









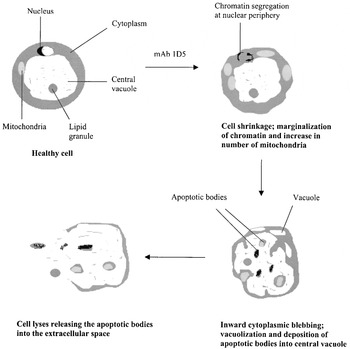


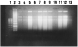





Crossref Citations
This article has been cited by the following publications. This list is generated based on data provided by Crossref.
Tan, Kevin S.W
Singh, Mulkit
and
Yap, Eu Hian
2002.
Recent advances in Blastocystis hominis research: hot spots in terra incognita.
International Journal for Parasitology,
Vol. 32,
Issue. 7,
p.
789.
Wang, Ke-Xia
2002.
Epidemiological survey ofBlastocystis hominisin Huainan City, Anhui Province, China.
World Journal of Gastroenterology,
Vol. 8,
Issue. 5,
p.
928.
Flores-Pérez, Ivan
Gonzalez, Gladis Fragoso
Sciutto, Edda
and
de Aluja, Aline S.
2003.
Apoptosis induced by gamma irradiation of Taenia solium metacestodes.
Parasitology Research,
Vol. 90,
Issue. 3,
p.
203.
Kobayashi, T
and
Endoh, H
2003.
Caspase-like activity in programmed nuclear death during conjugation of Tetrahymena thermophila.
Cell Death & Differentiation,
Vol. 10,
Issue. 6,
p.
634.
Tan, Kevin S.W.
2004.
Blastocystis in humans and animals: new insights using modern methodologies.
Veterinary Parasitology,
Vol. 126,
Issue. 1-2,
p.
121.
Moreira, Maria Elisabete C.
and
Barcinski, Marcello A.
2004.
Apoptotic cell and phagocyte interplay: recognition and consequences in different cell systems.
Anais da Academia Brasileira de Ciências,
Vol. 76,
Issue. 1,
p.
93.
Nasirudeen, A.M.A.
and
Tan, Kevin S.W.
2004.
Caspase-3-like protease influences but is not essential for DNA fragmentation in Blastocystis undergoing apoptosis.
European Journal of Cell Biology,
Vol. 83,
Issue. 9,
p.
477.
Argüello-García, Raúl
Cruz-Soto, Maricela
Romero-Montoya, Lydia
and
Ortega-Pierres, Guadalupe
2004.
Variability and variation in drug susceptibility among Giardia duodenalis isolates and clones exposed to 5-nitroimidazoles and benzimidazoles in vitro.
Journal of Antimicrobial Chemotherapy,
Vol. 54,
Issue. 4,
p.
711.
Nasirudeen, A. M. A.
Hian, Yap Eu
Singh, Mulkit
and
Tan, Kevin S. W.
2004.
Metronidazole induces programmed cell death in the protozoan parasite Blastocystis hominis.
Microbiology,
Vol. 150,
Issue. 1,
p.
33.
Nasirudeen, A.M.A.
and
Tan, Kevin S.W.
2005.
Programmed cell death in Blastocystis hominis occurs independently of caspase and mitochondrial pathways.
Biochimie,
Vol. 87,
Issue. 6,
p.
489.
Figarella, K
Rawer, M
Uzcategui, N L
Kubata, B K
Lauber, K
Madeo, F
Wesselborg, S
and
Duszenko, M
2005.
Prostaglandin D2 induces programmed cell death in Trypanosoma brucei bloodstream form.
Cell Death & Differentiation,
Vol. 12,
Issue. 4,
p.
335.
van der Giezen, Mark
Tovar, Jorge
and
Clark, C. Graham
2005.
A Survey of Cell Biology.
Vol. 244,
Issue. ,
p.
175.
Tan, Kevin S.W.
and
Nasirudeen, A.M.A.
2005.
Protozoan programmed cell death – insights from Blastocystis deathstyles.
Trends in Parasitology,
Vol. 21,
Issue. 12,
p.
547.
Durand, Pierre Marcel
and
Coetzer, Theresa Louise
2008.
Utility of Computational Methods to Identify the Apoptosis Machinery in Unicellular Eukaryotes.
Bioinformatics and Biology Insights,
Vol. 2,
Issue. ,
p.
BBI.S430.
González, Ana Martín
Díaz, Silvia
Gallego, Andrea
and
Gutiérrez, Juan C.
2008.
Programmed Cell Death in Protozoa.
p.
143.
Yarlett, Nigel
and
Tan, Kevin S. W.
2008.
Hydrogenosomes and Mitosomes: Mitochondria of Anaerobic Eukaryotes.
Vol. 9,
Issue. ,
p.
255.
Tan, Kevin S. W.
2008.
New Insights on Classification, Identification, and Clinical Relevance of
Blastocystis
spp
.
Clinical Microbiology Reviews,
Vol. 21,
Issue. 4,
p.
639.
Deponte, Marcel
2008.
Programmed cell death in protists.
Biochimica et Biophysica Acta (BBA) - Molecular Cell Research,
Vol. 1783,
Issue. 7,
p.
1396.
Tan, Kevin S. W.
2008.
Programmed Cell Death in Protozoa.
p.
116.
Puthia, Manoj K.
Lu, Jia
and
Tan, Kevin S. W.
2008.
Blastocystis rattiContains Cysteine Proteases That Mediate Interleukin-8 Response from Human Intestinal Epithelial Cells in an NF-κB-Dependent Manner.
Eukaryotic Cell,
Vol. 7,
Issue. 3,
p.
435.