



Introduction
The conodont species Histiodella labiosa Bauer, Reference Bauer2010, was erected based on a large collection of ~997 specimens from 27 samples from the middle to upper part of the Oil Creek Formation exposed near US Highway 77 and Interstate 35 in southern Oklahoma (Fig. 1). Based on the distinctive morphological features of this species, Bauer (Reference Bauer2010) established the H. labiosa Biozone in the Oil Creek Formation for a stratigraphic interval between the H. sinuosa and H. holodentata biozones. However, outside North America, H. labiosa has only been recorded from the lower Darriwilian in Australia. It is likely that the morphological entity represented by H. labiosa was lumped either in H. holodentata Ethington and Clark, Reference Ethington and Clark1982, or in H. serrata Harris, Reference Harris1962. To overcome this drawback and inconsistencies in taxonomy, H. labiosa is revised herein based on the study of originally illustrated types of this species, including a newly selected lectotype (Bauer, Reference Bauer2010, pl. 2, fig. 13), other paralectotypes (Bauer, Reference Bauer2010, pl. 2, figs. 10, 11, 14), and re-examination of the original and additional topotype material from the type locality. Abundant topotype material of H. labiosa has illustrated the morphological variation of its Pa, Pb, and M elements. Detailed study of this morphological variation allows us to analyze the phylogenetic relationships of H. labiosa with its closely related species and to better elucidate the evolution of the Histiodella species through late Dapingian to middle Darriwilian during the Middle Ordovician.
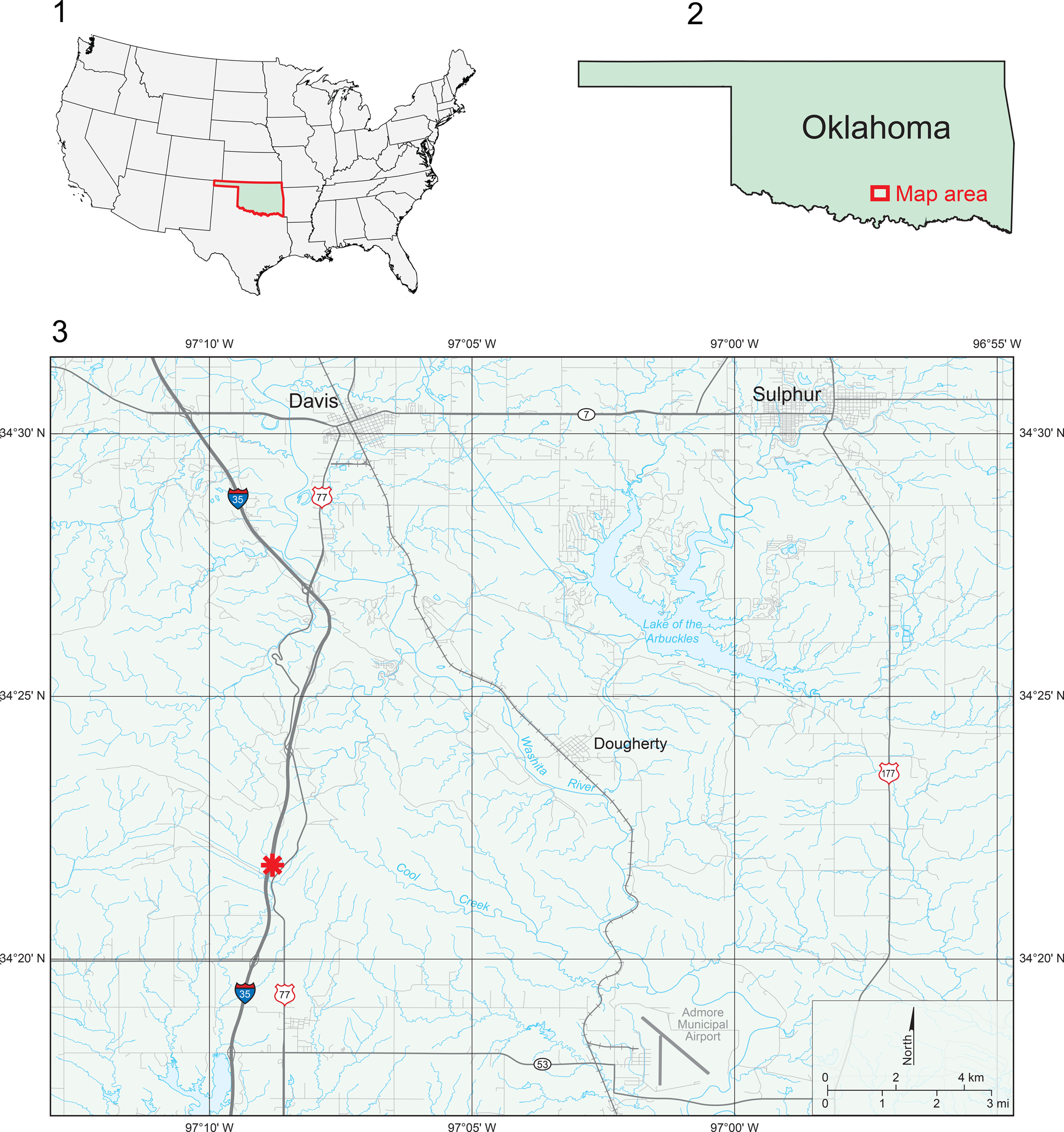
Figure 1. Location maps showing the study area in Oklahoma of the USA (1), location of study area in southern Oklahoma (2), and (3) the detailed location of the sampled section (marked by *) of the Oil Creek Formation with samples from the section bearing designations 72SC (collected by Bergström, Jaanusson, and Sweet in 1972) and 89JA (collected by Bauer in 1989), which is exposed near US Highway 77 and Interstate 35 in southern Oklahoma.
Geological setting and regional stratigraphy
The Arbuckle Mountains region of southern Oklahoma contains one of the best and most-continuous exposures (nearly 3350 m in total thickness) of fossiliferous upper Cambrian to Devonian rocks in the Midcontinent of the United States (Ham, Reference Ham1973; Carlucci et al., Reference Carlucci, Goldman, Brett, Westrop and Leslie2015). The sequences were uplifted regionally in association with folding and faulting during the Ouachita Orogeny, a mountain building event in the Pennsylvanian and Permian (Thomas, Reference Thomas1991; Ettensohn et al., Reference Ettensohn, Pashin, Gilliam and Miall2019).
The Simpson Group is a Middle to Late Ordovician carbonate and clastic succession reaching a maximum thickness of >760 m. It is widely distributed in southern Oklahoma and extends farther southwest into West Texas and southeastern New Mexico. In southern Oklahoma, the Simpson Group includes, in ascending order, the Joins, Oil Creek, McLish, Tulip Creek, and Bromide formations (Decker and Merritt, Reference Decker and Merritt1931). It was deposited during a major marine transgression and followed deposition of the upper (Lower to lowest Middle Ordovician) Arbuckle Group (Derby et al., Reference Derby, Bauer, Creath, Dresbach, Ethington, Loch, Stitt, McHargue, Miller, Miller and Repetski1991; Ethington et al., Reference Ethington, Repetski, Derby, Derby, Fritz, Longacre, Morgan and Sternbach2012; Morgan, Reference Morgan, Derby, Fritz, Longacre, Morgan and Sternbach2012; Read and Repetski, Reference Read, Repetski, Derby, Fritz, Longacre, Morgan and Sternbach2012; Taylor et al., Reference Taylor, Repetski, Loch, Leslie, Derby, Fritz, Longacre, Morgan and Sternbach2012). The Middle and Upper Ordovician are characterized by intervals of predominantly shales and clay-rich carbonates deposited during maximum flooding, whereas sandstone dominates intervals laid down during the low standing of sea level fluctuations.
The Joins Formation, with a total thickness of close to 90 m, is primarily thin- to medium-bedded limestone with shale interlayers that overlies (locally unconformable) the West Spring Creek Formation of the Arbuckle Group of latest Cambrian to earliest Middle Ordovician age (Derby et al., Reference Derby, Bauer, Creath, Dresbach, Ethington, Loch, Stitt, McHargue, Miller, Miller and Repetski1991; Ethington et al., Reference Ethington, Repetski, Derby, Derby, Fritz, Longacre, Morgan and Sternbach2012; Morgan, Reference Morgan, Derby, Fritz, Longacre, Morgan and Sternbach2012; Taylor et al., Reference Taylor, Repetski, Loch, Leslie, Derby, Fritz, Longacre, Morgan and Sternbach2012). The conformably succeeding Oil Creek Formation, with a maximum thickness of just over 300 m, consists of a lower sandstone member and an upper limestone member of thin- to medium-bedded limestone and interbedded greenish gray shales that gradationally become more dominant towards the top (Henry, Reference Henry1988). At the type locality (section 72SC) of H. labiosa, which is the focus of this study, the Oil Creek Formation has a total thickness of 193 m (Fig. 2).

Figure 2. Composite stratigraphic section of the 72SB (Joins Formation) and 72SC (Oil Creek Formation) showing the stratigraphic horizon of the lectotype (from sample 72SC-500, marked with red font and a red star) and horizons of the 10 conodont samples examined in this study. Conodont biozones and stratigraphic occurrences and ranges of Histiodella species recognized in the Joins and Oil Creek formations at this section in southern Oklahoma are based on Bauer (Reference Bauer2010). Da. = Dapingian.
As the type localities and type strata of several species of Histiodella Harris, Reference Harris1962, the Joins Formation and the overlying Oil Creek Formation in the Arbuckle Mountains region are fundamentally important in the study of this morphologically distinctive Middle Ordovician conodont genus. These two carbonate-dominated units of southern Oklahoma may represent the most continuous Middle Ordovician succession in the world that yields abundant material of several Histiodella species. They have been the focus of several classical works in the past six decades (e.g., Harris, Reference Harris1962; Mound, Reference Mound1965; McHargue, Reference McHargue1975, Reference McHargue1982; Bauer, Reference Bauer2010), particularly to study species of Histiodella in establishing a finely defined biozonal scheme, which is now widely used to subdivide the late Dapingian to middle Darriwilian strata worldwide (Bergström and Ferretti, Reference Bergström and Ferretti2017; Goldman et al., Reference Goldman, Sadler, Leslie, Gradstein, Ogg, Schmitz and Ogg2020; Zhen, Reference Zhen2021).
Materials and methods
The topotypes of H. labiosa used in this study represent a total of 427 discrete specimens recovered from four samples from the Oil Creek Formation. They include sample 72SC-450 (137.16 m above the base of the Oil Creek Fm., with 128 specimens recovered), sample 72SC-460 (140.21 m above the base of the Oil Creek Fm., with 53 specimens recovered), sample 72SC-510 (155.45 m above the base of the Oil Creek Fm., with 79 specimens recovered), and sample 72SC-520 (158.5 m above the base of the Oil Creek Fm., with 167 specimens recovered). Section 72SC starts at coordinates 34.365000°N, 97.145833°W, and ends at the top of the Oil Creek Formation at coordinates 34.362500°N, 97.146667°W. The section is exposed in several low ridges between US Highway 77 and Interstate 35 (SE1/4, sec. 24, T.2S., R.1E., Carter County), in southern Oklahoma. The lectotype (Bauer, Reference Bauer2010, pl. 2, fig. 13) is from sample 72SC-500, collected 152.4 m above the base of the Oil Creek Fm., with 75 specimens of H. labiosa recovered. They represent the most productive samples of H. labiosa among the 27 samples that yielded this species when Bauer (Reference Bauer2010) originally studied and erected H. labiosa. All samples were collected by S.M. Bergström, the late V. Jaanusson, and the late Prof. W.C. Sweet in 1972. Topotype material of H. labiosa was recovered by Jeffrey A. Bauer in 1989 in a nearby section exposed along the east side of Interstate 35 (Fig. 2) from six additional samples: 89JA-178 (175 m above the base of the Oil Creek Fm., with 310 specimens recovered), 89JA-184.5 (181.5 m above the base of the Oil Creek Fm., with six specimens recovered), 89JA-180 (177 m above the base of the Oil Creek Fm., with seven specimens recovered), 89JA-150 (147 m above the base of the Oil Creek Fm., with two specimens recovered), 89JA-146 (143 m above the base of the Oil Creek Fm., with one specimen recovered), and 89JA-198 (195 m above the base of the Oil Creek Fm., with one specimen recovered). The S and M elements of the Histiodella species are generally smaller than the Pa and Pb elements. Repetski (personal communication, 2021) indicated that a 200-mesh bottom sieve would catch S and M elements, whereas most or all of these elements would pass through a 140-mesh sieve. When the samples of this study were processed, unfortunately a 140-mesh sieve was used, which might be responsible for the bias of the element types recovered.
Repository and institutional abbreviation
Thirty topotype specimens of H. labiosa illustrated in Figures 3–6 bearing the prefix OSU (OSU 54951–54980, inclusive) are deposited in the Type Collection of the Orton Geological Museum, Orton Hall, School of Earth Sciences, The Ohio State University, Columbus, USA. All images illustrated are SEM digital photomicrographs (numbers with the prefix IY are the file names of the digital images).
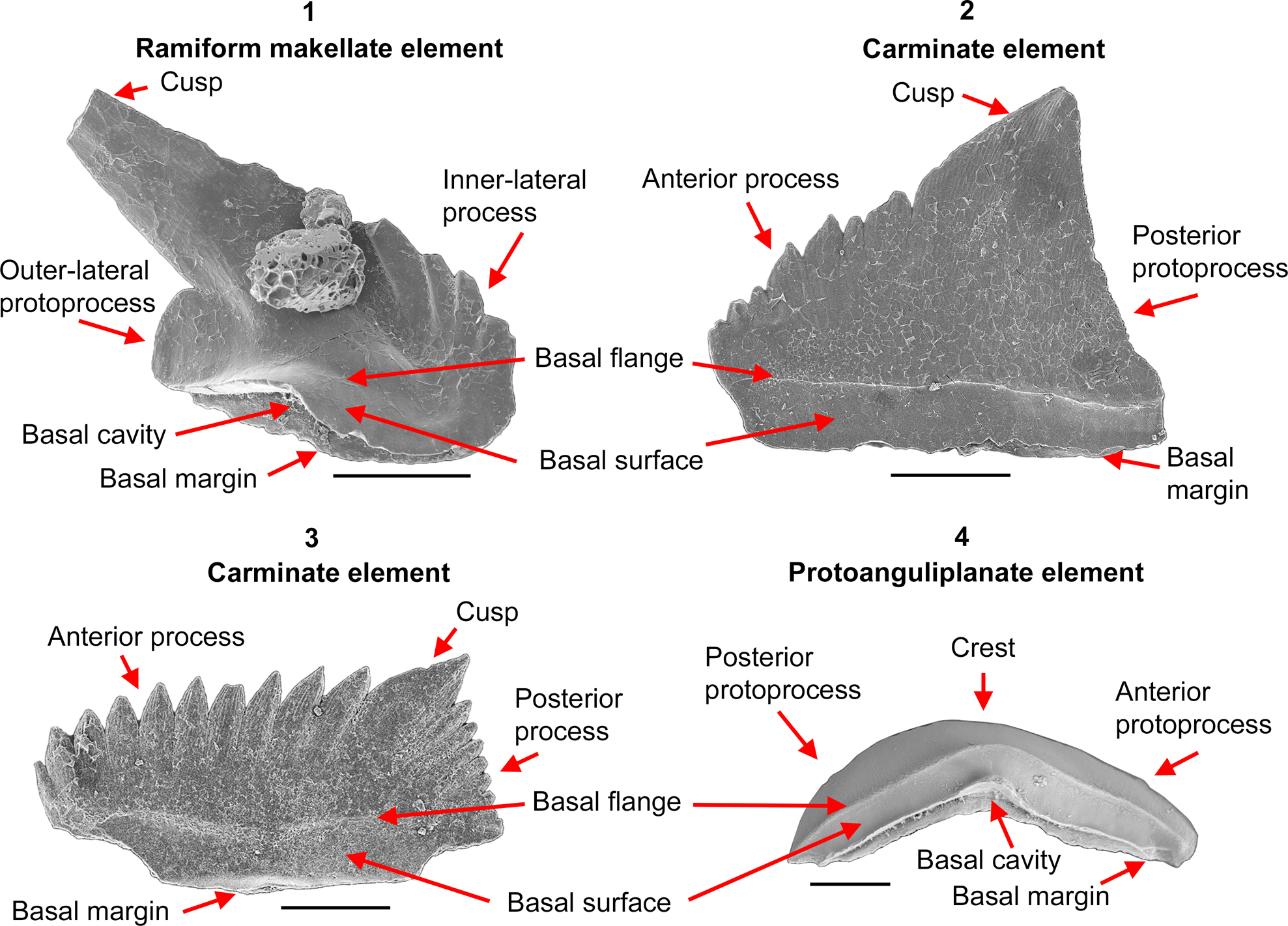
Figure 3. Orientation, morphology, and structural terms for Histiodella and related genera: (1) M element of Histiodella labiosa Bauer, Reference Bauer2010; (2) Pb element of Histiodella labiosa Bauer, Reference Bauer2010; (3) Pa element of Histiodella holodentata Ethington and Clark, Reference Ethington and Clark1982; (4) Protoanguliplanate P element of Cooperignathus nyinti (Cooper, Reference Cooper1981). All scale bars represent 100 μm.

Figure 4. Histiodella labiosa Bauer, Reference Bauer2010, from the Oil Creek Formation in the section 72SC exposed in southern Oklahoma: Pa element. (1) OSU 54951, 72SC-450, inner-lateral view (IY406-15); (2) OSU 54952, 72SC-520, apical-outer-lateral view (IY406-03); (3) OSU 54953, 72SC-460, outer-lateral view (IY406-17); (4, 5) OSU 54954, 72SC-510, (4) inner-lateral view (IY406-29), (5) close up showing surface striation (IY406-30); (6) OSU 54955, 72SC-520, inner-lateral view (IY406-01); (7) OSU 54956, 89JA-178, basal view (IY416-19); (8) OSU 54957, 89JA-178, apical view (IY416-05); (9) OSU 54958, 72SC-510, outer-lateral view (IY406-22); (10) OSU 54959, 72SC-510, inner-lateral view (IY406-23); (11) OSU 54960, 72SC-520, outer-lateral view (IY406-02); (12) OSU 54961, 89JA-178, outer-lateral view (IY416-01). All scale bars represent 100 μm, unless otherwise indicated.

Figure 5. Histiodella labiosa Bauer, Reference Bauer2010, from the Oil Creek Formation in the section 72SC exposed in southern Oklahoma: Pb element. (1) OSU 54962, 72SC-460, outer-lateral view (IY406-06); (2) OSU 54963, 72SC-520, inner-lateral view (IY406-06); (3, 4) OSU 54964, 72SC-510, (3) inner-lateral view (IY406-25), (4) close up showing minutely serrated posterior edge (IY406-26); (5) OSU 54965, 72SC-450, inner-lateral view (IY406-14); (6) OSU 54966, 72SC-460, outer-lateral view (IY406-21); (7) OSU 54967, 72SC-520, inner-lateral view (IY406-09); (8) OSU 54968, 89JA-178, apical view (IY416-13); (9) OSU 54969, 89JA-178, outer-lateral view (IY416-17); (10) OSU 54970, 89JA-178, outer-lateral view (IY416-12); (11) OSU 54971, 72SC-460, outer-lateral view (IY406-20). All scale bars represent 100 μm, unless otherwise indicated.

Figure 6. Histiodella labiosa Bauer, Reference Bauer2010, from the Oil Creek Formation in the section 72SC exposed in southern Oklahoma. (1) Sc element, OSU 54972, 89JA-178, outer-lateral view (IY416-20); (2–9) M element: (2) OSU 54973, 89JA-178, basal-posterior view (IY416-24); (3) OSU 54974, 72SC-510, posterior view (IY406-28); (4) OSU 54975, 89JA-178, apical view (IY416-27); (5) OSU 54976, 89JA-178, posterior view (IY416-30); (6) OSU 54977, 89JA-178, posterior view (IY416-29); (7) OSU 54978, 89JA-178, posterior view (IY416-25); (8) OSU 54979, 89JA-178, anterior view (IY416-26); (9) OSU 54980, 89JA-178, anterior view (IY416-28). All scale bars represent 100 μm.
Systematic paleontology
Morphological and apparatus notational terms defined in the conodont Treatise (Clark et al., Reference Clark, Sweet, Bergström, Klapper, Austin, Rhodes, Müller, Ziegler, Lindström, Miller, Harris and Robison1981) are followed in this contribution, except for the following technical terms, which are briefly defined herein (Fig. 3).
Basal flange
A costa-like structure presented as a prominent shelf-like projection, which extends around the element parallel to, and slightly above, the basal margin to define the upper limits of the basal surface. This character is well developed in species of Cooperignathus, Jumudontus, and Histiodella, as well as in some species of Protoprioniodus, Oelandodus (likely a junior synonym of Protoprioniodus), and Fahraeusodus.
Basal surface
The surface adjacent to the basal margin of the elements located between the basal margin and the basal flange.
Makellate element
A term referring to most types of the elements taking the M position in the apparatus. The most distinctive feature of a makellate element is its orientation as an antero-posteriorly compressed ramiform, protoramiform, or coniform element (Sweet, Reference Sweet1988; Nicoll, Reference Nicoll1990, Reference Nicoll1992).
Protoprocess
Adentate anterior, posterior, or lateral projections of protoramiform, ramiform, protopectiniform, or pectiniform elements.
Protocarminate element
A blade-like protopectiniform element with a prominent cusp, posterior and anterior protoprocesses, and a straight or nearly straight basal margin.
Protoanguliplanate element
An arched protopectiniform element with the upper surface developed as a thin, antero-posteriorly extended, blade-like crest, which lacks a recognizable cusp and denticles; typically represented by the P element of Cooperignathus nyinti (Cooper, Reference Cooper1981) and C. aranda (Cooper, Reference Cooper1981).
Class Conodonta Pander, Reference Pander1856
Order Ozarkodinida Dzik, Reference Dzik1976
Suborder Plectodinida Dzik, Reference Dzik1991
Superfamily Oistodontacea Lindström, Reference Lindström1970
Family Oistodontidae Lindström, Reference Lindström1970
Genus Histiodella Harris, Reference Harris1962
Type species
Histiodella altifrons Harris, Reference Harris1962.
Diagnosis
Seximembrate (or septimembrate) apparatus of adentate (in early species) to denticulate (in later species) elements, including geniculate makellate M element, protoramiform or ramiform S elements, which form a symmetry transition series from alate prototriform or triform Sa, asymmetrical prototertiopedate (or modified tertiopedate) Sb, to asymmetrical and laterally compressed protobipennate or modified bipennate Sc and possibly protoquadriramate or quadriramate Sd, and protocarminate or carminate P elements typically with well-defined basal surface between the basal flange and basal margin (modified after McHargue, Reference McHargue1982, p. 1430–1431, and Stouge, Reference Stouge2012, p. 82).
Remarks
Two species of Histiodella, including the type species of the genus (H. altifrons) and H. serrata, were erected by Harris (Reference Harris1962) as form species based on the Pa elements (protocarminate for H. altifrons and carminate for H. serrata) from the Joins Formation exposed in the Arbuckle Mountains of southern Oklahoma. Harris (Reference Harris1962, p. 209) distinguished the latter by its larger size with denticulate upper margin of the anterior and posterior processes. Mound (Reference Mound1965) subsequently erected H. minutiserrata and H. triquetra as two additional form species from the Joins Formation of the Arbuckle Mountains, Oklahoma. The former was based on the protocarminate Pa element differing from H. altifrons in having a minutely serrated upper margin, and H. triquetra was based on the prototriform alate element. Histiodella sinuosa (Graves and Ellison, Reference Graves and Ellison1941) was established as a form species based on the carminate Pa element characterized by having a denticulate anterior process and adentate posterior process. The type material of H. sinuosa, which is from the Fort Peña Formation of West Texas (Graves and Ellison, Reference Graves and Ellison1941; Bradshaw, Reference Bradshaw1969), was also reported from upper part of the Joins Formation and lower and middle part of the Oil Creek Formation in Arbuckle Mountains of southern Oklahoma (McHargue,1982; Bauer, 2010) and correlative units in Utah (Ethington and Clark, Reference Ethington and Clark1982) and Nevada (Harris et al., Reference Harris, Bergström, Ethington and Ross1979; Sweet et al., Reference Sweet, Ethington and Harris2005) of North America. Bradshaw (Reference Bradshaw1969) adopted a broader species concept of H. sinuosa and synonymized all the known species of Histiodella at the time, including H. altifrons Harris, Reference Harris1962, H. serrata Harris, Reference Harris1962, H. minutiserrata Mound, Reference Mound1965, and H. triquetra Mound, Reference Mound1965.
McHargue (Reference McHargue1975, Reference McHargue1982) was the first to apply a multielement species concept to the species of Histiodella based on the study of a large collection from the Joins Formation in the Arbuckle Mountains, southern Oklahoma. Although Pb (= short bryantodontiform of McHargue, Reference McHargue1982), ramiform S (including Sa, Sb, and Sc = trichonodelliform, zygognathiform, and twisted bryantodontiform, respectively, of McHargue, Reference McHargue1982) and geniculate makellate M (= oistodontiform of McHargue, Reference McHargue1982) elements were recognized to form the species apparatuses (McHargue, Reference McHargue1982, text-fig. 2), the seximembrate species concept and distinction among the four species (H. altifrons, H. minutiserrata, H. serrata, and H. sinuosa) recovered from the Joins Formation were mainly based on the protocarminate or carminate Pa elements (= bryantodontiform of McHargue, Reference McHargue1982) of these species, because the S elements were much less frequently represented in the Joins Formation samples and were difficult to assign confidently to species based on their morphology and known stratigraphic range (McHargue, Reference McHargue1982, table 1).
Table 1. Data matrix for the 10 species of Histiodella and outgroup, Cooperignathus nyinti (Cooper, Reference Cooper1981) (composite coding, with 12 binary characters, N = inapplicable). See text for descriptions of characters 1–12.

Ethington and Clark (Reference Ethington and Clark1982) accepted the species concept proposed by McHargue (Reference McHargue1982) and recovered H. altifrons, H. minutiserrata, and H. sinuosa from the Kanosh Shale, and a new species, H. holodentata Ethington and Clark, Reference Ethington and Clark1982, from the overlying Lehman Formation of Utah. Similarly, the 516 original type specimens defining H. holodentata from the Ibex area belong to the carminate Pa element, except for a single ramiform element assignable to the triform Sa element. Subsequently, three species of Histiodella, including H. bellburnensis Stouge, Reference Stouge1984, H. kristinae Stouge, Reference Stouge1984, and H. tableheadensis Stouge, Reference Stouge1984 (a junior synonym of H. holodentata) were erected based on the material from the Table Head Group of Newfoundland. Stouge (Reference Stouge1984, p. 87) suggested that the apparatuses of the Histiodella species from the Table Head Group consisted of six morphologically distinctive elements, but the Pb (= ozarkodiniform element of Stouge, Reference Stouge1984), geniculate M, and ramiform S elements were relatively rare, smaller in size, and were neither adequately described nor illustrated. Among these three species recognized in the Table Head Group, H. bellburnensis is represented by only the carminate Pa element (Stouge, Reference Stouge1984, p. 87, pl. 17, figs. 20, 21), H. tableheadensis by the carminate Pa (Stouge, Reference Stouge1984, p. 87, 88, pl. 18, figs. 8, 12–14) and unillustrated geniculate M and ramiform S elements, and H. kristinae by the carminate Pa (Stouge, Reference Stouge1984, p. 87, pl. 18, figs. 2, 3, 6, 7), geniculate M (Stouge, Reference Stouge1984, p. 87, pl. 18, figs. 1, 4), and ramiform S (Stouge, Reference Stouge1984, p. 87, pl. 18, figs. 5, 9–11) elements.
Several species of Histiodella were also erected from the middle Darriwilian of North China, but they were based solely on the carminate Pa elements. Histiodella triangularis (Wang and Lou, Reference Wang and Lou1984) and H. wuhaiensis Wang, Bergström, Zhen, Zhang, Wu, and Chen, Reference Wang, Bergström, Zhen, Zhang, Wu and Chen2013, were characterized by having an inconspicuous cusp and are considered as valid, showing closer phylogenetic ties with H. bellburnensis. Histiodella infrequensa An in An et al., Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and Yang1983, from the Beianzhuang Formation of North China is considered as a junior synonym of H. holodentata, and H. intertexa An in An et al., Reference An, Du, Gao, Chen and Li1981 (illustrated only as a nomen nudum in An et al., Reference An, Du, Gao, Chen and Li1981, Reference An, Du and Gao1985, and formally defined in An, Reference An1987) from the Kuniutian Formation in South China as a junior synonym of H. kristinae. A likely new species recently reported from middle Darriwilian cherts in New South Wales and referred to as Histiodella sp. cf. H. kristinae is also solely based on the carminate Pa element, which is like that of H. kristinae, but with a more extended anterior process (Zhen and Rutledge, Reference Zhen and Rutledge2020, fig. 10g–i; Zhen et al., Reference Zhen, Percival, Gilmore, Rutledge and Deyssing2021b, fig. 5s).
It is now generally agreed that Histiodella species have a seximembrate or septimembrate apparatus, a view supported by recovery of Pb, M, and S elements in association with various species of Histiodella, as discussed above (e.g., Ethington and Clark, Reference Ethington and Clark1982; McHargue, Reference McHargue1982; Stouge, Reference Stouge1984, Reference Stouge2012; Sweet, Reference Sweet1988; Zhen and Percival, Reference Zhen and Percival2004; Du et al., Reference Du, Zhao, Huang, Tan, Wang, Yang, Zhang and Xiao2005; Bauer, Reference Bauer2010; Feltes et al., Reference Feltes, Albanesi and Bergström2016). However, their morphologic relationships within an apparatus, particularly between the rare and delicate ramiform S elements and the constituent Pa element, are still poorly understood and often variably interpreted. Therefore, differentiation of a Histiodella species is practically still based mainly, if not solely, on the Pa element.
Histiodella donnae Repetski, Reference Repetski1982, from the lower part (upper Tremadocian) of the El Paso Group of westernmost Texas, is represented by a symmetrical element (figured paratype, Repetski, Reference Repetski1982, pl. 8, fig. 7; interpreted herein as representing the Sa element) and an asymmetrical element (holotype, Repetski, Reference Repetski1982, pl. 8, fig. 6; interpreted herein as representing the Sb element). Two additional elements belonging to this species subsequently were illustrated by Repetski et al. (1998, fig. 2I–L), including a geniculate M (Repetski et al., Reference Repetski, Loch, Ethington, Santucci and McClelland1998, fig. 2J) and a laterally compressed coniform Sc (Repetski et al., Reference Repetski, Loch, Ethington, Santucci and McClelland1998, fig. 2L). A posteriorly curved cusp and a mid-carina on the broadly concave posterior face of the Sa and Sb elements of H. donnae suggest a closer phylogenetic relationship with Rossodus Repetski and Ethington, Reference Repetski and Ethington1983. However, because the P elements have not been recovered for H. donnae, the morphological and phylogenetic relationship between these leaf-shaped blades of H. donnae and Histiodella sensu stricto from the Middle Ordovician remains unclear. Therefore, H. donnae is excluded herein from Histiodella sensu stricto. Histiodella angulata Moskalenko, Reference Moskalenko and Sokolov1982 (see Moskalenko, Reference Moskalenko1983; Tolmacheva et al., Reference Tolmacheva, Dronov, Alekseev, Danukalova, Larionov, Obut, Sennikov and Kipriyanova2019), reported from lower Darriwilian of the Siberian Platform, is considered herein as a possible species of Jumudontus Cooper, Reference Cooper1981. Histiodella levis Tolmacheva, Reference Tolmacheva2014, from the lower Darriwilian of central Asia is an adentate species consisting of a seximembrate apparatus that is comparable with that of H. altifrons, but the Pa element of H. levis (see Tolmacheva, Reference Tolmacheva2014, pl. 22, figs. 1–9, 18) has a much shorter anterior protoprocess, a more open basal cavity, a prominent platform-like projection on the outer-lateral side, and surface granular ornamentation on the basal flange and basal surface, which resemble typical Jumudontus species. More specifically, the strong resemblance between the granular ornamentation (Tolmacheva, Reference Tolmacheva2014, pl. 22, fig. 18) of H. levis and the small pustules developed on the P elements of Jumudontus gananda, as documented by Nicoll (Reference Nicoll1992), suggests a closer relationship of these species.
Species assigned to Histiodella
Given the remarks above, only the 10 named species discussed below are confirmatively included in Histiodella.
Histiodella altifrons Harris, Reference Harris1962.—Middle Ordovician, holotype (OMC 115, figured by Harris, Reference Harris1962, pl. 1, fig. 4) from the horizon 11.89 m (39 feet) above the base of the Joins Formation at the Simpson section located along the west side of U.S. Highway 77, on the south side of Arbuckle Mountains, Oklahoma (= Histiodella triquetra Mound, Reference Mound1965).
Histiodella bellburnensis Stouge, Reference Stouge1984
Middle Darriwilian, holotype (ROM 39716, figured by Stouge, pl. 17, fig. 20) from the Middle Member of the Table Head Group exposed at Table Point of the Great Northern Peninsula, Newfoundland.
Histiodella holodentata Ethington and Clark, Reference Ethington and Clark1982.—Middle Darriwilian, holotype (UMC 1094-2, figured by Ethington and Clark, Reference Ethington and Clark1982, pl. 4, fig. 3) from the Lehman Formation in the Ibex area of western Utah (= Histiodella infrequensa An in An et al., Reference An, Zhang, Xiang, Zhang, Xu, Zhang, Jiang, Yang, Lin, Cui and Yang1983; = Histiodella tableheadensis Stouge, Reference Stouge1984). A morphologically transitional form between H. holodentata and H. kristinae is characterized by having the tips of the cusp and the tallest denticle on the anterior process at more or less the same height. This form was treated by Stouge (Reference Stouge2012) as a separate species, referred to as Histiodella cf. holodentata, while other authors (e.g., Zhen, Reference Zhen2020; Zhen et al., Reference Zhen, Normore, Dent and Percival2020a) considered it as representing an advanced variant of H. holodentata.
Histiodella kristinae Stouge, Reference Stouge1984
Middle Darriwilian, holotype (ROM 39718, figured by Stouge, pl. 18, fig. 3) from the Middle Member of the Table Head Group exposed at Table Point of the Great Northern Peninsula, Newfoundland (= H. intertexa An in An et al., Reference An, Du, Gao, Chen and Li1981, which was introduced as a nomen nudum).
Histiodella labiosa Bauer, Reference Bauer2010
Early Darriwilian, lectotype (hereby specimen OSU 52034, originally figured by Bauer, Reference Bauer2010, pl. 2, fig. 13, is designated as lectotype) from sample 72SC-500 at the horizon 152.4 m above the base of the Oil Creek Formation in section 72SC exposed in southern Oklahoma.
Histiodella minutiserrata Mound, Reference Mound1965
Middle Ordovician, holotype (USNM 146971, figured by Mound, Reference Mound1965, pl. 3, fig. 2) from the Joins Formation at the Simpson section (same locality and section of Harris, Reference Harris1962, as indicated by Mound, Reference Mound1965, p. 2) located along the west side of U.S. Highway 77, on the south side of the Arbuckle Mountains, Oklahoma.
Histiodella serrata Harris, Reference Harris1962
Middle Ordovician, holotype (OMC 118, figured by Harris, Reference Harris1962, pl. 1, fig. 3) from the horizon 57 m (187 feet) above the base of the Joins Formation at the Simpson section, located along the west side of U.S. Highway 77, on the south side of the Arbuckle Mountains, Oklahoma (= ?Histiodella triquetra Mound, Reference Mound1965, which might represent the Sa element of H. serrata).
Histiodella sinuosa (Graves and Ellison, Reference Graves and Ellison1941)
Middle Ordovician, holotype (MSM 7125, G414, figured by Graves and Ellison, Reference Graves and Ellison1941, pl. 2, fig. 13) from the Fort Peña Formation exposed on east side of road in bed of Alsate Creek, six miles southwest of Marathon, Texas.
Histiodella triangularis (Wang and Lou, Reference Wang and Lou1984)
Middle Darriwilian, holotype (NIGP 52199, figured by Wang and Lou, Reference Wang and Lou1984, pl. 12, fig. 17) from the Zotzeshan Formation of Inner Mongolia, North China.
Histiodella wuhaiensis Wang, Bergström, Zhen, Zhang, Wu, and Chen, Reference Wang, Bergström, Zhen, Zhang, Wu and Chen2013
Middle Darriwilian, holotype (NIGP 156446, figured by Wang et al., Reference Wang, Bergström, Zhen, Zhang, Wu and Chen2013, pl. 2, fig. 8) from the lower Klimoli Formation of Inner Mongolia, North China.
Histiodella labiosa Bauer, Reference Bauer2010
Figures 3.1, 3.2, 4–6
- ?Reference Bergström1979
Histiodella n. sp. Bergström, fig. 2C.
- Reference Harris, Bergström, Ethington and Ross1979
Histiodella n. sp. 2 Harris et al., pl. 4, figs. 12, 13.
- Reference Bauer2010
Histiodella labiosa Bauer, p. 13, pl. 2, figs. 10, 11, 13, 14 (see for synonymy).
- ?Reference Stouge2012
Histiodella holodentata; Stouge, part, fig. 9AA.
- Reference Zhen2020
Histiodella labiosa; Zhen and Rutledge, fig. 8a–c.
- Reference Zhen2021
Histiodella labiosa; Zhen, fig. 5n.
- Reference Zhen, Normore, Dent and Percival2021a
Histiodella labiosa; Zhen et al., p. 273, fig. 12M.
- Reference Zhen, Zhang, Harper, Zhan, Fang, Wang, Yu and Li2021b
Histiodella labiosa; Zhen et al., fig. 5p.
Type series
Lectotype (designated herein OSU 52034, originally figured by Bauer, Reference Bauer2010, pl. 2, fig. 13) and paralectotypes (OSU 52031 = M from sample 72SC-500; OSU 52032 = Sa from sample 72SC-470; OSU 52035 = Pb from sample 72SC-620, originally figured by Bauer, Reference Bauer2010, pl. 2, figs. 10, 11, 14, respectively).
Diagnosis
Species of Histiodella consisting of carminate Pa and Pb elements, geniculate makellate M element, and ramiform S elements; Pa and Pb elements triangular in outline with a prominent cusp, a longer anterior process and with well-defined basal flange; Pa element with a narrow platform-like projection developed near mid-length on the outer-lateral side, and confluent denticles on anterior and posterior processes; axes of the denticles on both processes parallel or sub-parallel to the axis of cusp; Pb element having a denticulate anterior process and a shorter posterior protoprocess with minutely serrated upper margin and a weakly developed platform-like projection on the outer-lateral side; M element typically with short denticles along inner-lateral margin (modified after Bauer, Reference Bauer2010, p. 13).
Description
Species consisting of likely seximembrate apparatus of denticulate elements, including geniculate and typically denticulate makellate M, ramiform S (alate triform Sa, modified bipennate Sc, and possibly also including a modified tertiopedate Sb) and carminate Pa and Pb (both with a well-developed basal flange) elements. Carminate Pa element triangular in outline (H/L ratio varying from 0.52–0.86) with a prominent cusp and the denticulate anterior and posterior processes triangular in outline (Fig. 4); cusp suberect to weakly reclined, forming the apex of the element, laterally compressed and biconvex with sharp anterior and posterior edges, and width of three to four times as wide as the adjacent denticles in lateral view; anterior process longer, bearing 7–10 long, confluent denticles in full-growth specimens; posterior process slightly shorter, bearing 5–9 long, confluent denticles; cusp and denticles ornamented with fine striae; denticles on the two processes highest next to the cusp and progressively shorter distally; axes of denticles extending parallel or sub-parallel to the axis of cusp (more or less perpendicular to basal margin); platform-like projection prominent on the outer-lateral side, triangular in outline in apical view (Fig. 4.2, 4.8, 4.12); basal cavity shallow, slightly expanded underneath cusp and extending as a narrow groove to the distal end of the processes (Fig. 4.7).
Carminate Pb element shorter triangular in outline and shorter in length compared to the Pa (H/L ratio varying from 0.74–0.96) with a prominent cusp, a denticulate anterior process, and a shorter posterior protoprocess (Figs. 3.2, 5); cusp suberect to weakly reclined to form apex of the element, laterally compressed and biconvex with sharp anterior and posterior edges, and with a width of three to four times that of the adjacent denticles on the anterior process in lateral view; anterior process longer with a slightly convexly curved upper margin, bearing 4–9 long, confluent denticles that are highest next to the cusp and progressively shorter towards distal end; axes of denticles extending parallel or sub-parallel to axis of cusp; posterior margin of the cusp and upper margin of the posterior protoprocess extending smoothly into each other, straight (Fig. 5.5, 5.7) to slightly concavely curved (Fig. 5.6, 5.10) and minutely serrated in lateral view (Fig. 5.4); surface of cusp and denticles ornamented with fine striae; platform-like projection weakly developed on the outer-lateral side (Fig. 5.8).
Geniculate M element antero-posteriorly compressed with a robust and outer-laterally recurved cusp and a low, short base (Figs. 3.1, 6.2–6.9); cusp short and slightly bowed posteriorly, biconvex with sharp outer-lateral and inner-lateral margins, a more broadly convex anterior face and a broad carina on the posterior face (Fig. 6.2, 6.3, 6.5); inner-lateral margin gently curved and typically ornamented with 3–8 short denticles, less commonly denticles may be rudimentary (Bauer, Reference Bauer2010, pl. 2, fig. 10) or even absent; base expanding posteriorly to form a triangular outline in apical or basal view (Fig. 6.4) with a basal cavity of moderate depth (Fig. 6.2).
Ramiform S elements represented by alate triform Sa and modified bipennate Sc elements; Sa element bearing a denticulate blade-like lateral process on each side and sharp blade-like costa on the posterior face (Bauer, Reference Bauer2010, pl. 2, fig. 11); Sc element strongly compressed laterally and asymmetrical with a prominent cusp and an anterior process bearing five denticles (Fig. 6.1); cusp biconvex with sharp anterior and posterior margins.
Material illustrated
Thirty topotypes (OSU 54951–54980), including Pa (11 specimens, Fig. 4) and Pb (10 specimens, Fig. 5), M (eight specimens, Fig. 6.2–6.9), and Sc (one specimen, Fig. 6.1) elements from the Oil Creek Formation, section 72SC, southern Oklahoma.
Remarks
Bauer (Reference Bauer2010) recognized carminate Pa and Pb elements, geniculate M, and ramiform Sa and Sb elements, but S elements are extremely rare with only eight specimens represented among a total of 997 specimens in his original study of H. labiosa. Reexamination of the current topotype material from sample 89JA-178 produced a single specimen representing the modified bipennate Sc (Fig. 6.1) of this species, supporting a possible seximembrate species apparatus for H. labiosa. However, Bauer (Reference Bauer2010) included and described the Sb element as like alate Sa but with asymmetrically disposed lateral processes, but it was neither illustrated by him from the original type material nor recovered from the current samples of this study.
The Pa element of H. labiosa morphologically resembles that of H. serrata and H. holodentata, but differs from the latter two species by having a longer posterior process and by having denticles on the posterior process extending with their axes parallel or nearly parallel to the axis of the cusp. Bauer (Reference Bauer2010) also noted the development of a narrow platform on the outer-lateral side of the Pa (Fig. 4.2, 4.8, 4.12) and Pb (Fig. 5.8) elements of H. labiosa—a distinctive character that has not been reported occurring in any other species of Histiodella.
Phylogeny and evolution of Histiodella
Although many species of Histiodella are cosmopolitan and widely used as biozonal index species for worldwide correlation of the upper Dapingian and lower to middle Darriwilian strata, their origin and evolution are still poorly understood. By including H. donnae as the most primitive species of Histiodella, Repetski and Repetski (Reference Repetski and Repetski2002) suggested that Histiodella might be directly evolved from a Rossodus species. Variable views exist to suggest that Histiodella evolved from an ancestral stock within Oistodontidae, but relating to different genera, including Rossodus, Juanognathus, Tripodus, Protoprioniodus, and Cooperignathus, by several authors (e.g., Ethington and Clark, Reference Ethington and Clark1982; Dzik, Reference Dzik1983; Sweet, Reference Sweet1988; Repetski and Repetski, Reference Repetski and Repetski2002). Among these taxa considered to be the likely ancestor of Histiodella, only Protoprioniodus and Cooperignathus have protopectiniform P elements in their species apparatuses, which share a number of the characters with the P elements of Histiodella species. To test these hypotheses, a phylogenetic analysis was conducted in this study by selecting Cooperignathus nyinti as the outgroup, which resulted in a well-resolved hierarchical clustering that infers the evolutionary relationship of all the 10 confirmed species of Histiodella (Figs. 7–9). Rossodus, Juanognathus, and Histiodella donnae are less closely related to Histiodella. Having coniform P elements (in Rossodus) or lacking P elements (in Juanognathus and Histiodella donnae) in their species reconstructions prevents a direct morphological comparison with the protopectiniform or pectiniform Pa elements of the Histiodella species. Tripodus species have the protopastinate P elements in their species apparatuses (Sweet, Reference Sweet1988, fig. 5.12), which suggests a closer affinity with Prioniodontidae. Therefore, these taxa have been excluded from the analysis conducted herein.
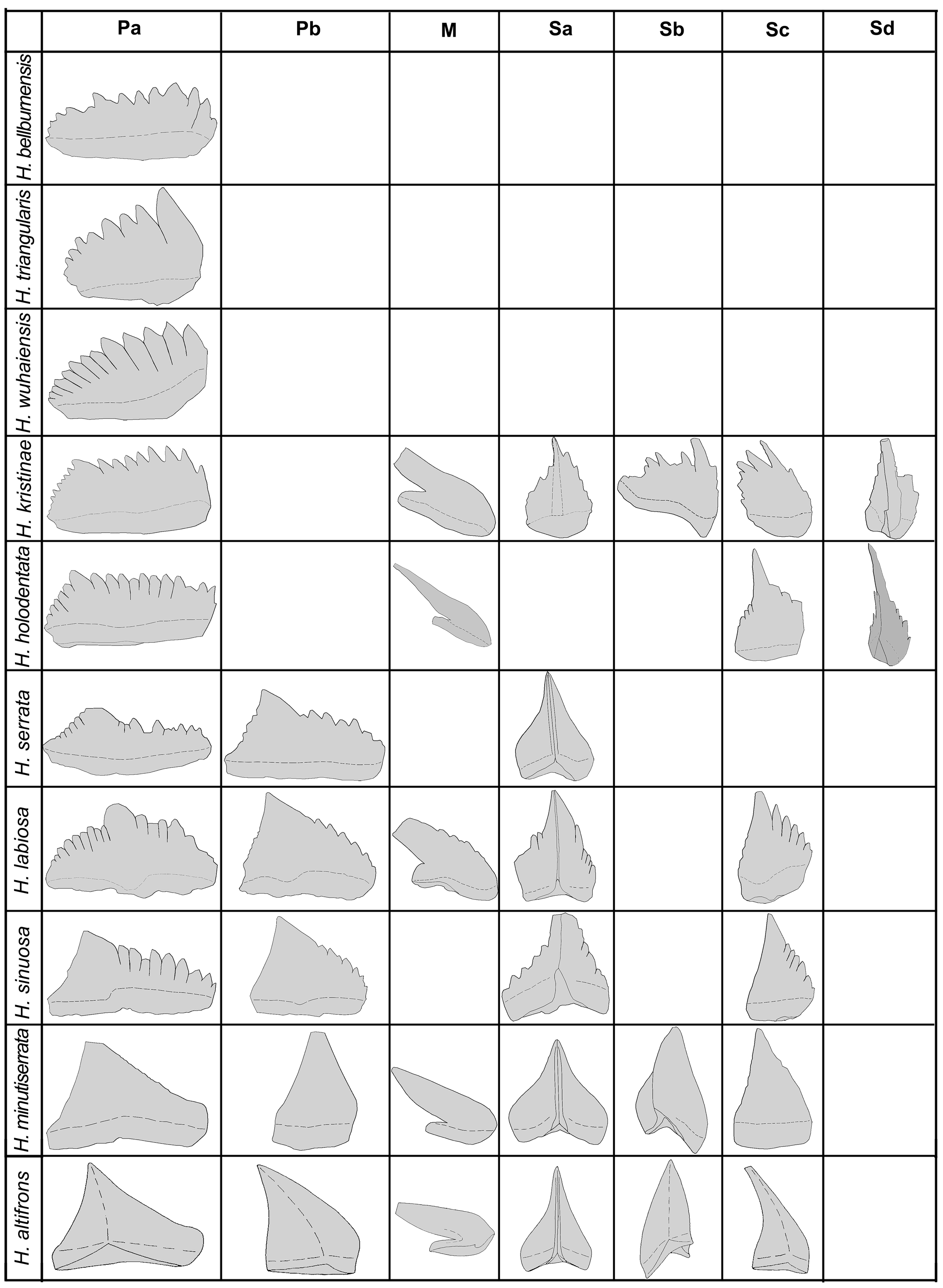
Figure 7. Species apparatus and comparison of the constituent elements of the 10 Histiodella species (sources of data from Harris, Reference Harris1962; Mound, Reference Mound1965; Ethington and Clark, Reference Ethington and Clark1982; McHargue, Reference McHargue1982; Stouge, Reference Stouge1984, Reference Stouge2012; Wang and Lou, Reference Wang and Lou1984; Bauer, Reference Bauer2010; Wang et al., Reference Wang, Bergström, Zhen, Zhang, Wu and Chen2013; this study).
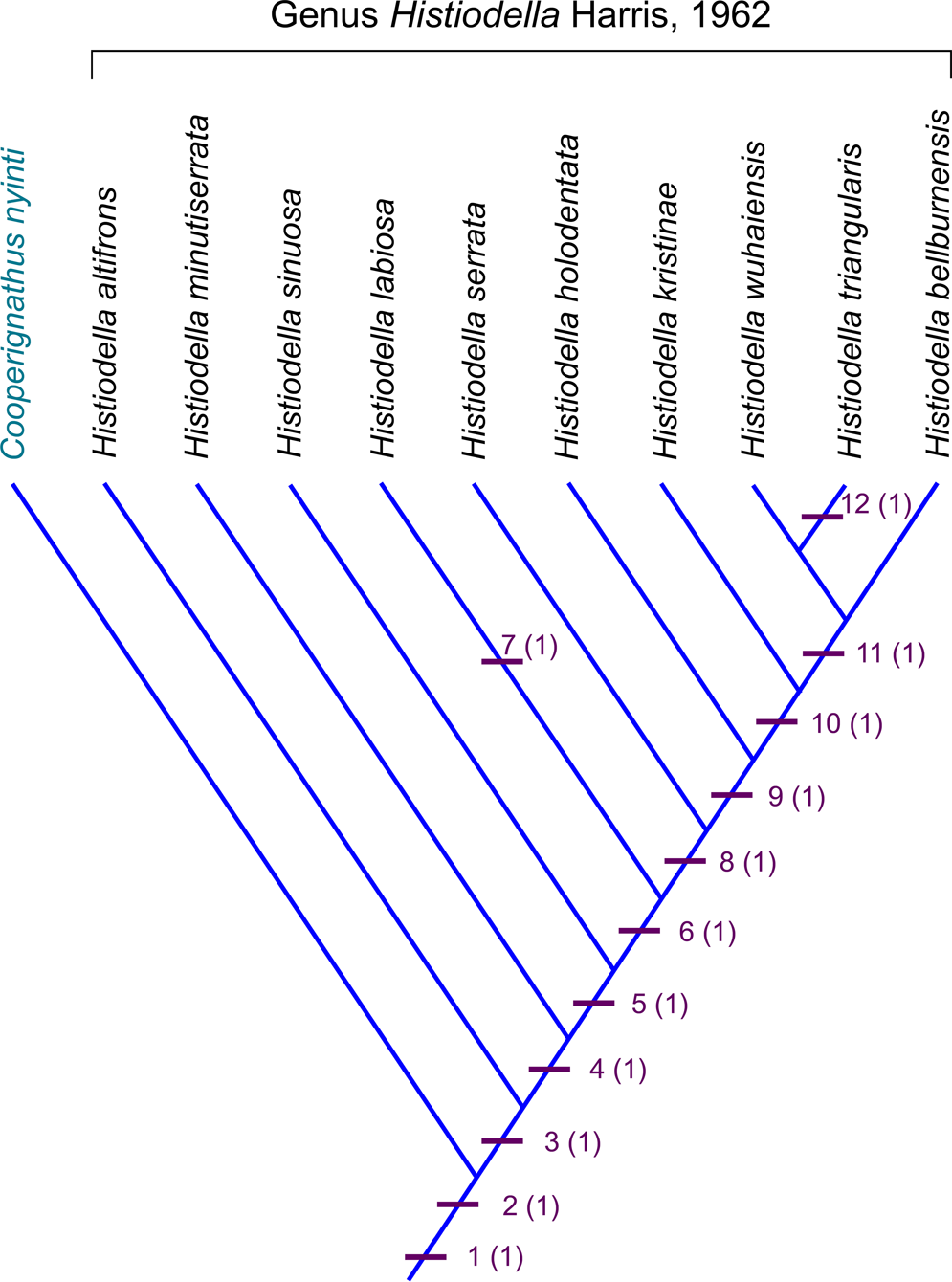
Figure 8. Cladogram showing the inferred phylogenetic relationships of the 10 Histiodella species by using data analysis program PAST (as unweighted and unordered; Algorithm: Branch-and-bound; Optimization: Wagner) with Cooperignathus nyinti (Cooper, Reference Cooper1981) as the outgroup; dataset coded with 12 binary characters of the Pa elements (see text for description).

Figure 9. Stratigraphic ranges of the 10 Histiodella species and their inferred species evolution. Up. = Upper Ordovician; Lower = Lower Ordovician.
Our analysis shows a closer relationship between Histiodella and Cooperignathus of the Early Ordovician. The protoanguliplanate P elements of Cooperignathus (Fig. 3.4) may represent the earliest geological record of true pectiniform elements that have several shared synapomorphies with Histiodella species, including the occurrence of a basal flange and basal surface (Fig. 7). Both Cooperignathus nyinti and C. aranda also have a narrow platform-like projection on the outer-lateral face of the P elements (Zhen et al., Reference Zhen, Percival and Webby2003, Reference Zhen, Laurie, Percival, Nicoll and Cooperin press), which is comparable with that developed on the outer-lateral face of the Pa element of H. labiosa (Fig. 4.2, 4.8, 4.12).
Based on the morphometric analysis of the Histiodella species recovered from the Joins Formation of the Arbuckle Mountains of Oklahoma, McHargue (Reference McHargue1982) established two evolutionary trends, namely (1) evolving from adentate early species (H. altifrons) to partially denticulate (e.g., H. sinuosa with only anterior process) and then to fully denticulate (e.g., H. serrata and H. labiosa with anterior and posterior processes), and (2) from a relatively high blade morphology in the early species (e.g., H. altifrons and H. minutiserrata) to more elongated shape of the late species (e.g., H. bellburnensis) by decreasing height/length ratio of the Pa elements. Our current study of the morphology and their stratigraphic ranges of the 10 Histiodella species also shows that the general shape of their Pa elements has evolved from a triangular outline of the early species (e.g., H. altifrons, H. minutiserrata, H. sinuosa, and H. labiosa) to a more or less rectangular form of the late species (e.g., H. kristinae and H. bellburnensis; Fig. 7). The apex of the Pa elements also changed from a location towards posterior in the early species (e.g., H. altifrons and H. minutiserrata) to more or less centrally positioned (H. labiosa) and then to anteriorly positioned in the late species (e.g., H. kristinae and H. bellburnensis). The cusp evolved from robust and high in the early species to inconspicuous in the late species (Fig. 7).
Cladistic analysis
To test the validity of these inferred phylogenetic relationships, the 10 species of Histiodella, plus Cooperignathus nyinti acting as the outgroup, were scored using the following 12 binary characters of the Pa elements and were then cladistically analyzed by applying reductive coding strategy (Table 1).
Character 1.—Basal flange and basal surface: absent (0), present (1). All 10 species of Histiodella and the out-group are coded as (1).
Character 2.—Protopectiniform or pectiniform: absent (0), present (1). All 10 species of Histiodella and the out-group are coded as (1).
Character 3.—Presence of cusp: absent (0), present (1). This is a character shared by all 10 species of Histiodella, with C. nyinti coded as (0).
Character 4.—Denticulated or serrated: absent (0), present (1). This is a character shared by nine species of Histiodella that are coded as (1), with H. altifrons and C. nyinti coded as (0).
Character 5.—Denticles on anterior process: absent (0), present (1). This is a character shared by eight species of Histiodella that are coded as (1), with H. minutiserrata, H. altifrons, and C. nyinti coded as (0).
Character 6.—Denticle on posterior process: absent (0), present (1). This character is shared by seven species of Histiodella that are coded as (1), with H. sinuosa, H. minutiserrata, H. altifrons, and C. nyinti coded as (0).
Character 7.—Axes of denticles on posterior process parallel to axis of the cusp: absent (0), present (1). This character features H. labiosa coded as (1). It is inapplicable for H. minutiserrata, H. altifrons, and C. nyinti, which are coded as (N). Others are coded as (0).
Character 8.—Reclined denticles on posterior process: absent (0), present (1). This character is shared by six species of Histiodella that are coded as (1), with H. labiosa coded as (0). It is inapplicable for H. sinuosa, H. minutiserrata, H. altifrons, and C. nyinti, which are coded as (N).
Character 9.—Rectangular outline in lateral view: absent (0), present (1). This character is shared by five species of Histiodella that are coded as (1), with the other five species of Histiodella coded as (0). It is inapplicable for C. nyinti, which is coded as (N).
Character 10.—Apex of the unit is located toward anterior end: absent (0), present (1). This character is shared by four species of Histiodella that are coded as (1), with the other six species of Histiodella coded as (0). It is inapplicable for C. nyinti, which is coded as (N).
Character 11.—Small or inconspicuous cusp: absent (0), present (1). This character is shared by three species of Histiodella that are coded as (1), with the other seven species of Histiodella coded as (0). It is inapplicable for C. nyinti, which is coded as (N).
Character 12.—Strongly reduced anterior process: absent (0), present (1). This character features in H. triangularis, which is coded as (1), with other species of Histiodella coded as (0). It is inapplicable for C. nyinti, which is coded as (N).
Histiodella cladogram
The cladogram produced is highly resolved (Fig. 8). It depicts the origin of this biostratigraphically valuable genus in the Middle Ordovician and its close phylogenetic affinity with late Floian (Early Ordovician) Cooperignathus, suggesting that both genera might share a common adentate ancestor (within Oistodontidae). It also supports the monophyletic grouping of the 10 Histiodella species and the evolutionary trends that have been revealed from the analyses of morphological changes and morphometrics among these species (e.g., McHargue, Reference McHargue1982; Feltes and Albanesi, Reference Feltes, Albanesi, Liao and Valenzuela-Rios2017). Our study is consistent with the results derived from the geometric morphometric analysis of five Histiodella species conducted by Feltes and Albanesi (Reference Feltes, Albanesi, Liao and Valenzuela-Rios2017), demonstrating that the Histiodella species evolved through continuous changes in shape of the element outlines and size and height of denticles of the Pa elements.
Distribution and biostratigraphy
With their distinctive morphology and short stratigraphic ranges, Histiodella species are recognized as excellent markers in subdivision and correlation of the lower and middle Darriwilian strata (Fig. 9). Occurrence of the six Histiodella species in the Joins and Oil Creek formations exposed in southern Oklahoma represents the best record of these species worldwide. Five conodont biozones are defined, with the H. altifrons Biozone recognized in the lower part of the Joins Formation (Figs. 2, 9). The succeeding H. minutiserrata Biozone is marked by the FAD (First Appearance Datum) of the eponymous species as the base and the FAD of H. sinuosa as the top (Figs. 2, 9). As the primitive forms of Histiodella, H. altifrons is the only adentate species and H. minutiserrata, which is characterized by having minutely serrated upper margin of the Pa element, might be directly evolved from it. Apart from their wide distribution in North America (e.g., Harris, Reference Harris1962; Mound, Reference Mound1965; Bradshaw, Reference Bradshaw1969; Harris et al., Reference Harris, Bergström, Ethington and Ross1979; Ethington and Clark, Reference Ethington and Clark1982; McHargue, Reference McHargue1982; Norford et al., Reference Norford, Jackson and Nowlan2002; Bauer, Reference Bauer2010), these two species are generally found as minor faunal components in the Argentine Precordillera (Lehnert, Reference Lehnert1995; Albanesi et al., Reference Albanesi, Carrera, Cañas and Saltzman2006; Voldman et al., Reference Voldman, Albanesi, Alonso, Fernández, Banchig, Cardó, Ortega and Vallaure2015), central Asian Fold Belt (Tolmacheva, Reference Tolmacheva2014, referred to as Histiodella сf. H. altifrons and Histiodella sp. 1, respectively), and Australia (Nicoll, Reference Nicoll1992, Reference Nicoll1993; Zhen et al., Reference Zhen, Normore, Dent and Percival2020a, Reference Zhen, Zhang, Harper, Zhan, Fang, Wang, Yu and Lib). The eponymous species of the H. sinuosa Biozone is the oldest species of Histiodella bearing true denticles (but only on the anterior process, Fig. 7). This biozone is utilized to correlate lower Darriwilian strata in Laurentia (Fig. 9), and is recognized in the Argentine Precordillera (e.g., Carrera et al., Reference Carrera, Fenoglio, Albanesi, Voldman, Albanesi and Ortega2013; Serra et al., Reference Serra, Albanesi, Ortega and Bergström2015, Reference Serra, Feltes, Albanesi and Goldman2019, Reference Serra, Feltes, Mango, Henderson, Albanesi and Ortega2020; Feltes et al., Reference Feltes, Albanesi and Bergström2016) and central Asian Fold Belt (Tolmacheva, Reference Tolmacheva2014).
Bauer (Reference Bauer2010) established the H. labiosa Biozone based on the appearance of H. labiosa and its stratigraphic range in the Oil Creek Formation within the section 72SC in southern Oklahoma. It was confined by the FAD of the eponymous species as the base and the FAD of H. holodentata as the top. Histiodella labiosa first occurs in sample 72SC-40 at a horizon 12.19 m above the base of the Oil Creek Formation (Bauer, Reference Bauer2010, table 1) and extends to the top of the Oil Creek Formation. Histiodella holodentata is very rare in the Oil Creek Formation at section 72SC. Bauer (Reference Bauer2010, pl. 2, fig. 9; Fig. 2) illustrated a specimen representing the Pa element of H. holodentata recovered from sample 72SC-620 (at a horizon 188.98 m above the base of the Oil Creek Formation). Two additional specimens representing the Pa element of this species were also recovered from sample 72SC-440 (at a horizon 134.11 m above the base of the Oil Creek Formation). Therefore, the boundary between the H. holodentata/H. labiosa biozones in section 72SC is drawn in the middle of the Oil Creek Formation, 134.11 m above the base of the Oil Creek Formation (Fig. 2). Abundant H. labiosa and rare H. holodentata in the upper Oil Creek Formation suggested that two species might have favored different habitats. Distribution of H. labiosa in North America was detailed by Bauer (Reference Bauer2010), and the synonym list in this study also includes a recent report of this species from the top part of the Willara Formation in the Canning Basin of Western Australia (Zhen et al., Reference Zhen, Nicoll, Normore, Percival, Laurie and Dent2021a) and from the chert beds within the Abercrombie Formation of the turbiditic sequences in New South Wales (Zhen and Rutledge, Reference Zhen and Rutledge2020; Zhen, Reference Zhen2021; Zhen et al., Reference Zhen, Percival, Gilmore, Rutledge and Deyssing2021b). Occurrence of this species from two separate paleobiogeographic domains in the Ordovician tropical zone (Laurentia and eastern Gondwana) indicates that H. labiosa may have a wide geographical distribution and be potentially useful for correlation of the lower Darriwilian Stage worldwide.
Histiodella serrata, H. holodentata, and H. kristinae are cosmopolitan, with wider distribution geographically than other species of Histiodella, and have played an important role in the detailed biostratigraphic correlation of early and middle Darriwilian worldwide (Bergström and Ferretti, Reference Bergström and Ferretti2017; Goldman et al., Reference Goldman, Sadler, Leslie, Gradstein, Ogg, Schmitz and Ogg2020; Zhen, Reference Zhen2021). Continuous succession consisting of the H. holodentata, H. kristinae, and H. bellburnensis biozones were well recorded from the Table Head Group of Newfoundland (Stouge, Reference Stouge1984, Reference Stouge2012; Fig. 9). Apart from their wide occurrence in North America, H. holodentata and H. kristinae also occur in the Argentine Precordillera (e.g., Albanesi, Reference Albanesi1998; Albanesi and Ortega, Reference Albanesi, Ortega and Aceñolaza2003, Reference Albanesi, Ortega and Montenari2016; Mestre, Reference Mestre2012; Serra et al., Reference Serra, Albanesi, Ortega and Bergström2015, Reference Serra, Feltes, Henderson and Albanesi2017, Reference Serra, Feltes, Albanesi and Goldman2019, Reference Serra, Feltes, Mango, Henderson, Albanesi and Ortega2020; Feltes et al., Reference Feltes, Albanesi and Bergström2016), Australasia (e.g., Zhen and Percival, Reference Zhen and Percival2004; Zhen et al., Reference Zhen, Percival, Cooper, Simes and Wright2009, Reference Zhen, Normore, Dent and Percival2020a, Reference Zhen, Zhang, Harper, Zhan, Fang, Wang, Yu and Li2021b; Zhen and Rutledge, Reference Zhen and Rutledge2020), major Chinese terranes (e.g., An and Zheng, Reference An and Zheng1990; Du et al., Reference Du, Zhao, Huang, Tan, Wang, Yang, Zhang and Xiao2005; Wang et al., Reference Wang, Qi and Wu2011; Zhen et al., Reference Zhen, Wang, Zhang, Bergström, Percival and Chen2011, Reference Zhen, Zhang, Harper, Zhan, Fang, Wang, Yu and Li2020b), southern Asia (Agematsu et al., Reference Agematsu, Sashida, Salyapongse and Sardsud2006, Reference Agematsu, Sashida and Sardsud2008), the central Asian Fold Belt (e.g., Tolmacheva, Reference Tolmacheva2014), and Baltoscandia (e.g., Rasmussen, Reference Rasmussen2001; Löfgren, Reference Löfgren2004; Viira, Reference Viira2011). The H. holodentata and H. kristinae biozones of the middle Darriwilian (= Dw2 stage slice of Bergström et al., Reference Bergström, Chen, Gutiérrez-Marco and Dronov2009) now can be correlated precisely across shallow-water carbonate platform to deep-water slope settings, and well integrated with graptolite successions (Maletz, Reference Maletz2009; Stouge, Reference Stouge2012). Histiodella bellburnensis, representing one of the most derived forms of the genus, also has been reported from the Tarim Basin (Du et al., Reference Du, Zhao, Huang, Tan, Wang, Yang, Zhang and Xiao2005), North China (Jing et al., Reference Jing, Zhou, Wang, Yang, Fang, Wang and Fan2020), and the Argentine Precordillera (Serra et al., Reference Serra, Albanesi, Ortega and Bergström2015, Reference Serra, Feltes, Albanesi and Goldman2019). Histiodella triangularis and H. wuhaiensis, representing the two other most-derived forms, originally were reported from North China and recently were recorded from Australia (Zhen, Reference Zhen2021).
Conclusions
With formal designation of a lectotype, a revised diagnosis, detailed description, and morphological comparisons with its direct ancestor and descendant, Histiodella labiosa Bauer, Reference Bauer2010, is now accepted as a valid species, which is morphologically distinctive and biostratigraphically valuable for correlation of the lower Darriwilian Stage. Analysis of the morphological changes among the 10 known species of Histiodella has revealed that Histiodella species evolved through continuous changes in shape of element outlines, size and height of denticles, and cusp size and position of the Pa elements. The phylogenetic and morphometric analysis conducted in this study indicates that the 10 known species of Histiodella represent a monophyletic group, which is especially important to support the wide use of these species in subdividing and correlating late Dapingian to middle Darriwilian strata worldwide.
Acknowledgments
Research of the senior author was supported by the Geological Survey of New South Wales. P. Carter (Geological Survey of New South Wales) assisted with the final drafting of Figure 1. SEM images were prepared in the Electron Microscope Unit of Macquarie University with assistance from S. Lindsay. J. Repetski and G. Albanesi are thanked for their careful and constructive reviews of the manuscript. Y. Y. Zhen publishes with permission of the Executive Director, Geological Survey of New South Wales. This is a contribution to IGCP Project 735 (Rocks and the Rise of Ordovician Life) and IGCP Project 668 (Equatorial Gondwana History and Early Paleozoic Evolutionary Dynamics).
Disclosure statement
No potential conflict of interest was reported by the authors.










