


1. Introduction
Upper Devonian deposits were first recognized in the Saoura Valley (Algeria, Ougarta Basin) by Haug (Reference Haug1903) on the basis of cephalopods (goniatites and clymeniids), which are particularly abundant in some levels (see also Menchikoff, Reference Menchikoff1930). The first comprehensive study on the geology of this area was by Gautier (Reference Gautier1906) and refined by Alimen et al. (Reference Alimen, Le Maître, Menchikoff, Petter and Poueyto1952), who described several important sections. The richness in ammonoids enabled Petter (Reference Petter1959) and Göddertz (Reference Göddertz1987) to discriminate the zonation established by Wedekind (Reference Wedekind1908, Reference Wedekind and Salomon1926) in Germany and subsequently modified by several authors (e.g. Korn, Klug & Reisdorf, Reference Korn, Klug and Reisdorf2000; Korn, Reference Korn2004).
Lower and Middle Devonian brachiopod faunas from the Ougarta were first described by Le Maître (Reference Le Maître1952) and revised in part by Boumendjel et al. (Reference Boumendjel, Brice, Copper, Gourvennec, Jahnke, Lardeux, Le Menn, Melou, Morzadec, Paris, Plusquellec and Rachebœuf1997), Ouali Mehadji et al. (Reference Ouali Mehadji, Elmi, Rachebœuf and Mekhali2004), and Brice et al. (Reference Brice, Boumendjel, Rachebœuf and Mottequin2011) but, until recently, Upper Devonian brachiopods were poorly known (e.g. Brice, Legrand-Blain & Nicollin, Reference Brice, Legrand-blain and Nicollin2005, Reference Brice, Legrand-blain, Nicollin, Becker and Kirchgasser2007). Brachiopods are of interest because they flourished along the northern margin of Gondwana in neritic facies sometimes unfavourable to ammonoids and conodonts, and are therefore valuable tools for biostratigraphy in such palaeoenvironmental contexts, especially regarding the Devonian–Carboniferous boundary (Mottequin, Brice & Legrand-Blain, Reference Mottequin, Brice and Legrand-Blain2014).
This paper is the first comprehensive study of the diverse and small rhynchonellide brachiopods from the Famennian (Marhouma Formation) Saoura Valley where they are the dominant element in the brachiopod assemblages, although they also occur in benthic communities (cf. Crônier et al. Reference Crônier, Malti, François, Benyoucef and Brice2013 for trilobites). Their diversity and palaeoecology are also discussed.
2. Geological setting and material
The material described in this paper comes from the Saoura Valley in the eastern part of the Ougarta Basin (Algerian Sahara), about 350 km SSW of Béchar (Fig. 1). The rhynchonellides were collected by F. Z. Malti in 2006–2008 during fieldwork for her PhD thesis on the Marhouma Formation, a unit first described by Poueyto (unpub. internal report, Société nationale de Recherche et d’Exploitation de Pétrole en Algérie (SN-REPAL), 1965) and Bastien et al. (unpub. internal report, SN-REPAL, 1965) (e.g. Ouali Mehadji et al. Reference Ouali Mehadji, Bendella, Atif, Belkacemi and Abbache2012 for more details), following a NNW–SSE transect along the Saoura Valley and more particularly in Ouarourout (30°10′30″N; 2°14′30″W), Gara Diba (30°7′38″N; 2°12′30″W), Cheffar El Ahmar (29°57′25″N; 2°7′15″W) and Tamtert-Zereg (29°54′30″N; 1°49′30″W) (Figs 1–3). She gathered abundant and diverse marine macro- and microfaunas, including brachiopods, ammonoids (goniatites and clymeniids identified by D. Korn), trilobites, conodonts, and spores.
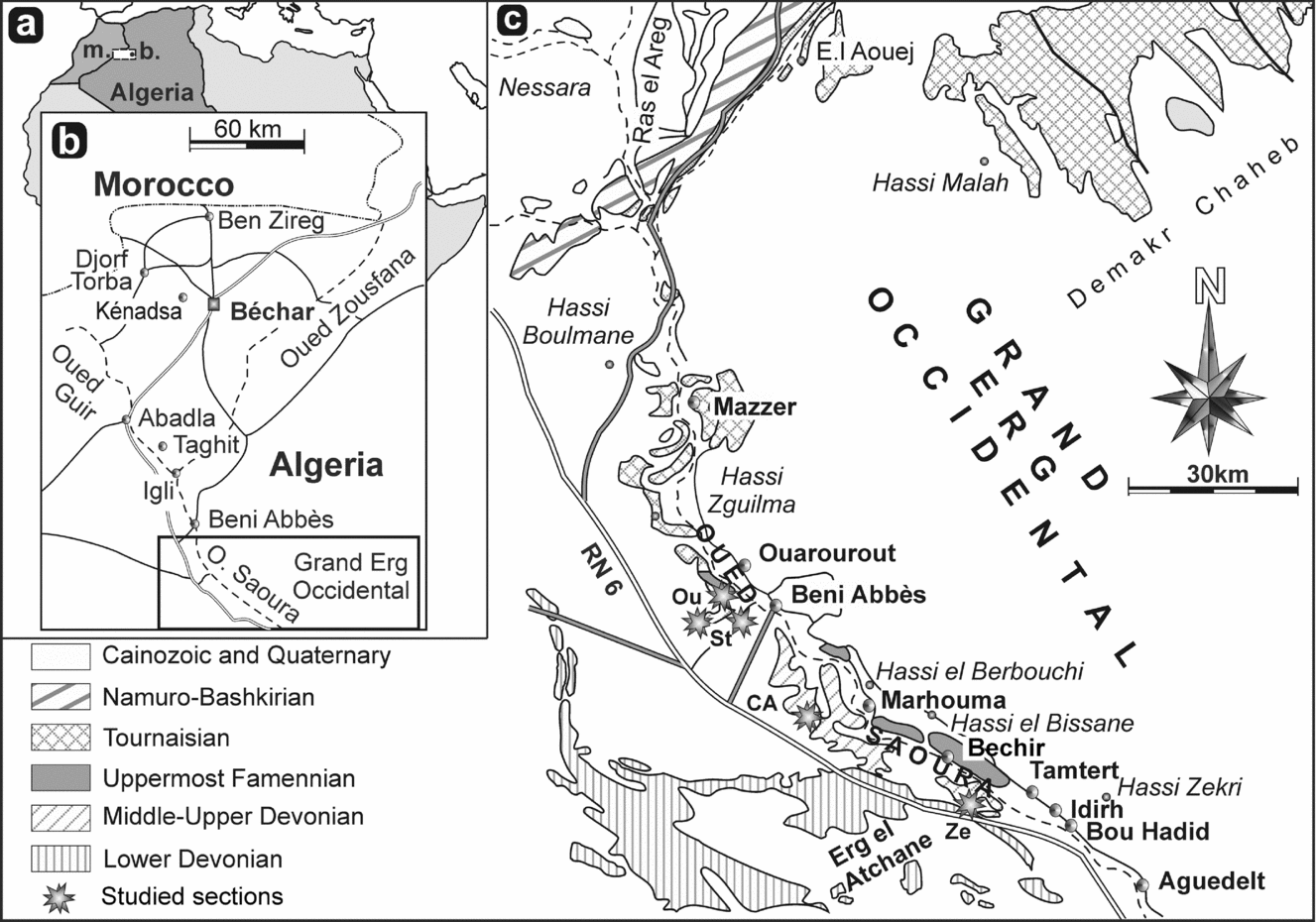
Figure 1. (a, b) Geographic locations of the Saoura Valley in the Ougarta Basin in Algeria. Abbreviations: b. – Béchar; m. – Morocco. (c) Location of studied sections (CA – Cheffar El Ahmar; St – Gara Diba; Ou – Ouarourout; Ze – Tamtert–Zereg).

Figure 2. Distribution of rhynchonellides, ammonoids, conodonts and palynomorphs within the Marhouma Formation in the Saoura Valley (Ougarta Basin, Algeria). Abbreviations: G. – Grandispora; P. – Palmatolepis; U. – Umbellasphaericum devonicum.

Figure 3. Distribution of rhynchonellides within the Marhouma Formation in the Cheffar El Ahmar, Gara Diba, Ouarourout and Tamtert–Zereg sections.
Subdivisions of the Famennian Stage are those used by Becker, El Hassani & Tahiri (Reference Becker, Hartenfels, Aboussalam, Tragelehn, Brice and Hassani2013, fig. 2) in alignments between the conodont and ammonoid zonations (see also Clausen, Weddige & Ziegler, Reference Clausen, Weddige and Ziegler1993; Korn, Reference Korn2002; Crônier et al. Reference Crônier, Malti, François, Benyoucef and Brice2013). The acronyms do I to do VI refer to successive Frasnian–Famennian cephalopod zones (Wedekind, Reference Wedekind1908) with ‘do’ meaning Upper Devonian.
F.Z. Malti (unpub. PhD thesis, Oran University and Abou Bekr Belkaid University, 2012) discriminated four successive members in the Marhouma Formation (Figs 2–3), the latter totalling about 260 m according to Ouali Mehadji et al. (Reference Ouali Mehadji, Bendella, Atif, Belkacemi and Abbache2012); it is overlain by sandstones of the uppermost Famennian Ourarourout Formation (Fabre, Kazi-Tani & Moussine-Pouchkine, Reference Fabre, Kazi-Tani, Moussine-Pouchkine and Fabre2005).
Member 1 consists of calcareous shales devoid of rhynchonellides; it is Late Frasnian in age (do Iβ) on the basis of goniatites (Manticoceras sp.) and entomozoaceans (Casier, Reference Casier1983).
Member 2 includes silty shales with ‘griotte’ nodules; it is divided into two submembers (a and b). The term ‘griotte’ is applied to nodular to pseudonodular limestones rich in ammonoid cephalopods (Benhamou et al. Reference Benhamou, Abbache, Elmi, Mekhali, Rachebœuf, Ouali-Mahadji and Boumendjel2004). Submember a comprises silty shales with numerous specimens of Evanidisinurostrum saouraense sp. nov. in the Gara Diba section and a few Sphaeridiorhynchus sp. in the Cheffar El Ahmar section in thin limestone lenses rich in horizontal burrows. The age of this submember cannot be precisely determined due to the lack of conodonts and goniatites; the rhynchonellides suggest a possible early Famennian age. Submember b consists of alternating shales and ‘griotte’ limestone levels, sometimes in a succession of centimetre-thick beds. A limestone bed of this submember in the Gara Diba section has produced the conodont Palmatolepis minuta subtilis in the interval spanning the P. triangularis to P. trachytera zones (do II). An interval of ‘griotte’ limestone in the Cheffar El Ahmar section produced Leptoterorhynchus sp. associated with Maenoceras; this rhynchonellide was previously known from the middle Famennian.
Member 3 includes the ‘griotte’ limestone which is divided into three submembers (a, b, c); one ‘griotte’ level produced several conodonts indicative of the middle–upper Palmatolepis expansa Zone (upper–uppermost Famennian). This submember in the Tamtert–Zereg section is late Famennian on the basis of Platyclymenia Zone ammonoids. It has a diverse rhynchonellide fauna: Centrorhynchus sp., Leptoterorhynchus sp., Phacoiderhynchus aff. antiatlasicus Sartenaer, Reference Sartenaer2000, Hadyrhyncha cf. hadyensis Havlíček, Reference Havlíček1979 and Novaplatirostrum sp.
Member 4 consists of shales with some ‘griotte’ levels, but also sandy and silty deposits of turbiditic origin; these are particularly thick at Ouarourout and Tamtert–Zereg. It has three submembers (a, b, c), but only the oldest submember produced rhynchonellides (Pugnaria sp.).
3. Systematic palaeontology
The material described and figured here is housed at the Museum of the Central University of Algiers (MUA) and was investigated by Denise Brice. The supraspecific classification follows Savage et al. (Reference Savage, Manceñido, Owen, Carlson, Grant, Dagys, Sun and Kaesler2002) and Savage (Reference Savage and Kaesler2007) for the Order Rhynchonellida (except if stated otherwise); where used, open nomenclature follows Feldman's (Reference Feldman1994) rules. Measurements of specimens assigned to Evanidisinurostrum saouraense sp. nov. are presented in Appendix 1 (see online supplementary data at http://journals.cambridge.org/geo).
Abbreviations. L, shell length; lc, lateral costae; mc, median costae; T, shell thickness; W, shell width; Ws, sulcus width.
Order rhynchonellida Kuhn, Reference Kuhn1949 Superfamily rhynchotrematoidea Schuchert, Reference Schuchert and Eastman1913 Family trigonirhynchiidae Schmidt, Reference Schmidt1965 Subfamily trigonirhynchiinae Schmidt, Reference Schmidt1965 Genus Centrorhynchus Sartenaer, Reference Sartenaer1970
Type species. Camarotoechia baitalensis Reed, Reference Reed1922, Famennian, Pamir.
Centrorhynchus sp.
Figure 4a–b
Material. One incomplete articulated specimen (Ze 2/8) from the Tamtert–Zereg section, Marhouma Formation, Member 3, Submember c.
Description. Shell small (16 mm in width), probably widest at mid-length, subpentagonal in outline (aa = 106°). Valves covered by angular, simple costae arising at beaks; four median and eight lateral costae in ventral valve (not known in dorsal valve).Ventral valve almost flat with obsolete, poorly defined sulcus; beak suberect. Dorsal valve (preserved only posteriorly) convex.
Remarks. This specimen is assigned to Centrorhynchus on the basis of its external morphology but additional material is required for a species-level identification. Centrorhynchus is a cosmopolitan genus assigned to the Trigonirhynchiinae by Nicollin & Brice (Reference Nicollin and Brice2004); it occurs first in the lower Famennian (Baliński, Reference Baliński1995). In North Africa, the genus has been recognized in the eastern part of the southern margin of the Tindouf Basin by Brice (in Gourvennec, Bitam & Robardet, Reference Gourvennec, Bitam and Robardet1997) and on its northern flank (Brice in Brice, Legrand-Blain & Nicollin, Reference Brice, Legrand-blain and Nicollin2005) and doubtfully in the uppermost Famennian of Tafilalt by Brice (in Brice, Legrand-Blain & Nicollin, Reference Brice, Legrand-blain and Nicollin2005 and in Becker et al. Reference Becker, El Hassani and Tahiri2013). According to ammonoids from the Tamtert–Zereg section, our specimen is of late Famennian age (do IV or IV/V, upper Platyclymenia Zone).
Subfamily hemitoechiinae Savage, Reference Savage, Copper and Jin1996
Genus Paurogastroderhynchus Sartenaer, Reference Sartenaer1970
Type species. Camarotoechia (?) nalivkini Abramian, Reference Abramian1957, Famennian, Armenia.
Paurogastroderhynchus presaharensis Brice in Brice, Legrand-Blain & Nicollin, Reference Brice, Legrand-blain and Nicollin2005
Figure 4c–d

Figure 4. Rhynchonellide brachiopods from the Marhouma Formation, Upper Devonian (Famennian), Saoura Valley, Algeria (see Fig. 1). (a, b) Centrorhynchus sp., MUA/1094/020: incomplete specimen in ventral and posterior views. (c, d) Paurogastroderhynchus presaharensis Brice in Brice, Legrand-Blain & Nicollin, Reference Brice, Legrand-blain and Nicollin2005, MUA/1094/021: incomplete specimen in ventral and lateral views. (e–g) Tenuisinurostrum? sp., MUA/1094/022: crushed specimen in ventral, dorsal and lateral views. (h–l) Sphaeridiorhynchus sp., MUA/1094/023: complete specimen in ventral, dorsal, lateral, posterior and anterior views. Scale bars: 5 mm.
2005 Paurogastroderhynchus presaharensis: Brice in Brice et al., p. 15–16, pl. 2, figs 1–7, 25a–b.
Material. One incomplete specimen from Tamtert–Zereg section (Ze 2/8), Marhouma Formation, Member 3, Submember c.
Description. Shell large (33.6 mm in width), dorsibiconvex, wider than long, rounded subpentagonal in outline. Valves covered by simple, narrow and angular costae near beaks, becoming wider and subangular anteriorly and flattened on flanks; five median and eight ventral lateral costae (not observed on dorsal valve due to poor preservation); intercostal grooves narrower than costae. Ventral valve slightly convex in umbonal area; apical angle 112°; tongue not preserved; traces of short, divergent dental plates. Traces of dorsal median septum.
Remarks. This single specimen is assigned to Paurogastroderhynchus presaharensis on the basis of its external features (shape and ornamentation). This species was reported previously from the upper Famennian deposits of Algeria (Timimoun region; Sartenaer, Reference Sartenaer1975), Ahnet area (material from the Khenig Sandstones collected by M. Legrand-Blain; D. Brice, unpub. data) and of the Western Anti-Atlas in Morocco (Kheneg Lakahal section; Brice in Brice, Legrand-Blain & Nicollin, Reference Brice, Legrand-blain and Nicollin2005) where it is abundant. It is also represented by some remains from the upper Famennian Hassi Rharouar section, SE of Tinfouchy, and the Zemoul area (Morocco) (Brice in Brice, Legrand-Blain & Nicollin, Reference Brice, Legrand-blain and Nicollin2005). The specimen from Ze 2/8 of the Tamtert–Zereg section is of late Famennian age (upper Platyclymenia Zone).
Superfamily camarotoechioidea Schuchert, Reference Schuchert and Pompeckj1929
Family leiorhynchidae Stainbrook, Reference Stainbrook1945
Subfamily leiorhynchinae Stainbrook, Reference Stainbrook1945
Genus Tenuisinurostrum Sartenaer, Reference Sartenaer1967
Type species. Camarophoria crenulata Gosselet, Reference Gosselet1877, lower Famennian, southern Belgium.
Tenuisinurostrum? sp.
Figure 4e–g
Material. One incomplete, partly deformed specimen from Gara Diba, Marhouma Formation, Member 2, Submember b.
Description. Shell of medium size (L?, W = 19 mm, T = 8.2 mm), suboval in outline, weakly dorsibiconvex; lateral commissures acute; anterior commissure uniplicate; anterior margin flat. Shell with rounded, irregular ribs present in anterior region, devoid of lateral ribs, with a weak ventral median groove originating in the umbonal region. Ventral valve with suberect beak; sulcus arising at about mid-length, poorly defined, flat-bottomed at front, shallow (?); tongue very low; apical angle 125°; possible traces of a dental plate left of the beak. Dorsal valve with greatest convexity slightly posterior to mid-length; fold weakly developed, low, flat-topped at front, originating at about mid-length; trace of a short median septum.
Remarks. Although poorly preserved, this single specimen is tentatively assigned to Tenuisinurostrum mainly on the basis of its suboval outline, its irregular ribs and its low tongue. It displays a trace of a rudimentary dental plate, though such structures are not reported in this genus (see Sartenaer, Reference Sartenaer1967). Externally, it is closer to T. crenulatum rather than to T. subcrenulatum Biernat, Reference Biernat1970, which is more dorsibiconvex and wider than long. Paromoeopygma Sartenaer, Reference Sartenaer1968 (Pugnacoidea) possesses dental plates and displays a similar outline in ventral view but differs by its more inflated dorsal valve, its higher tongue and absence of a dorsal median septum. Further material is therefore required for confident identification.
The type species of Tenuisinurostrum was described from the lower Palmatolepis crepida Zone (Sartenaer, Reference Sartenaer1984) in southern Belgium and northern France and T. subcrenulatum Biernat, Reference Biernat1970 is known from the P. triangularis to P. marginifera zones in the Holy Cross Mountains of Poland (Biernat, Reference Biernat1970, Reference Biernat1983; Sartenaer, Reference Sartenaer1985), though the latter needs to be revaluated from the taxonomic viewpoint (Sartenaer, Reference Sartenaer1984, Reference Sartenaer1985, Reference Sartenaer1987). Our badly preserved specimen of Tenuisinurostrum? sp. is of early Famennian age (do II) on the basis of Palmatolepis minuta subtilis (P. triangularis to P. trachytera zones). Tenuisinurostrum is not known with certainty from North Africa (cf. Sartenaer, Reference Sartenaer1984, Reference Sartenaer1987).
Genus Sphaeridiorhynchus Sartenaer, Pushkin & Kotlyar, Reference Sartenaer, Pushkin and Kotlyar1997
Type species. Sphaeridiorhynchus kuzmichiensis Sartenaer, Pushkin & Kotlyar, Reference Sartenaer, Pushkin and Kotlyar1997, lower Famennian, Belarus.
Sphaeridiorhynchus sp.
Figure 4h–l
Material. Eight specimens including one juvenile from the Cheffar El Hamar (CA 1a) section, Marhouma Formation, Member 2, Submember a.
Description. Shell of medium size (up to 20.7 mm in width), wider than long, widest near mid-length, biconvex or weakly dorsibiconvex, suborbicular to subpentagonal in outline; anterior commissure uniplicate, anterior margin straight. Shell smooth, but two specimens display median ribs arising in the anterior region of the shell (mc = 0/3 and 5/?). Ventral valve inflated; beak erect; sulcus weak, arising in umbonal region or at about mid-length, flat-bottomed; tongue nearly perpendicular to commissure plane, low, trapezoidal. Dorsal valve strongly inflated with greatest convexity slightly posterior to mid-length; fold weak, arising in umbonal area, flat-topped at front; septum lacking.
Remarks. These specimens are assigned to Sphaeridiorhynchus on the basis of their markedly biconvex, smooth and medium-sized shell with suborbicular to subpentagonal outline, and by the weak fold and sulcus. They are clearly less globular than specimens of its type species, but their specific assignment requires additional material.
Until now, Sphaeridiorhynchus was known only from the lower Famennian succession of Belarus (Pripyat’ Depression) and Ukraine (Dnepr–Donets Depression) (Sartenaer, Pushkin & Kotlyar, Reference Sartenaer, Pushkin and Kotlyar1997), where it is represented by S. kuzmichiensis. Discovery of Sphaeridiorhynchus sp. in the lower Famennian succession of the Saoura Valley thus represents the first occurrence of this genus outside the Eastern European Platform.
Superfamily pugnacoidea Rzhonsnitskaia, Reference Rzhonsnitskaia and Thalmann1956
Family pugnacidae Rzhonsnitskaia, Reference Rzhonsnitskaia and Thalmann1956
Genus Evanidisinurostrum Sartenaer, Reference Sartenaer1987
Type species. Pseudoleiorhynchus? zemoulensis Drot, Reference Drot1964, lower Famennian, plains of Drah-el-Kelba, pre-Saharan Morocco.
Evanidisinurostrum saouraense sp. nov.
Figure 5a–t

Figure 5. Rhynchonellide brachiopods from the Marhouma Formation, Upper Devonian (Famennian), Saoura Valley, Algeria (see Fig. 1). (a–t) Evanidisinurostrum saouraense sp. nov. from the Marhouma Formation, Upper Devonian (Famennian), Saoura Valley, Algeria (see Fig. 1). (a–e) holotype, MUA/1094/016: complete specimen in ventral, dorsal, lateral, posterior and anterior views. (f–j) MUA/1094/018: complete specimen in ventral, dorsal, lateral, posterior and anterior views. (k–o) MUA/1094/017: complete specimen in ventral, dorsal, lateral, posterior and anterior views. (p–t) MUA/1094/19: complete juvenile specimen in ventral, dorsal, lateral, posterior and anterior views. (u–y) Hadyrhyncha cf. hadyensis Havlíček, Reference Havlíček1979, MUA/1094/024: complete specimen in ventral, dorsal, lateral, posterior and anterior views. (z–ii) Leptoterorhynchus sp.: (z–dd) MUA/1094/025: complete specimen in ventral, dorsal, lateral, posterior and anterior views; (ee–ii) MUA/1094/026: complete specimen in ventral, dorsal, lateral, posterior and anterior views. Scale bar: 5 mm.
Derivatio nominis. In reference to the Saoura Valley (Ougarta Basin, Algeria).
Holotype. A complete steinkern, MUA/1094/016.
Locus typicus. Gara Diba section (30°7′38″N; 2°12′30″W), Saoura Valley, eastern part of the Ougarta Basin, Algerian Sahara.
Stratum typicum. Marhouma Formation (lower Famennian), Member 2, Submember a.
Studied material. About 400 specimens (steinkerns), 125 well preserved (of which 42 were measured), 27 decorticated, 21 incomplete, 227 juveniles and numerous isolated valves and fragments from the Gara Diba section.
Diagnosis. A small species of Evanidisinurostrum (up to c. 17 mm in width) with shell subpentagonal in outline, wider than long, dorsibiconvex, widest near mid-length or more anteriorly. Fold and sulcus starting posterior to mid-length. Tongue moderately high, subtrapezoidal in outline, perpendicular or almost perpendicular to the commissural plane. Most of shells smooth with rounded median costae arising at or posterior to mid-length (mc = 2–3/1–2, usually 2/1); lateral costae usually absent or rare (0–1/0–2).
Description. Shell small (up to 17.2 mm in width), dorsibiconvex, wider than long, widest near mid-length or more anteriorly, subpentagonal in outline; anterior commissure uniplicate, anterior margin, in ventral view, indented by a strong median costa. Most shells are smooth on the flanks with rounded median costae arising at or posterior to mid-length (mc = 2–3/1–2, usually 2/1); lateral costae usually absent or rare (0–1/0–2). Ventral valve gently inflated in both posterior and lateral profiles with greatest convexity posterior to mid-length; beak small; apical angle between 110° and 120°; sulcus wide, deep, originating at about mid-valve, with prominent median costa; tongue wider than long, almost perpendicular to commissural plane, trapezoidal; dental plates absent; muscle field impressed, triangular in outline. Dorsal valve evenly convex in both lateral and posterior profiles, highest at front, impressed; fold originating at mid-length, relatively flat-topped at front (apart from costae); no median septum; myophragm short.
Dimensions in mm (n = 42 for all; see Appendix 1 in online supplementary data, available at http://journals.cambridge.org/geo). Width range 9.75–17.2, average 12.5; length range 8.75–13.55, average 10.6; thickness range 4.2–7.1, average 5.5; width of sulcus range 4–13.1, average 8.18; width/length ratio range 1.01–1.37, average 1.18; width/thickness ratio range 1.78–2.71, average 2.27; sulcus width/width ratio range 0.41–0.82, average 0.64.
Remarks. As all the specimens are steinkerns, it was not possible to investigate the interiors by serial sections. These specimens are assigned to Evanidisinurostrum rather than Perrarisinurostrum Sartenaer, Reference Sartenaer1984 and Tenuisinurostrum Sartenaer, Reference Sartenaer1967 (see Sartenaer, Reference Sartenaer1987 for discussion of the differences between these three genera) as they have similarities with E. zemoulense (Drot, Reference Drot1964) such as the outline, the similar development of fold and sulcus and the absence of dental plates and median septum. Nevertheless, the specimens from the Saoura Valley can be discriminated from Drot's (Reference Drot1964) species by their smaller size, their maximum width near the mid-length or anteriorly, their different W/L ratio, their larger apical angle and their different median ornament.
Evanidisinurostrum was previously known with certainty from the lower Famennian IIβ by its type species in the Zemoul and south of Akka in the Dra Plains of pre-Saharan Morocco (Drot, Reference Drot1964; Sartenaer, Reference Sartenaer1987) and on the southern margin of the Tindouf Basin (Algeria; Drot, Reference Drot1964). Furthermore, Sartenaer (Reference Sartenaer1987, p. 135) suggested that Tenuisinurostrum subcrenulatum Biernat, Reference Biernat1970 from the Holy Cross Mountains in Poland (middle Palmatolepis triangularis to lower P. marginifera zones) may belong to Evanidisinurostrum, but this needs confirmation.
Family rozmanariidae Havlíček, Reference Havlíček1982
Genus Hadyrhyncha Havlíček, Reference Havlíček1979
Type species. Hadyrhyncha hadyensis Havlíček, Reference Havlíček1979, upper Famennian, Moravia, Czech Republic.
Hadyrhyncha cf. hadyensis Havlíček, Reference Havlíček1979
Figure 5u–y
cf. 1979 Hadyrhyncha hadyensis: Havlíček, p. 99, pl. 2, figs 6–9, text-fig. 8.
Material. Two specimens, of which one is incomplete, from the Marhouma Formation, Member 3, Submember c, of the Tamtert–Zereg section (Ze 2/8, Ze 2/8b).
Description. Shell small (up to 15.7 mm in width), wider than long, widest near mid-length, transversely elliptic in outline, slightly ventribiconvex; anterior commissure unisulcate; anterior margin straight. Low, rounded median costae originating near umbones (mc = 3/4), the central one bifurcating both in the sulcus and on the fold; lateral costae badly preserved (lc = 6/5–8), corresponding to the division of 2 or 3 costae present in the posterior region. Ventral valve with greatest convexity near mid-length; beak small, erect; fold low, flat-topped at front, arising in posterior part of the valve; tongue low, subtrapezoidal, perpendicular to commissural plane. Dorsal valve with greatest convexity posterior to mid-length; sulcus arising near umbo, well-defined, flat-bottomed at front.
Remarks. These specimens share numerous external similarities with Hadyrhyncha hadyensis, such as their transversally elliptic outline, the similar development of their dorsal sulcus and ventral fold, their sharp commissures and their ornament. However, as the internal morphology of the Algerian material remains unknown, they are tentatively compared to Havlíček's (Reference Havlíček1979) species. They differ from Hadyrhyncha meridionalis Sartenaer, Reference Sartenaer1998a from Morocco by their smaller size, their narrower apical angle, their ornament and their different L/W, T/W and T/L ratios. They are separated from Hadyrhyncha sp. from Poland (Halamski & Baliński, Reference Halamski and Baliński2009) by their smaller size and less transverse outline.
Our specimens are associated with ammonoids of the upper Famennian Platyclymenia Zone (do IV or IV/V). Hadyrhyncha hadyensis is known from the upper or uppermost Famennian (do V or do VI according to Havlíček, Reference Havlíček1979) of Moravia (Czech Republic) and from the middle part of do V in south-easternThuringia (Germany) (Bartzsch & Weyer, Reference Bartzsch and Weyer1986). H. meridionalis Sartenaer, Reference Sartenaer1998a occurs in the upper Famennian (do V) of southern Morocco (Dra Valley, Maider, Tafilalt, Zemoul); the genus is also reported from Poland (Holy Cross Mountains) in the upper Palmatolepis marginifera to upper Siphonodella praesulcata zones (Halamski & Baliński, Reference Halamski and Baliński2009).
Genus Leptoterorhynchus Sartenaer, Reference Sartenaer1998b
Type species. Rozmanaria magna Biernat & Racki, Reference Biernat and Racki1986, middle Famennian, Holy Cross Mountains, Poland.
Leptoterorhynchus sp.
Figure 5z–ii
Material. Three specimens from the Marhouma Formation, Member 3, Submember c of the Tamtert–Zereg (Ze 2/8) section and two from the Marhouma Formation, Member 2, Submember b of the Cheffar El Ahmar (CA 1c, CA2) section.
Description. Shell small (up to 13.1 mm in width), smooth, ventribiconvex, wider than long, widest about mid-length and transversally elliptical in outline; anterior commissure unisulcate; anterior margin straight. Ventral valve with greatest convexity near mid-length; beak small, erect; fold poorly developed, low, round-topped at front; tongue high, bent ventrally. Dorsal valve with greatest convexity near valve mid-length; sulcus wide, originating in posterior part of the valve, shallow, flat-bottomed at front; tongue low, rounded, perpendicular to commissural plane or bent dorsally; internal features not observed; fold poorly developed, low, round-topped at front; no median septum.
Remarks. These specimens are assigned to Leptoterorhynchus as they share many external similarities with the type species L. magnus (Biernat & Racki, Reference Biernat and Racki1986) such as the outline, the slightly developed and smooth sulcus and fold, and the sharp commissure. They probably belong to a new species which is not named because of insufficient material. However, they can be discriminated from the type species by their smaller size, their narrower apical angle, their different W/L, W/T and T/L ratios, their not-so-well-defined sulcus and fold and the absence of a median depression on the fold.
Leptoterorhynchus magnus (Biernat & Racki, Reference Biernat and Racki1986) is known only from the middle Famennian deposits of the southern Holy Cross Mountains, Poland according to Halamski & Baliński (Reference Halamski and Baliński2009). An unidentified species was reported in the Famennian (UD IIIC–IVB interval) of north-western Sauerland in Germany by Sartenaer (Reference Sartenaer1998b ). In the Saoura Valley, Leptoterorhynchus sp. is known from the upper Famennian Tamtert–Zereg section on the basis of Platyclymenia annulata and from the middle Famennian (on the basis of Maenoceras) Cheffar El Ahmar section.
Genus Novaplatirostrum Sartenaer, Reference Sartenaer1997
Type species. Novaplatirostrum sauerlandense Sartenaer, Reference Sartenaer1997, upper Famennian, north-western Sauerland, Germany.
Novaplatirostrum sp.
Figure 6a–j

Figure 6. Rhynchonellide brachiopods from the Marhouma Formation, Upper Devonian (Famennian), Saoura Valley, Algeria (see Fig. 1). (a–j) Novaplatirostrum sp.: (a–e) MUA/1024/027: complete specimen in ventral, dorsal, lateral, posterior and anterior views; (f–j) MUA/1094/028: complete specimen in ventral, dorsal, lateral, posterior and anterior views. (k–o) Pugnaria sp., MUA/1094/029: complete specimen in ventral, dorsal, lateral, posterior and anterior views. (p–x) Phacoiderhynchus aff. antiatlasicus Sartenaer, Reference Sartenaer2000: (p–s) MUA/1094/030: almost complete specimen in ventral, dorsal, lateral and posterior views; (t–x) MUA/1094/031: incomplete specimen in ventral, dorsal, lateral, posterior and anterior views. Scale bars: 5 mm.
Material. Five specimens from the Tamtert–Zereg section (Ze 2/8, 2/8b, 2/9) and one from “Plat” between Ouarourout and Gara Diba; Marhouma Formation, Member 3, Submember c.
Description. Shell small (up to 16.2 mm in width), flatly biconvex, suborbicular in outline; anterior commissure uniplicate; anterior margin slightly rounded, with sharp commissures (juveniles uniplicate, flattly biconvex, transversely subelliptical in outline). Low rounded median costae (3/2 or 4/3) near anterior commissure; no lateral costae. Ventral valve with greatest convexity in umbonal area; sulcus absent or wide (Ws/W: 0.68; n = 1), very shallow, starting from mid-length, flat-bottomed; tongue low, not perpendicular to commissural plane; maximal apical angle of 127°. Dorsal valve with greatest convexity posterior to mid-valve; fold obsolete, flat-bottomed.
Remarks. The Algerian specimens are externally close to Novaplatirostrum sauerlandense Sartenaer, Reference Sartenaer1997, especially paratypes F and G illustrated by Sartenaer (Reference Sartenaer1997, pl. 1, figs 31–40) but they are smaller and flatter than the latter and devoid of lateral costae. Our specimens display less median costae, have an obsolete fold and a smaller apical angle.
In the Saoura Valley, Novaplatirostrum sp. is known from the Tamtert–Zereg section where it is associated with goniatites and clymeniids of late Famennian age in levels Ze 2/8 and Ze 2/8b (upper Platyclymenia Zone) and in level Ze 2/9 (Gonioclymenia Zone). The age of the latter could be younger on the basis of a conodont fauna characteristic of the early upper Palmatolepis expansa Zone. N. sauerlandense occurs in deep-water environments in the German upper Famennian deposits (southern Sauerland and south-eastern Thuringia; Sartenaer, Reference Sartenaer1997) where it is associated with goniatites, but also in the Holy Cross Mountains in Poland in levels of probable do V–VI age (Halamski & Baliński, Reference Halamski and Baliński2009; A. Halamski, pers. comm., 2014).
Genus Pugnaria Biernat & Racki, Reference Biernat and Racki1986
Type species. Pugnaria plana Biernat & Racki, Reference Biernat and Racki1986, middle Famennian, Holy Cross Mountains, Poland.
Pugnaria sp.
Figure 6h–o
Material. Three specimens (one incomplete adult and two juveniles) from the Tamtert–Zereg (Ze 2/8) section (Marhouma Formation, Member 3, Submember c) and one adult from the Ouarourout (Ou 1/15) section (Marhouma Formation, Member 4, Submember a).
Description. Shell small (19.2 mm in width), wider than long, flatly dorsibiconvex, transversely elliptical in outline; anterior commissure rectimarginate; anterior margin slightly rounded. Shell smooth, growth lines thin, irregularly distributed, observed on ventral valve. Ventral valve with greatest convexity posterior to mid-length; beak small, curved; sulcus lacking. Dorsal valve with greatest convexity posterior to mid-length; fold lacking.
Remarks. Despite lack of knowledge about the internal features, our specimens are externally very close to Pugnaria plana Biernat & Racki, Reference Biernat and Racki1986 but are smaller, devoid of a fold and display a wider apical angle. Additional material is required for accurate specific identification.
Pugnaria sp. occurs in do III–do IV in the Tamtert–Zereg section (Ze 2/8) and is also known from the Ouarourout section (Ou1/15) where it is associated with ammonoids of probable do V age. In the Kowala section (Holy Cross Mountains, Poland), the range of P. plana in terms of conodont zones spans the Palmatolepis marginifera to Siphonodella praesulcata zones (Halamski & Baliński, Reference Halamski and Baliński2009).
Genus Phacoiderhynchus Sartenaer, Reference Sartenaer2000
Type species. Phacoiderhynchus antiatlasicus Sartenaer, Reference Sartenaer2000, middle Famennian, Maider, Morocco.
Phacoiderhynchus aff. antiatlasicus Sartenaer, Reference Sartenaer2000
Figure 6p–x
aff. 2000 Phacoiderhynchus antiatlasicus n. gen., n. sp.: Sartenaer, p. 79–84, pl. 1, figs 1–30, pl. 2, figs 31–65.
Material. Eleven incomplete specimens from the Tamtert–Zereg section (Ze 2/7: one adult and one juvenile; Ze 2/8: six adults, one juvenile; Ze 2/8b: two adults), Marhouma Formation, Member 3, Submember c.
Description. Shell of medium-size (up to 29 mm in width), wider than long, flatly biconvex, transversally elliptical in outline, widest at mid-length; commissure sharp; anterior commissure uniplicate; anterior margin flattened. Shells mostly smooth with rounded and wide median costae sometimes divided near the frontal margin (mc: 3–4/2–4); lateral costae not preserved. Ventral valve with greatest convexity close to mid-length; beak very small; sulcus shallow, flat-bottomed at front; tongue very low, not perpendicular to commissural plane. Dorsal valve with maximum convexity near mid-valve; fold scarcely developed, originating anterior to mid-length.
Remarks. These incomplete specimens are tentatively assigned to Phacoiderhynchus antiatlasicus Sartenaer, Reference Sartenaer2000 on the basis of their external morphology, although they can be distinguished by their smaller size, their sulcus and fold starting a little more anteriorly and their less numerous median costae (the lateral ones are absent or not preserved). The specimens from the Saoura Valley have a narrower apical angle (134–145° versus 147–157°) and may represent a new species, but specific assignment requires additional material.
Phacoiderhynchus aff. antiatlasicus is known from the upper Famennian Tamtert–Zereg section on the basis of ammonoids (upper Platyclymenia Zone). P. antiatlasicus occurs with certainty in middle–upper Famennian IIIB to IVB deposits from southern Morocco (Maïder and eastern Draa Plains) according to Sartenaer (Reference Sartenaer2000), and probably in the Immouzer du Kandar area south of Fès (central Morocco; see Brice et al. Reference Brice, Charriere, Drot and Regnault1984).
4. Discussion
4.a. Stratigraphic summary and general comments on brachiopod diversity during Famennian time
The stratigraphic range of the rhynchonellides within the Famennian part of the Marhouma Formation across the Saoura Valley (Ouarourout, Gara Diba, Cheffar Amar and Tamtert–Zereg sections) is presented in Figures 2 and 3.
As the Frasnian–Famennian (F–F) boundary, and thus the position of the Upper Kellwasser Event (Palmatolepis linguiformis Zone), is not precisely established within the Marhouma Formation, it is difficult to discuss in detail the post-Kellwasser recovery of the brachiopods in this part of the Ougarta Basin. Note that in regions where environmental conditions were more favourable to brachiopods, such as the Namur–Dinant Basin in southern Belgium and the Polish epicontinental basin, their recovery started in the Palmatolepis triangularis Zone very soon after the F–F boundary, in which athyridides, rhynchonellides and spiriferides played a significant role (Baliński Reference Baliński, Copper and Jin1996, Reference Baliński2002; Mottequin Reference Mottequin2008a , Reference Mottequin b ). On a worldwide scale, brachiopod diversity declined significantly during Frasnian time with a significant reduction in generic diversity; this was accentuated during Famennian time, possibly due to increasing faunal cosmopolitanism related to major sea-level changes and other parameters that suppressed faunal barriers and allowed interchange between remote areas (e.g. Copper, Reference Copper1998; Brice et al. Reference Brice, Carls, Cocks, Copper, García-Alcade, Godefroid and Rachebœuf2000). From the rhynchonellide viewpoint the Famennian Stage is marked by great generic diversity, contrasting with other brachiopod orders (e.g. orthides); this is similar to the situation reported during Emsian time (Curry & Brunton, Reference Curry, Brunton and Kaesler2007) and the occurrence of numerous cosmopolitan genera, including some of those identified in the Marhouma Formation (see discussion in Section 4.c). This Devonian stage is also characterized by the predominance of the rhynchonellides within brachiopod faunas (Curry & Brunton, Reference Curry, Brunton and Kaesler2007).
4.b. Palaeoecology and taphonomy
Brachiopods obtained from several horizons of the Famennian part of the Marhouma Formation include, in order of abundance, rhynchonellides (439 specimens), orthides (small Aulacella; 45 specimens), athyridides (medium-size Composita; 4 specimens) and spiriferides (only one small specimen; affinities not apparent due to poor preservation). Rhynchonellides therefore represent 90% of the whole brachiopod assemblage, with 10 species. For comparison, 197 ammonoids (goniatites and clymeniids; D. Korn and F.Z. Malti, unpub. data) and 19 trilobites (Crônier et al. Reference Crônier, Malti, François, Benyoucef and Brice2013), including the material from two other sections of the Saoura Valley (Béchir and Idhir; see location in Crônier et al. Reference Crônier, Malti, François, Benyoucef and Brice2013), have been collected in the same levels of the Marhouma Formation. Within these lithostratigraphic units, the greatest rhynchonellide diversity is at the top of Submember c of Member 3, which corresponds to the top of the early upper P. expansa Zone, i.e. at the base of the upper Famennian deposits (Fig. 2). Development of the ‘griotte’ limestones took place during a sea-level rise (F.Z. Malti, unpub. data) that probably facilitated faunal exchanges between distant basins.
The low-diversity brachiopod fauna is clearly an in situ rhynchonellide-dominated brachiopod community with much lower diversity of other orders; the specimens are not disarticulated, sorted or broken. No strophomenides or productides occurred in association with the rhynchonellides. Deep-water conditions are indicated by cephalopods (goniatites and clymeniids) and trilobites; the latter comprise blind to reduced-eyed phacopides (Crônier et al. Reference Crônier, Malti, François, Benyoucef and Brice2013). One of the most striking features of this rhynchonellide fauna is the predominance of smooth, thin-shelled and small forms (except some scarce large Trigonirhynchiidae), also characterized by flattened shells. Moreover, among these rhynchonellides, leiorhynchids (Camarotoechioidea) and rozmanariids (Pugnacoidea) are by far the most abundant elements as is generally the case in late Palaeozoic dysaerobic communities, though the Pugnacoidea are represented by other families (cf. Bowen, Rhoads & McAlester, Reference Bowen, Rhoads and McAlester1974; Biernat & Racki, Reference Biernat and Racki1986; Racki, Reference Racki1989; Alexander, Reference Alexander1994; Mottequin & Legrand-Blain, Reference Mottequin and Legrand-Blain2010). We also draw attention to the occurrence of a monospecific assemblage with Evanidisinurostrum saouraense in which immature specimens are particularly abundant. Almost monospecific rhynchonellide assemblages of leiorhynchids have been reported elsewhere, notably in the Frasnian deposits of southern Belgium (e.g. Sartenaer, Reference Sartenaer1974) and Poland (Racki, Reference Racki1993; Racki & Szulczewski in Sartenaer, Racki & Szulczewski, Reference Sartenaer, Racki and Szulczewski1998). Overrepresentation of juveniles of E. saouraense is probably related to the combination of several biological (e.g. nutrient availability) and physicochemical controlling factors (including water oxygenation and depth) (see Pérez-Huerta & Sheldon, Reference Pérez-Huerta and Sheldon2006) that induced significant losses among immature specimens.
The state of preservation of our material does not permit precision as regards the size of the foramen for each species, but adults of the identified genera are characterized by a reduced foramen (cf. data in Savage et al. Reference Savage, Manceñido, Owen, Carlson, Grant, Dagys, Sun and Kaesler2002). We are therefore inclined to regard the rhynchonellides as lying unattached on the sea floor. Another significant feature is the aequibiconvexity of the shells, notably for Pugnaria, Novaplatisinurostrum and Phacoiderhynchus. According to Walker (Reference Walker and Ziegler1974) this is indicative of soft substrates; it was interpreted in this way by Biernat & Racki (Reference Biernat and Racki1986) for Famennian rhynchonellides. Relatively more biconvex shells such as those of Sphaeridiorhynchus may accord with a firmer substrate (see Racki & Szulczewski in Sartenaer, Racki & Szulczewski, Reference Sartenaer, Racki and Szulczewski1998). According to Racki (Reference Racki1998), the deep-water rhynchonellide biofacies displays apparent continuity in spite of generic losses across the Frasnian–Famennian boundary. One of the major changes in this biofacies is the huge development of the rozmanariids during Famennian time. They made their appearance during Pragian time (cf. Savage et al. Reference Savage, Manceñido, Owen, Carlson, Grant, Dagys, Sun and Kaesler2002; Savage, Reference Savage and Kaesler2007), before disappearing at the end of the Devonian period (Mottequin, Brice & Legrand-Blain, Reference Mottequin, Brice and Legrand-Blain2014).
4.c. Comparison with other rhynchonellide faunas from North Africa and biogeographic affinities
Despite the increased number of publications, Famennian brachiopod assemblages from North Africa, and therefore from the northern margin of Gondwana, are still poorly documented. This is due in part to most papers being generally focused on a particular group, as with this paper. Taxonomic data are available for most of the important brachiopod orders present in the Famennian deposits of Algeria and Morocco (see below for Libyan data): productides (e.g. Brousmiche, Reference Brousmiche1975; Legrand-Blain, Reference Legrand-Blain1995a , Reference Legrand-Blain, Arbey and Lorenz b ; Legrand-Blain in Brice, Legrand-Blain & Nicollin, Reference Brice, Legrand-blain and Nicollin2005), spiriferides (e.g. Drot, Reference Drot1964; Nicollin & Brice, Reference Nicollin and Brice2000; Brice & Nicollin in Brice, Legrand-Blain & Nicollin, Reference Brice, Legrand-blain and Nicollin2005), and rhynchonellides.
Five rhynchonellide genera, namely Centrorhynchus, Evanidisinurostrum, Hadyrhyncha, Paurogastroderhynchus and Phacoiderhynchus, occurring within the Marhouma Formation were previously reported from Famennian successions on the northern margin of Gondwana (Morocco and Algeria). Attention is drawn to two salient absences from the rhynchonellides of the Marhuma Formation: (1) Tetragonorhynchus Sartenaer, Reference Sartenaer1999, reported from the upper Famennian (UD V) deep-water facies from southern Morocco (Maïder) (Sartenaer, Reference Sartenaer1999); and (2) Dzieduszyckia Siemiradzki, Reference Siemiradzki1909 represented by two species in the Famennian deposits of the Middle Atlas of Morocco (Baliński & Biernat, Reference Baliński and Biernat2003). Dzieduszyckia has been considered to be typical of Upper Devonian vent-seep environments (Campbell & Bottjer, Reference Campbell and Bottjer1995), but Baliński & Biernat (Reference Baliński and Biernat2003) have challenged this judgement on the basis of geochemical analyses of rocks associated with these Moroccan occurrences, insisting that no particular environmental conditions can be discerned.
Famennian brachiopods from western Libya were described by Havlíček (Reference Havlíček, Seidl and Röhlich1984), Havlíček & Röhlich (Reference Havlíček and Röhlich1987), and Mergl & Massa (Reference Mergl and Massa1992). They came from shallow-water environments characterized by siliciclastic deposits with benthic communities dominated by lingulide and rhynchonellide brachiopods (Mergl & Massa, Reference Mergl, Massa, Scola and Worsley2000). The Libyan rhynchonellides include species of Cupularostrum Sartenaer, Reference Sartenaer1961 and Libyaerhynchus Mergl & Massa, Reference Mergl and Massa1992 that are coarsely ribbed, and thus markedly different from those occurring in contemporaneous deep-water environments of Morocco and Algeria. As rightly reported by Mergl & Massa (Reference Mergl and Massa1992) and Mergl, Massa & Plauchut (Reference Mergl, Massa and Plauchut2001), the stratigraphic range of Cupularostrum, originally suggested as Givetian, should be reconsidered.
Four genera known from deep-water environments are recognized for the first time in North Africa: Pugnaria, Leptoterorhynchus, Sphaeridiorhynchus and Novaplatirostrum. They were previously known only from the south-eastern margin of Laurussia and were viewed as being restricted geographically: Leptoterorhynchus (Germany and Poland; Sartenaer, Reference Sartenaer1998b ), Novaplatirostrum (Germany and Poland; Sartenaer, Reference Sartenaer1997; Halamski & Baliński, Reference Halamski and Baliński2009), Pugnaria (Poland; Biernat & Racki, Reference Biernat and Racki1986), and Sphaeridiorhynchus (Belarus and Ukraine; Sartenaer, Pushkin & Kotlyar, Reference Sartenaer, Pushkin and Kotlyar1997). Tenuisinurostrum, previously reported from western and possibly eastern Europe (cf. Sartenaer, Reference Sartenaer1987), is here reported from the Famennian Marhouma Formation, but needs confirmation on the basis of well-preserved material.
Halamski & Baliński (Reference Halamski and Baliński2013) recently stressed the great similarities between Middle Devonian benthic faunas from the northern and southern shores of the Variscan Sea, which accords with a narrow oceanic zone between Laurussia and Gondwana (McKerrow et al. Reference McKerrow, Mac Niocaill, Ahlberg, Clayton, Cleal, Eagar, Franke, Haak, Oncken and Tanner2000) rather than an extensive Rheic Ocean (e.g. Torsvik & Cocks, Reference Torsvik, Cocks, Van Hinsbergen, Buiter, Torsvik, Gaina and Webb2011, Reference Torsvik and Cocks2013; Stampfli et al. Reference Stampfli, Hochard, Vérard, Wilhem and von Raumer2013). Legrand-Blain (Reference Legrand-Blain1995a ) also noted clear affinities between the productides, rhynchonellides and spiriferides of North Africa and Southern Europe with those from Armenia and the Urals across the Devonian–Carboniferous boundary. Close affinities between the Famennian rhynchonellides from central Algeria and those from western and eastern Europe is consistent with the hypothesis of prolonged proximity between both continental masses, but island arcs between Laurussia and Gondwana (Stampfli et al. Reference Stampfli, Hochard, Vérard, Wilhem and von Raumer2013; Torsvik & Cocks, Reference Torsvik and Cocks2013) could have facilitated dissemination and interaction of the brachiopod faunas on both sides of the Rheic Ocean.
5. Conclusions
In the Saoura Valley (Ougarta Basin, Algeria), the Famennian part of the Marhouma Formation (members 2–4) contains rhynchonellide-dominated brachiopod assemblages in which a generally low diversity of small athyridides (Composita), orthides (Aulacella) and spiriferides occur. The rhynchonellides, leiorhynchids and rozmanariids are the most abundant and the most diverse elements, represented by small, thin-shelled species. These brachiopods are generally found in association with cephalopods and blind or eye-reduced phacopide trilobites (Crônier et al. Reference Crônier, Malti, François, Benyoucef and Brice2013), interpreted to have lived in deep, oxygen-depleted environments. Such rhynchonellide assemblages are interpreted as being characteristic of dysaerobic environments found on epicontinental platforms during Late Devonian time, especially in Laurussia and North Gondwana. The leiorhynchid Sphaeridiorhynchus and the rozmanariids Leptoterorhynchus, Pugnaria and Novaplatirostrum are reported for the first time outside Europe; their occurrence on the margin of Gondwana reinforces the biostratigraphic value of these four genera and underscores the global character of the Famennian benthic faunas.
Acknowledgements
We are greatly indebted to Dieter Korn for having provided identifications of the cephalopods cited in the text, to Thierry Hubin for technical assistance and to Adam T. Halamski, Andrzej Baliński and John Talent for reviewing the manuscript. This paper is a contribution to the International Geoscience Programme (IGCP) Project 596, Climate change and biodiversity patterns in the Mid Palaeozoic.
Supplementary material
To view supplementary material for this article, please visit http://dx.doi.org/10.1017/S0016756814000697








