


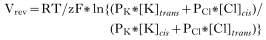



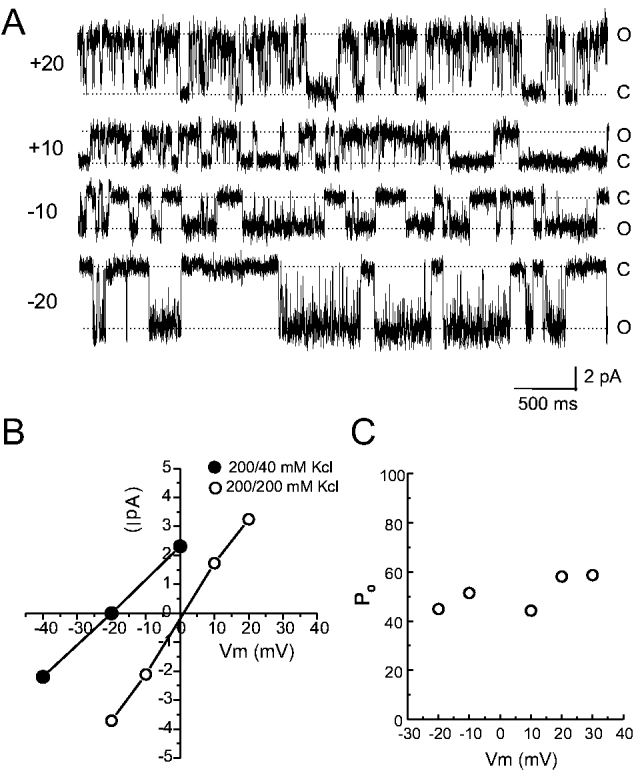










Crossref Citations
This article has been cited by the following publications. This list is generated based on data provided by Crossref.
Park, Jin Bong
Son, Sook Jin
Lee, Gyu Seung
Cho, Pyo Yun
Song, Ky Sun
Ryu, Pan Dong
Kang, Shin-Yong
and
Hong, Sung-Jong
2005.
Molecular and electrophysiological characterization of nucleotide-sensitive chloride current-inducing protein of Fasciola hepatica.
Molecular and Biochemical Parasitology,
Vol. 140,
Issue. 2,
p.
197.
Jimenez, Veronica
Henriquez, Mauricio
Galanti, Norbel
and
Riquelme, Gloria
2011.
Electrophysiological characterization of potassium conductive pathways in Trypanosoma cruzi.
Journal of Cellular Biochemistry,
Vol. 112,
Issue. 4,
p.
1093.
Jang, Jung Hee
Park, Jin Bong
Kim, Sun Don
Lee, So Yeong
Hong, Sung-Jong
and
Ryu, Pan Dong
2012.
Property of large conductance Ca2+-activated K+ channels from Fasciola hepatica incorporated into planar lipid bilayer.
Veterinary Parasitology,
Vol. 186,
Issue. 3-4,
p.
281.
Jimenez, Veronica
Docampo, Roberto
and
Carruthers, Vern B.
2012.
Molecular and Electrophysiological Characterization of a Novel Cation Channel of Trypanosoma cruzi.
PLoS Pathogens,
Vol. 8,
Issue. 6,
p.
e1002750.
Park, Kwon Moo
Kim, Sun-Don
Park, Jin Bong
Hong, Sung-Jong
and
Ryu, Pan Dong
2021.
Electrophysiological Properties of Ion Channels in Ascaris suum Tissue Incorporated into Planar Lipid Bilayers.
The Korean Journal of Parasitology,
Vol. 59,
Issue. 4,
p.
329.
Chakroborty, Anand
Pritchard, Deiniol
Bouillon, Marc E.
Cervi, Anna
Cookson, Alan
Wild, Charlotte
Fenn, Caroline
Payne, Joseph
Holdsworth, Peter
Capner, Colin
O’Neill, Jenna
Padalino, Gilda
Forde-Thomas, Josephine
Gupta, Sandeep
Smith, Brendan G.
Fisher, Maggie
Lahmann, Martina
Baird, Mark S.
and
Hoffmann, Karl F.
2022.
Flukicidal effects of abietane diterpenoid derived analogues against the food borne pathogen Fasciola hepatica..
Veterinary Parasitology,
Vol. 309,
Issue. ,
p.
109766.