


1. Introduction
The Anning family, especially their most famous member, Mary Anning, collected thousands of fossils from Lyme Regis on the Dorset coast, and helped to establish Lyme Regis as a fossil haven for both professionals and amateurs (Torrens, Reference Torrens1995). The first ichthyosaur scientifically identified and described was collected by Mary Anning and her brother Joseph (Torrens, Reference Torrens1995). Mary Anning's later finds were just as significant, including the first relatively complete skeleton of a plesiosaur, the first British pterosaur, and the first fossilized ink sac of a cephalopod, all from her collecting sites in Lyme Regis (Torrens, Reference Torrens1995). Awareness of her fossil discoveries reached the wider palaeontological community, including such prominent figures as William Buckland, William Conybeare, Henry De la Beche, Georges Cuvier and Gideon Mantell, as well as less renowned collectors who also were interested in acquiring specimens (Torrens, Reference Torrens1995). Specimens collected and prepared by Anning were sold to collectors, and many were subsequently donated to museums in the UK and elsewhere (Torrens, Reference Torrens1995). Notable collections of Mary Anning specimens are housed in the Natural History Museum, London, and the Sedgwick Museum, Cambridge. In fact, several of Anning's fossils were purchased by Adam Sedgwick for the Woodwardian Museum, now the Sedgwick Museum, Cambridge (Price, Reference Price1986).
Although museums were careful to credit the donors of their ichthyosaur specimens, the collector was rarely acknowledged for the discovery, as was the case with Mary Anning (Torrens, Reference Torrens1995). It was not until 1986 that Anning specimens in the Sedgwick Museum were identified and figured (Price, Reference Price1986). The Anning specimens included several ichthyosaur skulls and nearly complete skeletons (Price, Reference Price1986). This paper describes one of those specimens, which was purchased by Adam Sedgwick in 1832 for £35 (Price, Reference Price1986). The presacral portion of the specimen is nearly complete and well preserved, and illustrates features of the species that have not previously been documented.
Institutional abbreviations. CAMSM – Sedgwick Museum, Cambridge University, UK; NHMUK – (formerly BMNH) Natural History Museum, London, UK.
2. Materials and methods
CAMSMX.50187 was first figured by Price (Reference Price1986, fig. 4A; the specimen was unregistered at the time). In a letter to Adam Sedgwick (20 May, 1832), Mary Anning described a recently discovered Ichthyosaurus communis about five feet long, consisting of a skull, vertebrae, ribs, sternum, both hindfins, and pelvis; and noted that it was ‘the only specimen yet discovered having the pelvis clearly developed’ (Price, Reference Price1986). Price (Reference Price1986) identified the specimen described herein as the one that Sedgwick purchased from Mary Anning, based on the preservation, length of the specimen, and the unusually well preserved pelvis. He also noted that the specimen is the only one in the Sedgwick Museum collection that fits Anning's description. Price's (1986) measurement of the specimen (about five feet long, including the tail) was in agreement with that of Anning. The tail appears continuous with the presacral portion of the skeleton, although the centra in the anterior caudal region belong to another individual and were added to the specimen (see below).
Because this is a Mary Anning specimen, the provenance is almost certainly Lyme Regis, west Dorset, her prime collecting location (Torrens, Reference Torrens1995). Given the practice when it was collected, information on the specimen's stratigraphic horizon is unrecorded, although most of the ichthyosaurs were probably collected from the Lower Jurassic, lower Sinemurian bucklandi and semicostatum zones (McGowan, Reference McGowan1974a ). I. breviceps and other species from Lyme Regis that are in historical collections, are given a range from the upper Hettangian to the Lower Sinemurian (McGowan, Reference McGowan1974a , Reference McGowan b ; McGowan & Motani, Reference McGowan and Motani2003).
CAMSMX.50187 is a nearly complete skeleton lying on its left side (Fig. 1). The presacral skeleton is preserved in two blocks of matrix that are separated by a large crack. The two presacral blocks and the tail have been set in plaster in a wooden frame, and in some areas, chisel-like markings were added to the plaster. The original matrix, as well as the gastralia and two ammonites, have been painted over with a thick grey paint, presumably to make the mount more aesthetically pleasing, but the edges of some of the bones (e.g. the scapulae) have been covered by paint. The specimen had previously been on display (Price, Reference Price1986).

Figure 1. Complete specimen of Ichthyosaurus breviceps, CAMSMX.50187. Scale bar represents 10 cm. © Sedgwick Museum of Earth Sciences, University of Cambridge, 2012. Reproduced with permission.
The large crack that separates the two presacral blocks is filled entirely with plaster. Ribs are continuous across the crack, although sections of the ribs have been lost and there are no impressions in the plaster filling. Slightly posterior to the pelvic bones is a break in the vertebral column, after the 44th centrum, which marks the edge of the second block. Posterior to the break, in the anterior caudal region, the centra appear to have been added, although the poorly preserved centra in the distal portion of the tail may be original and in situ. The latter are in line with the curve of the tail, although partly disarticulated.
The ten centra immediately posterior to the break are rotated through 180° with respect to the presacral skeleton, so that the ventral aspect is oriented dorsally, i.e. upside down relative to the presacral skeleton (Fig. 2). Immediately distal to this series is another sequence of eight centra that are rotated through 90° so that the neural canals face the right side of the skeleton. The two series of centra are surrounded by plaster (Fig. 2). The positions of the rib articulations indicate that the anterior series is from the posterior dorsal region, and the centrum shape suggests that they belong to a different species (see below). Thus these eighteen ‘caudal’ centra do not belong to the presacral skeleton.

Figure 2. Caudal region of CAMSMX.50187, with centra completely surrounded by plaster. The anterior two series of 10 and 8 centra are most likely replacements for the originals (see text). Scale bar represents 10 cm. © 2012. Sedgwick Museum of Earth Sciences, University of Cambridge. Reproduced with permission.
Mary Anning is not known for composite specimens, and in fact did not approve of collectors who altered specimens (see letter in Lang, Reference Lang1945, p. 171). Moreover, one of her specimens, a complete Plesiosaurus skeleton discovered in 1823, was the subject of an intense debate questioning its authenticity (Torrens, Reference Torrens1995). Having been unjustly accused of forgery (if only by implication) early in her career, it is unlikely that she would have altered a specimen that she offered for sale. Mary Anning's original length measurement was about five feet (Price, Reference Price1986), but the length of the presacral skeleton is only about 109 cm (3.6 ft). Thus the tail must have been included in the original measurement. The distal caudal centra appear to belong to the specimen, based on their colour and shape, but they are very poorly preserved. Perhaps the introduced centra replaced or covered original, similarly poorly preserved ones to make a better display specimen. Interestingly, another ichthyosaur specimen previously on display in the Sedgwick Museum (CAMSM J 13579), donated by Thomas Hawkins, also has a series of centra that do not belong to it. In this specimen, caudal centra were placed upside-down in the dorsal region of a Leptonectes from the Lias of Street (Lomax & Massare, Reference Lomax and Massare2012).
Our measurements of the specimen were taken using digital callipers for features smaller than 15 cm, and recorded to the nearest 0.1 mm. For larger features, a measuring tape was used, and measurements were recorded to the nearest 1 mm.
3. Description
3.a. Skull and jaw
Our measurements of the skull and jaw (Table 1) follow those of McGowan (Reference McGowan1973). The skull is 30–31 cm long, but the posterior portion is crushed from the top, so a precise measurement is not possible. The jaw is missing the anterior tip, although the premaxilla is complete so that the jaw length can be measured accurately (Figs 3, 4). The lachrymal is pushed forward because the skull is crushed from above. We attempted to account for this in the measurement of the preorbital length by measuring from the posterior, inner edge of the lachrymal rather than from the edge of the orbit, a difference of about 1 cm. This produces a relative snout length (ratio of preorbital length/jaw length, after McGowan, Reference McGowan1973) of 0.53, which is well within the range for I. breviceps (< 0.57; McGowan, Reference McGowan1974b ). The maxilla is curved along the tooth row, as in the holotype of I. breviceps, NHMUK 43006 (McGowan, Reference McGowan1974b , fig. 7A). The premaxilla is slightly downturned at the tip, but not markedly curved as in the holotype; however the downturn does not occur in all I. breviceps specimens (e.g. NHMUK R 216, NHMUK 33277). The orbit is 9 cm long, but it is deformed because of the crushing so the length might be high. Thus the ratio of the orbit length to the jaw length of 0.27 is a maximum value. It is within the range for I. breviceps (≥ 0.26), but also within the range of I. conybeari (< 0.28) and close to the range for I. communis (< 0.26; McGowan & Motani, Reference McGowan and Motani2003). Given the small differences in ratios and the potential for distortion by crushing, the use of an orbit ratio might not be very reliable.
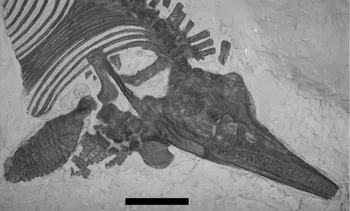
Figure 3. Skull, pectoral elements, and forefin of CAMSMX.50187. Note the high neural spines on the cervical centra. Scale bar represents 10 cm. © 2012. Sedgwick Museum of Earth Sciences, University of Cambridge. Reproduced with permission.
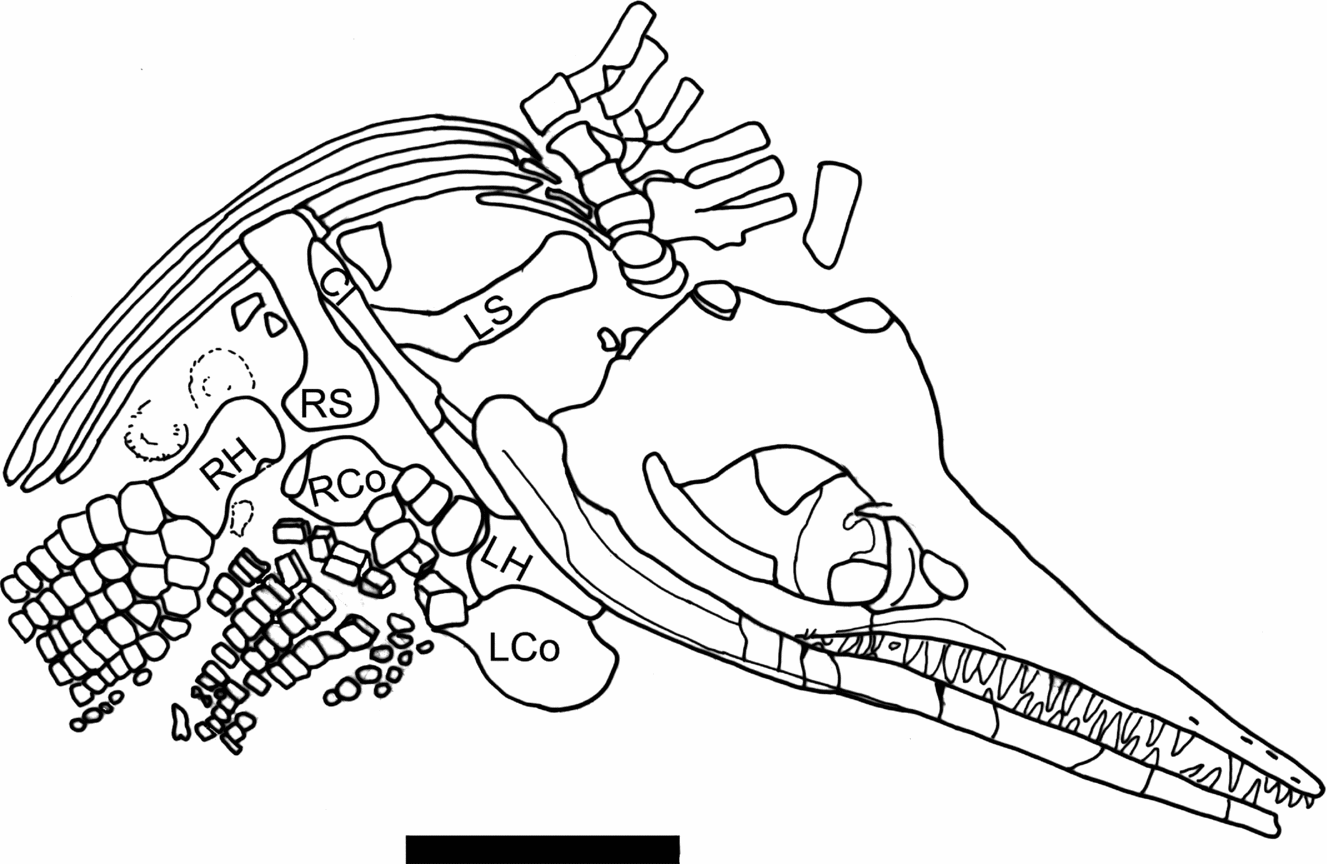
Figure 4. Line drawing of a portion of Figure 3, identifying the pectoral girdle elements and humeri. LH – left humerus; RH – right humerus; LS – left scapula; RH – right scapula; LCo – left coracoid; RCo – right coracoid; Cl – clavicle, probably the right. The two circular objects between the ribs and right humerus are ammonites covered in thick paint. Scale bar represents 10 cm.
Table 1. Skull measurement of CAMSMX.50187

Lengths use definitions of McGowan (Reference McGowan1973).
*Estimated length.
There are at least ten fairly large teeth in the maxilla, with about three teeth per 2 cm of length, although the roots are exposed making the teeth appear larger. The anterior maxillary teeth, however, are considerably larger than the premaxillary teeth. There are at least 18 teeth in the premaxilla, spaced at 3–4 teeth per 2 cm length. The longest maxillary tooth crown is at least 0.9 cm long (not including the tip which is buried). The longest premaxillary tooth crown is 0.6 cm long, not including the tip, which is missing. All teeth have large, somewhat bulbous roots, and the crowns have coarse longitudinal striations.
3.b. Pectoral girdle and forefin
Both scapulae are present, but only the right one is exposed clearly. The scapula is long relative to its width (Table 2). The proximal end looks rounded in Figures 3 and 4, but it is covered by paint so its true shape is unclear. A complete clavicle 9.5 cm long is preserved partially overlying the distal end of the right scapula, so we assume that it is the right clavicle. The articular surface of the right coracoid is exposed and appears to be touching the proximal end of the right scapula. Most of the shaft and distal end of the left scapula is exposed anterior to the clavicle. The left coracoid is preserved under the left humerus, but only the posterior medial portion of it is exposed (Figs 3, 4).
Table 2. Measurements of elements of the appendicular skeleton of CAMSMX.50187
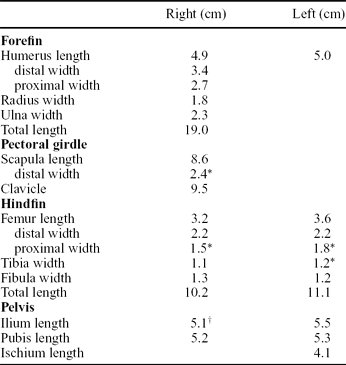
Note: We assume that the pelvic elements closest to each hindfin are from the same side of the animal as the hindfin (e.g. the elements nearest the left hindfin are the left pelvic elements).
*Estimated width because edge of the bone is covered or poorly defined.
†Right ilium is broken and partially covered.
A nearly complete right forefin, 19 cm long, is preserved in dorsal aspect (Fig. 3). The morphology is typical of Ichthyosaurus, especially in the number of digits and the anterior digital bifurcation (Motani, Reference Motani1999). The right humerus is about 1.5 times the length of the right femur (Table 2). The anterior edge of the shaft is partly covered by paint, but an anterior facet about 1 cm high is visible on the distal edge. None of the elements of the forefin are notched. The radius and ulna are both wider than long, and meet along a broad, straight contact. The ulna is wider than the radius (Table 2). The ulnare is rectangular, and wider than either the intermedium or radiale, which are about equal in size. The forefin is somewhat unusual in that the digital bifurcation, typical of Ichthyosaurus forefins, occurs in the metacarpal row. It is difficult to determine which digit bifurcates, as the first element of the additional digit is situated between metacarpals 2 and 3 (using the terminology of Motani, Reference Motani1999) and articulates with both along straight facets (Fig. 3). The anteriormost digit is slightly narrower than the next two. The phalanges are closely packed and rectangular, but become rounded towards the distal end of the forefin, at the 6th or 7th phalanx in Digit V and the 9th or 10th phalanx in Digit II. One anterior accessory digit, with rounded elements, begins at the second row of phalanges. No posterior accessory digits are evident, although the posterior edge of the forefin abuts against a thick coating of paint. A crack runs across the distal end of the forefin, and the preservation looks different on either side of the crack. The accessory digit and Digits IV and V line up well across the crack, so it is unlikely that the forefin is a composite. A partial, largely disarticulated, left forefin is also preserved with numerous rectangular-shaped phalanges. It includes the humerus, which is partially overlain by the lower jaw (Figs. 3, 4). The left humerus is about 1.4 times the length of the left femur (Table 2). Preserved on the same side of the left forefin are two digits running side by side, both with rounded phalanges. The smaller of the two is an accessory, but the larger one could be another accessory or the distal portion of one of the primary digits. It is impossible to tell for certain whether the digits are on the anterior or posterior side of the forefin.
3.c. Pelvic girdle and hindfin
We assume that the pelvic bones on the right side of Figure 5 are the left pelvic elements as they are closest to the left hindfin (see below); similarly, the pelvic bones on the left side of Figure 5 are assumed to be the right pelvic elements. The pelvis is poorly known in I. breviceps (McGowan, Reference McGowan1974b ), but this specimen preserves all three bones from both sides of the pelvic girdle, although the right ischium is partly covered by the right pubis (Fig. 5). The ilium is a long and slightly curved rod, and is the longest of the three elements. The pubis is noticeably longer and wider than the ischium, and only slightly shorter than the ilium (Table 2). The pubis is somewhat expanded at both ends, whereas the ischium is only slightly expanded and is more rod-like. All three bones are longer than the femur.
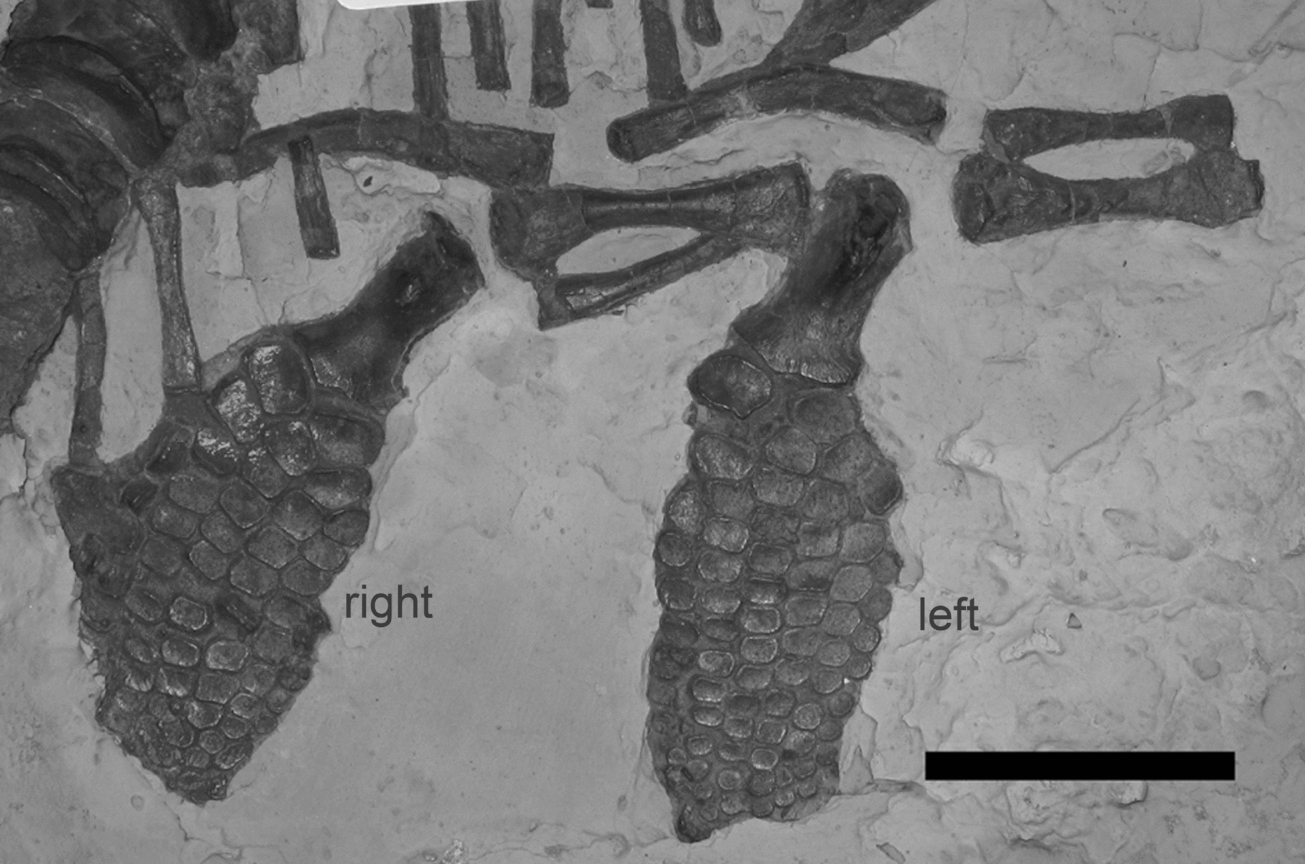
Figure 5. Pelvis and hindfins of CAMSMX.50187. The left hindfin, in ventral view, and pelvic bones are to the right in the photo. Note that the pubis is larger than the ischium, and the ilium is larger than the pubis. The right hindfin, in dorsal view, is on the left, closer to the vertebral column. Anterior is to the right for both hindfins. Scale bar represents 5 cm. © 2012. Sedgwick Museum of Earth Sciences, University of Cambridge. Reproduced with permission.
The right hindfin, the more posterior of the two, is preserved in dorsal view, whereas the left hindfin is in ventral view (Fig. 5; Maxwell, Zammit & Druckenmiller, Reference Maxwell, Zammit and Druckenmiller2012). A vertical facet is present on the distal anterior edge of both femora (on the right side of each in Figure 5). The femur is much longer than wide, and is wider at the distal end than at the proximal end (Table 2). On the right hindfin, the tibia and fibula meet along a narrow contact, with the intermedium separating the distal halves of the bones. They are about the same width, but the tibia is more equidimensional in shape than the fibula. The fibula is displaced on the left hindfin, and the fibulare is missing. None of the elements of the hindfin are notched. Both hindfins have a total of six digits, four primary digits, and an anterior and posterior accessory digit, both of which originate at the first row of phalanges. The left hindfin has 10 phalanges in the longest digit. The entire left hindfin is 11.1 cm long and the right is 10.2 cm long, about two thirds the length of the right forefin.
3.d. Vertebral column
Twenty-two centra are on the anterior block of the specimen. The 15th centrum is lying flat, but the others on either side are articulated. The 23rd centrum is missing at the crack between the blocks, as indicated by the ribs that are articulated, or nearly so, with the centra. An additional 21 centra are present on the posterior block, bringing the total count to 44. The 44th centrum shows touching parapophysis and diapophysis. We estimate that the first unicipital rib would have occurred at the 45th or 46th vertebrae, which agrees with the presacral count in the diagnosis of I. breviceps (<46) in McGowan (Reference McGowan1974b) and Owen (Reference Owen1881), but is slightly higher than the count (<44) given in McGowan & Motani (Reference McGowan and Motani2003). The posterior four ribs (articulating with 41st to 44th centra) are short compared to the others, indicating their proximity to the pelvic region.
The presacral neural spines (including the neural arch) are quite high where they are preserved, consistent with the original diagnosis of Ichthyosaurus breviceps (Owen, Reference Owen1881). The first neural spine, isolated in the matrix dorsal to the skull, is twice as wide as the second, which suggests that it belongs to the atlas-axis. The height of the neural spine is 1.7–2.0 times the height of the centrum on mid-dorsal centra (25th–29th). Those on the more anterior centra are at least as high. The presacral centra are long relative to their height, giving them a short, fat profile in lateral view (Fig. 6). The centra have distinct posterior and anterior rims that stand out markedly from the sides of the centrum. For the posterior 20 presacral centra (on the second block), the average height/length ratio is 1.8 (Table 3). By comparison, the first three ‘caudal’ centra have height/length ratios of 3.0, 3.1, and 3.3, respectively. Furthermore, these centra show separated diapophysis and parapophysis, whereas the last centrum of the presacral block shows them touching. Thus the series of ten ‘anterior caudal’ centra of this mount are actually posterior dorsal centra from a different individual, and probably from a different species as well, judging by their shape (Fig. 2). The next series of eight ‘caudal’ centra are markedly smaller and are rotated relative to the anterior series so that the dorsal surface is facing laterally. Their correct position in the vertebral column cannot be determined because rib articulations are buried. The bone colour is lighter and the preservation looks better than that of the more posterior centra, so it is likely that these have also been added. Following a small gap are four articulated centra and three centra lying flat on the matrix. These are poorly preserved, but the colour is similar to that of the presacral skeleton. The four are shorter relative to their height than the presacral centra, however. There is then a gap of 8 cm, in which there are indentations in the plaster for four or five additional centra. The proximity of these indentations to the ‘chisel marks’ in the plaster raises the question of whether these impressions are original. Following this large gap are 13 small, articulated centra, poorly preserved but of the same dark colour as the presacral centra. Thus a total of 40 centra are present in the caudal region, ignoring what may be missing in the gaps and the four or five questionable impressions in the plaster.
Table 3. Centrum dimensions and centrum height/length ratio (H/L) for mid-dorsal and posterior dorsal centra of CAMSMX.50187

Note: Height was measured at the rim of the centrum. Length was measured from rim to rim of the centrum. The last three centra in the table (45–47) are in the anterior caudal position on the specimen, but are posterior dorsal centra belonging to another individual.
*Estimated measurement because centrum is poorly preserved or partially buried.

Figure 6. Dorsal centra of CAMSMX.50187 are long relative to their height, producing a short, fat profile in lateral view. Scale bar represents 10 cm. © 2012. Sedgwick Museum of Earth Sciences, University of Cambridge. Reproduced with permission.
The length of the presacral vertebral column, measured along the column, is 78 cm. If the distal caudals do indeed belong to this specimen, then the caudal portion, measured along the vertebral column and across the gaps, is about 45 cm. Adding an additional 30 cm for the length of the skull results in a total length of 153 cm, about five feet long, as originally described by Anning (Price, Reference Price1986). Thus it is very likely that at least the two series of rotated centra in the anterior and mid-caudal regions were set there to replace the original caudal centra. The vertebral column stops just short of the bend in the tail (see Section 4). Given the counts above, more than 84 preflexural centra are present on this specimen, which is much higher than what is given in the diagnosis for I. breviceps (<74; McGowan & Motani, Reference McGowan and Motani2003). This discrepancy can be explained if the original, longer caudal centra were replaced with shorter, better preserved ones now seen on the specimen. The distal portion of the tail is probably authentic.
4. Discussion
We assign specimen CAMSMX.50187 to I. breviceps, not to I. communis as Mary Anning identified it. I. breviceps was first recognised by Owen (Reference Owen1881) more than thirty years after Anning's death (Torrens, Reference Torrens1995). The relatively short snout and relatively large orbit, as given in the ratios mentioned above, are key features in the diagnosis of I. breviceps (Owen, Reference Owen1881; McGowan, Reference McGowan1974b ; Maisch & Matzke, Reference Maisch and Matzke2000; McGowan & Motani, Reference McGowan and Motani2003). Owen (Reference Owen1881) also noted the very high neural spines on the dorsal vertebrae, which are present on this specimen, although subsequent workers did not use this characteristic in species diagnoses.
This is the largest specimen of I. breviceps reported in the literature. McGowan (Reference McGowan1974b , fig. 1) graphed a maximum observed jaw length of about 25 cm for I. breviceps, whereas this specimen has a jaw length of 33.5 cm, about one third larger. The length of this specimen, measured along the vertebral column is approximately 153 cm, but this is probably not the full preflexural length. Buchholtz (Reference Buchholtz2001, fig. 5) indicated that the preflexural length of the tail in Ichthyosaurus is 60–65% of the length of the presacral vertebral column. With a presacral length of 78 cm, this individual would have had a preflexural tail length of 47–51 cm, slightly longer than what is preserved. Adding the skull length of 30–31 cm would give a total length of 155–160 cm, or approximately 1.6 m. This is within the range given by McGowan & Motani (Reference McGowan and Motani2003), but slightly larger than the maximum length of 1.5 m given by McGowan (Reference McGowan1974b).
CAMSMX.50187 is noteworthy in that it preserves a complete pelvic girdle, which provides new information on the pelvis morphology of I. breviceps. The ilium is markedly longer than the ischium and pubis. McGowan & Motani (Reference McGowan and Motani2003, fig. 60) showed the ilium length at about three quarters of the length of the pubis and ischium in I. communis, although McGowan (Reference McGowan1974b , fig. 6B) illustrated the three bones at about the same length. Furthermore, in CAMSMX.50187, the pubis is much longer than the ischium, whereas in I. communis, the two bones are nearly the same size (McGowan & Motani, Reference McGowan and Motani2003, fig 60). The pelvis is not preserved often and has not been used very much taxonomically to distinguish species within genera, although it has been used in diagnoses of genera. This specimen, however, suggests that the relative size of the pelvic bones may be a feature that distinguishes I. breviceps and I. communis. We are investigating this further in our research in progress on the genus Ichthyosaurus.
A second feature worthy of further study is the shape of the centra (Fig. 6). The posterior dorsal centra have unusually low height/length ratios, comparable to the ratios for Eurhinosaurus and Suevoleviathan for the same part of the vertebral column (Massare et al. Reference Massare, Buchholtz, Kenney and Chomat2006, fig. 5). The aforementioned genera have more flexibility in their vertebral columns and are therefore more manoeuvrable but slower swimmers (Buchholtz, Reference Buchholtz2001; Massare et al. Reference Massare, Buchholtz, Kenney and Chomat2006), and this could possibly apply to I. breviceps. In any case, the short, ‘fat’ lateral profile of the centra may prove to be a taxonomic character for I. breviceps.
Acknowledgements
Matt Riley and Sarah Finney of the Sedgwick Museum, Cambridge were extremely helpful in giving us access to the specimen, and we are grateful for their assistance. Images of the specimen are reproduced with permission of the Sedgwick Museum of Earth Sciences, University of Cambridge. We thank Michael A. Taylor for providing useful historical information and references, as well as reviewing an earlier version of the manuscript; and Hugh Torrens for comments about Mary Anning. Finally we would like to thank Judith Pardo-Pérez and an anonymous reviewer for their helpful suggestions. JAM travel was supported in part by a United University Professions Faculty Development Grant, SUNY Brockport. DRL travel costs were covered in part by a Karl Hirsch Memorial Grant from the Western Interior Paleontological Society.









