

























Crossref Citations
This article has been cited by the following publications. This list is generated based on data provided by Crossref.
Krasnov, Boris R.
Morand, Serge
Hawlena, Hadas
Khokhlova, Irina S.
and
Shenbrot, Georgy I.
2005.
Sex-biased parasitism, seasonality and sexual size dimorphism in desert rodents.
Oecologia,
Vol. 146,
Issue. 2,
p.
209.
Bajer, A.
Harris, P. D.
Behnke, J. M.
Bednarska, M.
Barnard, C. J.
Sherif, N.
Clifford, S.
Gilbert, F. S.
Siński, E.
and
Zalat, S.
2006.
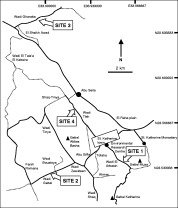
Local variation of haemoparasites and arthropod vectors, and intestinal protozoans in spiny mice (Acomys dimidiatus) from four montane wadis in the St Katherine Protectorate, Sinai, Egypt.
Journal of Zoology,
Vol. 270,
Issue. 1,
p.
9.
BORDES, F.
and
MORAND, S.
2008.
Helminth species diversity of mammals: parasite species richness is a host species attribute.
Parasitology,
Vol. 135,
Issue. 14,
p.
1701.
Miño, M.H.
2008.
Infection pattern of the spirurid nematodeProtospirura numidica criceticolain the cricetid rodentAkodon azaraeon poultry farms of central Argentina.
Journal of Helminthology,
Vol. 82,
Issue. 2,
p.
153.
BEHNKE, J. M.
2008.
Structure in parasite component communities in wild rodents: predictability, stability, associations and interactions .... or pure randomness?.
Parasitology,
Vol. 135,
Issue. 7,
p.
751.
BEHNKE, J. M.
BAJER, A.
HARRIS, P. D.
NEWINGTON, L.
PIDGEON, E.
ROWLANDS, G.
SHERIFF, C.
KULIŚ-MALKOWSKA, K.
SIŃSKI, E.
GILBERT, F. S.
and
BARNARD, C. J.
2008.
Temporal and between-site variation in helminth communities of bank voles (Myodes glareolus) from N.E. Poland. 1. Regional fauna and component community levels.
Parasitology,
Vol. 135,
Issue. 8,
p.
985.
BRADLEY, J. E.
and
JACKSON, J. A.
2008.
Measuring immune system variation to help understand host-pathogen community dynamics.
Parasitology,
Vol. 135,
Issue. 7,
p.
807.
BEHNKE, J. M.
BAJER, A.
HARRIS, P. D.
NEWINGTON, L.
PIDGEON, E.
ROWLANDS, G.
SHERIFF, C.
KULIŚ-MALKOWSKA, K.
SIńSKI, E.
GILBERT, F. S.
and
BARNARD, C. J.
2008.
Temporal and between-site variation in helminth communities of bank voles (Myodes glareolus) from N.E. Poland. 2. The infracommunity level.
Parasitology,
Vol. 135,
Issue. 8,
p.
999.
Smales, L. R.
Harris, P. D.
and
Behnke, J. M.
2009.
A redescription of Protospirura muricola Gedoelst, 1916 (Nematoda: Spiruridae), a parasite of murid rodents.
Systematic Parasitology,
Vol. 72,
Issue. 1,
p.
15.
Bordes, F.
and
Morand, S.
2009.
Parasite diversity: an overlooked metric of parasite pressures?.
Oikos,
Vol. 118,
Issue. 6,
p.
801.
Schuster, Rolf K.
Thomas, Katja
Sivakumar, Saritha
and
O’Donovan, Declan
2009.
The parasite fauna of stray domestic cats (Felis catus) in Dubai, United Arab Emirates.
Parasitology Research,
Vol. 105,
Issue. 1,
p.
125.
Abu-Madi, Marawan A.
Behnke, Jerzy M.
Prabhaker, K.S.
Al-Ibrahim, Roda
and
Lewis, John W.
2010.
Intestinal helminths of feral cat populations from urban and suburban districts of Qatar.
Veterinary Parasitology,
Vol. 168,
Issue. 3-4,
p.
284.
Simões, R.O.
Souza, J.G.R.
Maldonado, A.
and
Luque, J.L.
2011.
Variation in the helminth community structure of three sympatric sigmodontine rodents from the coastal Atlantic Forest of Rio de Janeiro, Brazil.
Journal of Helminthology,
Vol. 85,
Issue. 2,
p.
171.
Miño, M.H.
Rojas Herrera, E.J.
Notarnicola, J.
Robles, M. del R.
and
Navone, G.T.
2012.
Diversity of the helminth community of the Pampean grassland mouse (Akodon azarae) on poultry farms in central Argentina.
Journal of Helminthology,
Vol. 86,
Issue. 1,
p.
46.
Chesh, Adrian S.
Mabry, Karen E.
Keane, Brian
Noe, Douglas A.
and
Solomon, Nancy G.
2012.
Are body mass and parasite load related to social partnerships and mating inMicrotus ochrogaster?.
Journal of Mammalogy,
Vol. 93,
Issue. 1,
p.
229.
Alfadala, Sanad
Dawson, Deborah A.
Horsburgh, Gavin J.
Behnke, Jerzy M.
Bajer, Anna
Mohallal, Eman M. E.
Zalat, Samy
and
Slate, Jon
2013.
Large-scale isolation of Eastern spiny mouse Acomys dimidiatus microsatellite loci through GS-FLX 454 titanium sequencing.
Conservation Genetics Resources,
Vol. 5,
Issue. 2,
p.
519.
Lutermann, Heike
Medger, Katarina
and
Junker, Kerstin
2014.
Endoparasites of the Spiny Mouse (Acomys spinosissimus) from South Africa.
Journal of Parasitology,
Vol. 100,
Issue. 1,
p.
144.
Palmeirim, Marta
Bordes, Frédéric
Chaisiri, Kittipong
Siribat, Praphaiphat
Ribas, Alexis
and
Morand, Serge
2014.
Helminth parasite species richness in rodents from Southeast Asia: role of host species and habitat.
Parasitology Research,
Vol. 113,
Issue. 10,
p.
3713.
Bajer, Anna
Alsarraf, Mohammed
Bednarska, Małgorzata
Mohallal, Eman ME
Mierzejewska, Ewa J
Behnke-Borowczyk, Jolanta
Zalat, Sammy
Gilbert, Francis
and
Welc-Falęciak, Renata
2014.
Babesia behnkei sp. nov., a novel Babesia species infecting isolated populations of Wagner’s gerbil, Dipodillus dasyurus, from the Sinai Mountains, Egypt.
Parasites & Vectors,
Vol. 7,
Issue. 1,
Schutgens, M.
Cook, B.
Gilbert, F.
and
Behnke, J.M.
2015.
Behavioural changes in the flour beetleTribolium confusuminfected with the spirurid nematodeProtospirura muricola.
Journal of Helminthology,
Vol. 89,
Issue. 1,
p.
68.