

The fossil record of sponges is incomplete, complex, and increasingly important for understanding early animal evolution (Botting & Muir Reference Botting and Muir2018). One of the key indicators of our limited state of knowledge in Ordovician and Silurian sponge diversification is the large proportion of highly problematic taxa being described (e.g., Botting & Rhebergen Reference Botting and Rhebergen2011; Botting & Zhang Reference Botting, Muir and Lin2013). This implies long phylogenetic gaps between known taxa, which severely hinders a full understanding of how the diversification occurred. In particular, a key point is the difficulty in recognising the last common ancestors of the class-level crown groups. For demosponges this is now becoming clear, but for hexactinellids is still highly problematic, with numerous implications for how to interpret the earliest hexactin-bearing sponge fossils (Botting & Muir Reference Botting, Zhang and Muir2018). New taxa that may represent early examples of crown-group hexactinellids are, thus, vital to developing our knowledge in this area. This paper adds to the literature by describing a new, surprisingly modern-looking hexactinellid from the lower Silurian rocks of the Pentland Hills, Scotland.
The first geological investigations of the Pentland Hills date to as far back as the early 19th Century, when French expatriate Boué (Reference Boué1820) noted the presence of rocks older than the Old Red Sandstone (ORS). However, he did not record the presence of any fossils at that time, and it was not until 1836 that the first fossils were found by Charles MacLaren (MacLaren Reference MacLaren1839). The latter recorded two poorly preserved specimens unidentifiable at the generic level: a cephalopod and a trilobite. The first fossils from the locality now known as R82 were collected and described by Salter (in Howell & Geikie Reference Howell and Geikie1861) during the first mapping of the area by the Geological Survey. Work continued in the North Esk Inlier until the end of the century, culminating with the Geological Survey Memoir by Peach & Horne (Reference Peach and Horne1899), a work of lasting quality that played a prominent role in British geology and palaeontology for decades (although the Silurian beds were still regarded there as late Silurian age). Following this, little attention was dedicated to the Pentland Hills, and it was not until Lamont (Reference Lamont1947), in which he concludes a late Llandovery age for the North Esk Inlier strata, that decisive works were starting to be published again. From then on, a more steady flux of palaeontological and geological studies were published (see Clarkson et al. Reference Clarkson, Harper, Taylor and Anderson2007 for a summary) that contributed to a better understanding of the area.
The sponges constitute a quantitatively minor component of the North Esk Inlier faunas, due largely to the fragility of their skeletons, which results in only localised conditions suitable for their preservation. This is particularly true in shallow-water environments, such as those represented in these deposits, because of elevated turbulence and high silica solubility. There has also been relatively little research on the sponge fauna. The first taxonomic descriptions of sponges from the Pentland Hills were undertaken by Hinde (Reference Hinde1883, Reference Hinde1887–1912), who described two species (Amphispongia oblonga and Plectoderma scitulum) from the Wether Law Linn Formation, and these remained the only studies until Botting (Reference Botting2004, Reference Botting, Clarkson, Harper, Taylor and Anderson2007a) reviewed and updated the fauna. In the North Esk inlier, four species of sponges are known from locality R82 [NT 149586] in the Whether Law Linn Formation and one species from a single locality on the Gutterford Burn, which is otherwise unfossiliferous (Botting Reference Botting, Clarkson, Harper, Taylor and Anderson2007a). Recent fieldwork has additionally uncovered isolated hexactine spicules from an unidentified taxon at locality R260 (lateral stratigraphic equivalent to R82), along the Deerhope Burn. This assemblage of sponges in the North Esk Inlier is preserved in many cases fully articulated, providing a perspective that is highly unusual due to both the rarity of conditions required for abrupt burial, and the transience of shallow-water siliciclastics (Botting Reference Botting, Clarkson, Harper, Taylor and Anderson2007a). The uniqueness of this shallow-water community is reflected in the fact that the majority of the species, and some genera, are only known from their type locality.
The Pentland sponge fauna is not only taxonomically distinct from similar-aged assemblages, but also morphologically distinct. In particular, the hexactine-bearing Amphispongia oblonga cannot be related to any other known species, and the distinctive architecture of Plectoderma is also difficult to relate to other reticulosans. The uniqueness of this aberrant fauna is reinforced by the discovery of another bizarre new species of sponge, described in the section on Systematic Palaeontology.
The identification and description was by JPB and YC; the specimen was prepared by VC and photographed by WRBC.
1. Geological setting
The North Esk Inlier (Fig. 1) is the largest and most westerly of the three Silurian inliers in the Pentland Hills. It forms a 6.8km2 elongate area at the head of the North Esk River. Exposure is reasonably good but is often restricted to stream valleys. Within the inlier, the sediments are steeply inclined to vertical, striking in a NE to SW direction, younging to the NW. The Silurian succession is 1500m thick. The inlier is ringed primarily by a greywacke-conglomerate of early ORS age, but a fault truncates the north-western part of the inlier and throws down Lower Carboniferous sediment of ORS facies (Clarkson & Taylor Reference Clarkson, Taylor, Clarkson, Harper, Taylor and Anderson2007).

Figure 1 (a) Location of the Pentland Hills in Scotland. (b) Geological sketch map showing the three Silurian inliers in relation to the surrounding geology. (c) Geological sketch map of the central part of the North Esk Inlier showing the most important localities (including locality R82), as described by Robertson (Reference Robertson1985). Grid reference in margin. Redrawn from Clarkson & Taylor (Reference Clarkson, Taylor, Clarkson, Harper, Taylor and Anderson2007) and Candela & Crighton (Reference Candela and Crighton2015).
The Wether Law Linn Formation is composed of calcareous mudstones, and is up to 265m thick. It has been divided by Robertson (Reference Robertson1985) into three members, mainly on the basis of their abundant, well-preserved, and varied faunas. Robertson (Reference Robertson1985, Reference Robertson1989) and Clarkson & Taylor (Reference Clarkson, Taylor, Clarkson, Harper, Taylor and Anderson2007) provided a description of the lithology. The upper part of the lower member is exposed some 50m N of the junction of Wether Law Linn with the North Esk River at locality R82 [NT 149586]. This exposure was dug up by Gary Robertson in 1983 and although partially fallen in, excellent fossil specimens may be collected from the scree (Clarkson & Taylor Reference Clarkson, Taylor, Clarkson, Harper, Taylor and Anderson2007). The base of the exposure is characterised by the Eoplectodonta penkillensis Association (Robertson Reference Robertson1989), updated as the Eoplectodonta–Visbyella Association by Clarkson & Taylor (Reference Clarkson, Taylor, Clarkson, Harper, Taylor and Anderson2007), although Visbyella is very rare and disappears after about 2m. Some 5m from the base of this section, a 0.5m-thick horizon is dominated by the brachiopod Coolinia pecten, bivalves (Palaeoneilo and Leptodesma), the peculiar mollusc Spirina, and gastropods such as Liospira. This is described as the Liospira(?) simulans–Synek(?) Association (Robertson Reference Robertson1989). Through the exposures, molluscs become more important as brachiopods decline, which indicates an upwards trend towards changing or variable salinity. The sediments of the Wether Law Linn Formation are interpreted as being deposited in a lagoonal environment that presumably became more isolated with time.
The specimen described here was collected from a loose block lying at the bottom of the succession, so its exact stratigraphic provenance is uncertain. However, the associated fauna consist of the reticulosan sponge Plectoderma scitulum (Hinde Reference Hinde1883), which is only known from locality R82 (Botting Reference Botting, Clarkson, Harper, Taylor and Anderson2007a, Reference Bottingb) in the Pentland Hills and, therefore, the loose block is indigenous, and the relatively abundant brachiopods Eoplectodonta penkillensis (Reed Reference Reed1917) and Coolinia applanata (Salter In M'Coy Reference M'Coy1846), and the ostracode Craspedobolbina (Mitrobeyrichia) impendens (Haswell Reference Haswell1865). As the brachiopods dominate the associated fauna, this block was almost certainly derived from the adjacent outcrops at the base of the section, possibly from the interval in which Visbyella disappears.
2. Repository
The specimen described here, consisting of NMS G.2010.38.1 and its counterpart NMS G.2010.38.2, is catalogued and stored at the National Museums Scotland (NMS), Edinburgh.
3. Systematic palaeontology
Class Hexactinellida Schmidt, Reference Schmidt1870
Remarks. The presence of hexactins is itself not restricted to Hexactinellida (Botting & Muir Reference Botting, Zhang and Muir2018), but the complexity of the body form and architecture suggests a more derived position than the stem-silicean reticulosans. The new sponge has typical hexactinellid-type spicules (including hypodermal pentactins, restricted to a single living family) but its complex architecture resembles only some members of the same family. Although certain assignment to the Rossellidae is not possible with the current material, it appears very likely, and a crown-group hexactinellid position seems to be reliable.
Family Rossellidae? Schulze, Reference Schulze1885
Eoghanospongia gen. nov.
Derivation of name. Named after Eoghan Candela, son of the second author, who has helped many times with collecting in the Pentland Hills.
Type species. By monotypy; Eoghanospongia carlinslowpensis sp. nov., from the Wether Law Linn Formation (Telychian), locality R82 [NT 149586], Pentland Hills, Scotland.
Diagnosis. Sponge body in form of large, shallowly convex to undulating ovoid disc with cuspate margin deflexed irregularly up or down; array of longitudinal (apparently radial) prostalial spicules with greatly enlarged proximal ray, and other rays reduced or absent, the spicule centre extruded beyond the sponge margin; fine spicule mesh of irregular tracts of small, fine hexactines and perhaps abundant fine monaxons, mostly concentric and longitudinal but sometimes oblique; upper surface of sponge with irregular sub-radial and concentric array of small depressions. An associated structure appears to be a long basal peduncle with predominantly fine monaxon spiculation.
Eoghanospongia carlinslowpensis sp. nov.

Figure 2 Eoghanospongia carlinslowpensis gen. et sp. nov. (a) NMS G.2010.38.1 (part) and (b) NMS G.2010.38.2 (counterpart), with indications of positions of detail images. (c) Detail of part showing rounded distal termination of large prostalial style. (d) Detail of counterpart showing lateral rays of prostalial pentactin projecting just beyond body wall; position shown by an arrow on image (b). (e) Central part of disc with insertion points of major spicules, and fine spicular skeleton surrounding rounded depressions (presumed exhalent canals, see arrows). (f) Axial canal remnants on longitudinal section of prostalial spicule, shown by vertical arrow. (g) Sub-helical arrangement of fine monaxons in proximal part of stalk/peduncle; position shown by an arrow on image (b). Detail of part showing: (h) diverging fine monaxons in distal part of the stalk (position shown by an arrow on image (a)); (i) transverse spicules (oriented top–bottom) and probable small hexactins in central part of the stalk (position shown by an arrow on image (a)). Scale bars=10mm (a, b); 1mm (e–i); 500μm (c, d).

Figure 3 (a) Detail from Figure 2e, including magnified box showing small hexactin-based spicules, with (b) interpretative drawing to highlight visible spicules. Only the two vertically oriented prostalia are preserved with recrystallised silica. Largest spicules with greatest relief (dark stippling) include several, with only centre of cross visible, the rest embedded in the matrix. Unstippled areas interpreted as openings of canals. Most spicules are not recognisable, and only the clearest are included. Scale bar=500μm.
Derivation of name. Derived from Carlins Lowp (meaning Witches' Leap), Scots name for the town of Carlops, the closest town to the fossil locality.
Holotype. NMS G.2010.38.1 and counterpart NMS G.2010.38.2. Only known specimen.
Diagnosis. As for the genus.
Locality. Locality R82 [NT 149586], 50m N of the junction of the Wether Law Linn and the North Esk River, Pentland Hills, Scotland.
Description. Sponge preserved as part and counterpart broken through the thickness of an oxidised (presumed originally pyritised) body wall; spicules normally dissolved, except for largest which retain siliceous composition and occasionally traces of axial canals.
Body consists of a large, slightly convex, ellipsoidal disc (Fig. 2a, b), the major axis measuring 50mm and the minor axis 40mm. Margin cuspate and irregular, with edge upturned in parts and downturned in others (polarity uncertain, but presumed majority is downturned). Disc with central apex preserved as a low mound; presumed underside not seen. Slight inflection in the surface of the disc present at nearly half-radius. Outer part of disc with strong inflexion downwards, 3–4mm high at most, before extending into marginal frill with cuspate edge, up to 5mm wide; marginal frill visible inconsistently due to uneven preservation or development.
Most obvious spicules are large, smooth, longitudinal rays up to 20mm long and up to 0.75mm in diameter, extending beyond the periphery of the disc. Some appear to be monaxons with rounded distal terminations (styles) (Fig. 2c), but at least one shows four perpendicular rays (pentactine) (Fig. 2d) that branch from a point beyond the sponge wall margin. Number of primary spicules difficult to assess with confidence due to breakage; we estimate there to have been approximately 30, not regularly spaced. Proximal end of these spicules inserted at around 1/3 of the disc radius, with some finer spicules (ray diameter 0.4mm) inserted more distally. Primary spicule rays parallel to the surface of the disc, including curvature at the marginal inflexion. The distal ends of the coronal spicules extend up to 5mm beyond the sponge margin in some cases; usually probably less.
Fine skeleton (Fig. 3) poorly preserved and difficult to characterise, but consists of at least some very fine-rayed hexactines (ray diameter approximately 30μm), and probably abundant fine monaxons. Hexactin-based spicules include numerous sizes, including some with ray length less than 100μm. Fine spicules with irregular longitudinal and concentric tract development locally, but oblique spicules or tracts also present, and overall skeletal organisation is structured around openings to canals on oral surface (Fig. 3). Dense longitudinal monaxon array visible at margins of frill. Any regular skeletal architecture is distorted by the presence of numerous small depressions on the upper surface (often corresponding to similar small mounds on lower surface). Depressions with local radial or concentric alignment, but not consistent. Diameter of depressions up to approximately 1mm (increasing slightly in average towards outer part), and spacing varies from six in 5mm centrally, to four in 5mm in the outer part of the disc.
Central quarter of the disc characterised by densely packed, irregularly arranged depressions only; skeletal structure unclear. Middle part of disc characterised by the insertion points of the major spicules, and clearest fine spicule tract development; depressions locally in concentric arrangement (Figs 2e, 3). Outer section of primary disc with depression arranged radially, constrained by primary spicule rays, often with two rows of depressions between each pair of spicule rays.
Primary spicules typically show a laminar internal structure, although the fine-scale laminae present in extant species of hexactinellid sponges like Euplectella (Aizenberg et al. Reference Aizenberg, Weaver, Thanawala, Sundar, Morse and Fratzl2005) are not preserved. Evidence for an axial canal is preserved in only a few examples (Fig. 2f); the rest are destroyed by recrystallisation.
Associated with the sponge body, curving towards and perhaps attached to the centre of the underside (but unexposed) is either a second unique sponge specimen, or more probably a peduncle or stalk. Exposed length is 80mm, with implied length exceeding 110mm, if connected with the sponge body. Spicules are predominantly fine monaxons, 2–2.5mm long and 15–20μm in diameter, with helical arrangement in narrowest part (Fig. 2g), diverging unevenly distally (Fig. 2h). Some transverse spicules may also be present, but are inconclusive (Fig. 2i). It is not possible to determine whether the spicules are fused, due to the coarse preservation. The margins of the structure are more heavily mineralised than the interior, and spicules extend through the (very thin) sediment thickness rather than being restricted to a planar wall, but do not show the dense internal spiculation of an anchoring root tuft. The spicules are much finer and less densely packed than in the proximal part of Plectoderma, which occurs at the same locality. This combination of features demonstrates that the structure is a pedunculate stalk composed of spicules and soft tissue, rather than a dense, purely spiculate basal tuft.
Remarks. Uncertainty over whether the stalk-like structure is part of the sponge body hinders the interpretation, but it is unlikely that two unique specimens would co-occur on the same slab in such a way by chance. The preservation of this structure, including the presence of sediment inside it, supports an interpretation as a peduncle or stalk structure that is very different to those known for co-occurring species (Botting Reference Botting, Clarkson, Harper, Taylor and Anderson2007a), and implies that it was part of the sponge body and with spicules embedded in soft tissue, rather a purely spicular basal root tuft. This interpretation is used for the reconstruction (Fig. 4). The morphology of the sponge body, being flat and without an apical osculum, is also best interpreted as being raised off the sea floor (by analogy with modern hexactinellids such as Caulophacus Schulze, Reference Schulze1886). Because of all these factors, assigning this structure to Eoghanospongia is highly likely, but cannot be considered categorical from the material available, and cannot be confirmed without significantly damaging the specimen.
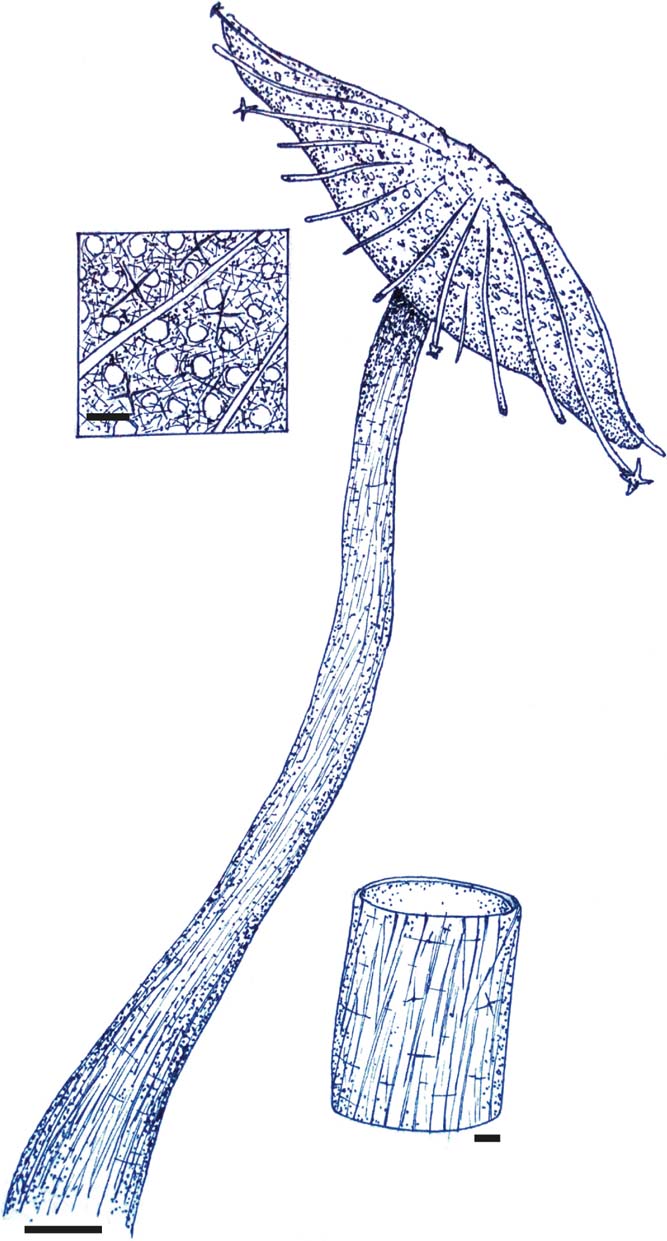
Figure 4 Interpretative reconstruction of Eoghanospongia carlinslowpensis gen. et sp. nov., including the probable peduncle, the connection of which to the sponge body cannot be confirmed without damaging the holotype. Upper detail views show the structure of the upper (convex) surface, including presumed exhalent canal openings between large prostalial styles/pentactins (smallest spicules only rarely preserved). Lower detail shows hollow peduncle/stalk, with dominantly sub-longitudinal monaxon (diactin) spiculation in thin wall. Scale bars=10 mm in main image, 1 mm in detail views.
Although only a single specimen is yet known, the good preservation characteristic of the Wether Law Linn Formation at locality R82 allows a reasonable characterisation of this new taxon, which is distinct from all previously described sponges in several ways. Classification of the new species into higher groups is challenging because of the unique character combination, and the lack of any similar described forms, and we therefore discuss several known groups as possible candidates for comparison.
Superficially, the morphology of the specimen described here is crudely similar to Choia and Choiaella, two members of the cosmopolitan (but probably paraphyletic; Botting et al. Reference Botting and Muir2013) family Choiidae, the geographic range of which extended from South China, Siberia, and Laurentia (including peri-Laurentian margins) to Gondwana (Finks & Rigby Reference Finks, Rigby and Kaesler2004a; Botting Reference Botting, Clarkson, Harper, Taylor and Anderson2007a, Reference Bottingb; Beresi et al. Reference Beresi, Botting and Clarkson2010). Choia consisted of a flattened, circular to elliptical bowl, with a central disc of longitudinal (apparently radial) monaxons, enhanced by an array of large coronal monaxons radiating from the peripheral part and extending beyond the central disc. Choiaella, first discovered from Cambrian rocks in China, was described (Beresi et al. Reference Beresi, Botting and Clarkson2010) from the Late Ordovician of the Southern Uplands, Scotland, as well as from the locality R82 discussed here (Botting Reference Botting, Clarkson, Harper, Taylor and Anderson2007a). Choiaella is morphologically similar to Choia in possessing a radiating, thatched central disc, but lacks coronal spicules.
In contrast, Eoghanospongia gen. nov. possesses a radial array of large spicules, with an irregular, sub-concentric fine skeletal mesh rather than a simple radiating thatch of small spicules covering the central disc; furthermore, the coronal spicules are larger and less abundant than in Choia. In at least some cases, these spicules are also pentactins, implying that the projecting styles also originated as pentactins with suppressed rays. Although small hexactins are present within the central disc of some Choia-like sponges (Botting et al. Reference Botting and Muir2013, p. 9), there is no indication of the coronal spicules being anything other than monaxons in any species. The similarity with choiids is, therefore, entirely superficial.
The new species is characterised, in part, by thin, parallel, concentric monaxon (and hexactine) spicules combined with a radial fabric to form a loosely quadrate mesh. This feature is loosely comparable to the structure of dictyospongioid hexactinellids (Finks & Rigby Reference Finks, Rigby and Kaesler2004b), although much less regularly organised, and the robust radial spicules have never previously been described in a dictyospongioid.
Hall & Clarke (Reference Hall and Clarke1898) described and illustrated numerous reticulate sponges of the family Dictyospongidae, most of which share a very similar fundamental structure, and differ primarily in body form. Their fine skeletal structure is often poorly preserved, but in overall form, some show a degree of similarity to the new species. For example, the basal area of Phragmodictya (a genus restricted to the Carboniferous) is characterised by an oval disc with a ‘very fine reticulum' (Hall & Clarke Reference Hall and Clarke1898, p. 172, pl. 64, fig. 5). As in all dictyosponges, the reticulum is composed of a strictly radial and concentric spicular grid, the radial striae converging toward the centre of the disc. Phragmodictya also lacks the primary radial spicules and the depressions diagnostic of Eoghanospongia.
Not only does the fine skeletal architecture of the disc in Eoghanospongia lack the regularity of the dictyosponges, it also cannot be easily reconciled with the simple, homogeneously expanding growth pattern of reticulosans generally (Botting Reference Botting2003). There are also no plausible counterparts in known dictyosponges for the large radial pentactines. Despite the superficial similarity in the fundamental body form and apparent wall structure, the detailed differences refute any assignment relating the new sponge to dictyosponges.
The body form of the new sponge, if the associated stalk region is a part of the sponge, is strikingly similar to the living rossellid hexactinellid Caulophacus (Schulze Reference Schulze1886; Tabachnick Reference Tabachnick, Hooper and Van Soest2002). In this case, the stalk is essential to raising the flattened, mushroom-shaped-to-convolute discoidal body above the sea floor, and the same could be presumed for the body form described here. Caulophacus also has an irregular surface sculpture, with numerous small depressions associated with oscula. However, the skeletal structure of Caulophacus consists of a regular, fine grid of hexactins that is quite distinct from the tract development seen here. Although there is no exact counterpart for the large projecting pentactins in Caulophacus, such hypodermal pentactine prostalia are indeed limited in modern faunas to the Rossellidae (Tabachnick Reference Tabachnick, Hooper and Van Soest2002). Although the origins of the family are believed to be in the Late Palaeozoic based on molecular clock data (Dohrmann et al. Reference Dohrmann, Vargas, Janussen, Collins and Wörheide2013), a putative stem-group rossellid, Matteolaspongia hemiglobosa, also featuring pentactine prostalia, has now been described by the Late Ordovician of China (Botting et al. Reference Botting and Muir2018). A rossellid affinity is, therefore, possible, despite the early date and unusual shallow-water environment. The extremely robust prostalia in Eoghanospongia differ from all described rossellid examples, but there is a wide variation across the family. The morphology (including multiple exhalent depressions on the upper surface), stalk structure (composed dominantly of monaxons forming a thin wall), and presence of these prostalia all suggest a remarkable similarity to Caulophacus, or to related sponges within the rossellid crown group. Without fine detail of the skeletal architecture and spicule morphology (ideally including microscleres), however, this morphology is strongly suggestive but cannot be considered conclusive.
No other described fossil or modern sponges show particular similarity to the new species, and Eoghanospongia must, therefore, be left in open taxonomy at this stage, with strong suggestions of a rossellid affinity.
4. Discussion
Interpreting the life position and autecology of uncommon fossils without close living relatives is often problematic. The new genus is described based on only a single specimen, and the proposed interpretation is, therefore, subject to some uncertainty. There are particular difficulties in the interpretation of fossils where the morphology of the sponge differs from the normal vasiform body form with a clear osculum; Choia, a Cambro–Ordovician genus that is similar in outline to Eoghanospongia, is a textbook example of the problems encountered. Although many specimens have been collected and described since Walcott (Reference Walcott1920), it was not until Botting (Reference Botting2007b) that the conventional reconstructed orientation (see Rigby Reference Rigby1986) was inverted. A similar issue exists with the new taxon, in that there are no definitive indications of the life orientation (except for the stalk, if attached), and it could be assumed that the stable orientation (convex upward) represents the life position. However, the functionality of the sponge body plan makes such an orientation very difficult, especially as the direction of flexure of the rim varies around the margin. If the associated stalk is also a part of the sponge, then the orientation is fixed as being convex-upwards, as in the modern Caulophacus.
The depressions that may represent oscula or exhalent canal openings are regularly developed over much of the surface of the disc, arranged radially and concentrically. However, the central 20% of the disc does not show any clear geometric pattern. Some of the depressions appear to be locally radially arranged, whereas others are organised in short arcs. If these do represent oscula, then the morphological similarity to the extant Caulophacus is reinforced.
The evidence listed above favours the sponge being reconstructed with the convex apex upwards, and a stalked mode of life resembling that of Caulophacus. This would be unprecedented in the Palaeozoic, or in shallow-water environments, and indicates a highly derived morphology. Its co-occurrence with other unique sponges such as Amphispongia (Botting Reference Botting2004) indicates that significant evolution of unusual morphologies was occurring in shallow-water environments, as represented by the Pentland Hills deposits. Normally, such environments are hostile to sponge preservation due to dissolution and turbulence, but unusual chemical conditions and rapid burial occasionally yield well-preserved fossils in even the most unlikely environments (see Botting Reference Botting2005).
The new species further supports previous suggestions that dramatic diversification was occurring among sponges in shallow-water habitats during the Ordovician (Botting Reference Botting2005; Muir et al. Reference Muir, Botting, Carrera, Beresi, Harper and Servais2013), and highlights the limitations of our current knowledge of sponge evolution within these groups. The differences between the new species and any potential ancestors or close relatives, as with Amphispongia (see Botting Reference Botting2004), Haljalospongia (Botting & Rhebergen Reference Botting and Rhebergen2011), and others (e.g., Rigby & Webby Reference Rigby and Webby1988), implies that we currently have no knowledge of large parts of sponge evolution from this interval, especially regarding the stem group (and perhaps early crown group) of Hexactinellida. If we are correct in interpreting the specimen as a plausible crown-group rossellid allied to Caulophacus, or at least crown-group lyssacinosidan, then this builds on other results from China (Botting et al. Reference Botting and Muir2018) that exacerbate the problems of the missing record of sponge diversification, as it implies that many modern groups must have arisen cryptically, and left almost no trace in the fossil record. The presence of a possible derived hexactinellid in a shallow-water, lagoonal setting is in complete contradiction of their modern distribution pattern (deep oceans and caves, both palaeontologically inaccessible), and makes explanation of the missing record even more problematic. Changing environment inhabitation is a common phenomenon within hexactinellid-like sponges generally (Mehl Reference Mehl1996; Mehl-Janussen Reference Mehl-Janussen1999), but for modern-type hexactinellids, shallow-water colonisation is tied to particular intervals of high sea level, such as the Jurassic and Cretaceous periods.
5. Conclusions
A new fossil sponge, Eoghanospongia carlinslowpensis gen. et sp. nov., is described from the Telychian (early Silurian) of the Pentland Hills, Scotland. It has a morphology unique in the fossil record, and among living taxa most closely resembles that seen in rossellid hexactinellid Caulophacus. However, there are significant differences that preclude a close relationship to known taxa, and the only fossil record of Caulophacus is from spicules in Tertiary radiolarite deposits from New Zealand (Hinde & Holmes Reference Hinde and Holmes1892). Although a long and cryptic deep-sea history of such a genus is possible, we prefer to consider the new fossil to represent an early rossellid that may have evolved a similar morphology convergently. The implications of the new species include further support for the contention that the family Rossellidae has an extremely deep history, and reiteration of the incompleteness of some aspects of the sponge fossil record. The most surprising aspect is the presence of what is now a deep-water (abyssal) sponge in a shallow-water lagoon. This may be an ecological analogue of the recent discovery of living hexactinellids in shallow-water caves in the Mediterranean (Vacelet Reference Vacelet, Uiblein, Ott and Stachowitsch1996), a habitat that mimics the dark and quiet-water environment of the deep oceans.
6. Acknowledgements
JPB thanks the National Museums Scotland for support through a Visiting Research Fellowship. YC thanks Lyall Anderson for helping with Scots vocabulary. The manuscript was improved by constructive comments from two anonymous referees.




