Introduction
Phthalates are a family of chemicals utilized in the plastics production process to render plastic flexible. 1 Human phthalate exposure may occur through ingestion, inhalation, and or dermal contact and can readily cross the placental barrier. Reference Benjamin, Masai, Kamimura, Takahashi, Anderson and Faisal2 Di-2-ethylhexyl phthalate (DEHP) is among the most commonly used of the phthalates. Reference Wittassek and Angerer3,Reference Latini, De Felice and Verrotti4 Following exposure, DEHP is metabolized in the gastrointestinal tract into a variety of metabolites, including the bioactive metabolite mono-ethylhexyl phthalic acid (MEHP). Reference Frederiksen, Skakkebaek and Andersson5 MEHP has been identified as particularly toxic through both clinical and animal studies, Reference Inada, Chihara and Yamashita6–Reference Sant, Dolinoy, Jilek, Sartor and Harris10 shown to impact reproductive development such as disruption of male urogenital tract development. Reference Swan, Sathyanarayana and Barrett11 Other research found significant associations between prenatal phthalate exposure and increased birth weight Reference Sathyanarayana, Barrett, Nguyen, Redmon, Haaland and Swan12 as well as childhood and longitudinal body mass index. Reference Harley, Berger and Rauch13 Therefore, phthalates have been repeatedly characterized as “obesogens,” a term representing chemicals known to induce obesity, weight gain, and dyslipidemia (reviewed in refs. Reference Heindel14–Reference Grün and Blumberg17 ). On a molecular level, MEHP has been shown to cause damage both by inducing oxidative stress and by acting as an endocrine disruptor. Reference Wang, Craig, Basavarajappa, Hafner and Flaws18–Reference Zhai, Huang, Chen, Feng, Li and Li21
The cap “n” collar basic leucine zipper (CNC b-ZIP) transcription factor family plays a critical role in processes such as mitigating oxidative stress, cellular differentiation, carcinogenesis, and aging. Reference Shelton and Jaiswal22–Reference Sykiotis and Bohmann24 The CNC b-ZIP family includes nuclear factor erythroid 2 (Nfe2) and three related factors: nuclear erthryoid-1-related factor (Nrf1), nuclear erthryoid-2-related factor (Nrf2), and nuclear erthryoid-3-related factor (Nrf3). In the presence of oxidative stress, Nrf2 translocates to the nucleus whereby it heterodimerizes with small MAF proteins and binds to a cis-promoter element called the antioxidant response element (ARE). Reference Ma25 Binding to the ARE regulates a large family of cytoprotective genes. Reference Espinosa-Diez, Miguel and Mennerich26 In zebrafish, the nrf2a paralog is increasingly expressed during development, Reference Timme-Laragy, Karchner and Franks27 its expression is inducible by chemically stimulated oxidative stress, Reference Timme-Laragy, Karchner and Franks27 and is the primary inducer of cytoprotective gene expression. Reference Timme-Laragy, Karchner and Franks27–Reference Sant30 Ultimately, inadequate Nrf2 antioxidant and cytoprotective function can result in the generation of excessive reactive oxygen species (ROS) and cytotoxic biochemical changes such as lipid peroxidation and DNA damage.
In mammalian models, the reduction of ROS has led to improvement of disease states and symptoms including diabetes, hepatic steatosis, and hyperlipidemia. Reference Furukawa, Fujita and Shimabukuro31 Additionally, in obese humans, there is evidence of systemic oxidative stress, and reduction of oxidative stress has been shown to decrease the prevalence of metabolic syndrome. Reference Marseglia, Manti and D’Angelo32 The molecular mechanism of metabolic disorder phenotypes in response to oxidative stress is more unclear. The absence of Nrf2 was shown to decrease adiposity and adipocyte differentiation, Reference Pi, Leung and Xue19 potentially due to stimulation of glutathione metabolism, Reference Takahashi33 and yet another study has demonstrated no significant relationship. Reference Shin, Wakabayashi and Yates34 Likewise, there are inconsistent results regarding relationship between Nrf2 and obesity, and insulin resistance. Reference Seo and Lee35 These inconsistencies could be the result of strategies investigating different tissues, cells lines, or animal models at varying windows of time throughout the life course.
The liver plays a critical role in metabolic processes and is a major site of phase I and phase II metabolism of toxicants. 36 In addition to xenobiotic metabolism, liver cells are central to a number of processes such as the production of bile, breakdown of fats, and enzymatic control of blood sugars via glycolysis and gluconeogenesis. These roles define the liver as a crucial metabolic regulator and indicate different pathways that may be impacted by liver damage. Non-alcoholic fatty liver disease (NAFLD) encompasses a spectrum of conditions ranging in severity. In its most mild form, NAFLD presents as simple steatosis (abnormal retention of lipid) in the liver. Reference Benedict and Zhang37 Incurring further damage may lead to hepatitis (chronic liver inflammation), fibrosis, cirrhosis, and sometimes hepatocellular carcinoma. In addition to the implication of various metabolic processes, NAFLD is associated with both liver and cardiovascular related mortality. Reference Benedict and Zhang37 Some studies have suggested that NAFLD rates are between 20% and 30% in the western world. Reference Sayiner38 Although a high-fat diet in conjunction with a sedentary life is generally pointed to as the key risk factors for NAFLD, toxicant exposure has also been shown to induce NAFLD. Reference Arciello, Gori and Maggio39
A significant portion of the research investigating the role of Nrf transcription factors in metabolic disorder utilizes knockout models in conjunction with diet-induced obesity. Markedly less research has investigated the role of Nrf transcription factors in toxicant-induced metabolic disorder and obesity. Fewer studies yet have examined the embryonic period as a sensitive window of susceptibility during which exposures could induce pathological and physiological changes later in the life course. Herein, we examine the role of the Nrf2a transcription factor in mediating lipid bioaccumulation, namely, hepatic steatosis, in juvenile zebrafish that had been developmentally exposed to MEHP during the embryonic period.
Methods
Chemicals
MEHP of the highest purity was purchased from AccuStandard (New Haven, CT, USA). Dimethyl sulfoxide (DMSO) was purchased from Fisher Scientific (Pittsburg, PA, USA). An MEHP stock solution of 2 mg/ml was prepared by dilution in the vehicle, DMSO. All solutions were stored in amber-tinted vials at −20°C and vortexed before use. All other chemicals used in this study were purchased directly from Fisher Scientific.
Animals
Homozygous Nrf2a wild-type (WT) and mutant (nrf2a fh318−/− ) fish crossed onto an AB strain background were obtained from Dr. Mark Hahn (Woods Hole Oceanographic Institution, Woods Hold, MA, USA). This strain of zebrafish was generated through the TILLING mutagenesis Project (R01 HD076585) and originally obtained by Dr. Hahn as embryos from the Moens Laboratory (Fred Hutchinson Cancer Research Center, Seattle, WA, USA). The nrf2a fh318−/− genotype is considered a loss-of-function mutation, since the point mutation produces a mutant amino acid sequence in the DNA-binding domain of the Nrf2a protein, impairing its transcriptional activity. This mutation was originally characterized in ref. Reference Mukaigasa40 and further examined by our laboratory. Reference Jacobs, Sant, Basnet, Williams, Moss and Timme-Laragy9,Reference Sant29,Reference Rousseau41
All animal use and care was conducted in strict accordance with protocols approved by the University of Massachusetts Amherst and Bates College Institutional Animal Care and Use Committees (Animal Welfare Assurance Number UMASS A3551-01, Bates A3320-01). Adult breeding populations were housed in an Aquaneering automated zebrafish habitat at 28.5 °C and following a 14 h light:10 h dark cycle daily. Adult fish were fed the recommended amount of GEMMA Micro 300 (Skretting, Westbrook, ME, USA) once daily in the morning. Breeding populations were housed at an appropriate density with a 2:1 female-to-male ratio. Embryos for experiments were collected within 1 h post-fertilization (hpf) from homozygous genotyped tanks, washed thoroughly, and microscopically confirmed for fertilization prior to experimentation.
Exposures
WT and mutant embryos were exposed to either DMSO (vehicle) or 200 μg/l MEHP through immersion beginning at 6 hpf (gastrula period, shield stage) and concluding at 120 hpf (fully developed larvae). This concentration of MEHP has been previously utilized and optimized in previous zebrafish studies, and we have previously shown that it impacts the expression of genes in the Nrf2 signaling pathway. Reference Jacobs, Sant, Basnet, Williams, Moss and Timme-Laragy9,Reference Zhai, Huang, Chen, Feng, Li and Li21 MEHP or DMSO was added (0.01% v/v) to 0.3X Danieau’s medium (17 mM NaCl, 2 mM KCl, 0.12 mM MgSO4, 1.8 mM Ca(NO3)2, 1.5 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), pH 7.6) for immersion. Media was 50% replaced every 24 h and re-dosed with either DMSO or MEHP. Exposure took place in 20 ml glass scintillation vials with five embryos per vial. For each experiment, 3–5 vials were used per group. Experiments were performed in replicate to minimize clutch effects. At 96 hpf, some larvae were removed for analysis of yolk size and histology. At 120 hpf, remaining larvae were individually placed in 150 ml beakers in a 1:1 mix of 0.3X Danieau’s medium to clean system (adult breeding facility) water. Fish were transitioned to 100% system water by refreshing 50% system water daily until 15 days post fertilization (dpf) on a 12 h light:12 h dark cycle at 28.5 °C and being fed Gemma Micro 75 (Skretting).
Microscopy
For larvae collected at 96 hpf, animals were anesthetized in 0.3X Danieau’s medium containing tricaine mesylate (MS-222) (a 2% solution prepared from 4 mg/ml tricaine powder in water, pH buffered and stored at −20 °C until use) and staged in 3% methylcellulose. They were then imaged using a Leica M165 FC, and the area of the yolk was quantified using Leica Application Suite X (LAS X; Buffalo Grove, IL).
At 15 dpf, fish were imaged for length using the methods described above. To capture images most useful for length analysis, fish were mounted on their side. Images of nrf2a mutant larvae were captured utilizing an upright Olympus compound microscope with a Zeiss Axiocam 503 camera and Zen analysis software (Zeiss, USA). All images were captured under 20× magnification. Following imaging, larvae were washed thoroughly in 0.3× Danieau’s medium and either preserved for lipid staining or used for RNA isolation.
Histology
At 96 hpf, DMSO (control)-treated and MEHP-treated WT animals were fixed in 4% formaldehyde in 1× phosphate buffer, dehydrated in ethanol, and stored in 75% ethanol until embedding. Larvae were sent to Environmental Pathology Laboratories (Sterling, VA, USA) where they were embedded laterally into paraffin. Sagittal sections were made serially every 2 µm. Sections were mounted on superfrost glass slides and stained with hematoxylin and eosin. Histopathology was conducted by an Olympus BX 40 with an Olympus DP25 (Waltham, MA) camera system.
Lipid staining via oil red O
To spatially quantify larval lipid content, 15 dpf larvae were stained with Oil Red O. Reference Schlegel and Stainier42,Reference Fraher43 Following length imaging, three pools of five larvae per treatment group were reserved for gene expression analysis, and the remaining were preserved in 4% paraformaldehyde (PFA) overnight. Larvae preserved in PFA were then washed three times in 1× phosphate-buffer saline (PBS) for a total of 6 min. Larvae were transferred to individual 2 ml glass vials, briefly bleached, and then washed thoroughly with PBS (five washes). Larvae were then washed with 80% propylene glycol for 10 min, followed by 100% propylene glycol. Propylene glycol was removed, and larvae were stained with Oil Red O overnight. Fish were washed twice more with 80% and 100% propylene glycol following staining. Zebrafish were mounted on their side in 100% glycerol for imaging. Following imaging, stained larvae were stored indefinitely in glycerol.
RNA isolation and conversion to cDNA
Three pools of five larvae from each treatment group were used for RNA isolation. Following imaging at 15 dpf, RNA Later (Fisher Scientific) was added to five larvae per treatment group and stored at −80 °C until RNA isolation. Though individually housed from 5 to 15 dpf, larvae were randomized from the initial exposure vials to minimize batch effects. RNA was isolated using the GeneJET RNA purification Kit (Fischer Scientific, Waltham, MA, USA). The isolation protocol followed was developed for use of whole-tissue purification. The concentration of RNA and the purity of samples were analyzed using a µLITE spectrophotometer (BioDrop, Cambridge, UK), and all samples had quality A260/A280 ratios ranging from 1.8 to 2.1. After quantifying RNA, 500 ng RNA was reverse transcribed into cDNA with an iScript cDNA Synthesis Kit (Bio-Rad, Hercules, CA). These samples were diluted to a concentration of 0.25 ng/μl of cDNA in nuclease-free water and stored at −20 °C until use.
Gene expression
Quantitative real-time polymerase chain reaction (PCR) was conducted on the previously synthesized cDNA samples. Specifically, the Agilent Mx3000 qPCR (Agilent, Santa Clara, CA, USA) was used. Master Mix with Brilliant II SYBR Green was used in quantitative PCR with the genes described above as well as on several housekeeping genes. Target genes were compared to the arithmetic mean of three housekeeping genes 18 s, b2m, and beta actin utilizing the 2−ΔΔC T method. Reference Livak and Schmittgen44,Reference McCurley and Callard45
Triplicates of SYBR Master Mix and 10 ng of cDNA were performed for each sample. The conditions used in our qPCR were 95°C for 10 min followed by 35 cycles of 95°C for 30 s, 55°C (65°C for beta actin) for 60 s, and 72°C for 60 s. A melt curve was utilized to confirm the amplification of a singular product, and primers were tested for amplification of a single product prior to use. Primer sequences and temperatures have been previously published and are listed in Supplemental Table 1 Reference Timme-Laragy, Karchner and Franks27,Reference Sant29,Reference McCurley and Callard45–Reference Williams50 . All primer pairs were optimized in house using standard curves (amplification efficiencies ranged from 90% to 100%) and temperature gradients (±1 °C).
Data analyses
Data analyses were performed using IBM SPSS Statistics (v.25), Armonk, NY. To select appropriate statistical tests, tests for skewness and Shapiro–Wilk tests for normality were performed. The relationships between fish length, genotype, and exposure were analyzed using a non-parametric Kruskal–Wallis test with Games–Howell post hoc tests. To analyze the liver lipid content, red color intensity was normalized to image background using ImageJ (version 1.5; National Institutes of Health). These intensities were also compared across treatment and genotype using Kruskal–Wallis test with Games–Howell post hoc tests. To analyze the relationship between genotype, treatment, and gene expression, qPCR data were analyzed using a two-way analysis of variance (ANOVA) with Tukey’s multiple comparisons. A confidence level of 5% (α = 0.05) was used for all analyses.
Results
Embryonic deformities
To first assess whether knockdown of Nfe2 and Nrf family transcription factors, in combination with MEHP exposure, can alter embryogenesis, we examined 96 hpf embryos exposed to MEHP after knockdown using morpholinos (Supplemental Figure 1). MEHP increased swim bladder abnormalities in control, nrf1a, and nrf2a morphants compared to DMSO controls. Heart rate was increased in nrf2b and nrf3 morphants exposed to MEHP. No other structural abnormalities were observed due to Nfe2 signaling or MEHP exposure (p > 0.05). No changes were observed in yolk area between MEHP-exposed and control embryos (p > 0.05).
Histology
At 96 hpf, WT animals were assessed at the tissue level, including the brain, neural cord, eyes, muscle, swim bladder, gut, pancreas, liver, gill, ear labyrinth, yolk sac, notochord, and heart (sinus venous, atrium, ventricle, and bulbus). Upon examination of three animals per group (DMSO control and MEHP treatment), the only noted difference was seen in the MEHP treated animals where the liver was highly vacuolated with large vacuoles as compared to DMSO control animals where no vacuoles were seen (Fig. 1). Sections were scored to determine the presence of vacuolation and the type of vacuolation (Supplemental Table 2). Sections were scored to determine the incidence of vacuolation in the liver, but also the degree of vacuolation within cells. Embryos exposed to MEHP has significantly greater scores for both vacuolation (p = 0.001) and type of vacuolation (p = 0.004) in the liver (Table 1).

Fig. 1. Photomicrographs of histological sections through MEHP-exposed and DMSO-exposed (control) larval 96 hpf zebrafish. Images shown are 2 µm sections of paraffin-embedded larval tissues stained with hematoxylin and eosin. (a) WT larvae treated with DMSO (control). Hepatocytes (arrow) show homogenous, non-vacuolated cytoplasm. (b) WT larvae treated with MEHP. Hepatocytes (indicated by an arrow) show large vacuoles expanding the cytoplasm.
Table 1. MEHP exposure increases incidence and severity of vacuolation in the embryonic liver

Fish length and lipid accumulation at 15 dpf
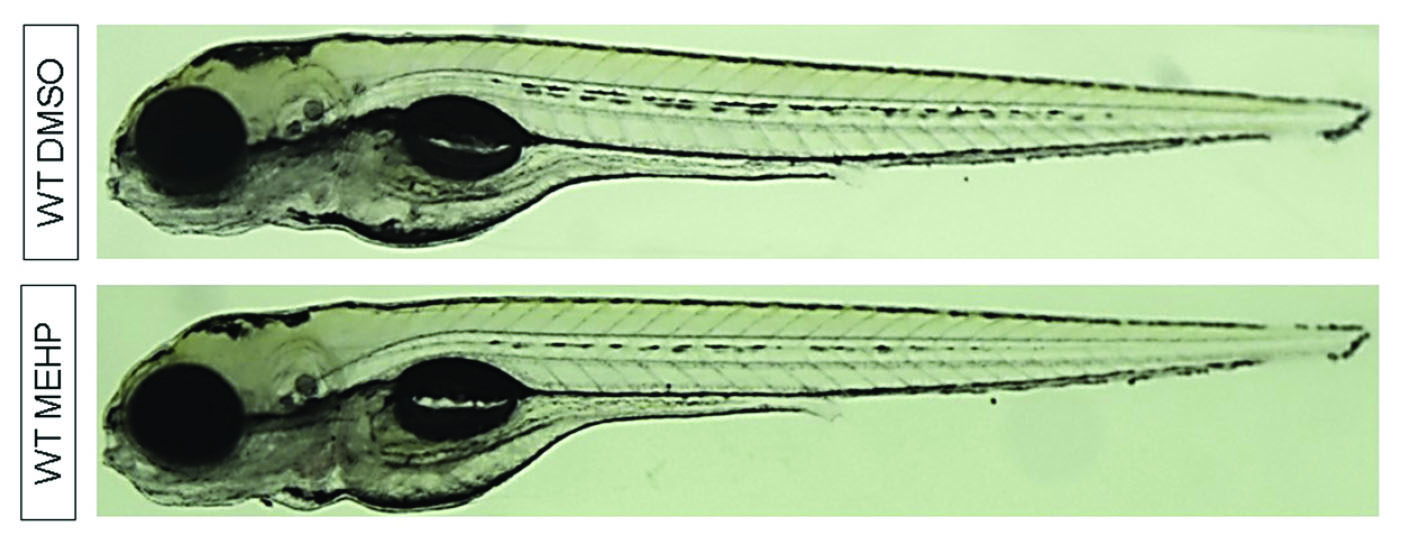
Following exposures from days 1 to 5, fish were then reared in clean conditions until day 15. Fish length measurements were taken at 15 dpf as a proxy for overall larval growth (Fig. 2a). Overall, MEHP exposure did not significantly impact larval growth (p > 0.05). In WT larvae, MEHP reduced fish length by 2.4%, though this decrease was not statistically significant (p > 0.05). Compared to unexposed WT larvae, mutant larvae were shorter by 8% (p < 0.001).

Fig. 2. Fish growth and adiposity vary with MEHP-exposure and Nrf2a genotype at 15 dpf. Images were analyzed via ImageJ. Values are means ± standard error of the mean. (a) For fish length, ANOVA with Tukey post hoc tests was used to assess exposure and genotyping changes in growth. Oil Red O staining intensity in the liver (b) and brain (c) was quantified and assessed using Kruskal–Wallis tests with Games–Howell post hoc tests. (d) Representative images of fish are shown, all at 50× magnification and the same light intensity. n = 48−53 for all treatment groups. Asterisks (*) indicate a statistically significant change due to exposure, within each genotype. Daggers (†) indicate a statistically significant change due to genotype, within each exposure group. p < 0.05.
Oil red O stain was utilized to spatially and quantitatively visualize neutral lipid accumulation in larvae at 15 dpf (Fig. 2b and 2c). Developmental MEHP exposure significantly increased lipid content within the liver (p = 0.002; Fig. 2b). Although genotype did not statistically modify these relationships, the magnitude of change due to exposure in mutant larvae was greater. In the brain, staining was more intense in MEHP-exposed larvae compared to controls, although this trend was only statistically significant in WT larvae (p = 0.016; Fig. 2c).
Gene expression – antioxidant response and Nrf2 interaction
Quantitative PCR was utilized to assess changes in gene expression resulting from MEHP exposures and impaired Nrf2a signaling (Fig. 3). These genes were selected as targets of Nrf2 signaling or interaction with the Nrf2 signaling pathway. Glutathione-S-transferase Pi (gstp) is a sensitive Nrf2-target and a well-characterized indicator of Nrf2 induction. The transcription factor Nfe2 (nfe2), like Nrf2, is a member of the CNC b-ZIP transcription factor family and also mediates the antioxidant response in the embryo. Reference Williams50,Reference Williams51 Enzymes cytochrome P450 2E1 (cyp2e1) and histone deacetylase (hdac) are involved in a myriad of cellular processes including chromatin structure and the hepatic xenobiotic response, and both enzymes have been shown to have interactions with Nrf2 signaling. Reference Cederbaum and Dey52

Fig. 3. Expression of gstp1, nfe2, cyp2e1, and hdac in WT and Nrf2a mutant zebrafish. Zebrafish embryos and larvae were exposed to DMSO or MEHP from 6 to 120 hpf, and gene expression was assessed at 15 dpf. Data are presented as mean fold change normalized to the mean of housekeeping genes 18 s, b2m, and beta actin ± SEM. Data were analyzed using two-way ANOVA with Tukey's multiple comparisons. Asterisks (*) indicate a statistically significant change due to exposure, within each genotype. Daggers (†) indicate a statistically significant change due to genotype, within each exposure group. p < 0.05; n = 3 pools of five larvae.
Gene expression of gstp was significantly attenuated by approximately 60% in Nrf2a mutant larvae, regardless of exposure (p < 0.05). No significant changes in gstp gene expression were observed due to MEHP exposure for either genotype (p > 0.05). No statistically significant effects due to exposure or genotype were observed for nfe2 or cyp2e1 (p > 0.05; Fig. 3). There was also a subtle increase in nfe2 gene expression due to MEHP-exposure for both genotypes, and Nrf2a mutants had elevated expression compared to WT larvae (p > 0.05). Expression of cyp2e1 and hdac was moderately increased in DMSO-exposed Nrf2a mutant larvae compared to WT larvae, but these trends were not statistically significant (p > 0.05; Fig. 3).
Gene expression – peroxisome proliferator-activated receptor signaling
Gene expression of peroxisome proliferator-activated receptor (PPAR) gamma (pparγ) and alpha (pparaa) and several of their targets were assessed at 15 dpf (Fig. 4). PPARs are transcription factors with multifaceted regulatory domains over metabolic processes including, but not limited to, glycolysis and gluconeogenesis, lipid metabolism and catabolism, and lipid transport. Expression of isoforms pparaa and pparg was measured as well as several of their gene targets, notably lipid transporter apolipoprotein A1 (apoa1a) and fatty acid-binding proteins (FABP) 1a and 1b (fabp1a, fabp1b.1). The gene apoa1a is a hepatic and yolk syncytial layer transporter involved in cholesterol and lipid transport and is responsive to both PPARα and PPARγ signaling. Reference Thisse53 FABPs are intracellular carrier proteins responsible for lipid binding and transport in a myriad of tissues and systems. Reference Fagerberg54 Expression of fabp1a1 and fabp1b.1 is subfunctionalized in zebrafish, with fabp1a1 being responsive to PPARα signaling and fabp1b.1 being responsive to PPARγ signaling. Reference Laprairie, Denovan-Wright and Wright48 Numerous studies have demonstrated that MEHP is an activator of both PPARα and PPARγ signaling. Reference Feige55,Reference Hurst and Waxman56 However, it was unknown whether this activation would persist in larvae at 15 dpf after only developmental exposures ceasing at 5 dpf.

Fig. 4. Expression of pparaα, pparg, fabp1a1, fabp1b1, and apoa1a in WT and nrf2a mutant zebrafish. Zebrafish embryos and larvae were exposed to DMSO or MEHP from 6 to 120 hpf, and gene expression was assessed at 15 dpf. Data are presented as mean fold change normalized to the mean of housekeeping genes 18 s, b2m, and beta actin ± SEM. Data were analyzed using two-way ANOVA with Tukey’s multiple comparisons. Asterisks (*) indicate a statistically significant change due to exposure, within each genotype. Daggers (†) indicate a statistically significant change due to genotype, within each exposure group. p < 0.05; n = 3 pools of five larvae.
No statistically significant changes in gene expression were observed for either the pparaa or pparg transcription factors, nor for targets apoa1a or fabp1b.1 (p > 0.05; Fig. 4). However, there were notable decreasing trends in apoa1a and fabp1b.1 in Nrf2a mutants and Nrf2a mutants embryonically exposed to MEHP (p > 0.05). Expression of fabp1a1 was increased by MEHP exposure in WT larvae, and Nrf2a mutants were elevated compared to WT larvae regardless of exposure (p < 0.05).
Zebrafish embryos and larvae were exposed to DMSO or MEHP from 6 to 120 hpf, and gene expression was assessed at 15 dpf. Data are presented as mean fold change normalized to the mean of housekeeping genes 18 s, b2m, and beta actin ± SEM. Data were analyzed using two-way ANOVA with Tukey’s multiple comparisons. Asterisks (*) indicate a statistically significant change due to exposure, within each genotype. Daggers (†) indicate a statistically significant change due to genotype, within each exposure group. p < 0.05; n = 3 pools of five larvae.
Discussion
The purpose of this study was to investigate the potential interaction between developmental exposure to MEHP and the antioxidant response and their impact on larval steatosis. We hypothesized that embryonic MEHP exposure would increase larval lipid accumulation and that impaired Nrf2a function would exacerbate this phenotype. Our data indicate that developmental MEHP exposure increases risk for hepatic steatosis in larval zebrafish. Significant decreases in gstp gene expression are concordant with previously reported understandings of the role of Nrf2a in cytoprotection, and increased fabp1a1 in mutants and due to MEHP exposures suggests a PPARα-mediated adaptive response to steatosis. The exacerbation of steatosis as well as increased fabp1a1 expression in Nrf2a mutant larvae due to MEHP exposures suggests that Nrf2a may function in hepatoprotection.
In this study, developmental MEHP exposure reduced mean fish length by 2.2% in WT larvae, though this decrease was not statistically significant (p > 0.05). However, unexposed Nrf2a mutant larvae were 7.7% shorter than WT larvae (Fig. 2), suggesting that Nrf2a may play a role in overall growth and development. Most significantly, these phenotypic effects were observed at 15 dpf – after being only exposed from 6 to 120 hpf. We have previously observed this decrease in total fish length due to other developmental toxicant exposures (perfluorooctane sulfonic acid Reference Sant57 ;), but not in Nrf2a mutant fish examined at earlier timepoints (7 dpf Reference Sant29 ;). Only a modest 4%–6% decrease in human fetal length exists between the median infant length and fetuses small for gestational age (SGA), which is defined as the lowest 10th percentile for fetal growth according to the Fenton growth chart. Reference Fenton and Kim58 Therefore, the 7.7% reduction in fish length observed in this study is physiologically relevant and may propagate other phenotypic and metabolic changes commonly associated with SGA or low-birth-weight classifications (reviewed in references Reference Dulloo59–Reference Simmons61 ).
The prevalence of pediatric obesity and overweight children has been steadily increasing in recent decades. Reference Pan62,Reference Fryar, Carroll and Ogden63 Here, we assessed how a ubiquitous environmental toxicant, MEHP, impacts developmental steatosis and examined the modification of this relationship by the antioxidant response pathway. We confirmed the results of previous studies, demonstrating that MEHP exposures increased hepatic steatosis, even as early as 120 hpf (Fig. 1). Hepatic steatosis at 15 dpf was significantly increased by developmental MEHP exposure in both WT and Nrf2a mutant larvae (Fig. 2). These data indicate that early developmental exposures alone to MEHP may increase risk for juvenile hepatic steatosis, even if exposures are remediated during the larval period. Several studies have previously shown that exposures to MEHP or parent compound DEHP increased lipid accumulation in vitro or hepatic steatosis in vivo. Reference Bai64–Reference Pereira, Mapuskar and Rao66 However, to our knowledge, this is the first study demonstrating that exposures during the embryonic period alone are capable of inducing lasting steatosis into the larval period.
We have previously shown that embryonic exposures to MEHP impacted the expression of Nrf2a targets gsr and gstp1 in 96 hpf zebrafish, but that the embryonic glutathione redox couple was not significantly impacted. Reference Jacobs, Sant, Basnet, Williams, Moss and Timme-Laragy9 We also previously demonstrated that the oxidative response to MEHP is rapid, occurring primarily within the first few hours after exposure, in mouse whole embryo culture. Reference Sant, Dolinoy, Jilek, Sartor and Harris10 Therefore, the timing of glutathione and antioxidant enzyme assessment relative to exposure is an important variable in MEHP-induced oxidative stress research. In this study, we examined the gene expression of antioxidant enzymes at 15 dpf following developmental exposures (from 6 to 120 hpf). This delayed assessment of the antioxidant response allowed us to probe the more programmatic or longitudinal impacts of developmental exposures rather than an acute response. Here, we found that gstp1 expression was decreased by impaired Nrf2a signaling, but that MEHP had no impact on gstp1 expression (Fig. 3). Therefore, it is unlikely that developmental exposures to MEHP induce acute oxidative effects, but that this acute oxidative stress does not produce any direct chronic oxidative effects.
The endogenous antioxidant response is coordinated largely by the Nfe2 family of transcription factors, including Nrf1a, Nrf1b, Nrf2a, Nrf2b, Nrf3, and Nfe2 in the zebrafish model. We have previously published several studies examining the oxidative stress response and embryonic phenotypes of deficient signaling by this antioxidant family and have observed that several of these family members have redundant functions and may compensate for another’s impairment. Reference Timme-Laragy, Karchner and Franks27,Reference Sant30,Reference Williams50 Here, we observed decreased gstp1 gene expression in Nrf2a mutant larvae and stable gene expression of family member nfe2 regardless of genotype or exposure (Fig. 3). Therefore, it is unlikely that compensation of deficient Nrf2a signaling is occurring, at least not via the Nfe family of antioxidant transcription factors.
PPAR activity has been widely implicated in environmental determinants of metabolic dysfunction due to its regulatory role in processes such as carbohydrate and lipid metabolism and storage. In zebrafish, PPARs are expressed in a myriad of tissues throughout the entire life course, suggesting constitutive gene expression. Reference Ibabe, Bilbao and Cajaraville67 However, the differential expression of PPAR genes in various tissues and in response to metabolic conditions also demonstrates the potential for induction of this signaling pathway. PPARα agonism has been widely explored and developed as potential therapeutic for hepatic steatosis, and increased expression has as an adaptive response to ameliorate hepatic steatosis (reviewed in reference Reference Liss and Finck68 ). We had previously found that developmental exposures to another environmental toxicant, perfluorooctanesulfonic acid, could induce PPAR gene expression in developing embryos but had not assessed any more longitudinal changes in expression after 96 hpf. Reference Sant29 In this study, gene expression of pparg and pparaa was not changed at 15 dpf despite increased hepatic steatosis resulting from both developmental MEHP exposure and deficient Nrf2a signaling (Fig. 4). However, expression of PPARα-target fabp1a1 was increased by both MEHP exposure and impaired Nrf2a signaling, suggesting that PPARα activity, rather than expression, may be modulated by exposure and genotype. Therefore, future work should complement these measures with protein expression, cellular localization, and transcriptional regulation (such as by Chromatin ImmunoPrecipitation and sequencing) data to probe the activity and induction of PPARα.
The potential for crosstalk between the Nrf2 and PPAR signaling pathways needs to be explored. We previously identified putative AREs within the promoter of pparaa and pparg in zebrafish, as well as PPAR-response elements in the promoters of Nfe2 family members and their targets. Reference Sant29 Likewise, we had found that the expression of pparaa and pparg to exposure was only induced in nrf2a mutant embryos. Reference Sant29 In the present study, we observed no statistically significant changes in gene expression of pparaa or pparg in Nrf2a mutant larvae (Fig. 4). These data suggest that any potential Nrf2-PPAR crosstalk is unlikely to play a significant role in MEHP-induced steatosis.
A limitation of this study is the inability to observe sex-based differences in hepatic steatosis in larval zebrafish. Unlike mammalian models, zebrafish sexual differentiation is polygenic, meaning that it is not simply determined by a singular genetic locus. Reference Liew69 Due to this complexity, sexual determination does not occur in zebrafish until approximately 30 dpf, marking the change from the larval to the juvenile period. Reference Uchida70 Studies utilizing rodents to investigate metabolic outcomes of phthalate exposure in juveniles found sexually dimorphic responses in numerous phenotypes, expected due to the endocrine disrupting properties of phthalates. There is ample epidemiologic and model data demonstrating that males are at a higher risk for obesity, diabetes, and NAFLD than women and that these relationships are modified by age. Reference Phillips71–Reference Bertolotti73 Therefore, future longitudinal studies beyond the larval stage need to explore the role of biological sex in MEHP-induced hepatic steatosis.
In conclusion, this study demonstrated that early developmental exposures to MEHP increased hepatic steatosis during the mid-larval period more than 10 days after exposure. These data suggest that there are lasting metabolic effects from embryonic exposures and provide further evidence that MEHP may contribute to the developmental origins of metabolic dysfunction. Nrf2a antioxidant signaling did not play a significant role in MEHP-induced steatosis, though decreased Nrf2a signaling increased fabp1a gene expression regardless of exposure. Overall, this study demonstrates the significance of embryonic-only exposures in the developmental origins of metabolic dysfunction.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/S2040174420000057
Acknowledgments
This work was possible due to the exceptional animal care and laboratory assistance of Christopher Clark, Sarah Conlin, Mary Hughes, Sadia Islam, Archit Rastogi, Monika Roy, Olivia Venezia, and Michael Young.
Financial Support
This work was funded by an Institutional Development Award from the National Institute of General Medical Sciences of the National Institutes of Health (P20 GM103423 to Bates College) as well as support from the National Institute of Environmental Health Sciences (R01 ES025748 and R01 ES028201 to AT-L; F32 ES028085 to KES).
Conflict of Interest
None.
Ethical Standards
The authors assert that all procedures contributing to this work comply with the ethical standards of the AAALAC Guidance on the Housing and Care of Zebrafish (Danio rerio) and has been approved by the University of Massachusetts Amherst and Bates College Institutional Animal Care and Use Committees (Animal Welfare Assurance Number UMASS A3551-01, Bates A3320-01).