, regular in thickness, chondroid, with anticlinal hyphae. Parathecium sometimes visible, small, 10–30 μm and brownish at the outer rim, filled with granules especially in the outer part. Epihymenium pale yellowish brown, medium to dark brown, sometimens olive pigmented, with many fine and more stout, yellowish granules, N+ reddish violet. Hymenium hyaline, with orange-brown granules, 30–50 μm. Hypothecium hyaline to rarely slightly yellowish, to 120 μm high. Paraphyses simple to branched and anastomosing, top hyaline to rarely brown and bluish grey, up to c. 3 μm wide. Ascospores ellipsoid, simple, 6·7–8·4 (−10·2) × 3·1–3·6 μm.
Micropycnidia rarely present, immersed, top free and brown to greenish blue, 40–90 × 40–60 μm; microconidia short-bacilliform, 4·8–6·2 × 1·0–1·3 μm. Mesopycnidia often present, black at least in upper half, sessile, 120–130 × 100–110 μm; mesoconidia ellipsoid to short cylindrical 2·8–3·4 × 1·4–1·6 μm.
Chemistry. K± yellow, C−, KC+ yellow Pd·. Usnic acid (major); ± brialmontin 1 (minor) (TLC & HPLC).
Ecology and distribution. Lecanora subintricata occurs mainly on bark or decorticated wood of conifers (Pinus cembra, P. nigra, Larix) mainly in the montane and subalpine zones (1000–2000 m) in Europe. Accompanying species are often Hypogymnia physodes, Lecanora varia, Parmelia sulcata, Parmeliopsis ambigua and Pseudevernia furfuracea, and sometimes Hypocenomyce scalaris, Trapeliopsis flexuosa and Xylographa vitiligo.
Notes. Lecanora subintricata is distinguished from all the other species mentioned in this study by the presence of usnic acid as a major compound. However, according to Printzen (Reference Printzen2001) Lecanora albellula found in North America can have usnic acid as a major component. Furthermore L. subintricata differs from other species with usnic acid by its much smaller ascospores (except L. coppinsii and the western American species L. latens), smaller apothecia and shorter microconidia or mesoconidia.
The measurements of conidia reported in the literature: 4–7 × 1 μm (Clauzade & Roux Reference Clauzade and Roux1985; Wirth Reference Wirth1995); 3–7 × 1–1·5 μm (Hawksworth & Dalby Reference Hawksworth, Dalby, Purvis, Coppins, Hawksworth, James and Moore1992) and 5–8 × 1 μm (Hinteregger Reference Hinteregger1994) most probably refer to microconidia or a mixture of micro- and mesoconidia. Although the specimens we studied have ascospores <8·5 μm (very rarely 8·5–9·5 μm) long, Hinteregger (Reference Hinteregger1994) recorded them as 8–11·5(−13) μm long and sometimes 1-septate. However, we have never found septate ascospores in L. subintricata and cannot explain these measurements. In one British specimen, Ellis L235, both micro-conidia and mesoconidia were found and their measurements agree with those in our description. However the specimen is poorly developed and we could not detect any lichen substance by TLC. Isousnic acid has been reported from most American specimens (Printzen Reference Printzen2001); this compound is probably identical with the compound formerly named by us as ‘subintricata unknown’ and subsequently identified by J. A. Elix as brialmontin 1.
Selected specimens examined. Norway: Oppland: Rondane, 3 km E of Doralseter, open fjell, on dead wood of Betula, 9°51′E, 62°0.6′N, 1000 m, 2003, M. Brand 47408 (hb. Brand); Sel, Horgesetrene, W above Rosten, towards Mt. Horgenosi, 9°22′E, 61°52′N, 1000 m, Betula nana-Cladina-heath, just above tree limit, on isolated Betula tree trunk, 1999, H. Sipman 44165 (B). Buskerud: Hemsedal, Mörkedalen, W slope of Slettind, on twigs of Salix glauca, 8°9.6′E, 60°58.2′N, 1100 m, 2003, M. Brand 48205 (hb. Brand).—Sweden: Torne Lappmark: Kiruna, near Jieprenjakkstugan hut, on N shore of Lake Torneträsk, subalpine Betula tortuosa forest, on trunk of Betula, 18°52′E, 68°27′N, 350 m, 1986, H. Sipman 21385 (B).—Finland: Lapponia enontekiensis: Pallastunturi, S of Pallaskero, 24°6.4′E, 68°1.6′N, 400 m, Salix capraea on wet place in Pinus forest, 1980, M. Brand 9873, 9869 (hb. Brand); 19 km NE of Muonio, area Matkailukeskus, open heathland with scattered Betula and Picea, on wood of sauna house, 470 m, 1988, P. van den Boom 7394 (hb. v. d. Boom). Lapponia inarensis: parish Utsjoki, valley of Kevojoki river, 1–2 km before outlet into Lake Kevojärvi, 100 m, light mossy Betula forest in valley on E slope, on dead wood of fallen tree, 1988, H. Sipman 23550, Tan & E. Reiniko (B).—Great Britain: Scotland: NO 483788, Aspen stand, in pasture close to north banks of the River Esk, nr East Migvie, on Populus tremula, 200 m, 2004, RBGE Aspen Epiphyte Project, C. J. Ellis L235 (E).—Switzerland: Valais: Val d'Hérens, 0·5 km SW of Evolène, 7°29.4′E, 46°6.6′N, 1400 m, Larix at edge of forest, M. Brand 24365 (hb. Brand); 0·7 km N of Arolla, 7°27.3′E, 46°1.7′N, 2120 m, decayed wood of dead log in open Larix–Pinus cembra wood on E slope, 1990, M. Brand 24876 (hb. Brand). Valais: 7 km S of Kandersteg, S edge of Sagiwald, on decorticated wood of Pinus mugo, 2000 m, 1990, M. Brand 24031 (hb. Brand). Graubünden: Engadin, Val Susauna, 9°59.5′E, 46°38.9′N, 1690 m, on rotten wood in meadow near edge of forest; 1977, M. Brand 6685b (hb. Brand).—Austria: Tirol: Innsbruck, Patscherkofel, between Boschede and Isshütte, Pinus cembra forest on N slope, 11°27′E, 47°12′N, on dead branch, 1900 m, 1981, M Brand 31808 (hb. Brand). Styria: S of Trieben, W of Hohentauern, trail in Picea forest, on stump, 1350 m, 2002, P. & B. van den Boom 28616, 28627 (hb. v. d. Boom). Carinthia: Gailtaler Alpen, 12 km NW of Hermagor, area of Weissbriach, Alnus woodland, on wood of bird observation post, 820 m, 1994, P. van den Boom 15728 (hb. v. d. Boom).—Italy: Lombardia: Como, 7·5 km W of Gravedona, Val S. Iorio, 9°12′E, 46°9′N, 1500 m, Larix in Fagus-Larix forest, just under timberline, 1993, M. Brand 30263 (hb. Brand). Trentino-Alto Adige: Bolzano, 54 km WSW of Merano, NP Stilfser Joch, road Prad-Stilfserjoch, SW of Trafoi, SE slope, with Picea wood and outcrops, on Picea, 1850 m, 1990, P. van den Boom 10836 (hb. v. d. Boom).—France: Hautes Alpes: c. 27 km SE of Briancon, near Fontgillarde, 6°52′E, 44°43′N, 1970 m, on Larix in open wood on N slope, 1984, M. Brand 11678b (hb. Brand). Alpes Maritimes, 1·6 km ESE of Col de Cayolle, Pinus and Larix in open wood near tree line, 6°45.9′W, 44°15′N, 2000 m, 2002, M. Brand 45336 (hb. Brand). Pyrénées Atlantiques: St Engrâce, 0·5 km N of Calla, 0°49.3′W, 43°6.0′N, 600 m, 1992, M. Brand 28205 (hb. Brand).—Spain: Navarra: Pireneos, 3·5 km ENE of Isaba, Barranco de Belabarce, on Pinus, 1000 m, 1992, M. Brand 28537 (hb. Brand).—Slovakia: Hautes Tatras: Tichá Dolina, partie super. de la vallée, tronc mort de Picea, dans la pessière claire, 1300 m, 1966, J. Lambinon 66/T/1100 (LG); the Carpathians, Muránska planina plateau, Mt Kl'ak, N-NE facing slope, nearby Machnatá cave, 19°58′E, 48°47′N, on hard wood of a stump, 1300 m, 2000, A. Guttová, J. Halda, V. Orthová & Z. Palice 4819 (hb. Palice, hb. v.d. Boom).
Exsiccata examined: Lumbsch & Feige, Lich. exs. 92 (B); Magnusson, Lich. exs. 404 (B); Räsänen, Lich. Fenn. 474 (B). Schade, Stolle & Riehmer, Lich. sax. exs. 489 (B); Vezda, Lich. selec. exs. 1858 (B).
Lecanora subravida Nyl
Flora 55: 250 (1872); type: Switzerland, Schaer. Lich. Helv. 544 (M—lectotype, designated here; M—isolectotype!).
(Figs 8C & 11)

Fig. 11. Lecanora subravida, habitus (isolectotype). Scale: 0·5 mm.
Thallus endosubstratal, rather large, patches to 8 cm wide or with scattered small, warted parts, warts to 0·1 mm, in section thin, up to c. 80 μm thick, without a clear cortex and with granules throughout the thallus, upper surface yellowish brown-grey, matt. Photobiont chlorococcoid, cells c. 12–15 μm diam. Prothallus not observed.
Apothecia rather large, constricted at the base, c. 0·5–0·8 mm diam.; margin conspicuous, clearly separated, persistent, c. 0·04–0·1 mm wide, in older apothecia knobbly or folded; disc plane, persistent, pale to medium brown, epruinose. Thalline excipulum well developed, c. 160–190 μm high in section (from upper margin of the parathecium); cortex lacking at the upper part of the excipulum, strongly developed at the lower part, chondroid, with anticlinal, conglutinated hyphae, hyaline, c. 40–80 μm thick, medulla of excipulum filled with algae and rather large granules. Parathecium not developed. Epihymenium pale brown, with fine yellowish granules at the upper surface, N−. Hymenium hyaline, orange-brown granules lacking, c. 50–65 μm high. Hypothecium hyaline, 50–70 μm high. Paraphyses simple to rarely branched, ± constricted at septum, 1·5–2 μm diam., midhymenial cells 6–12 μm long, partly orange-brown pigmented at apices. Asci Lecanora-type, c. 45–65 ×12–14 μm. Ascospores broadly ellipsoid, simple, 9·4–11·0 × 5·0–6·0(−6·2) μm.
Macropycnidia rare, semi-immersed, hyaline at base, dark bluish green-grey c. 120–140 × 90–120 μm; macroconidia curved, pointed at both ends, non-septate (falcate), 12·5–13 × 2·8–3·0 μm. Micropycnidia very rare, immersed, c. 50–70 μm diam.; micro-conidia, curved, c. 7–8 × 0·8 μm. Leptopycnidia abundant, globose, outer rim partly brown, towards the top bluish green-grey pigmented, c. 80 μm diam.; leptoconidia curved, c. 10·8–11 × 0·7–0·9 μm.
Chemistry. K+ yellow, KC+ yellow, P−; usnic acid (major/minor); placodiolic acid (major/minor), ±isousnic acid (minor/ trace), squamatic acid (minor), (TLC and HPLC).
Ecology and distribution. Lecanora subravida occurs on bark or wood, sometimes on rotting trunks of mature Picea, in Germany and Switzerland (19th century collections). Nylander (Reference Nylander1872) reported that he knew this species from Switzerland, Bayern and Savoye (Sabaudia, France). The only recent collection (P. v. d. Boom 8487) is lignicolous on a stump.
Notes. Lecanora subravida is clearly a distinct taxon, not readily confused with any other species in the Lecanora saligna group. Its habitus sometimes resembles L. varia, but that species is never endoxylic, and it even resembles L. conizaeoides, for example, by the well-developed cortex in the apothecia. Its macroconidia are comparable with those of L. quercicola, although these are more attenuate and longer in L. subravida. The ascospores are clearly wider than in all related species. In the recent literature the name L. subravida has appeared only in the checklist for Germany (Scholz Reference Scholz2000). Lecanora subravida was described by Nylander (Reference Nylander1872: 250) who gave only a very short description and noted that the exsiccatum (Lich. Helv. Exs.) Schaerer 544 differs from L. sarcopis Wahlenberg, but he referred to Arnold (Reference Arnold1872: 76), who described L. hageni var. sarcopis Schaerer non Wahlenb. as “sporae ovales 12 μm long, 6–7 μm lat.; spermatia acicularia curvata 12–15 × 1 μm”. Printzen (Reference Printzen2001) regarded L. subravida Nyl. (1872) as a nom. nud. and cited Lecanora varia var. subravida (Nyl.) Nyl. (in Malbranche, Bull. Soc. Amis. Scienc. Nat. Rouen 1881: 120.) as the first valid description of the epithet. However, Nylander (Reference Nylander1872) made an indirect reference to Arnold (Reference Arnold1872) who gave a brief description without publishing a new name for it. This affects the choice of a lectotype, which has to be chosen from material in M seen by Arnold and not from H-Nyl. The measurements of the ascospores and the leptoconidia are somewhat larger in Arnold's description than our own. Studying more material, could probably give a wider range of measurements.
Squamatic acid has been detected in L. subravida, which is a very rare compound in the L. saligna group and has never been found in any other related species.
Specimen examined. France: Vaucluse: NE of Mt. Ventoux, 5 km E of Bédoin, E of St. Colombe, in Quercus forest, on stump, 475 m, 1989, P. van den Boom 8487 (hb. v. d. Boom).
Exsiccata specimens examined. Arnold, Lich. Mon. 32 (L) [Alter Fichtenstrunk im Walde östlich zwischen Gauting und Mühlthal. München, 18 viii 1889, leg. Arnold]; Arnold, Lich. Mon. 1384 (L) [Am morschen Holze alter Fichtenstrunke im Walde nördlich von Hohenbrunn bei München, 13 v 1888, leg. Arnold]; Krypt. exs. Vindob. 2758 (BR). Schaerer, Lich. Helv. 544 (H-Nyl.26246).
Notes on the chemistry of the L. saligna/subintricata agg. [mainly major substances]
There are differences in acetone extracts which may be useful for identification. They may be even more suitable for spot tests and clearer than spot tests on the thallus, especially for the ‘major’ substances. In this section some notes and additional techniques are described to check the identification of some important substances.
Isousnic acid: acetone extract weakly gelatinous, with broadened, clustered granules to spiny globules (growing as crystals from within a droplet); K− yellowish, KC− P−, FeCl3+ red-brown; in light weakly yellowish, UV−. In GE, elongate broadened yellow granules (Huneck & Yoshimura Reference Huneck and Yoshimura1996, fig. 48).
Usnic acid: acetone extract ± dry, with clearly feather-shaped crystals; K+ yellow, C−, KC+ yellow, P−; in light yellowish, UV−. In GE, straight yellow needles (Huneck & Yoshimura Reference Huneck and Yoshimura1996, fig. 50).
Pseudoplacodiolic acid: acetone extract ± comparable with isousnic acid, less gelatinous with broadened, clustered, elongated granules; K+ yellow, C−, KC++ yellow, P−, FeCl3+ red-brown; in light weakly yellowish, UV−. In GE, hexagonal prisms (Huneck & Yoshimura Reference Huneck and Yoshimura1996, fig. 66).
Neousnic acid: always together with isousnic acid, extract not different; in light yellowish, UV−.
7-O-methylnorascomatic acid: UV+ white, Squamatic acid: acetone extract with prisms, FeCl3+ purple. In GE rhombic prisms (Huneck & Yoshimura Reference Huneck and Yoshimura1996, fig. 35). It is a very rare compound in the L. saligna-group.
Paraensic acid D (major), paraensic acid C (minor): acetone extract with nonpolarizing, radiating pattern; in the thallus and apothecia visible as very small oil droplets (<4 μm).
We warmly thank Prof. Jack Elix for his help with the identification of several chemical compounds. We are grateful to the keepers of the following herbaria for the loan of specimens: Prof/Drs E Sérusiaux (Liège), P. Diederich (Strassen), J. Etayo (Pamplona), H. Sipman (Berlin) and the curators of BR, E, L, LUX, M, H-Nyl, GZU and MAF. We are also very grateful to Dr H. Sipman for his contribution regarding photographs, for help with Latin translations and for valuable remarks about an earlier version of the manuscript and to the referees Dr B. Coppins and Dr C. Printzen for constructive reviews of the manuscript.