

















Crossref Citations
This article has been cited by the following publications. This list is generated based on data provided by Crossref.
STOLK, W. A.
RAMAIAH, K. D.
VAN OORTMARSSEN, G. J.
DAS, P. K.
HABBEMA, J. D. F.
and
DE VLAS, S. J.
2004.
Meta-analysis of age-prevalence patterns in lymphatic filariasis: no decline in microfilaraemia prevalence in older age groups as predicted by models with acquired immunity.
Parasitology,
Vol. 129,
Issue. 5,
p.
605.
Ramaiah, Kapa D.
and
Das, Pradeep K.
2004.
Mass drug administration to eliminate lymphatic filariasis in India.
Trends in Parasitology,
Vol. 20,
Issue. 11,
p.
499.
EGAN, C. A.
HOUSTON, K. M.
ALCOCER, M. J. C.
SOLOVYOVA, A.
TATE, R.
LOCHNIT, G.
McINNES, I. B.
HARNETT, M. M.
GEYER, R.
BYRON, O.
and
HARNETT, W.
2005.
Lack of immunological cross-reactivity between parasite-derived and recombinant forms of ES-62, a secreted protein of Acanthocheilonema viteae.
Parasitology,
Vol. 132,
Issue. 02,
p.
263.
Dreyer, Gerusa
Addiss, David
and
Norões, Joaquim
2005.
Does longevity of adult Wuchereria bancrofti increase with decreasing intensity of parasite transmission? Insights from clinical observations.
Transactions of the Royal Society of Tropical Medicine and Hygiene,
Vol. 99,
Issue. 12,
p.
883.
Stolk, Wilma A
de Vlas, Sake J
and
Habbema, J Dik F
2005.
Anti-Wolbachia treatment for lymphatic filariasis.
The Lancet,
Vol. 365,
Issue. 9477,
p.
2067.
Goodridge, Helen S.
Stepek, Gillian
Harnett, William
and
Harnett, Margaret M.
2005.
Signalling mechanisms underlying subversion of the immune response by the filarial nematode secreted product ES‐62.
Immunology,
Vol. 115,
Issue. 3,
p.
296.
HARNETT, W.
GOODRIDGE, H. S.
and
HARNETT, M. M.
2005.
Subversion of immune cell signal transduction pathways by the secreted filarial nematode product, ES-62.
Parasitology,
Vol. 130,
Issue. S1,
p.
S63.
Oqueka, Tim
Supali, Taniawati
Ismid, Is Suhariah
Purnomo
Rückert, Paul
Bradley, Mark
and
Fischer, Peter
2005.
Impact of two rounds of mass drug administration using diethylcarbamazine combined with albendazole on the prevalence of Brugia timoriand of intestinal helminths on Alor Island, Indonesia.
Filaria Journal,
Vol. 4,
Issue. 1,
Stolk, Wilma A
de Vlas, Sake J
and
Habbema, J Dik F
2006.
Advances and challenges in predicting the impact of lymphatic filariasis elimination programmes by mathematical modelling.
Filaria Journal,
Vol. 5,
Issue. 1,
De Kraker, Marlieke E. A.
Stolk, Wilma A.
Van Oortmarssen, Gerrit J.
and
Habbema, J. Dik F.
2006.
Model‐based analysis of trial data: microfilaria and worm‐productivity loss after diethylcarbamazine–albendazole or ivermectin–albendazole combination therapy against Wuchereria bancrofti.
Tropical Medicine & International Health,
Vol. 11,
Issue. 5,
p.
718.
Harnett, William
and
Harnett, Margaret M
2006.
FILARIAL NEMATODE SECRETED PRODUCT ES‐62 IS AN ANTI‐INFLAMMATORY AGENT: THERAPEUTIC POTENTIAL OF SMALL MOLECULE DERIVATIVES AND ES‐62 PEPTIDE MIMETICS.
Clinical and Experimental Pharmacology and Physiology,
Vol. 33,
Issue. 5-6,
p.
511.
Ramaiah, K.D.
Vanamail, P.
and
Das, P.K.
2007.
Changes in Wuchereria bancrofti infection in a highly endemic community following 10 rounds of mass administration of diethylcarbamazine.
Transactions of the Royal Society of Tropical Medicine and Hygiene,
Vol. 101,
Issue. 3,
p.
250.
Hise, Amy G.
Daehnel, Katrin
Gillette-Ferguson, Illona
Cho, Eun
McGarry, Helen F.
Taylor, Mark J.
Golenbock, Douglas T.
Fitzgerald, Katherine A.
Kazura, James W.
and
Pearlman, Eric
2007.
Innate Immune Responses to Endosymbiotic Wolbachia Bacteria in Brugia malayi and Onchocerca volvulus Are Dependent on TLR2, TLR6, MyD88, and Mal, but Not TLR4, TRIF, or TRAM.
The Journal of Immunology,
Vol. 178,
Issue. 2,
p.
1068.
Harnett, William
and
Harnett, Margaret M.
2008.
Therapeutic immunomodulators from nematode parasites.
Expert Reviews in Molecular Medicine,
Vol. 10,
Issue. ,
Harnett, William
and
Harnett, Margaret M.
2008.
Parasitic nematode modulation of allergic disease.
Current Allergy and Asthma Reports,
Vol. 8,
Issue. 5,
p.
392.
STOLK, WILMA A.
DE VLAS, SAKE J.
BORSBOOM, GERARD J. J. M.
and
HABBEMA, J. DIK F.
2008.
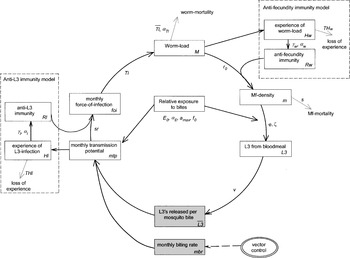
LYMFASIM, a simulation model for predicting the impact of lymphatic filariasis control: quantification for African villages.
Parasitology,
Vol. 135,
Issue. 13,
p.
1583.
HARNETT, W.
and
HARNETT, M. M.
2008.
Lymphocyte hyporesponsiveness during filarial nematode infection.
Parasite Immunology,
Vol. 30,
Issue. 9,
p.
447.
Grabitzki, Julia
and
Lochnit, Günter
2009.
Immunomodulation by phosphocholine—Biosynthesis, structures and immunological implications of parasitic PC-epitopes.
Molecular Immunology,
Vol. 47,
Issue. 2-3,
p.
149.
Harnett, William
and
Harnett, Margaret M.
2009.
Pathogen-Derived Immunomodulatory Molecules.
Vol. 666,
Issue. ,
p.
88.
Harnett, M M
Melendez, A J
and
Harnett, W
2010.
The therapeutic potential of the filarial nematode-derived immunodulator, ES-62 in inflammatory disease.
Clinical and Experimental Immunology,
Vol. 159,
Issue. 3,
p.
256.