Introduction
Sirex noctilio Fabricius (Hymenoptera, Siricidae) is a woodwasp native to Eurasia (Taeger & Blank, Reference Taeger and Blank2008). It has been accidentally introduced into multiple locations in the Southern Hemisphere, where it has frequently become an important pest in plantations of pine species that originate from the Northern Hemisphere (Slippers et al., Reference Slippers, de Groot and Wingfield2012). Most recently, in 2004, the species was also discovered in the USA (Hoebeke et al., Reference Hoebeke, Haugen and Haack2005) and Canada (de Groot et al., Reference de Groot, Nystrom and Scarr2006). Despite a century of research and well-established ongoing management programmes, control of S. noctilio in the Southern Hemisphere remains unpredictable and spread continues to new areas (Slippers et al., Reference Slippers, Hurley and Wingfield2015). Understanding why a species is not a pest in one ecosystem can help to understand how it becomes a pest in other ecosystems (Ayres et al., Reference Ayres, Pena, Lombardo and Lombardero2014).
S. noctilio larvae rely for their development on the alteration of wood by their specialized symbiotic fungus, Amylostereum areolatum (Chaillet ex Fries) Boidin, a basidiomycete (Talbot, Reference Talbot1977; Madden, Reference Madden and Berryman1988; Thompson et al., Reference Thompson, Bodart, McEwen and Gruner2014). This fungal species is inoculated into trees during oviposition, together with a phytotoxic venom containing noctilisin (Coutts, Reference Coutts1969a , Reference Coutts b ; Coutts & Dolezal, Reference Coutts and Dolezal1969; Bordeaux et al., Reference Bordeaux, Lorenz, Johnson, Badgett, Glushka, Orlando and Dean2014). The combined action of the insect and its symbiont may cause up to 80% mortality in pine plantations of the invaded area (Talbot, Reference Talbot1977; Neumann et al., Reference Neumann, Morey and McKimm1987). Control strategies have included removal of infested trees and silvicultural practices that limit tree competition (Neumann et al., Reference Neumann, Morey and McKimm1987; Dodds et al., Reference Dodds, Cooke and Gilmore2007, Reference Dodds, Cooke and Hanavan2014), but the most widely used strategy has been classic biological control (Hurley et al., Reference Hurley, Slippers and Wingfield2007). Several species of enemies have been introduced to control populations of S. noctilio. Hymenoptera parasitoids that have been introduced from the native range of S. noctilio include Ibalia leucospoides leucospoides (Hochenwarth) and Rhyssa persuasoria persuasoria (Linnaeus) and from North America Megarhysa nortoni nortoni (Cresson). These species have become established in most regions where they were introduced and are thought to play a role in reducing populations of S. noctilio (Hurley et al., Reference Hurley, Slippers and Wingfield2007; Collett & Elms, Reference Collett and Elms2009; Cameron, Reference Cameron, Slippers, de Groot and Wingfield2012). However, the most widely employed control agent is the parasitic nematode Deladenus (Beddingia) siricidicola Bedding, which can produce parasitism of up to 90%, although with substantial variation among years and areas (Hurley et al., Reference Hurley, Slippers and Wingfield2007).
There has been great growth in knowledge of S. noctilio and its associated species in the Southern Hemisphere (e.g., Slippers et al., Reference Slippers, de Groot and Wingfield2012, Reference Slippers, Hurley and Wingfield2015). However, there have been relatively few studies of S. noctilio in its native range and the species has been consistently characterized as rare and at most a secondary pest that colonizes suppressed trees or trees weakened from primary damage by other insects, fungi or abiotic effects. Ratzeburg (Reference Ratzeburg1844) reported woodwasps from a pine forest that was experiencing severe defoliation; he regarded them as secondary pests but suggested that they could occasionally kill trees if they reach high local abundance. Similarly, Hartig (Reference Hartig1860) described woodwasps as far less important than bark beetles. Later descriptions from Europe have continued to describe siricids as mainly scavengers of trees dying for other reasons (Judeich & Nitsche, Reference Judeich, Nitsche, Judeich, Ratzeburg and Nitsche1895; Chrystal, Reference Chrystal1928; Benson, Reference Benson1943; Wolf, Reference Wolf1969a ). Spradbery & Kirk (Reference Spradbery and Kirk1978) provided the most comprehensive study of S. noctilio in Europe. More recently, Tarasco & Notte (Reference Tarasco and Notte2003) reported some damage by S. noctilio in Pinus pinea in Italy, but Wermelinger et al. (Reference Wermelinger, Rigling, Mathis and Dobbertin2008) described them as restricted to greatly weakened trees.
There is similarly limited knowledge of siricids on the Iberian Peninsula. The most prominent works are by Hall (Reference Hall1968) and Spradbery & Kirk (Reference Spradbery and Kirk1978). Previous literature referred to occasional records of Siricidae on the Iberian Peninsula, but often with questionable or erroneous identifications (Llorente & Gabuyo, Reference Llorente and Gabuyo1984), as has also occurred in other areas of Europe (Chrystal, Reference Chrystal1928). In Spain, S. noctilio and S. juvencus have often been confused (Ceballos, Reference Ceballos1963). The main hosts of S. juvencus are various species of Picea (Wolf, Reference Wolf1969a ; Spradbery & Kirk, Reference Spradbery and Kirk1978; Schiff et al., Reference Schiff, Goulet, Smith, Boudreault, Wilson and Scheffler2012) and these tree species are not naturally distributed in Spain except as ornamentals in gardens and some small recent plantations in the Basque Country and Pyrenees (IFN4, 2013). According to Kirk (Reference Kirk1974) and Spradbery & Kirk (Reference Spradbery and Kirk1978), S. noctilio is the only species of Sirex occurring in the Mediterranean bioclimatic area. The Fauna Europaea Organization cites S. juvencus as absent in the Spanish mainland (Taeger & Blank, Reference Taeger and Blank2008).
Although it is frequently assumed that most invasive species exhibit enhanced performance in their introduced range, there are few tests (Parker et al., Reference Parker, Torchin, Hufbauer, Lemoine, Alba, Blumenthal, Bossdorf, Byers, Dunn, Heckman, Hejda, Jarošík, Kanarek, Martin, Perkins, Pyšek, Schierenbeck, Schlöder, van Klinken, Vaughn, Williams and Wolfe2013). In the case of S. noctilio these comparisons are currently impossible because the biology of native populations is too poorly known. Here we report studies of the abundance, basic biology, host use patterns and natural enemies of S. noctilio on the Iberian Peninsula, where it is native and rare. We address the question of why it is rare by summarizing the information in a preliminary life table that permits comparisons of effects on abundance from different sources. This information may help to understand what is different in the population dynamics of S. noctilio in the Southern Hemisphere, which may suggest strategies for control and management.
Material and methods
Study area
Our studies were conducted in Galicia (located in Northwestern Spain), which is a major region of commercial forestry. The most common pine species in the area are the native maritime pine (Pinus pinaster Aiton) which occupies 217,281 ha of pure stands, followed by the non-native Pinus radiata D. Don with 96,177 ha and finally the native Pinus sylvestris L. with 32,736 ha (IFN4, 2013). To locate study sites, we searched pine stands of all three species during spring of 2013 to locate stands supporting detectable populations of S. noctilio (as evidenced by emergence holes and resin drips from attacks by S. noctilio; Ayres et al., Reference Ayres, Pena, Lombardo and Lombardero2014). We located populations of S. noctilio within four different stands with pine plantations in the province of Lugo. Two stands contained P. pinaster: Pena de Rodas (43.121°N, 7.679°W; tree age = 16 years, DBH ± SD of attacked trees = 12 ± 2 cm) and Cova da Serpe (43.097°N, 7.753°W; 23 years old, DBH = 12 ± 2 cm). One stand contained P. radiata: Begonte (43.201°N, 7.691°W; 19 years old, DBH = 10 ± 3 cm), and one contained P. sylvestris: Faro (42.631°N, 7.892°W; 23 years old, DBH = 11 ± 3 cm). In March of 2013, and again in March of 2014, we located individual trees within each stand that had been attacked by S. noctilio during the previous flight season (late summer or early autumn of 2012 or 2013). A total of 134 attacked trees were located based on external drippings of resin; we confirmed that the attacks were from S. noctilio based on characteristic drillings of the inner bark from female ovipositors. In July of 2013 and again in 2014, we noted whether or not each tree was still alive or had died since March, then felled the attacked trees, cut them into logs and transported them to emergence containers within a building in Lugo. The number of study trees in 2013 and 2014 (respectively) were 26 and 26 from Cova da Serpe, 23 and 0 from Pena de Rodas, 13 and 14 from Begonte, and 19 and 13 from Faro. The bole of each tree (from just above ground to the base of the live crown) was cut into 8–23 logs (depending on tree height) of 60–70 cm length. Each log was labelled individually to record its position in the bole of the tree. Before being placed in emergence cans, the logs were measured (length and diameter), waxed on both ends and examined to confirm the absence of emergence holes.
Genetic analyses of S. noctilio
A subset of the collected Galician wasps was sequenced to confirm species identity. This was done using the mitochondrial cytochrome c oxidase subunit I (COI) locus as described by Wooding et al. (Reference Wooding, Wingfield, Hurley, Garnas, de Groot and Slippers2013). Sequences were deposited in GenBank under accession numbers KT780457, KT780458, KT780459 and KT780460. Sequencing data were parsed by JModelTest v. 0.1.1 (Posada, Reference Posada2008) to identify the best nucleotide substitution model, followed by the maximum likelihood analysis through PhyML 3.0.1 inputting the model parameters obtained (Guindon & Gascuel, Reference Guindon and Gascuel2003).
Flight period, natural enemies and body size
Cans were checked for emerging insects every two days after emergence started until late November when emergences ceased. We resumed checking for new emergences the following summer after it became evident that additional S. noctilio were emerging in the second flight season after oviposition. We collected all individual insects emerging from logs, including S. noctilio, its natural enemies and associates. For each adult S. noctilio, we measured body size with three metrics (length from head to cornus, tibia length, and pronotum width) and dissected the abdomens to search for parasitic nematodes in the gonadal tissue. We collected additional measurements on a subset of 46 females to count the total number of eggs produced and examine the eggs for infective nematodes.
Log dissections
In the spring of 2014, from the attacked trees that we felled in 2013, we examined and dissected a subset of 171 bolts from 11–13 trees per site. Most trees were represented by 2–3 logs, but for 1–3 trees per site we dissected all of the logs (9–13). Before dissection, we: (1) estimated the xylem moisture content (≈percentage) of each log as the average of 2 measurements per log using a Delmhorst RDM-3 moisture meter (Delmhorst Instrument Co. Towaco NJ); and (2) recorded the number of points on the surface of the log that had produced drippings of resin from drillings by S. noctilio. Then we removed the bark, counted the number of oviposition galleries from individual pairs of the bark beetle Tomicus piniperda (L.) (Coleoptera, Curculionidae), and counted and identified any other insects that had colonized the inner bark. Then, by examination of the full exterior surface of the xylem on each log, we marked and counted the individual attacks from S. noctilio (each from one female wasp at one location on the tree) and recorded the number of drillings (usually 1–3) associated with each attack. Each drilling was scored as filled with oleoresin or not. We also estimated the total proportion of surface area of each log with visible blue staining (characteristic of Ophiostomatoid fungi). Some of the drills from S. noctilio induced the formation of a visible lesion in the xylem tissue surrounding it. These lesions matched the description of Coutts & Dolezal (Reference Coutts and Dolezal1966), who interpreted them as a barrier of polyphenols that represent a defensive reaction of the trees against the mutualistic fungus of S. noctilio (A. areolatum). We scored each drill for whether it had or had not induced a lesion. We recorded the number of exit holes from each log, each representing the emergence of an adult S. noctilio or an adult of one of its parasitoids, which had developed from feeding on a larva of S. noctilio. We also measured the diameter of exit holes.
After measurements were completed from the outer surface of logs, we cut the logs twice transversely (at ≈20% of the total length from each end) to expose two cross-sectional surfaces that could be examined for the characteristic white staining of wood (‘white rot’) from A. areolatum and the presence of blue stain from Ophiostoma (blue–grey colour from pigments produced by O. minus and O. ips). We took a digital photograph of both exposed surfaces on each log and analysed them with ImageJ software (Schneider et al., Reference Schneider, Rasband and Eliceiri2012) to quantify the proportion of cross-sectional area occupied by both A. areolatum and Ophiostoma spp.
Statistical analyses
We used maximum likelihood estimation and log-likelihoods (VGAM package for R; Yee, Reference Yee2015) to compare the fit of the frequency distribution of wasp emergence holes per tree to four competing binomial models: Poisson, zero-inflated Poisson, negative binomial and zero-inflated negative binomial. We evaluated patterns among sites and trees in the adult size of S. noctilio with a statistical model that included site, sex and individual trees within site, with trees as a random effect. We tested for differences among sites in sex ratio of S. noctilio, and the incidence of nematode parasites, with Pearson chi-square statistics from contingency analyses. We estimated the proportion of S. noctilio killed by wasp parasitoids as the slope of a regression (with intercept = zero) of number of emerging parasitoids per tree vs. the number of S. noctilio plus parasitoids emerging per tree (Cochran, Reference Cochran1977).
Except as noted, we analysed results from log dissections with a general linear model that included site, tree status (alive or dead) and tree (as a random effect, nested within site and tree status); logs were nested within tree to avoid pseudoreplication. To satisfy assumptions of normality, we applied square root transformations to log-specific measurements of attacks by T. piniperda, attacks by S. noctilio, total drills, attacks/dm2, drills/dm2, and total exit holes, and we applied arcsin transformations to proportions of log area (surface area and cross-sections) with blue stain, proportion of log cross-sections with white rot, and proportions of drills that were filled with resin or produced lesions. We compared the average number of drills per attack among sites with a linear regression ratio estimator (Cochran, Reference Cochran1977) using logs as sample units. To further evaluate hierarchical patterning in drills per attack, we also performed an analysis that explicitly considered each of 3,256 attacks distributed across 171 logs from 11 to 13 trees at each of four sites; the response variable, number of drills per attack, was evaluated with a generalized linear mixed model (Poisson link function) that included site, tree (as a random effect nested within site) and log within tree (as a random effect nested within site, tree status and tree) (Bates et al., Reference Bates, Maechler, Bolker and Walker2014). Except where indicated, other analyses were performed with JMP, version 11.0, SAS Institute, Cary, NC. Because we have unequal sample size we reported least-square means (lsmeans) throughout.
Results
Adult morphology and life history of S. noctilio in Galicia
Adult females from our study population had darkly tinted wings, black femora and pale tibiae, as has been described for other populations of S. noctilio from Southern Europe (Schiff et al., Reference Schiff, Goulet, Smith, Boudreault, Wilson and Scheffler2012). Sequencing of the COI locus and subsequent maximum-likelihood analyses confirmed that the wasps obtained were S. noctilio.
Generation development time of S. noctilio in our study populations was usually 1 year (72%), but some individuals required 2 years (24%) or even 3 years (4%). In fall of 2013, 1 year after our study trees were attacked, we collected a total of 187 emerging adults (138 males and 49 females): median emergence dates (and 10th–90th percentiles) for males and females were 20 September (2 September–23 October) and 30 September (4 September–30 October), respectively. The next year, from 17 July to 7 September (logs held at room temperature), a second emergence occurred from the same logs (47 males and 15 females). During the winter of 2014–15, while dissecting the same logs for other measurements, we observed another ten S. noctilio larvae that were early to mid-instar (2–5 mm long) and destined to emerge no sooner than summer of 2015 (3 years after oviposition by parental females).
Adult body size was quite variable with females tending to be longer than males: overall means ± SE (10th–90th percentile) = 20.0 ± 0.5 (14–25) and 17.3 ± 0.3 (12–24) mm, respectively. Males emerging the 2nd year after attack (in 2014 from trees attacked in 2012) were larger than those that emerged from the same trees in the 1st year: mean ± SE = 14.3 ± 0.4 vs. 19.5 ± 0.8 mm for 1st- and 2nd-year emergence, respectively. However, female length did not differ between those emerging one vs. 2 years after oviposition: mean ± SE = 20.0 ± 0.7 vs. 20.5 ± 0.9 mm. There was no difference in body size of either sex between those emerging from trees attacked in 2012 vs. 2013. There were no apparent differences in body size among sites: F 3,137 < 1.1, P > 0.3 for effects of site and site × sex (for adults emerging in 2013 from trees attacked in 2012, where all sites and both sexes were represented). There was modest variation among trees in male body size (23% of total variance; 95% confidence interval for variance = 0.3–11.2; n = 172 males from 21 trees) but not in female body size (8% of total variance; estimated variance = −2.8 to 5.5; n = 64 females from 14 trees). The dbh of trees was unrelated to adult body size (P > 0.6; n = 14 and 21 trees for females and males) (fig. 1, Supplemental materials).

Fig. 1. Frequency distribution of the production of S. noctilio adults from attacked trees Data are for 139 trees representing four sites, three species and 2 years of attacks. Production per tree was standardized to the average volume of 50 dm3 of logs per attacked tree.
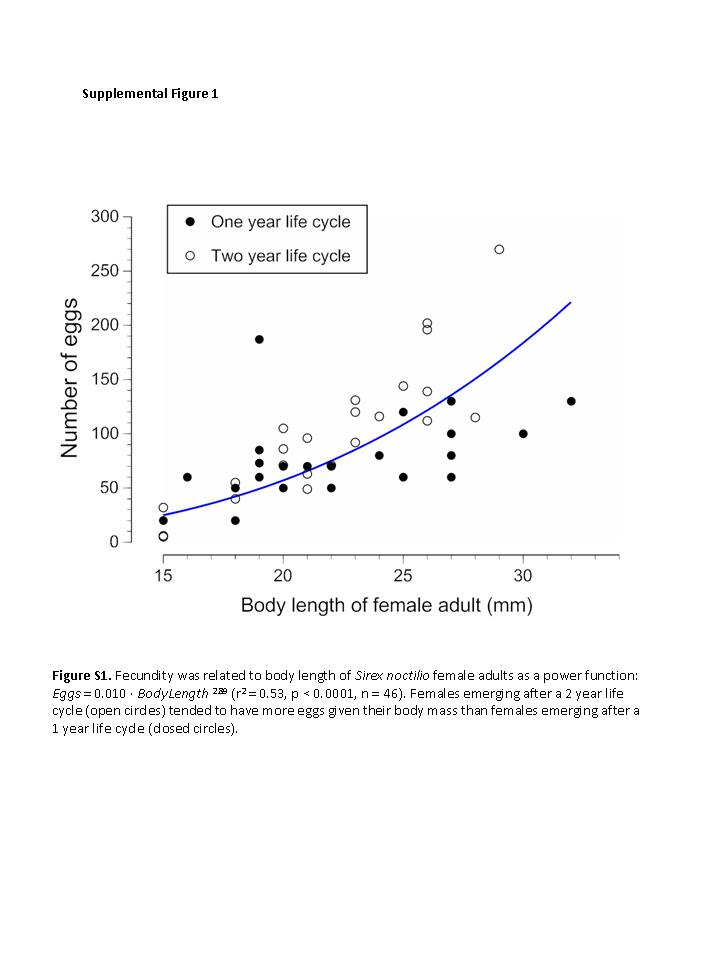
The number of eggs per female was highly variable, ranging from 0 to 270 (mean ± SD = 88 ± 54), and was related as a power function to all three linear dimensions of body size: Eggs = 0.010 · BodyLength 2.89 (r 2 = 0.53); Eggs = 7.49 · Pronotum 2.01 (r 2 = 0.33); Eggs = 1.83 · Tibia 2.37 (r 2 = 0.39) (P < 0.0001 and n = 46 females for all). Compared with females emerging 1 year after oviposition, females emerging 2 years after oviposition tended to have more eggs: 83 vs. 70 eggs given the same (average) body length of 22.1 mm (t = 2.28, df = 43, P = 0.027 for comparison of residuals; see fig. S1 in Supplementary Materials).
Overall sex ratio (females: males) of S. noctilio emerging from trees attacked in 2012 was 1: 2.9. Among the four plots, sex ratios ranged from 1: 2.3 (Begonte) to 1: 4.7 (Faro), but plots were not statistically distinguishable (χ2 = 1.66, df = 3, P = 0.65). The sex ratio did not differ between animals emerging 1 vs. 2 years after attack (1: 2.8 vs. 1: 3.1, respectively). The sex ratio of S. noctilio emerging from trees attacked in 2013 was similar to that of trees attacked in 2012 (1: 2.4).
Parasitism
We recorded two species of Hymenopteran parasitoids in Galicia: I. l. leucospoides (38 and 25 individuals from trees attacked in 2012 and 2013, respectively) and R. persuasoria persuasoria (26 from trees attacked in 2012). The first emergence of I. l. leucospoides (1 year after attack by S. noctilio) was slightly earlier than that of S. noctilio: median (10th–90th percentile) = 13 September 2013 (4 September–22 October) and 8 September 2014 (1 September–18 September). We also recorded emergence of some I. l. leucospoides 2 years after attack by S. noctilio (six individuals from trees attacked in 2012 emerging from 17 July to 7 September in 2014). Apparent parasitism of S. noctilio by I. l. leucospoides was 10 ± 3 and 6 ± 4% during emergence of 1 and 2 years, respectively, after attacks in autumn of 2012, and 30% during emergence of 1 year after attacks in autumn of 2013. Adults of R. p. persuasoria emerged about 1.5 years after attack by S. noctilio (during late April and early May of 2014 from trees attacked in fall of 2012); n = 26 individuals relative to 186 S. noctilio and 38 I. l. leucospoides emerging from the same material in the previous autumn.
We recorded frequent parasitism of S. noctilio by the nematode Deladenus siricidicola: 39% of 67 females and 36% of 181 males were parasitized, overall. The nematodes were sterilizing to females in that ≈90% of eggs within infected females contained 27–302 nematodes per egg. Animals parasitized by D. siricidicola were present at all four sites and in trees attacked in both 2012 and 2013. For animals emerging 1 year after attack, the frequency of parasitized adults was similar between years: 62 of 144 (43%) and 16 of 40 (40%) for trees attacked in 2012 and 2013, respectively. However, the frequency of parasitism by D. siricidicola was only 12 of 64 (19%) in adults emerging 2 years after attack. Parasitism was lower at Faro than at the other three sites: one of 14 adults at Faro vs. five of ten at Begonte, 36 of 89 at Cova da Serpe and 25 of 43 at Pena de Rodas (χ2 = 11.80, df = 3, P = 0.008). There was no difference in body size between parasitized and non-parasitized S. noctilio: F 1,177 < 1, P > 0.5 for effects of nematodes and nematode × sex; restricted to S. noctilio emerging 1 year after attack).
Reproductive success by species and tree
Slightly over half of the trees attacked by S. noctilio in the fall of 2012 had died by the time we cut them in July of 2013: 8 of 13 P. radiata from Begonte; 8 of 19 of P. sylvestris from Faro, 14 of 26 P. pinaster from Cova da Serpe and 18 of 23 P. pinaster in Pena de Rodas. Of the 48 trees that died following attack by S. noctilio in the fall, 41 were colonized in the winter after attack by the bark beetle T. piniperda. The other seven trees did not show any other causes of death other than attack by S. noctilio. Among trees attacked in fall of 2013, 12 of 14 P. radiata (Begonte) and 12 of 13 P. sylvestris (Faro), had died by the time we cut them the following July, but mortality of P. pinaster (Cova da Serpe) was only ten of 26.
The production of adult progeny of S. noctilio was highly concentrated in a minority of the attacked trees (fig. 1). About 70% of the attacked trees produced 0 adult progeny. More than 90% of the progeny emerged from less than 20% of the attacked trees. The probability density function for production by tree was well described by a zero-inflated negative binomial (ZINB) with ϕ = 0.41 (proportion of excess zeros), μ = 4.3 (central tendency for production), and k = 0.47 (overdispersion parameter for production). Most emergences came from P. pinaster: 15 of the 23 trees from Pena de Rodas had emergences and 17 of 52 trees from Cova da Serpe. Only five of 27 trees from Begonte (P. radiata) produced S. noctilio, and only one of 32 trees from Faro (P. sylvestris). Only five of the 48 trees that were still alive in the summer after attack produced adults of S. noctilio or their parasitoids, and this only accounted for 9% of total emergence.
Dissection of logs
In dissecting 171 logs, we found 3,256 attacks with 6,115 drillings. The number of attacks per 18 dm2 (the surface area of an average log) differed among study sites (F 3,40 = 4.47; P = 0.008; fig. 2) being highest in the two sites with P. pinaster (Pena de Rodas and Cova da Serpe) and lowest at Faro (P. sylvestris) and Begonte (P. radiata). The density of attacks was much higher (average of about 2×) in trees that were dead the summer after attack than in trees that were still alive (F 3,40 = 16.06; P = 0.0003). The average number of adult S. noctilio emerging per attack also varied among sites: 16 adults per 100 attacks at Cova da Serpe and 5–8 adults per 100 attacks at the other three sites (fig. 3). Logs that produced the most S. noctilio also tended to be most colonized by T. piniperda during January and February after the autumn when they were attacked: slope of T. piniperda attacks per log vs. S. noctilio attacks per log ± SE (both square root transformed) = 0.23 ± 0.07 (P = 0.001, n = 168 logs). We did not observe any tendency for attacks or emergences of S. noctilio to vary from the lower to upper bole of attacked trees.

Fig. 2. Density of attacks by S. noctilio in pine trees of three species from four sites in Galicia, Spain. Trees at each site were separated into those that did and did not die by mid-summer following attacks the previous fall (dead vs. alive). Attack densities were standardized to the surface area of an average log. The figure represents means and standard error bars which were back-transformed (from √x).

Fig. 3. Emergences relative to attacks for dissected logs from pines at four study sites in Galicia, Spain. Values indicate estimates of the population averages for emergences/attack ± SE at each study site (from slopes of regressions forced through the origin). Open symbols indicate trees that were still alive the summer after attack; others had died subsequent to attack.
The number of drillings per attack was 1, 2, 3 or occasionally more (to a maximum of 8): overall proportions for drills = 1: 2: 3: more = 0.43: 0.34: 0.18: 0.05. The average number of drillings per attack was highest in the two sites with P. pinaster, lowest at the site with P. sylvestris, and intermediate at the site with P. radiata (fig. 4). The average number of drills/attack was higher in trees that died by the following July compared with those that were still alive: least square means ± SE = 1.84 ± 0.07 vs. 1.53 ± 0.11 drills/attack (F 1, 52.9 = 5.26, P = 0.026; no interaction between site and tree status). These patterns notwithstanding, most of the variance in drills/attack was among nearby attacks within the same logs: ≈87% of the total random variance was within individual logs as opposed to among logs within trees or among trees within sites (from a nested analysis of log-transformed data). Similarly, the generalized linear mixed model indicated only modest variance among trees within sites and no variance among logs within trees.

Fig. 4. Frequency distributions of drills per attack by S. noctilio females attacking pines at four sites in Galicia, Spain. Values indicate mean ± SE of drills/attack.
About 62% of the total drillings were filled with resin and 35% induced lesions. The proportion of drills filled with resin was unrelated to whether trees died or not but varied among sites (F 3, 33.2 = 3.12, P = 0.039: lsmeans ± SE = 1.08 ± 0.08, 0.77 ± 0.08, 1.05 ± 0.13 and 0.81 ± 0.10 for Cova da Serpe, Pena de Rodas, Begonte and Faro, respectively; units = arcsin-transformed proportions). In contrast, the proportion of drills that produced lesions was lower in trees that died (F 1, 39.6 = 5.50, P = 0.024: lsmeans ± SE = 0.59 ± 0.06 vs. 0.86 ± 0.10, respectively) and did not vary among sites (F 3, 34.7 = 1.07, P = 0.27).
Whiteness of the xylem from disappearance of lignin (‘white rot’; apparently from A. areolatum) was only evident in a small part (average of 3% of cross-sectional area) of a minority of logs (55 of 171), and not until ≈6 months after S. noctilio emergence. There were no differences in the extent of white rot among sites (F 3, 75.4 = 0.90, P = 0.44), but white rot was significantly higher in trees that had died by the summer after attack (F 1, 76.2 = 6.03, P = 0.016: lsmeans ± SE = 0.091 ± 0.017 vs. 0.016 ± 0.025, respectively; units = arcsin-transformed proportions). The area of white rot was weakly but significantly related to attacks/dm2 (Pearson's r = 0.18, P = 0.014, n = 168 logs). Production of S. noctilio per log was positively correlated with the extent of white rot, as expected, but the relationship was weak and there were many logs with no visible white rot that had relatively high production of S. noctilio (fig. 5).

Fig. 5. Production per log of S. noctilio relative to visible extent of their mutualistic fungus, A. areolatum. Production tended to be higher in logs with more white rot, but there were also many logs with no visible white rot that still had relatively high production of S. noctilio. Values of 0.1, 0.4 and 0.7 on the x-axis correspond to 1, 15 and 41% of the cross-sectional area.
Blue stain from Ophiostoma spp. was ubiquitous within trees attacked by S. noctilio (visually evident in all but 2 of 171 logs). There was little or no variation in blue stain among sites (F 3, 36.6 = 1.37, P = 0.26). As with white rot, there was a relationship with whether trees were still alive in the summer after attack, but the pattern was opposite that of white rot: lsmeans ± SE = 0.85 ± 0.09 vs. 0.55 ± 0.07, for trees that were alive vs. dead, respectively (units = arcsin-transformed proportions; F 1, 36.6 = 7.13, P = 0.011). There was further conspicuous variation in blue stain among trees within site × status (42% of total random variance; estimated 95% confidence interval for variance among trees = 0.02–0.15). High blue stain within logs was correlated with lower extent of white rot (r = −0.16, P = 0.027) and lower productivity of S. noctilio (r = −0.36, P < 0.0001) (Pearson's correlation coefficients from n = 169 logs for both).
Determinants of S. noctilio emergence in P. pinaster
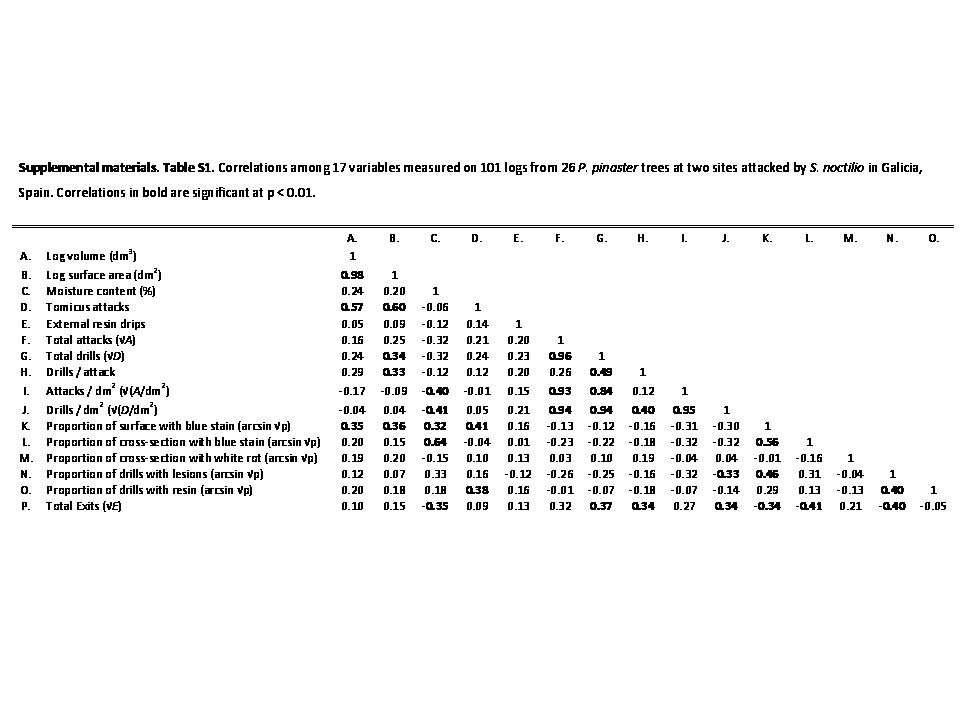
Our dissections included a total of 101 logs from P. pinaster, representing 26 trees attacked by S. noctilio. There was considerable structure to the correlation matrix of the various measurements (table S1, Supplemental materials). From these data, we identified a four-parameter model that explained 33% of the variance in emergences per log and in which the coefficients for all four parameters were significantly different from 0 (fig. 6). Emergence was positively related to total attacks per log (fig. 6a), but negatively related to the density of attacks (fig. 6d). Independent of total attacks and attack density, exits per log were negatively associated with the extent of blue stain and with the proportion of drills that induced lesions in the surrounding xylem tissue (fig. 6b, c). An alternative model in which total attacks and attacks/dm2 were replaced by total drills and drills/dm2 provided equivalent goodness of fit (delta AIC < 2; all four parameters also significant). There were no alternative models with equivalent or additional information from including any combination of the following other variables: extent of white rot, proportion of drills that were resin filled, attacks by T. piniperda, external resin drips from attacks by S. noctilio and moisture content of logs.

Fig. 6. Production of S. noctilio from 101 logs of P. pinaster relative to total attacks (a), proportion of blue stain (b), proportion of drills with lesions (c) and the density of attacks (d). Values within each frame indicate the coefficient ± SE for that parameter in a model that includes all four parameters (all significantly different from 0 at P < 0.05; intercept for the multiple linear regression = 2.32 ± 0.48; r 2 = 0.33). Exits per log in each panel are residuals after adjustment for effects of the other three parameters. The full model explained 33% of the variation in production of S. noctilio per log.
A preliminary life table of S. noctilio in Galicia
Data permitted a simple model of the demography of S. noctilio in Galicia (table 1). We estimated that an average female (with 88 eggs; data reported above) can generate up to 84 individual attacks on host trees (with drills per attack of 1–4 as in fig. 4, and eggs per drill as previously reported (Madden, Reference Madden1974; Spradbery, Reference Spradbery1977). Given the observed production of new adults per attack (fig. 3), this yields an average of about 7.9 potential adult progeny per parental female. The reduction from 88 eggs to 7.9 potential progeny was mostly attributable to larval mortality within the host tree and partly attributable to sterilization of 36% of eggs by the parasitic nematode, D. siricidicola. The reproductive potential of our study population was further depreciated by 20% incidence of Hymenoptera parasitoids. The male biased sex ratio (1: 2.8) further reduced λ by a factor of 0.48 relative to what it would be with a sex ratio of 1: 1 (table 1).
Table 1. A preliminary life table of S. noctilio in Galicia. An average female can generate up to about 84 attacks, with an average of about one egg per attack. With the consideration of impacts from parasitic nematodes, host suitability, parasitoids and skewed sex ratio, this suggests a maximum reproductive potential of λ = 1.65 (vs. 1.00 for replacement). The largest demographic impacts were from host suitability, followed by skewed sex ratio, parasitic nematodes and Hymenoptera parasitoids.

1 Proportional reduction in potential number of female progeny per female adult due to the corresponding demographic factor.
2 Eggs/attack = Σ(d·Pd·Ed), where d, number of drills per attack (1–4); Pd, proportion of attacks with drills = 1–4; and Ed, expected eggs/drill with drills = 1–4. Eggs/attack = 1.05 = midpoint of 0.64 and 1.45 as calculated using Pd = 0.43, 0.34, 0.18 and 0.05 from fig. 4 (this study) and Ed as reported by either Madden (1974; 0.04, 0.34, 0.52 and 0.55) or Spradbery (Reference Spradbery1977; 0.25, 0.63, 1.33 and 1.00).
3 Adult progeny/attack is lowered by: (1) parasitic nematodes that have sterilized the eggs introduced by attacking females; and (2) by mortality of immatures woodwasps within the host tree. With 84 attacks/female × 0.094 adult progeny/attack, there were 7.9 adult progeny/female; in the absence of 36% egg sterilization by nematodes, we can estimate that this would have been 10.7 adult progeny per attack. Thus, the estimated demographic impact from nematodes = 1–7.88/84 = 0.26, and the estimated impact from dying within the host tree = 1–10.7/84 = 0.87.
4 Relative to sex ratio of 1:1.
Discussion
Adult morphology and life history of S. noctilio in Galicia
Adult females of S. noctilio in our study area had dark femora, in contrast to the original species description and reports from most other areas (Schiff et al., Reference Schiff, Valley, LaBonte and Smith2006). This likely contributed to some earlier taxonomic confusion (Ceballos, Reference Ceballos1963; Llorente & Gabuyo, Reference Llorente and Gabuyo1984). Our specimens match descriptions of Schiff et al. (Reference Schiff, Goulet, Smith, Boudreault, Wilson and Scheffler2012) for S. noctilio in the Azores, Portugal, Spain, Italy and Turkey that are distinguishable from the more widespread pale legged form. Consistent with this, analyses of COI sequence data placed specimens from our study clearly within S. noctilio, but also within their own cluster separate from that of the pale legged S. noctilio in South Africa.
Generational development time of most S. noctilio in our study area was one year, but sometimes 2 years (24%) or even longer (≈4%). This is longer than reported by Spradbery & Kirk (Reference Spradbery and Kirk1978) for Europe (98% univoltine), shorter than reported for Tasmania (50% semivoltine; Neumann & Minko, Reference Neumann and Minko1981) and similar to New Zealand (Morgan, Reference Morgan1968) and Argentina (Corley & Villacide, Reference Corley, Villacide, Slippers, de Groot and Wingfield2012). Emergence of S. noctilio in Galicia started in mid-August and lasted about 3 months similar to Australia (Nahrung et al., Reference Nahrung, Ramsden and Griffiths2015). In other populations emergence is concentrated in a few weeks (Ryan & Hurley, Reference Ryan, Hurley, Slippers, de Groot and Wingfield2012). Voltinism and phenology can be expected to vary with climate (Neumann et al., Reference Neumann, Morey and McKimm1987; Corley et al., Reference Corley, Villacide and Bruzzone2007). The Galician climate is oceanic with a continental influence in the study plots (Martínez-Cortizas & Pérez-Alberti, Reference Martínez-Cortizas and Pérez-Alberti1999) with a mean annual temperature of ≈11°C and mean annual precipitation of ≈1,000 mm. Emergence times could be relatively protracted in Galicia because of cool rainy conditions in autumn when S. noctilio are emerging (Neumann et al., Reference Neumann, Morey and McKimm1987). Unlike Wolf (Reference Wolf1969a ) and Morgan (Reference Morgan1968) we did not find longer development times in lower trunks or larger trees.
There has been speculation (e.g., Hanson, Reference Hanson1939) about whether multi-year life cycles in Sirex spp. involve developmental quiescence (≈ diapause) vs. slow growth vs. extended growth. The occurrence in our material of early instars two years after oviposition provides strong evidence of prolonged developmental quiescence at the egg stage or early larval stage; this pattern is similar to reports of prolonged diapause in Argentina (Corley et al., Reference Corley, Villacide and Bruzzone2007). However, we also found evidence of extended growth in that male adults emerging in the 2nd year were about 50% longer than those emerging after one year, and females emerging in the 2nd year, though not longer, had ≈18% more eggs for their body size than females that were univoltine.
Our adults were smaller than reported from some areas: female lengths of 8–38 mm in Galicia contrast with 21–72 mm in Tasmania (Madden, Reference Madden1974) and 10–44 mm in South Africa (Hurley et al., Reference Hurley, Slippers, Croft, Hatting, van der Linde, Morris, Dyer and Wingfield2008), but similar to Victoria, Australia (Neumann & Minko, Reference Neumann and Minko1981). The diameters of exit holes from our material were also small compared with other reports for this species: 1–6 mm in Galicia vs. 3–7 mm in Zondag & Nuttall (Reference Zondag and Nuttall1977) and Schiff et al. (Reference Schiff, Goulet, Smith, Boudreault, Wilson and Scheffler2012). Concomitant with the smaller body size, fecundity of our population was lower than in the Southern Hemisphere: range for eggs/female = 0–270 vs. 30–450 (Madden, Reference Madden1974), 50–500 (Zondag & Nuttall, Reference Zondag and Nuttall1977), 21–458 (Neumann & Minko, Reference Neumann and Minko1981) and 40–466 (Corley et al., Reference Corley, Villacide and Bruzzone2007). We found an average of 88 eggs/female vs. 220 reported by Neumann et al. (Reference Neumann, Morey and McKimm1987). Compared with Tasmanian material (Madden, Reference Madden1974), Galician females had as many eggs as expected based on pronotum width, but only about half as many eggs on average because of their smaller size. In terms of table 1, the potential factor of population increase (λ) would be ≈3.3 instead of ≈1.6 if Galician females were as large as in Tasmania.
The overall sex ratio in Galicia was 1:2.9 (female: male), which is somewhat more male biased than reported by Spradbery & Kirk (Reference Spradbery and Kirk1978) for Europe (1:1.8); but not nearly as biased as in some invasive populations (1:32 in Brazil: Iede et al., Reference Iede, Penteado and Schaitza1998; 1:12 in South Africa: Hurley et al., Reference Hurley, Slippers, Croft, Hatting, van der Linde, Morris, Dyer and Wingfield2008). Even the relatively modest sex ratio bias in Galicia was enough to be the second strongest demographic factor of those we could parse (48% reduction in population growth potential compared to a 1: 1 sex ratio; table 1).
Reproduction success by tree in Galicia
Maybe the most remarkable result from our study was the low abundance of S. noctilio in Galicia. The species appeared to be restricted to suppressed trees (Hall, Reference Hall1968; Ayres et al., Reference Ayres, Pena, Lombardo and Lombardero2014), which are scarce in the actively managed forests that prevail in our study area. Most pine stands were devoid of resin drips from attacks or holes from emerging adults. Our four study plots contained the most conspicuous populations of S. noctilio that we could find, and still the abundances were very low: only 313 adult progeny from 134 attacked trees (compare for example to 2,558 progeny from 18 attacked trees in New York (Eager et al., Reference Eager, Allen, Frair and Fierke2011) or 7,018 individual from 60 attacked trees in Canada (Ryan et al., Reference Ryan, de Groot, Nott, Drabble, Ochoa, Davis, Smith and Turgeon2012)).
Even among the suppressed trees that were attacked by S. noctilio, it was only a fraction that produced progeny. We could not find comparable frequency distributions of emergence per tree from other locations. We hypothesize that outbreak populations have a lower fraction of attacked trees with 0 emergences than in Galicia (ϕ = 0.41 from ZINB). In terms of the ZINB fit to our data, a modest reduction in ϕ or increase in μ would increase emergences per attack, which could influence abundance of S. noctilio because of the sensitivity of λ to progeny per attack (table 1). Understanding host suitability seems important for understanding variable abundance of S. noctilio.
Regression analyses of dissected logs suggested some apparent drivers of host suitability for S. noctilio. The number of attacks (and drillings) per tree was positively related to emergence per tree. This is at least partly because more attacks means more eggs (estimated average of about 1 egg per attack). So emergences would be positively related to attacks given similar survival of egg to adult. However, more attacks and more drills also means more inoculation of the phytotoxin (noctilisin) and the fungus (A. areolatum), which should increase the probability of tree death, and thereby increase the success of the fungus and larvae within trees (Coutts & Dolezal, Reference Coutts and Dolezal1969; Madden, Reference Madden1977; Corley et al., Reference Corley, Villacide and Bruzzone2007). Indeed, trees that experienced higher attack densities tended to be those that died following attack and those that accounted for most production of S. noctilio. If there is positive feedback in the population dynamics from increasing attack success with increasing abundance (Corley et al., Reference Corley, Villacide and Bruzzone2007), it could explain the tendency for S. noctilio to switch quickly between low and high abundance. This is consistent with there being particularly low production of S. noctilio in our study sites where S. noctilio was least abundant (Faro and Begonte). However, our data do not demonstrate that higher attack densities cause higher tree mortality; the pattern could also indicate that S. noctilio attack at higher densities on trees that are already more likely to die. If outbreaks of S. noctilio are partly a result of positive feedback from larger populations having greater success in killing trees, then we could expect higher attack densities in outbreak populations than we observed in Galicia. Such data seem to be limited, but Coutts & Dolezal (Reference Coutts and Dolezal1966) reported up to ‘500 tunnels per square foot of bark surface’, which is ≈30-fold higher than our observations of no more than 1–2 attacks/dm2.
The suitability of P. pinaster for S. noctilio was inversely related to the tendency for induced production of polyphenols (lesions) around points of drilling by attacking females. Overall, more than a third of drillings (2,141 of 6,041) induced visible lesions. Our results support the hypothesis that these lesions are a defensive response against A. areolatum (Coutts & Dolezal, Reference Coutts and Dolezal1966; Hillis & Inoue, Reference Hillis and Inoue1968; Talbot, Reference Talbot1977; Madden, Reference Madden and Berryman1988; Thompson et al., Reference Thompson, Bodart, McEwen and Gruner2014). Notably, lesions were more frequent in trees that were still alive the summer after attacks.
The success of S. noctilio within logs was inversely related to the presence of blue stain fungi. This is consistent with the hypothesis of competition between blue stain fungi and A. areolatum (Wolf, Reference Wolf1969a ; Ryan et al., Reference Ryan, Moncalvo, de Groot and Smith2011; Hurley et al., Reference Hurley, Hatting, Wingfield, Klepzig and Slippers2012; Yousuf et al., Reference Yousuf, Carnegie, Bashford, Bedding, Nicol and Gurr2014a , Reference Yousuf, Gurr, Carnegie, Bedding, Bashford, Gitau and Nicol b ), but we question whether this interpretation applies in our case. The main vector of blue stain to our pines is the bark beetle, T. piniperda, which flies several months after S. noctilio. Thus the introduction of blue stain should generally come well after A. areolatum has had a chance to establish itself in host trees.
After accounting for effects of other factors, an additional negative correlation emerged between production per log and attack density, which suggests larval competition. Competition for space within the xylem was not evident in our dissections, but there could be competition for limited patches of A. areolatum, especially in trees with low carbohydrate content (Titze, Reference Titze1965). Another possibility is that concentrations of attacks within a region of pine bole could trigger more effective inducible defences in those regions.
Top-down effects
In addition to effects from host suitability and skewed sex ratios, there are strong effects on the abundance of S. noctilio in Galicia from natural enemies (table 1). The strongest top-down effects were from the parasitic nematode, D. siricidicola. This is consistent with the attention that has been afforded D. siricidicola as a biological control agent (Hurley et al., Reference Hurley, Slippers and Wingfield2007). Ours is the first work that we know since Wolf (Reference Wolf1969b ) reporting the frequency of D. siricidicola in a native population of S. noctilio. We found D. siricidicola to be ubiquitous (infecting 36–39% of adults at all study sites). This is consistent with earlier studies in Belgium (Wolf, Reference Wolf1969b ). Parasitism by D. siricidicola in invaded areas, where it has been repeatedly introduced as a control agent, can be both higher and lower than we found in Galicia. In Australia infection can reach 100% of the population within inoculated trees (Bedding & Akhurst, Reference Bedding and Akhurst1974, but see Nahrung et al., Reference Nahrung, Ramsden and Griffiths2015); infection levels in Brazil are 70 to 80% (Iede et al., Reference Iede, Penteado and Schaitza1998), and in South Africa infection ranges from 23 to 96% (Tribe & Cillié, Reference Tribe and Cillié2004). However there have been less successful introductions in Brazil (Fenili et al., Reference Fenili, Mendes, Miquelluti, Mariano-da-Silva, Xavier, Ribas and Furlan2000), Argentina (Eskiviski et al., Reference Eskiviski, de Coll, Faraldo and Bennesch2003) and South Africa (Hurley et al., Reference Hurley, Slippers and Wingfield2007). The abundance of D. siricidicola could be affected by environmental conditions – including any factors that influence growth of A. areolatum, on which the nematodes feed within trees before infecting larvae (Hurley et al., Reference Hurley, Slippers, Croft, Hatting, van der Linde, Morris, Dyer and Wingfield2008; Carnegie & Bashford, Reference Carnegie, Basford, Slippers, de Groot and Wingfield2012; Nahrung et al., Reference Nahrung, Ramsden and Griffiths2015).
D. siricidicola in Galicia readily entered female eggs and caused nearly complete sterilization of infected females in our study; this differs from North American populations (Kroll et al., Reference Kroll, Hajek, Morris and Long2013). Understanding the maintenance of sterilization in native, unmanaged, populations of D. siricidicola, such as in Galicia, could have value in managing the chronic loss of sterilization in populations of nematodes introduced for biological control (Bedding & Iede, Reference Bedding, Iede, Grewal, Ehlers and Shapiro-Ilan2005).
D. siricidicola offers a potential explanation for the prolonged life cycle displayed by about 35% of Galician S. noctilio. Females that emerged two years after attack tended to have about 35% higher fecundity, and also had only about half the risk of being sterilized by D. siricidicola. The reduced parasitism could be a result of higher nematode performance in moist wood (Bedding & Akhurst, Reference Bedding and Akhurst1974; Hurley et al., Reference Hurley, Slippers, Croft, Hatting, van der Linde, Morris, Dyer and Wingfield2008), and wood moisture decreases with time following attack by S. noctilio and colonization by A. areolatum (Coutts & Dolezal, Reference Coutts and Dolezal1965).
S. noctilio in Galicia are also regularly parasitized by two species of Hymenoptera. These same species of parasitoids have been reported at comparable frequencies elsewhere in Europe (Wolf, Reference Wolf1969b ; Spradbery & Kirk, Reference Spradbery and Kirk1978). The overall proportion of S. noctilio killed by parasitoids in Galicia (≈20%), was less than reported in several studies from the Southern Hemisphere (25–55% in New Zealand, Victoria, New South Wales and South America; Neumann et al., Reference Neumann, Morey and McKimm1987; Iede et al., Reference Iede, Penteado, Filho and Schaitza2000; Carnegie et al., Reference Carnegie, Eldridge and Waterson2005) and on the low end for reports from North America of ≈10–40% (Ryan et al., Reference Ryan, de Groot, Nott, Drabble, Ochoa, Davis, Smith and Turgeon2012; Zylstra & Mastro, Reference Zylstra and Mastro2012; Kroll et al., Reference Kroll, Hajek, Morris and Long2013). It is tempting based on table 1 and comparisons with other continents to infer that Hymenopteran parasitoids are less important than low host suitability and parasitic nematodes as explanations for low abundance of S. noctilio in Galicia in particular and Europe in general. However, we still have limited understanding how the factors identified in table 1 vary with the abundance of S. noctilio. Understanding the direction and strength of density-dependent feedbacks is an emerging challenge for understanding the global ecology and management of S. noctilio.
Supplementary material
The supplementary material for this article can be found at http://dx.doi.org/10.1017/S0007485316000043
Acknowledgements
Support was provided by USDA Forest Service International Programs. Andrew Liebhold contributed to research design, field work and analyses. Thanks to Jimbeaux Black, Daniel Canosa, Wales Carter, Lars Höger, Naidu Lombardero and Alicia Villamarín for assistance with field and laboratory work. Manuel Guaita provided tools and assistance for processing logs. The Consellería de Medio Rural e do Mar (Xunta de Galicia) gave permission to conduct studies in their forest.