Introduction
Predictable, incremental growth is an important tool for understanding many organisms and their environment (e.g., Hemming Reference Hemming1969; Pannella Reference Pannella1971; McLaughlin et al. Reference McLaughlin, Downing, Blasing, Cook and Adams1987). Lichen growth rates have been used to determine productivity and performance of lichen communities (e.g., Karenlampi Reference Karenlampi1971; Vasander Reference Vasander1981; Keon & Muir Reference Keon and Muir2002), as measures of lichen response to changes in their environment (e.g., Miller Reference Miller1973; Lechowicz Reference Lechowicz1981; Benedict Reference Benedict1990; Kytöviita & Crittenden Reference Kytöviita and Crittenden2002), or to estimate age of the substratum upon which they are growing (e.g., Armstrong Reference Armstrong1983; Innes Reference Innes1985; McCarthy Reference McCarthy1999; Solomina & Calkin Reference Solomina and Calkin2003). In addition, growth curves inferred from these rates can contribute to lichen population dynamics (Yarranton Reference Yarranton1975).
Most studies use marginal growth rates, assessing incremental change of thallus radius, perimeter or area over time (e.g., Armstrong Reference Armstrong1973, Reference Armstrong1993). For saxicolous crusts and foliose lichens these two-dimensional methods may be adequate, but not for fruticose lichens. Change in biomass or lichen height has been used to account for the three-dimensional growth of these lichens (e.g., Ahti Reference Ahti1959; Steen Reference Steen1965; Lechowicz Reference Lechowicz1981; McCune et al. Reference McCune, Derr, Muir, Shirazi, Sillett and Daly1996; Hyvarinen & Crittenden Reference Hyvärinen and Crittenden1998; Peck et al. Reference Peck, Ford, McCune and Daly2000). Another method implemented widely in Arctic systems to assess growth within a small group of Cladonia (those with regular branching, no cortex and a crustose primary thallus—formerly recognized as Cladina; Stenroos et al. Reference Stenroos, Hyvönon, Myllys, Thell and Ahti2002) is average annual linear growth rate (Scotter Reference Scotter1963; Karenlampi Reference Karenlampi1970; Prince Reference Prince1973; Lechowicz Reference Lechowicz1983). Although originally outlined by Salazkin (Reference Salazkin1937), Andreev (Reference Andreev1954) popularized this technique, which relies upon this group of Cladonia producing a whorl of branches each year, thus enabling growth rates to be derived by dividing total lichen height by the number of internodes.
More recently, accelerator mass spectrometry (AMS) was used to date precisely lichen fragments by measuring 14C/C ratios that compare the carbon content in a fragment to a radiocarbon standard (Clark et al. Reference Clark, Mangelson, St. Clair, Rees, Bench and Southon2000; Bench et al. Reference Bench, Clark, Mangelson, St. Clair, Rees, Grant and Southon2001; Clark Reference Clark2001). This technique was used successfully to age the crustose lichen, Caloplaca trachyphylla (Tuck.) Zahlbr (Clark et al. Reference Clark, Mangelson, St. Clair, Rees, Bench and Southon2000). However, similar work with the crustose lichen, Rhizocarpon geographicum (L.) DC, and fruticose lichen, Usnea longissima Ach., showed flat 14C profiles (Bench et al. Reference Bench, Clark, Mangelson, St. Clair, Rees, Grant and Southon2001; Clark Reference Clark2001; Bench et al. Reference Bench, Clark, Mangelson, St. Clair, Rees, Grant and Southon2002). To our knowledge, this methodology has not been implemented successfully with any fruticose lichen.
Our main objectives for this study were: (1) examine the utility of 14C/C ratios for estimating growth rates and ages of Cladonia stygia (Fr.) Ruoss. (2) estimate the average growth rate and absolute age of Cladonia stygia in north-western Alaska using both traditional methods (sensu Andreev) and 14C/C ratios, and compare their results.
Materials and Methods
Sampling and processing
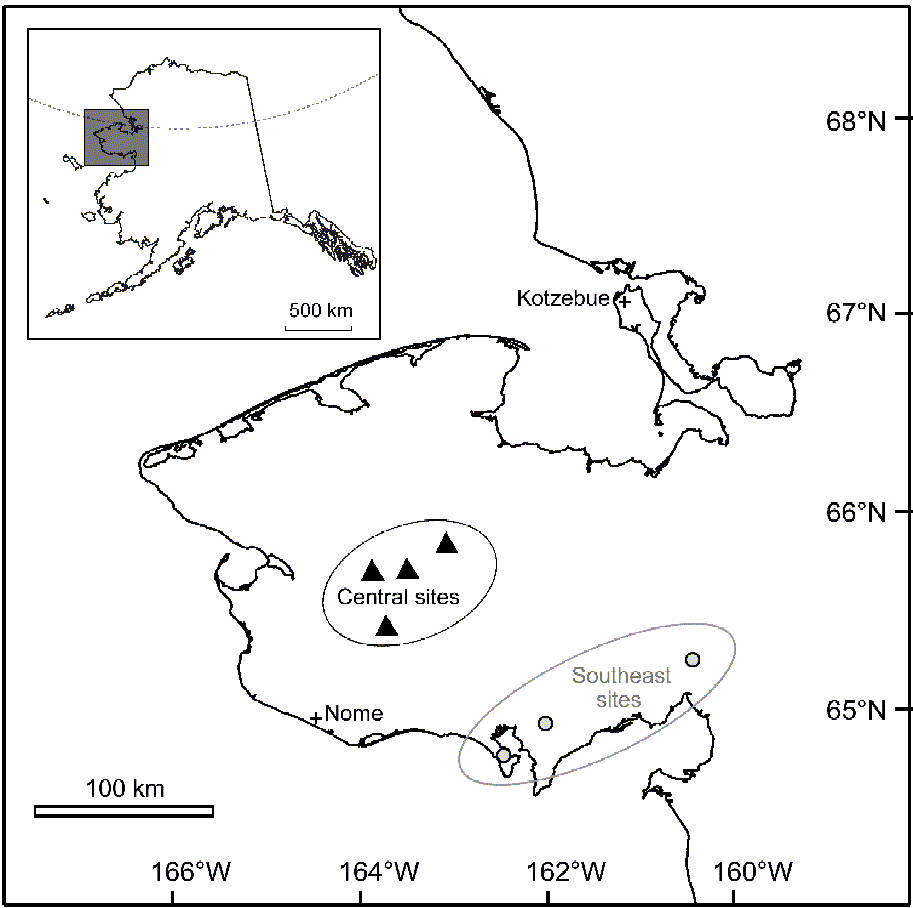
Cladonia stygia is a terricolous lichen endemic to open bog environments (Ahti & Hyvönen Reference Ahti and Hyvönen1985). This fruticose species occurs primarily in boreal and arctic regions, yet also extends into some temperate areas (Ahti & Hyvönen Reference Ahti and Hyvönen1985; Perlmutter Reference Perlmutter2005). During the 2003 and 2004 field seasons, 12 individual thalli were collected from north-western Alaska, USA (64°30′–65°30′N, 161°07′–165°05′W; 20–630 a.s.l.). To represent the climatic and environmental variability, thalli were collected from seven different sites on the Seward Peninsula, Alaska (Fig. 1). Robust, primarily unbroken thalli were collected and stored in glass vials and subsequently transported to Oregon State University for sectioning.

Fig. 1. Map of Seward Peninsula and seven sites from which lichens were collected. The dashed line in the Alaska inset in the upper left corner represents the Arctic Circle. Black triangles represent the four central sites (6 lichens were collected from these sites) and grey circles represent the three south-eastern sites (6 lichens). Plus signs indicate nearby towns of Nome and Kotzebue.
Thalli were washed with deionized water to remove large adhering fragments and to re-hydrate for ease of processing. While moist, we measured total length of the predominant main axis and total number of internodes, or spaces between branches, along this axis. All side branches were trimmed away and not sampled. From the base of the main axis towards the tip, fragments, ranging between 2–8 mm in length, were sampled approximately one centimetre apart. To minimize the effects of carbon turnover, the outer medulla and algal layer of each fragment were scraped away and considered as a separate sample. Small segments of the inner medulla, or sterome, were the primary samples used in our analyses. These samples were stored in glass vials and transported to the Center for Accelerator Mass Spectrometry (CAMS) at Lawrence Livermore National Laboratory.
At the CAMS facility, samples were washed in 1N HCl at 80°C to remove any remaining adhering carbonate. Lichen fragments were weighed and loaded with CuO oxidizer into quartz tubes. Loaded tubes were evacuated, sealed and combusted at 900°C. Catalyzed by iron powder, graphite was condensed from CO2 gas at a low temperature (Vogel et al. Reference Vogel, Southon, Nelson and Brown1984). The 14C/C isotope ratio in the graphitized sample was measured by AMS (Southon et al. Reference Southon, Caffee, Davis, Moore, Proctor, Schumacher and Vogel1990). Sample ages were calibrated with Levin & Kromer (Reference Levin and Kromer2004), one-year smoothing and one-year resolution using CaliBomb (Reimer et al. Reference Reimer, Brown and Reimer2004). For all analyses, we used the mean of the upper and lower calibrated ages.
Comparison of the isotopic ratios of lichen inner medullary fragments to ratios estimated by Levin & Kromer (Reference Levin and Kromer2004) allows us to determine the approximate date that carbon fixed by the photobiont was incorporated by the mycobiont. Although carbon turnover has been demonstrated in some lichens (Bench et al. Reference Bench, Clark, Mangelson, St. Clair, Rees, Grant and Southon2002), we believe removal of the photobiont helps reduce these effects. Preliminary analyses of samples containing both photobiont and outer medulla scraped from the inner medulla, demonstrated similar or no trends of age with distance from the lichen base, which substantiates our choice of using primarily inner medullary fragments for our analyses.
Analyses
Traditional estimates of lichen growth rate were determined using Andreev's (Reference Andreev1954) method of average annual linear growth rates (i.e., lichen height divided by the number of internodes on a single thallus). Absolute age was calculated by subtracting the number of internodes from the year of collection. Growth rates from 14C/C ratios were derived from slope estimates of regressing the approximate distance of each sample from the base of the lichen against the mean calendar year determined from 14C/C ratios. The absolute age from 14C/C ratios was the mean year of measurement for the basal-most sample of each lichen. Growth rates and ages were compared with paired and two-sample t-tests.
Results and Discussion
Growth rate
Using traditional methods (sensu Andreev Reference Andreev1954), the average growth rate of Cladonia stygia in north-western Alaska was 6·2 mm yr−1. Our estimates of growth are slightly faster than reported elsewhere for other species within this group of Cladonia using the same method (e.g., Andreev Reference Andreev1954; Scotter Reference Scotter1963; Pegau Reference Pegau1968; Prince Reference Prince1973; Boudreau & Payette Reference Boudreau and Payette2004). Vasander's (Reference Vasander1981) measurements of C. arbuscula (Wallr.) Flotow and C. rangiferina (L.) F. H. Wigg. in southern Finland, however, were fairly similar to our growth rates using this traditional internode method.
14C/C ratios measured in Cladonia stygia revealed linear trends in lichen growth rate (Fig. 2). The average growth rate of our lichens, using data from 14C/C ratios, was 6·5 mm yr−1. We found the two methods, internode-based growth rates and carbon ratios, did not significantly differ from one another (t = −0·75, P = 0·47). Moreover, the variability in rates was not systematically biased between the two methods (Fig. 3). Although we do not know the ‘true’ growth rate of these lichens, both methods appear to have roughly equal bias and error. Therefore, this concordance supports further use of Andreev's internode method as a reliable field practice to estimate growth rates of richly branched Cladonia.

Fig. 2. Scatterplot of distance of lichen segment from base, in millimeters, and age of that segment derived from 14C/C ratios. Lines represent least square regressions of twelve separate lichens (0·853 < R2 < 0·998). Black lines and triangles are lichens from central Seward Peninsula and dashed lines and open circles are from the south-eastern Seward Peninsula.

Fig. 3. Scatterplot of growth rates of 12 individual lichens using two different methods; internode growth rates (sensu Andreev Reference Andreev1954) and 14C/C ratios (slopes from Fig. 2). Points fall evenly on either side of the one-to-one line (shown as dashed line).
Absolute age
The oldest lichen we found, using either method, was 30 years old. We found lichen age did not differ significantly between the two methods (t = −0·59, P = 0·60); although, trends in the data suggest 14C/C ratios may underestimate age of younger lichens, while the internode method may underestimate age of older lichens (Fig. 4). Due to the indeterminate growth of lichens, these individuals may have been much older, yet the older portions at the base had decayed or broken off and were not sampled. The age divergence in young lichens may possibly be explained by local departures from the 14C bomb curve which is a hemisphere average.

Fig. 4. Scatterplot of lichen age of 12 individual Cladonia stygia specimens using two different methods: traditional internode method (sensu Andreev Reference Andreev1954) and 14C/C ratios. Bars represent the confidence interval surrounding the mean calibrated age based on 14C/C ratios from CaliBomb (Reimer et al. Reference Reimer, Brown and Reimer2004). Younger lichens tended to fall below the one-to-one line (shown as dashed line), while older lichens tended to fall above the one-to-one line.
Our 14C/C ratios for Cladonia stygia contrast with those reported for the fruticose lichen Usnea longissima, which showed flat 14C profiles (Clark Reference Clark2001). Clark (Reference Clark2001) proposed that complex carbon transfer and subsequent turnover in fruticose and foliose lichens manifest as uniform carbon profiles, inhibiting their usefulness in dating studies. These disparate results raise the question: is carbon simply not turning over as quickly in Cladonia, or does the climate of the surrounding environment [i.e., our sites were colder and shorter-seasoned than those of Clark (Reference Clark2001)] reduce the degree of turnover? Future studies incorporating different species, within this group of Cladonia and from other fruticose genera, may yield insights into this disparity of success with 14C/C ratios.
Geographic differences
Interestingly, lichen growth was not consistent across the geographic range sampled. Using growth rates derived from both 14C/C ratios and internode counts, we found lichens collected in the south-eastern portions of the Seward Peninsula had significantly faster growth rates, than those from the central portion of the Peninsula (t = 4·76, P < 0·01; Figs. 1 & 2). Average growth rates based on 14C/C ratios and internode counts for the south-eastern sites were 7·8 and 7·0 mm yr−1, and 5·2 and 5·5 mm yr−1 for the central sites, respectively. The south-eastern sites are located at the junction of the Seward Peninsula and mainland Alaska; thus, these sites tend to be more continental while those to the west are more oceanic. Elsewhere, lichen growth rates have been linked to continentality of climate (e.g., Andreev Reference Andreev1954); with faster lichen growth in more oceanic climates, where temperature and precipitation are more favourable. Conversely, our results depict increased growth rates with increased continentality. Due to lack of detailed local climate data for each collection site, we were unable to determine whether climate accounts for the observed differences in lichen growth rates. Previous studies of crustose species, however, support the view that climate significantly influences lichen growth (Armstrong Reference Armstrong1993, Reference Armstrong2005; Benedict Reference Benedict1991).
Furthermore, lichens from the central portion of the Seward Peninsula were older on average than those from south-eastern sites (comparing intercepts, t = −4·77, P < 0·01; Fig. 2). These geographic differences in age and growth rate may also be explained by biotic interactions. Average total lichen cover and lichen species richness of the south-eastern sites (52% cover and 32 species) tended to be greater than those in the central sites (14% cover and 23 species). Faster growing lichens in a highly competitive environment, such as sites to the southeast, would likely succeed more easily. Future studies, accounting for both climate and competition, are needed to clarify further disparities in lichen growth over large geographic areas.
Conclusions
14C/C ratios can successfully estimate growth rate and age of the fruticose lichen, Cladonia stygia. Although routine use of this technique is unrealistic, in both its financial and time expense, for most ecological studies, the exceptional value of our work was to identify an alternative method of dating fruticose lichens. We found growth rates derived from 14C/C ratios closely coincided with those from Andreev's internode method. This correspondence justifies future use of the internode method as a quick yet reliable field method. Additional support for Andreev's method may be welcomed by Arctic ecologists as many fruticose species occur in remote, inaccessible areas where repeated observations are impossible or would disrupt the community in a way that might provide inaccurate estimates of growth.
The authors thank S. Miller, S. Nunn and A. Hardman for their support in the field, and T. Guilderson and the CAMS staff for their help in the laboratory. A. Green provided comments on an earlier version of the manuscript. We are also grateful to P. Neitlich and the National Park Service who provided support and additional funding for this study. This work performed under the auspices of the U.S. Department of Energy by Lawrence Livermore National Laboratory under Contract DE-AC52-07NA27344.