


INTRODUCTION
Calanus sinicus is a calanoid copepod with wide distribution in continental shelf waters of the north-west Pacific Ocean (Hulsemann, Reference Hulsemann1994). Along the Chinese coast, the range of C. sinicus extends from the Bohai Sea and the Yellow Sea in the north to the northern part of the South China Sea in the south (Chen, Reference Chen1992). In the temperate waters of the Bohai Sea, the Yellow Sea and the East China Sea, C. sinicus occurs throughout the year and is a dominant member of the mesozooplankton in terms of numerical abundance and ecological importance (Chen, Reference Chen1992; Wang et al., Reference Wang, Zuo and Wang2003). In subtropical seas along the coast of southern China, C. sinicus occurs seasonally from late autumn to early spring (Chen, Reference Chen1992; Hwang & Wong, Reference Hwang and Wong2005). While C. sinicus is considered one of the key species of planktonic copepods in the coastal waters of China and Japan (Chen, Reference Chen1964), most previous studies on the seasonal dynamics and life history of C. sinicus have focused on populations in temperate oceans (Uye, Reference Uye2000; Zhang et al., Reference Zhang, Sun and Zhang2005; Xu & Chen, Reference Xu and Chen2007; Zhang et al., Reference Zhang, Sun and Yang2007). Far less information is available on populations in the subtropical waters near the southern edge of the species range.
Hong Kong is located in the southern part of the Chinese coast. A plume of estuarine water from the Zhujiang River covers the western side of Hong Kong during the rainy season from April to September. Oceanic conditions, characterized by high salinities, prevail on the eastern side of Hong Kong. The eastern side of Hong Kong consists of two semi-enclosed bays in the north and open sea area in the south (Figure 1). Tolo Harbour, a landlocked bay opens into Mirs Bay via a narrow tidal channel. Mean water depth in Tolo Harbour varies from ~10 m in the inner shallow basin to about 20 m in the channel. Because of the semi-enclosed topography, Tolo Harbour is characterized by low current velocity and is poorly flushed (Oakley & Cripps, Reference Oakley, Cripps and Ruivo1972). Bordered by Hong Kong on the west and mainland China on the north and east, Mirs Bay has an average water depth of about 25 m and is fully exposed to water currents from the South China Sea.

Fig. 1. Map of Hong Kong showing the locations of Tolo Harbour, Mirs Bay and the six sampling stations.
Results of a previous study revealed that the occurrence of Calanus sinicus in coastal oceans around Hong Kong and Taiwan is limited by temperature to winter and spring (Hwang & Wong, Reference Hwang and Wong2005). It was hypothesized that C. sinicus is carried into the subtropical oceans in the southern part of the species range every winter from population centres in the north by the China Coastal Current when the north-eastern monsoon is blowing (Chen, Reference Chen1992). Understanding the life history of C. sinicus in marginal habitats near the southern edge of its geographical range is important to researchers of the ecosystem dynamics of the South China Sea. In this study, seasonal variations in population structure and body size of C. sinicus are investigated along a transect extending from the inner part of Tolo Harbour to the outer part of Mirs Bay. The objectives are to: (i) investigate the population structure of C. sinicus in subtropical coastal oceans; (ii) identify factors that influence the seasonal and spatial distribution patterns of C. sinicus; and (iii) provide preliminary information on the recruitment mechanisms of C. sinicus in marginal habitats in the northern part of the South China Sea.
MATERIALS AND METHODS
Zooplankton was collected approximately semi-monthly from September 2003 to July 2004 at 6 sampling stations. Two stations, S1 (22°26′725″N, 114°14′566″E) and S2 (22°28′539″N, 114°18′034″E) were located inside Tolo Harbour. Two stations, S3 (22°29′468″N, 114°21′588″E) and S4 (22°26′670″N, 114°26′920″E) were located in Mirs Bay. Two stations, S5 (22°17′560″N, 114°26′920″E) and S6 (22°13′000″N, 114°26′920″E) were located in the coastal sea of south-eastern Hong Kong outside Mirs Bay (Figure 1). Water depths ranged from 15 m at S1, the innermost station, to ~30 m at S5 and S6, the two outermost stations. At each station, duplicate zooplankton samples were taken by hauling a conical plankton net (0.5 m mouth diameter; 125 µm mesh) vertically from ~2 m above bottom to the surface. All zooplankton samples were immediately preserved in 4% formalin seawater. Water temperature and salinity were measured at the surface (0.5 m) using a Hydrolab H20 water quality monitoring system (Austin, USA). Chlorophyll-a concentration in seawater samples collected at the surface (0.5 m) with a Van Dorn type bottle was measured fluorometrically using a Turner Designs 10-AU-000 Fluorometer (Sunnyvale, USA) (Strickland & Parsons, Reference Strickland and Parsons1968).
In the laboratory, C. sinicus in different developmental stages (eggs, nauplii (NI–NVI), copepodites (CI–CV), adult males and adult females) were identified and sorted under a stereomicroscope according to the descriptions of Li & Fang (Reference Li and Fang1983). Densities were determined by counting 25–100% of each sample under a stereomicroscope. Prosome lengths, from the anterior end of the cephalosome to the posterior lateral edge of the last metasome segment, of CVs, adult males and adult females were measured. At least 10 individuals of each developmental stage were measured for each sample.
RESULTS
Environmental conditions
Similar seasonal trends in temperature and salinity were observed at all sampling stations (Figure 2). Surface water temperature ranged from ~15°C in late January and early February to >25°C in May. Salinity ranged from a maximum of 37 at S2 in February to a minimum of 31 at S1 and S3 in early May (Figure 2). Salinity decreased markedly after March, especially in Tolo Harbour, with the approach of the rainy season.

Fig. 2. Temporal variations in water temperature and salinity in the study area during the period when Calanus sinicus was present. Values represent average of six sampling stations. Error bars denote the range.
Chlorophyll-a concentration was much higher in Tolo Harbour than in Mirs Bay (Figure 3). In Tolo Harbour, values spanned widely from 2.67 to 20.30 µg l−1 at S1 and from 0.84 to 8.70 µg l−1 at S2. In comparison, values recorded at the four stations outside Tolo Harbour varied from 0.17 to 4.36 µg l−1.

Fig. 3. Temporal variations in mean chlorophyll-a concentrations (error bars: standard deviation) at six sampling stations during the period when Calanus sinicus was present.
Population abundance and structure
Calanus sinicus was found only in samples collected between late December and early May (Figure 4). The highest densities were recorded in January and February. Averaged across all sampling stations, the density of adults increased from 1.1 ind. m−3 in late December to 78.8 ind. m−3 in early February, then decreased to 13.2 ind. m−3 in early March and <1.6 ind. m−3 in April. No adults were found in samples taken in May. Copepodites also appeared first in December and remained relatively abundant until early March. The maximum average density of 402 ind. m−3 was recorded in late January. Average density dropped abruptly from 384 ind. m−3 in early March to 38.3 ind. m−3 at the end of March. Samples collected in April and May contained <30 ind. m−3. Only a few nauplii of C. sinicus were found in samples collected in December. Average density increased markedly from <30 ind. m−3 in early January to >1100 ind. m−3 at the end of January, and then decreased more or less continuously to <10 ind. m−3 in samples taken in April and May. Eggs of C. sinicus appeared first in samples collected in December but did not become abundant until late January. The maximum average density of 485 ind. m−3 was recorded in early February. No eggs of C. sinicus were found in samples collected in May.

Fig. 4. Temporal variations in the density of Calanus sinicus of different developmental stages at six sampling stations. Solid lines connect average densities of six sampling stations.
Calanus sinicus exhibited spatial variations in abundance throughout the study period (Figure 4). Adults, copepodites, nauplii and eggs appeared at S5 and S6, the outermost stations in December, but did not reach Tolo Harbour until January. Adults and copepodites appeared at S5 and S6 in December, reached S3 and S4 in Mirs Bay in early January, and were found in Tolo Harbour at the end of January. Throughout January and February, the density of adults was much lower at S1 and S2 than at the four outer stations. A similar trend of shoreward decrease was observed in the density of nauplii and copepodites in January. Samples collected in Tolo Harbour in early January contained no nauplii. A few eggs were found in S2 in early January when the average density of eggs in the entire study area was still <50 m−3, but samples collected in Tolo Harbour in late January did not contain eggs even though egg density at S6, the outermost station, had reached a maximum of >1400 m−3. While the density of adults was always highest in the open waters outside Tolo Harbour, samples collected in March showed that eggs, nauplii and copepodites were often abundant at S2 in the outer part of Tolo Harbour and S3 in the inner part of Mirs Bay than at the other stations. Eggs and nauplii disappeared from Tolo Harbour before adults and copepodites. No eggs and nauplii were found in Tolo Harbour after March. In contrast, adults and copepodites remained at S1 until April and at S2 until early May.
Sex-ratio and female fecundity
The ratio of female:male was highest in December when the density of adults in the study area was still very low (Figure 5). Sex-ratio decreased after December. Females were outnumbered by males only in samples taken in early March. The number of eggs per female increased from <15 in December to a peak of 111 in early March. The number of eggs per female decreased markedly after March and no ovigerous females were found in samples taken in May.
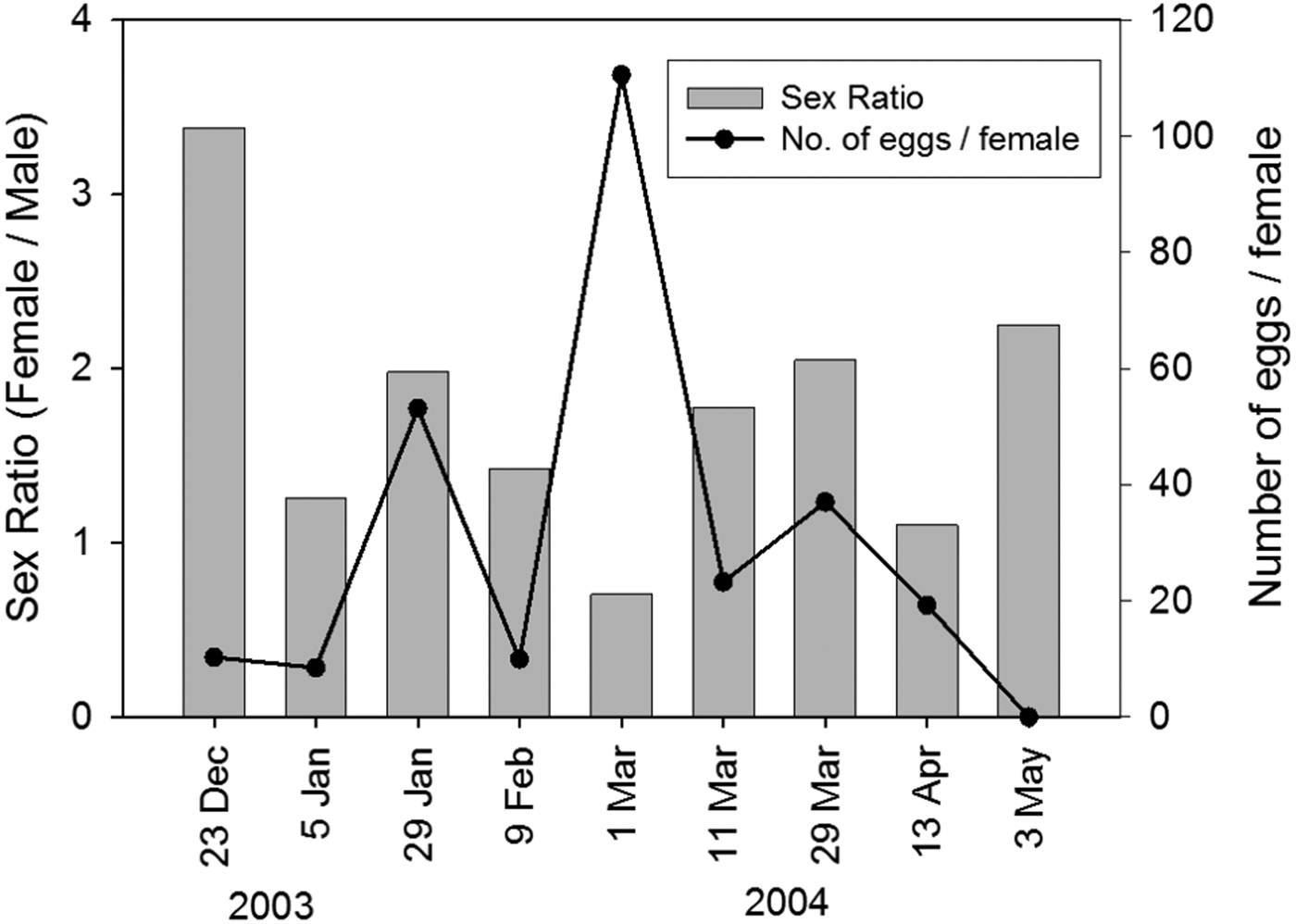
Fig. 5. Temporal variations in the sex-ratio and the number of eggs per female in Calanus sinicus. Values represent average of six sampling stations.
Prosome length
Temporal and spatial changes in the prosome lengths of adults and copepodites of C. sinicus are shown in Figure 6. The prosome length of females, males and CV varied significantly among stations and with time (2-way analysis of variance, P < 0.001). The overall pattern was that individuals from January and February were bigger than individuals from March, April and May (Tukey's multiple comparison tests, P < 0.05), and individuals from S3, S4, S5 and S6 were bigger than individuals from S1 and S2 (Tukey's multiple comparison tests, P < 0.05). Averaged across all stations, the prosome length of female increased from 2.29 mm in December to 2.34 mm in early February, and then decreased gradually to 1.98 mm in early May (Figure 7). Similarly, the average prosome length of male increased from 2.03 mm in December to 2.18 mm at the end of January, and then decreased to 1.86 mm in May (Figure 7). Only one CV individual was found in samples taken in December. The average prosome length of CV was 1.87 mm in early January, 1.99 mm at the end of January, and 1.67 mm in May (Figure 7).

Fig. 6. Temporal variations in the average prosome length of males, females and copepodites (CVs) at six sampling stations. Vertical bars denote the standard deviation. Broken horizontal lines mark the average prosome length at each sampling station.

Fig. 7. Temporal variations in the prosome length of males, females and copepodites (CVs). Values represent average of six sampling stations. Vertical bars denote the range.
DISCUSSION
Population dynamics
Calanus sinicus appears in the subtropical coastal waters of eastern Hong Kong in winter and early spring. Adults and juveniles first appear in the open waters of south-eastern Hong Kong and Mirs Bay at the end of December. Density increases rapidly in January. By the end of January, individuals of all developmental stages can be found in the innermost station in the shallow waters of the semi-enclosed Tolo Harbour. The highest density is recorded in late January and early February, when average water temperature has dropped to <15°C. Densities recorded in January and February are comparable to those encountered in the population centres in the Yellow Sea (Liu et al., Reference Liu, Sun, Wang, Zhang, Yang and Ji2003; Wang et al., Reference Wang, Zuo and Wang2003). Overall, observations from this study and an earlier study (Hwang & Wong, Reference Hwang and Wong2005) confirm that the occurrence of C. sinicus in coastal seas of southern China coincides with the north-eastern monsoon period in winter, when cold water masses carrying C. sinicus from the Yellow Sea and East China Sea are driven into the South China Sea through the Taiwan Strait (Hwang et al., Reference Hwang, Tu, Tseng, Fang, Souissi, Fang, Lo, Twan, Hsaio, Wu, Peng, Wei and Chen2004, Reference Hwang, Souissi, Tseng, Seuront, Schmitt, Fang, Peng, Wu, Hsiao, Twan, Wei, Kumar, Fang, Chen and Wong2006).
Data presented in this study and in many previous studies suggest that temperature is an important factor regulating the distribution of C. sinicus. In the Inland Sea of Japan, the biomass of C. sinicus reaches a maximum at ~20°C (Uye, Reference Uye2000). The upper thermal limit of C. sinicus in laboratory cultures is considered to be ~23°C (Huang & Zheng, Reference Huang and Zheng1986; Uye, Reference Uye1988). In Xiamen Harbour, C. sinicus disappears in June when water temperature reaches 24°C (Lin & Li, Reference Lin and Li1984). Zooplankton surveys conducted along the Chinese coast and the Taiwan Strait have generally confirmed that C. sinicus disappears when water temperature exceeds 26–27°C (Wang et al., Reference Wang, Zuo and Wang2003; Hwang et al., Reference Hwang, Souissi, Dahms, Tseng, Schmitt and Chen2009). In a more recent study, C. sinicus is found to be abundant in western and north-eastern Taiwan, but absent in eastern Taiwan where the main branch of the Kuroshio Current warms the surface water to 23.5°C in January to 29.2°C in July (Hsiao et al., Reference Hsiao, Fang, Shih and Hwang2011). Results presented here show that temperature influences the occurrence of C. sinicus in the coastal waters of eastern Hong Kong. There is a negative correlation between temperature and density in C. sinicus nauplii and adults (Table 1). The only increasing trend in the density of C. sinicus of all developmental stages occurs between December and early February when water temperature is <20°C and decreasing. Decline in the density of all developmental stages begins in February when water temperature is just starting to increase. Very few individuals remain in the water column in May when water temperature approaches 25°C. In general, it can be concluded that the density of C. sinicus increases when temperature is <20°C and decreases when surface water temperature begins to rise.
Table 1. Pearson correlation coefficients between the abundance and prosome length of Calanus sinicus of different developmental stages and various environmental parameters. Statistically significant correlations (P < 0.05, N = 24) are marked by asterisks.

Temperature probably also influences the size of the C. sinicus population that can be established in the subtropical coastal seas around Hong Kong. Densities recorded in this study were much higher than those recorded in a study conducted between 2000 and 2002 in an area near S5 and S6 (Hwang & Wong, Reference Hwang and Wong2005). While the previous study was conducted in an area nearer to the shore than S5 and S6, the lowest temperature encountered was also 2–3°C higher than the 15°C recorded in this study.
In temperate regions, from the Bohai Sea to the East China Sea, population size of C. sinicus decreases abruptly after June, when water temperature goes up to 15–20°C (Chen, Reference Chen1964). In the Yellow Sea, C. sinicus has been known to survive the summer in cold waters in the thermally stratified deep areas (Wang et al., Reference Wang, Zuo and Wang2003; Zhang et al., Reference Zhang, Sun and Yang2007). The duration of the occurrence of C. sinicus also increases northward along the Chinese coast. In Hong Kong, C. sinicus occurs from December to May. Further north in Xiamen Harbour in the Taiwan Strait, C. sinicus occurs between November and early June (Lin & Li, Reference Lin and Li1984; Dur et al., Reference Dur, Hwang, Souissi, Tseng, Wu, Hsiao and Chen2007; Hsiao et al., Reference Hsiao, Fang, Shih and Hwang2011). In the coastal seas of northern Taiwan, the East China Sea and the Yellow Sea, C. sinicus occurs throughout the year (Chen, Reference Chen1992; Wang et al., Reference Wang, Zuo and Wang2003; Hwang & Wong, Reference Hwang and Wong2005; Xu & Chen, Reference Xu and Chen2007). The north and middle parts of the Taiwan Strait are considered to be the southern limit for the perennial occurrence of C. sinicus (Chen, Reference Chen1964; Huang et al., Reference Huang, Li and Chen2002). Outside Xiamen Harbour, C. sinicus disappears completely between June and late November from nearshore areas, but persists in deep offshore waters (Huang et al., Reference Huang, Li and Chen2002). The population is able to reproduce and recruit rapidly when temperature becomes more favourable during autumn. Shantou is located 200 km south-west of Xiamen and 300 km north-east of Hong Kong (Figure 1). Temperature in the coastal areas outside Shantou drops to <22°C in November and reaches a minimum of 16.4°C in January (Jiang et al., Reference Jiang, Huang, Chen, Zhou and Dong2002). Calanus sinicus can be found in Shantou from late November to April (Huang, personal communication). The time of first appearance in Shantou is almost similar to that in Xiamen and approximately one month earlier than that in Hong Kong.
Correlation between salinity and density in C. sinicus adults is positive (Table 1). Nevertheless, the distribution of C. sinicus may not be strongly influenced by salinity. In Hong Kong, beginning of the rainy season and a gradual decrease in surface salinity coincide with the rise in temperature in spring (Figure 2). Calanus sinicus can tolerate a wide range of salinity (Huang & Zheng, Reference Huang and Zheng1986). In the Yellow Sea, C. sinicus is well adapted to salinities >29, but its distribution is probably not limited by salinity (Wang et al., Reference Wang, Zuo and Wang2003; Zhang et al., Reference Zhang, Sun and Yang2007; Hwang et al., Reference Hwang, Souissi, Dahms, Tseng, Schmitt and Chen2009). Surface water in the western part of Hong Kong is strongly influenced by fresh water discharged from the Zhujiang River during the summer. In estuarine water with salinity <30, C. sinicus has been found to occur in densities comparable to those recorded in this study (Wong et al., Reference Wong, Tam, Fu and Chen2000).
No significant correlation is found between chlorophyll-a concentration and the abundance of C. sinicus (Table 1). In the Yellow Sea, non-phytoplankton food such as ciliates formed a significant portion of the diet of C. sinicus (Huo et al., Reference Huo, Wang, Sun, Li and Liu2008), suggesting that the abundance of C. sinicus is not necessarily related to the supply of phytoplankton. On the other hand, the egg production rate of C. sinicus in the Inland Sea of Japan declined when chlorophyll-a concentration in phytoplankton >5 µm was <0.5 µg l−1 (Uye & Murase, Reference Uye and Murase1997). Chlorophyll-a concentrations at the two stations in the south-eastern corner of Hong Kong dropped below 0.5 µg l−1 in April and May, therefore decline in chlorophyll-a concentrations may contribute to the rapid disappearance of C. sinicus after March.
Prosome length
Seasonal changes in prosome length suggest that the body size of C. sinicus may be influenced by temperature. Between December and early March, the average prosome length of adult females varies between 2.29 and 2.34 mm, and is comparable to the 2.22–2.38 mm recorded for populations in the Yellow Sea (Zhang et al., Reference Zhang, Sun and Zhang2005). The average prosome length of females decreases to 2.14 mm in the middle of March when water temperature is rising but still <23°C. In May, when water temperature averages >25°C, the average prosome length of the few surviving females decreases to 1.98 mm. For comparison, it is worth noting that the prosome length of most females in the Yellow Sea is >2.1 mm, although small individuals with prosome length as short as 1.90 mm have also been recorded (Zhang et al., Reference Zhang, Sun and Zhang2005).
In regions with a wide temperature range, the body size of copepods may be affected by water temperature (Hirche, Reference Hirche1992; Ban, Reference Ban1994). The body size of calanoid copepods has been known to correlate negatively with temperature, with the smallest individuals occurring at the highest temperature (Halsband & Hirche, Reference Halsband and Hirche2001; Halsband-Lenk et al., Reference Halsband-Lenk, Hirche and Carlotti2002). Data presented here show a significant negative correlation between prosome length and temperature in C. sinicus CVs and adult males (Table 1). Prosome length of all developmental stages fluctuates between December and February, but follows a clear decreasing trend in March as water temperature begins to increase. In both CVs and adults, the smallest individuals appear in May when water temperature is high and the population is just about to disappear from the water column.
Size selective predation is known to play an important role in controlling the size composition of zooplankton communities (Brooks & Dodson, Reference Brooks and Dodson1965). Dense schools of the larval and juvenile fishes appear in Tolo Harbour in spring. Copepods form an important part of the diet of these small fishes (Nip et al., Reference Nip, Ho and Wong2003). Planktivorous fishes rely on vision to capture the biggest and most conspicuous prey (O'Brien, Reference O'Brien1979). In shallow areas where C. sinicus cannot rely on vertical migration to avoid fish predators, size selective predation may favour individuals with smaller body size.
Recruitment
Calanus sinicus is carried into the coastal seas around Hong Kong from population centres in the Yellow Sea and the East China Sea every winter by the China Coastal Current (Hwang & Wong, Reference Hwang and Wong2005). In the eastern part of Hong Kong, C. sinicus first appears in the open waters outside Mirs Bay in December and reaches the inner part of Tolo Harbour approximately one month later in January. This sequence of appearance suggests that C. sinicus in the northern part of the South China Sea was first recruited to the open seas around Hong Kong, and then to the shallow, semi-enclosed waters of Mirs Bay and Tolo Harbour. The delay in the appearance of C. sinicus in Tolo Harbour is probably due to the low level of water exchange between the semi-enclosed bay and the adjacent sea areas. The copepod community in Tolo Harbour is marked by a scarcity of large species (Zhang & Wong, Reference Zhang and Wong2011). Calanus sinicus is a prominent member of the zooplankton community in Tolo Harbour and Mirs Bay because of its large size. Calanus sinicus adults and copepodites that enter into the shallow waters of Tolo Harbour are probably removed rapidly by small fishes which make their appearance in early spring (Nip et al., Reference Nip, Ho and Wong2003). Indeed, there is a shoreward decrease in abundance of C. sinicus even during the period of peak densities in late January and early February, and this could be caused by patterns of the current as well as predation.
The presence of ovigerous females suggests that reproduction occurs between January and March. The maximum clutch size of 111 eggs per female was comparatively higher than those recorded for populations in the Yellow Sea (Zhang et al., Reference Zhang, Sun and Zhang2005) and the Inland Sea of Japan (Uye, Reference Uye2000). Since clutch size increases with prosome length (Zhang et al., Reference Zhang, Sun and Zhang2005), it is surprising to note that females produce their biggest clutches in early March when average prosome length is already decreasing. On the other hand, high food concentrations in February may allow females to increase their reproductive effort in March. At 20°C, eggs take 14 hours to hatch (Zhang et al., Reference Zhang, Sun and Zhang2005) and develop into adults in 16 days (Uye, Reference Uye1988). While there is sufficient time for eggs spawned locally to develop into nauplii and copepodites, study of the effect of mud on the survival of C. sinicus egg by Uye (Reference Uye2000) suggests that most eggs would die very quickly as they sink and come into contact with the sediment in the shallow waters of Tolo Harbour and Mirs Bay. Experiments in the Yellow Sea revealed that high temperature is detrimental to the hatching of C. sinicus (Zhang et al., Reference Zhang, Sun and Yang2007). Continual decline in the density of nauplii after March also suggests that most eggs do not hatch into nauplii.
Decrease in the prosome length of C. sinicus begins in March and the smallest females appear in May, just before the disappearance of the species from the study area. While selective predation by visual predators may favour smaller individuals, the observed seasonal pattern in body size suggests that environmental conditions, particularly high temperature, in the subtropical seas around Hong Kong are detrimental to C. sinicus after March. Change in the direction of the monsoon winds, discontinuation in the intrusion of cold water masses from the north, and influx of warm water masses from the South China Sea cause a rapid increase in surface water temperature in the northern part of the South China Sea. No over-summer populations of C. sinicus have been reported in the coastal seas of southern China. Copepods can retreat to the bottom when the water temperature at the surface becomes stressful. In the southern part of the Yellow Sea, C. sinicus retreats to the cold bottom waters during summer (Wang et al., Reference Wang, Zuo and Wang2003). A similar situation occurs in the Taiwan Strait, where C. sinicus disappears completely between June and late November from nearshore areas outside Xiamen Harbour, but persists in deep offshore waters (Huang et al., Reference Huang, Li and Chen2002). At present, no data are available to suggest that C. sinicus populations can survive in deeper areas in the northern part of the South China Sea. In eastern Taiwan where surface water is warmed by the Kuroshio Current, C. sinicus is absent even in areas with water depth up to 800 m (Hsiao et al., Reference Hsiao, Lee, Shih and Hwang2004).
CONCLUSIONS
In conclusion, the occurrence of C. sinicus in the coastal seas of eastern Hong Kong is restricted to the period between December and May. Densities comparable to those recorded in population centres in the Yellow Sea and East China Sea (Wang et al., Reference Wang, Zuo and Wang2003) are encountered only in late January and early February when water temperature drops to ~15°C. Females with eggs appear between December and April, but most eggs probably do not hatch into nauplii. Prosome length and clutch size decrease after March when water temperature begins to rise. The absence of a summer population and the observed seasonal change in prosome length, with individuals collected in March and April, just before the disappearance of the population, being much smaller than those collected in January when the population first appears, indicate that C. sinicus in the coastal waters of Hong Kong is recruited from populations in the northern part of the species range. The shallow seas of eastern Hong Kong provide only marginal habitats for C. sinicus populations which have been transported into subtropical oceans on the southern edge of the species range during winter and early spring by water currents.
ACKNOWLEDGEMENTS
We are grateful to Alle Lie, Eva Yau, Vivian Li and Y.H. Yung for helping with zooplankton sampling. We thank Pan Tse and K.C. Cheung for technical assistance. This work was supported by the Chinese Ecosystem Research Network (CERN), the Chinese National Ecosystem Observation and Research Network (CEORN), the Science and Technology Ministry of China (973-G1999043708 and 2006CB400606), and a Direct Grant for Research from The Chinese University of Hong Kong.








