INTRODUCTION
Aoridae (Crustacea: Amphipoda) are distributed all over the world from shallow estuarine to open marine habitats (Myers, Reference Myers2009) and are grouped into 26 genera. In European waters, Aoridae are represented by nine genera (Horton & De Broyer, Reference Horton, De Broyer, Horton, Lowry, De Broyer, Costello, Bouchet, Boxshall, Arvanitidis and Appeltans2015): Aora Krøyer, 1845, Arctolembos Myers, 1979, Autonoe Bruzelius, 1859, Bemlos Shoemaker, 1935, Chevreuxius Bonnier, 1896, Grandidierella Coutière, 1904, Lembos Bate, 1857, Microdeutopus Costa, 1853, Tethylembos Myers, 1988. The genus Aoroides contains 18 valid species (Lowry, Reference Lowry, Horton, Lowry and De Broyer2015) and is distributed only in the coastal regions of the Pacific Ocean (Barnard & Karaman, Reference Barnard and Karaman1991; Ariyama, Reference Ariyama2004; Myers, Reference Myers2005, Reference Myers2009). This paper reports on the first records of the genus Aoroides in European marine waters with the presence of three Asian species on the French Atlantic coasts: A. longimerus Ren & Zheng, 1996, A. semicurvatus Ariyama, Reference Ariyama2004 and A. curvipes Ariyama, Reference Ariyama2004. An identification key is provided for the three European species of the genus Aoroides.
MATERIALS AND METHODS
Study areas
The Bay of Brest (NW Brittany) is a 180 km2 semi-enclosed shallow basin (Figure 1A) connected to shelf waters (Iroise Sea) by a narrow (1.8 km wide) and deep (40 m) channel. Owing to strong tidal influence, the water masses are well-mixed (Le Pape et al., Reference Le Pape, Del Amo, Menesguen, Aminot, Queguiner and Treguer1996). In this bay, the main freshwater inputs (80%), due to the Elorn River in the northern part and to the Aulne River in the southern part, can influence salinity of surface seawater in localized areas. According to local hydrodynamic conditions, the Bay of Brest encompasses a mosaic of habitats from mud flats to coarse gravels and rocky substrata (Castric-Fey & Chassé, Reference Castric-Fey and Chassé1991; Grall & Glémarec, Reference Grall and Glémarec1997). Important cupped oyster (Crassostrea gigas (Thunberg, 1793)) farms (110 ha) are located in the south-eastern part of the bay. Brest is one of the main hotspots of military, commercial and recreational shipping on the Atlantic coast of France. A total of 2250 berths are available, distributed between the marinas of Moulin-Blanc in the eastern part and Château in the western part (Figure 1A).

Fig. 1. Sampling stations of Aoroides longimerus Ren & Zheng, 1996 (black stars), Aoroides semicurvatus Ariyama, Reference Ariyama2004 (black crosses) and both A. semicurvatus and Aoroides curvipes Ariyama, Reference Ariyama2004 (black dots) in: (A) Arcachon Bay; (B) Bay of Brest; (C) Hossegor Lake (see code legend in Table 1).
Arcachon Bay is a 180 km2 macrotidal coastal lagoon situated on the south-western coast of France (Figure 1B). This lagoon communicates with the Atlantic Ocean by a narrow channel and receives freshwater inputs in its south-eastern part (Leyre River). It is characterized by large intertidal flats (115 km2), whose lower parts are used for cupped oyster farming (720 ha). In the inner lagoon, tidal channels represent 71 km2, with 1.02 km2 occupied by eelgrass beds (Zostera marina Linnaeus, 1753) (Plus et al., Reference Plus, Dalloyau, Trut, Auby, de Montaudouin, Emery, Noel and Viala2010). Arcachon Bay is surrounded by marinas and numerous seasonal moorings representing a total accommodation capacity of more than 7800 leisure boats (Alzieu, Reference Alzieu2000).
Hossegor Lake (Figure 1C) is a small coastal lagoon (0.9 km2) communicating with the Atlantic Ocean by a very narrow channel and receiving fresh water by two small rivers (Bourret and Boudigau Rivers) in its southern part. Hossegor Lake is also connected to Capbreton harbour. Intertidal flats represent 0.28 km2 (Trut et al., Reference Trut, Dalloyau and Auby2009), with the south-eastern part devoted to oyster farming.
Material examined
Specimens of Aoroides were collected from different stations and habitats in the Bay of Brest, Arcachon Bay and Hossegor Lake from 2009 to 2014 (Figure 1, Table 1). In Arcachon Bay and Hossegor Lake, specimens were sampled during specific biodiversity surveys. In the Bay of Brest, the specimens were incidentally sampled as part of a study aiming at examining colonization patterns on hard substrates in three marinas in Brittany. Specimens were fixed in 4% formalin and some individuals stained with methyl green for further morphological examinations. Specimens were observed under a Nikon SMZ25 stereomicroscope and a Nikon Eclipse E400 microscope, and photographed with a Nikon DS-Ri2 camera. Body length (BL) was measured with NIS-Elements Analysis software from anterior margin of head to posterior end of telson. Drawings were carried out from pictures using Inkscape software. Specimens examined in the scope of this study were deposited in collections of the Muséum National d'Histoire Naturelle, Paris (Table 1) and of Arcachon and Brest Marine Stations.
Table 1. Records of the Aoroides species in Bay of Brest, Arcachon Bay and Hossegor Lake. Sites and stations: see Figure 1.

N, number of males collected.
a For floating pontoons, specimens were found on substrates at a constant water depth of 1 m.
RESULTS
SYSTEMATICS
Order AMPHIPODA Latreille, 1816
Suborder SENTICAUDATA Lowry & Myers, 2013
Infraorder Corophiida Leach, 1814
Parvorder Corophiidira Leach, 1814
Superfamily Aoroidea Stebbing, 1899
Family Aoridae Stebbing, 1899
Genus Aoroides Walker, 1898
Type species: Aoroides columbiae Walker, 1898
Specimens were identified as being members of the genus Aoroides, a genus not previously reported in European waters. Three species of Aoroides were sampled in three different sites between 2009 and 2014: Aoroides longimerus (Figure 2A) in Bay of Brest and Arcachon Bay, Aoroides semicurvatus (Figure 2B) in Arcachon Bay and Hossegor Lake and Aoroides curvipes (Figure 2C) in Arcachon Bay.

Fig. 2. Lateral views of (A) Aoroides longimerus Ren & Zheng, 1996: specimen from Brest, W France; (B) Aoroides semicurvatus Ariyama, Reference Ariyama2004: specimen from Arcachon Bay, SW France; (C) Aoroides curvipes Ariyama, Reference Ariyama2004: specimen from Arcachon Bay, SW France. Scale bars: 0.5 mm.
GENERIC DIAGNOSIS
According to Barnard & Karaman (Reference Barnard and Karaman1991), the genus Aoroides is characterized by: article 3 of antenna 1 shorter than article 1; accessory flagellum absent; mandibular palp article 3 rectolinear (except for A. palfreyensis Myers, Reference Myers2009 and A. vitiosus Myers, Reference Myers1995 without mandibular palp); gnathopod 1 grossly merochelate; gnathopod 2 not carpochelate; uropod 3 with 2 rami.
As the individuals of the family Aoridae are easily broken, specimens of Aoroides might be confused with Aora Krøyer, 1845, a common aorid genus in European waters. However, Aoroides can be easily separated from Aora by (1) the absence of the accessory flagellum and (2) the mandibular palp article 3 rectolinear (Barnard & Karaman, Reference Barnard and Karaman1991) (Figure 3).

Fig. 3. Comparison of antenna 1 and mandibular palp in the genera Aora and Aoroides. (A) Antenna 1, lateral view of Aoroides longimerus Ren & Zheng, 1996; specimen from Bay of Brest, W France; (B) Antenna 1, lateral view of Aora gracilis with black arrow showing the accessory flagellum; specimen from Arcachon Bay, SW France; (C) Left mandibular palp, posterior view of Aoroides longimerus; specimen from Bay of Brest; (D) Left mandibular palp, posterior view of Aora gracilis; specimen from Arcachon Bay. Scale bars: (A), (B): 0.2 mm; (C), (D) 0.1 mm.
Aoroides longimerus Ren & Zheng, 1996
MATERIAL EXAMINED
North-east Atlantic Ocean: France: Arcachon Bay, Bay of Brest.
DESCRIPTION
Based on male, BL = 4.4 mm, Château marina (Brest), August 2014.
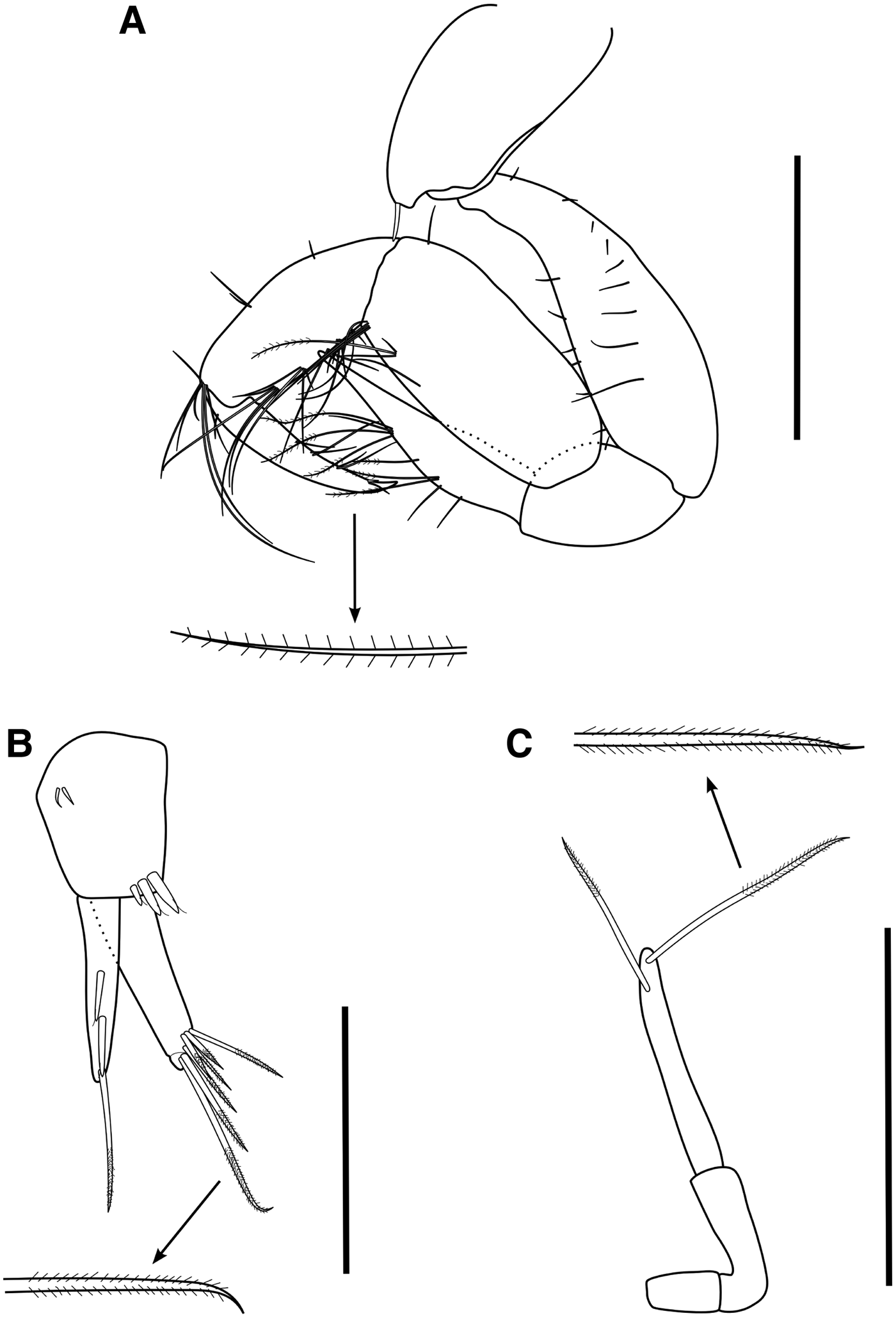
Antenna 1 accessory flagellum absent; mandibular palp article 3 with 2 distal setae (Figure 4D); coxa 1 with several plumose setae and one robust seta anteriorly; gnathopod 1 basis and carpus with many plumose setae (Figure 4A, B); palm of gnathopod 2 transverse; anterior margin of basis pereopods 3 and 4 with several simple setae; uropod 3 peduncle with 2 proximal and 4 distal sensory cuspidate setae (sensu Garm (Reference Garm2004): cuspidate setae with a sensory seta at its distal end), inner ramus with 3 dorsal sensory cuspidate setae, outer ramus with 2 dorsal and 1 distal sensory cuspidate setae (Figure 4C).

Fig. 4. Aoroides longimerus Ren & Zheng, 1996: specimen from Bay of Brest, W France (Body length = 4.4 mm): (A) left gnathopod 1, lateral view; (B) left gnathopod 1, setae of carpus and merus not all drawn, lateral view; (C) left uropod 3, dorsal view; (D) left mandibular palp, posterior view. Scale bars: (A), (B) 0.5 mm; (C), (D) 0.1 mm.
COLOURATION
Dorsal part of head, anterior and posterior margins of pereonite 1, posterior margins of pereonites 2–5 and 7 (sometimes 6), pleonites 1–3, uropodite 1, ventral parts of pleonites 1–3, ventral surface of coxa 1 brown or with dense brown dots (Figure 2A).
VARIABILITY
According to Ariyama's (Reference Ariyama2004) description, uropod 3 rami show a great variability with 1 to 3 dorsal sensory cuspidate setae on inner ramus and 1 to 3 dorsal sensory cuspidate setae on outer ramus.
REMARKS
Setae on distal part of uropod 3 rami and mandibular palp with short setules (difficult to see); this information is lacking for Japanese specimens (Ariyama, Reference Ariyama2004).
HABITAT
This species occurs in different types of sublittoral habitats, frequently associated with biogenic structures, living among seaweeds and on the surface of sessile animals (ascidians, hydroids, bryozoans and sponges). Rarely, A. longimerus could be found under stones in intertidal areas (Ariyama, Reference Ariyama2004). In this study, specimens were sampled on slipper limpet beds, oyster reefs and Zostera marina meadows (Arcachon) or associated with fouling communities dominated by mussels, ascidians and bryozoans under pontoons (Brest).
DISTRIBUTION
Japan, China (Ariyama, Reference Ariyama2004), France: Bay of Brest, Arcachon Bay (this study).
Aoroides semicurvatus Ariyama, Reference Ariyama2004
MATERIAL EXAMINED
North-east Atlantic Ocean: France: Arcachon Bay, Hossegor Lake.
DESCRIPTION
Based on male, BL = 3.9 mm, Les Hosses (Arcachon), October 2009.
Antenna 1 accessory flagellum absent; mandibular palp article 3 with 2 distal setae (Figure 5C); coxa 1 with only one robust seta anteriorly; gnathopod 1 basis and carpus with few simple and fine plumose setae respectively (Figure 5A); palm of gnathopod 2 curved; anterior margin of basis pereopods 3 and 4 with several simple setae; uropod 3 peduncle with 2 proximal simple and 3 distal sensory cuspidate setae, inner ramus with 2 dorsal sensory cuspidate setae, outer ramus without dorsal setae and with 1 distal robust seta (Figure 5B).

Fig. 5. Aoroides semicurvatus Ariyama, Reference Ariyama2004: specimen from Arcachon Bay, SW France (Body length = 3.94 mm): (A) left gnathopod 1, arrow showing plumose seta, lateral view; (B) right uropod 3, dorsal view; (C) left mandibular palp article 3 with a detailed seta, posterior view. Scale bars: A: 0.5 mm; (B), (C) 0.2 mm.
COLOURATION
Posterior part of head, ventral surface of coxa 1, posterior and lower parts of pereonites 1–5, posterior part of pereonite 7, posterior and ventral parts of pleonite1, ventral part of pleonite 2 brown (Figure 2B).
VARIABILITY
Uropod 3 peduncle: distal end with 2 or 3 sensory cuspidate setae, inner ramus with 1 or 2 dorsal sensory cuspidate setae.
REMARKS
Ariyama (Reference Ariyama2004) described A. semicurvatus with mandibular palp article 3 with 7 marginal and one terminal setae and uropod 3 peduncle without proximal setae and with 2 distal robust setae. Arcachon specimens have only 2 subdistal setae on mandibular palp article 3 and 2 proximal simple and 3 distal sensory cuspidate setae on uropod 3 peduncle. After reexamination of A. semicurvatus specimens in the Osaka Museum of Natural History collections, mandibular palp article 3 shows a great variability with 2 to 12 setae (H. Ariyama, pers. comm.). Setae on gnathopod 1 carpus finely plumose, setae on distal part of uropod 3 rami and mandibular palp with short setules (difficult to see); this information is lacking for Japanese specimens.
HABITAT
This species is present in the lower intertidal zone, under stones, rarely among algae (Ariyama, Reference Ariyama2004). In Arcachon Bay, specimens were typically associated with intertidal oyster reefs (this study).
DISTRIBUTION
Japan (Ariyama, Reference Ariyama2004), France: Arcachon Bay, Hossegor Lake (this study).
Aoroides curvipes Ariyama, Reference Ariyama2004
MATERIEL EXAMINED
North-east Atlantic Ocean: France: Arcachon Bay.
DESCRIPTION
Based on male, BL = 4.1 mm, Les Hosses (Arcachon), October 2009.
Antenna 1 accessory flagellum absent; mandibular palp article 3 with 2 distal setae (Figure 6C); coxa 1 with a robust and a simple setae anteriorly; gnathopod 1 basis with few simple setae, except a group of plumose setae in distal part, merus and carpus with few finely plumose setae (Figure 6A); palm of gnathopod 2 curved; anterior margin of basis pereopods 3 and 4 with several simple setae; uropod 3 peduncle with 3 proximal and 3 distal robust setae, inner ramus with 3 dorsal robust setae, outer ramus with 1 dorsal and 1 distal robust setae (Figure 6B).

Fig. 6. Aoroides curvipes Ariyama, Reference Ariyama2004: specimen from Arcachon Bay, SW France (Body length = 4.1 mm): (A) left gnathopod 1, lateral view; (B) left uropod 3, dorsal view; (C) left mandibular palp article 3, posterior view. Scale bars: A: 0.5 mm; (B), (C) 0.2 mm.
COLOURATION
Dorsal part of head, whole pereonites 1–5, lower part of pereonite 7, dorsal and lower parts of pleonite 1, lower part of pleonites 2–3, coxae 1–5 brown (Figure 2C).
VARIABILITY
Uropod 3 peduncle: distal end with 3 or 4 robust sensory setae; coxa 1 with 1 robust sensory setae and with or without 1 simple setae.
REMARKS
Ariyama (Reference Ariyama2004) described A. curvipes with mandibular palp article 3 with 5 setae and uropod 3 peduncle with 1 proximal and 2 distal robust setae. Arcachon specimens have only 2 subdistal setae on mandibular palp article 3, and 3 proximal and 3 distal robust setae on uropod 3 peduncle. After examination of A. curvipes specimens in the Osaka Museum of Natural History collections, mandibular palp article 3 shows great variability with 3 to 9 setae. Uropod 3 peduncle also shows a great variability with 1 or 2 proximal setae and 1 to 3 distal setae (H. Ariyama, pers. comm.). Setae on distal part of uropod 3 rami and mandibular palp with short setules (difficult to see). French specimens were found in intertidal areas whereas Japan specimens were found in the subtidal.
HABITAT
This species was found in intertidal areas and typically associated with oyster reefs.
DISTRIBUTION
Japan (Ariyama, Reference Ariyama2004), France: Arcachon Bay (this study).
DISCUSSION
In this study, we report the first records of the genus Aoroides in European waters, and assume that the three species recorded along the French Atlantic coast are non-indigenous. One of the species was unambiguously assigned to A. longimerus native to north-western Pacific and which has never been recorded elsewhere. The French specimens of the other two species differ from the Pacific species by very minor morphological details, mainly relative to the number of proximal setae on the uropod 3 peduncle: 2 setae (Atlantic) vs 0 (Pacific) and 3 setae (Atlantic) vs 1–2 (Pacific) in A. semicurvatus and A. curvipes, respectively. However, Japanese specimens also show a great variability in some characters (mandibular palp and uropod 3 ornamentations) (Ariyama, Reference Ariyama2004 and H. Ariyama, pers. comm.). Molecular studies could be helpful for confirming the species identifications (see remarks in the Results section). Taxonomic and systematic issues are common issues in biological invasion studies and non-native surveys (Carlton, Reference Carlton, Rilov and Crooks2009). DNA-based studies, namely molecular barcoding (Comtet et al., Reference Comtet, Sandionigi, Viard and Casiraghi2015), integrative taxonomy (e.g. Geoffroy et al., Reference Geoffroy, Le Gall and Destombe2012), phylogeography and phylogeny (e.g. Dijoux et al., Reference Dijoux, Viard and Payri2014) are efficient approaches to address this problem. Although some sequences are available (e.g. A. columbiae, Best & Stachowicz, Reference Best and Stachowicz2013), no molecular barcodes are yet publicly available for the genus Aoroides but, regarding the European specimens, a first step could be reached by comparing specimens from Arcachon Bay and Japan. This comparison would be valuable to confirm that the documented morphological differences are due to natural variability.
Repeated and numerous successful introductions of Asian species in Arcachon Bay have been well documented. Since the decimation of the Portuguese cupped oyster (Crassostrea angulata (Lamarck, 1819)) by a viral disease in the 1970s and the massive introduction of the Pacific cupped oyster C. gigas from Japan to sustain the local industry, several Asian species were recorded in the bay, including species with either indirect (Arcuatula senhousia (Benson in Cantor, 1842)) (Bachelet et al., Reference Bachelet, Blanchet, Cottet, Dang, de Montaudouin, de Moura Queirós, Gouillieux and Lavesque2009) or direct development (Paranthura japonica Richardson, 1909, Grandidierella japonica Stephensen, 1938) (Lavesque et al., Reference Lavesque, Sorbe, Bachelet, Gouillieux, de Montaudouin, Bonifácio, Blanchet and Dubois2013, Reference Lavesque, Gouillieux, de Montaudouin, Bachelet, Bonifácio and Simonet2014). Due to the presence of sandbanks at the entrance of Arcachon Bay, commercial shipping is absent and recreational boating restricted within the bay. Owing to the strong association of Aoroides semicurvatus and A. curvipes with oyster beds in the bay and the numerous introductions of non-native species with imports of C. gigas in European waters (Brenner et al., Reference Brenner, Fraser, Van Nieuwenhove, O'Beirn, Buck, Mazurié, Thorarinsdottir, Dolmer, Sanchez-Mata, Strand, Flimlin, Miossec and Kamermans2014), we assume that these species were introduced accidentally through aquaculture. The configuration of Hossegor Lake also prevents the entrance of ships in the lake and strengthens the hypothesis that A. semicurvatus was introduced through oyster transfers, probably from other shellfish farming areas in France. Unlike these intertidal species, A. longimerus has been observed in diverse subtidal habitats in Arcachon Bay and under pontoons in the Bay of Brest. In these bays, oysters are massively cultivated and farming may have been the cause of its initial introduction, but its current distribution and common association with biogenic structures suggest more a complex establishment and subsequent spread, in the absence of a larval stage (Ruiz et al., Reference Ruiz, Fofonoff, Carlton, Wonham and Hines2000; Clarke Murray et al., Reference Clarke Murray, Gartner, Gregr, Chan, Pakhomov and Therriault2014). Within Arcachon Bay, adults of A. longimerus may have dispersed away from oyster beds, through natural migration or transport as a fouling organism on the hull of recreational boats, and successfully established in seagrass meadows and slipper limpet beds. In the Bay of Brest, the species A. longimerus was incidentally sampled as part of a global study aiming at examining colonization patterns on hard substrates in three marinas in Brittany. It is noteworthy that A. longimerus was found living on the surface of fouling communities in a marina of Brest, as in its native range in Asia: these communities include several widespread native (Mytilus sp., Ascidiella aspersa (Müller, 1776), Ciona intestinalis (formerly known as C. intestinalis type B; Brunetti et al. Reference Brunetti, Gissi, Pennati, Caicci, Gasparini and Manni2015) and non-indigenous (e.g. Bugula neritina (Linnaeus, 1758), Watersipora subatra (Ortmann, 1890), C. robusta (formerly known as C. intestinalis type A; Brunetti et al. 2015)) species reflecting a boat-shipping network connecting marinas at regional and international scales (Bishop et al., Reference Bishop, Wood, Lévêque, Yunnie and Viard2014). In the Bay of Brest, A. longimerus has been recorded only in the Château marina, at least so far (present study and Le Garrec, unpublished data). In particular, conversely to other non-native taxa found in Château, it was absent from the surveys conducted in two other marinas in Northern Brittany (Leclerc & Viard, in preparation). As such, we may assume that the introduction of A. longimerus has occurred recently in the study area. With regards to the strong similarity of fouling communities across the English Channel marinas (Bishop et al., Reference Bishop, Wood, Lévêque, Yunnie and Viard2014) and its ubiquitous habitat affinity, A. longimerus might further establish and spread in the region as has been documented for other non-native invertebrates that first appeared on the coasts of Brittany (Bishop et al., Reference Bishop, Wood, Lévêque, Yunnie and Viard2014), e.g. the tunicate Asterocarpa humilis (Heller, 1878) (Bishop et al., Reference Bishop, Roby, Yunnie, Wood, Lévêque, Turon and Viard2013).
Aoroides species might be present in European waters for decades but misidentified by marine biologists for different reasons. The main one is that this genus is not listed in the taxonomic keys generally used for amphipod identification in European waters (Chevreux & Fage, Reference Chevreux and Fage1925; Lincoln, Reference Lincoln1979; Ruffo, Reference Ruffo1982). Until now, this genus was only recorded from the North Pacific. Moreover, the specimens are very fragile (loss of gnathopods and antennae) during the sieving process and are thus difficult to identify. Since only males of Aoridae could be identified to species level, most specimens without gnathopods are frequently classified as ‘Aoridae’. Finally, Aoroides species are present in particular habitats (oyster reefs, slipper limpet beds, fouling communities under pontoons) which are not usually sampled in benthic routine surveys using corers or grabs. These three species might be present in other locations in France and in Europe, particularly in shellfish farming areas and marinas. The transfer of C. gigas among regions has caused the introduction and spread of numerous alien species in northern Europe (Goulletquer et al., Reference Goulletquer, Bachelet, Sauriau, Noel, Leppäkoski, Gollasch and Olenin2002). Recreational boating has been only recently pointed as an important vector for both primary and secondary introductions for a diverse set of non-native taxa (Mineur et al., Reference Mineur, Johnson and Maggs2008; Clarke Murray et al., Reference Clarke Murray, Pakhomov and Therriault2011; Ros et al., Reference Ros, Vázquez-Luis and Guerra-García2013).
Identification key to adult males of Aoroides species from the French Atlantic coast (based on Ariyama's (Reference Ariyama2004) key):
-
1. Gnathopod 1 poorly setose. Intertidal species2
-
— Gnathopod 1 densely setose. Subtidal species A. longimerus Ren & Zheng, 1996
-
-
2. Outer ramus of uropod 3 with 1–2 spines, inner ramus with 2–3 spinesA. curvipes Ariyama, Reference Ariyama2004
-
— Outer ramus of uropod 3 marginally bare, inner ramus with 1–2 spines A. semicurvatus Ariyama, Reference Ariyama2004
-
ACKNOWLEDGEMENTS
The authors thank F. Prince and P. Dusoewoir, captain and crew member of RV ‘Planula IV’. We also thank X. de Montaudouin, L. Pascal and S. Gasmi for their help in field sampling. We are very grateful to J.C. Sorbe for his help to confirm the genus Aoroides. The authors would like to thank F. Vandenbosch, W. Thomas, F. Noisette, T. Androuin, G. Schaal, L. M.-L. J. Noël and Div&Co members, especially J. Coudret, T. Broquet, S. Le Cam, C. Roby and T. Comtet for fieldwork assistance and help with sample processing throughout the JCL/FV Commun'touf project. We also thank two anonymous reviewers for helpful comments on the manuscript. Finally, the authors are very grateful to H. Ariyama for his help to confirm our identifications of Aoroides specimens and for his observations of specimens of A. semicurvatus and A. curvipes in the Osaka Museum of Natural History collections.