


Introduction
Transgenic, herbicide-resistant oilseed rape (Brassica napus L.) has been commercially approved for more than 10 years and grown over 4–5 million hectares in recent years in North America, but is still under discussion in Europe. In China, several transgenic lines resistant to herbicides and insects are being tested in the laboratory and in field trials. Concern about gene flow to wild relatives and the impact of the introduction of new traits in plant communities has grown, since spontaneous hybridization with close wild relatives of transgenic oilseed was demonstrated with B. rapa L., B. juncea (L.) Czern, Hirschfeldia incana (L.) Lagrèze-Fossat, Raphanus raphanistrum L., Sinapis arvensis L. and B. nigra (L.) Koch (Ellstrand, Reference Ellstrand2003). Introgression between B. napus and B. rapa has been reported in natural populations (Hansen et al., Reference Hansen, Siegismund and Jørgensen2001), and evidence of transgene persistence in a weedy B. rapa population was provided recently (Warwick et al., Reference Warwick, Légère, Simard and James2008). Gene flow could enhance the weediness of weeds in arable fields and affect the biology of wild populations in natural habitats, since the transgenes provide adaptive traits such as resistance to herbicides, pests, diseases and various environmental stresses (Darmency, Reference Darmency1994). However, interspecific hybridization could result in developmental and reproductive costs due to genomic imbalance and, therefore, the establishment of hybrids and transgenes in wild populations could be largely delayed or even unsuccessful due to detrimental characteristics. The fate of the first-generation hybrids is crucial because subsequent introgression, if successful, may restore normal plant functions, allowing the transgene express fully and open the way for further evolution boosted by beneficial transgenes.
Natural hybrids in fields and on riversides were reported only for B. napus × B. rapa hybrids, named B. × harmsiana O.E. Schulz. (Stace, Reference Stace1975). There is no other evidence suggesting that the hybrids formed between B. napus and other wild relatives could establish in nature. Therefore, hybrid seed germination and seedling emergence may constitute limiting factors to hybrid establishment. Much research has been carried out on the fitness of B. napus hybrids with B. rapa. In all cases, growth and seed production were analysed on established hybrids growing in natural populations (Allainguillaume et al., Reference Allainguillaume, Alexander, Bullock, Saunders, Allender, King, Ford and Wilkinson2006) or transplanted from, and even completely grown in, growth cabinets (Hauser et al., Reference Hauser, Damgaard and Jørgensen2003; Ammitzbøll et al., Reference Ammitzbøll, Mikkelsen and Jørgensen2005; Halfhill et al., Reference Halfhill, Sutherland, Moon, Poppy, Warwick, Weissinger, Rufty, Raymer and Stewart2005, and previous work cited therein). The ability of hybrid seeds to germinate and establish as seedlings has seldom been studied. Guéritaine et al. (Reference Guéritaine, Sester, Eber, Chèvre and Darmency2002, Reference Guéritaine, Bazot and Darmency2003) discussed the possible role of hybrid seed quality in the reduced fitness of B. napus × R. raphanistrum hybrids in direct seeding experiments. Indeed, crosses between B. napus and B. rapa (Jørgensen and Andersen, Reference Jørgensen and Andersen1994), H. incana (Eber et al., Reference Eber, Chèvre, Baranger, Vallée, Tanguy and Renard1994; Chadoeuf et al., Reference Chadoeuf, Darmency, Maillet and Renard1998), R. raphanistrum (Eber et al., Reference Eber, Chèvre, Baranger, Vallée, Tanguy and Renard1994; Chadoeuf et al., Reference Chadoeuf, Darmency, Maillet and Renard1998) and B. juncea (Bing et al., Reference Bing, Downey and Rakow1996) only produced small seeds. Variations in seed size are of major ecological relevance. For instance, Stanton (Reference Stanton1984) quoted an early work by Galloway on the seed-size effect of the growth of wild radish. Small-sized seeds were assumed to escape predation better than large seeds and are easier to disperse further, but emerge later if buried deep in the soil (Harper, Reference Harper1977); this may decrease plant fitness in the field (Verdu and Traveset, Reference Verdu and Traveset2005). There is a positive relationship between seed size, higher rates of survival and final plant size among different plant species (Westoby et al., Reference Westoby, Falster, Moles, Vesk and Wright2002; Moles and Westoby, Reference Moles and Westoby2004).
In this study, we aimed to assess whether the small seed size of spontaneous hybrids affects the early development and establishment of hybrids between five wild Brassicaceae species and B. napus. Our work is the first study of seedling establishment of interspecific Brassica hybrids that takes the effect of seed size into consideration. We also addressed the effect of the parent seed size on the hybrid seeds and their subsequent development.
Materials and methods
Plant materials
The seeds of wild species B. juncea (2n = 36, AABB, Turkey, ref. X04-021), B. rapa (2n = 20, AA, Germany, ref. X04-023), B. nigra (2n = 16, BB, Germany, ref. X04-022), R. raphanistrum (2n = 18, RrRr, France, ref. R04-050), S. arvensis (2n = 18, SarSar, France, ref. S04-001) and H. incana (2n = 14, AdAd, France, ref. A04-001) were obtained from the Dijon laboratory collection of weeds. A sulphonylurea herbicide-resistant spring oilseed rape [B. napus cv. Brutor (CRS) (2n = 38, AACC)] and a cytoplasmic male sterile spring oilseed rape [cv. Drakkar (CMS)] were provided by M. Renard (INRA, Rennes, France). Spontaneous hybrids were produced using the male sterile CMS line of B. napus. There were several reasons for this choice. By using the same male sterile B. napus line as the mother plant: (1) the hybrid yield expected was higher than in the reciprocal crosses; (2) hybrid identification was easier than when the wild species served as the female parent; (3) the seed-size categories were determined by those used to detect the hybrids and therefore were homogeneous for all the species; (4) the resulting material was not influenced by maternal effects that could have been different for each wild species if the female had been the wild species, especially for germination and emergence; (5) all the differences found among hybrids from each wild species were thus due to the paternal species contribution and the interaction between paternal and maternal genomes; (6) male sterility is much used to breed hybrid varieties, but is not stably restored in the progeny, so that volunteers and feral plants on the field border will be real traps for foreign pollen; (7) composite cultivars containing 80% of male sterile plants have become more and more popular in Europe and can occur as volunteers and in field borders, and they represent the highest risk of producing interspecific hybrids because they are easily fertilized by surrounding wild relatives [Our previous work showed that a few male sterile plants 5 m away from the field produced many more hybrids than the whole field (Chèvre et al., Reference Chèvre, Eber, Darmency, Fleury, Picault, Letanneur and Renard2000)]; (8) hybridization with B. napus as the female parent is the unique transgene escape route for future transplastomic varieties that are supposed to reduce gene flow (Johannessen et al., Reference Johannessen, Andersen and Jørgensen2006).
Seeds were harvested in six 2 m × 3 m insect-proof cages in Dijon in 2004. A cage contained 35 plants belonging to one of the six wild species, 13 CMS plants as targets for foreign pollen to produce interspecific hybrids, and 2 CRS plants to provide control B. napus CMS × CSR hybrid seeds and self-pollinated CRS seeds. The cages were continuously supplied with houseflies (Musca domestica L.) during the flowering period to facilitate random cross-pollination. The seeds of each plant type were sieved into three categories by seed diameter: larger than 1.6 mm (L), smaller than 1.2 mm (S), and medium-sized between 1.2 and 1.6 mm (M). These categories were the same as those used in previous studies on the hybridization of B. napus with H. incana and R. raphanistrum (Chadoeuf et al., Reference Chadoeuf, Darmency, Maillet and Renard1998).
Transplant experiment
Seeds of all three size categories – belonging to the six wild relatives, CRS and CMS types – were sown in trays containing vermiculite in the greenhouse on 25 April 2005 in Dijon (22°C, natural light). Emergence rates were recorded, and the interspecific hybrids were easily identified: they had intermediate leaf shape and hairiness between those of the two parents, which made them very different from the surrounding CMS × CRS B. napus × B. napus hybrids which were all alike. Since these plants were always confirmed as true hybrids in previous work when using other identification tools (cytometry, isozymes and DNA markers) (e.g. Darmency et al., Reference Darmency, Lefol and Fleury1998; Chèvre et al., Reference Chèvre, Eber, Darmency, Fleury, Picault, Letanneur and Renard2000; Darmency and Fleury, Reference Darmency and Fleury2000), no further test was carried out. After the interspecific hybrids were removed from the trays, all but three CMS trays from the cage with S. arvensis were sprayed with 2.5 g a.i. ha− 1 chlorsulfuron in 300 litres of water (Glean, Du Pont de Nemours, France). All 12,800 CMS seedlings survived the herbicide treatment, thus indicating that they were all CMS × CRS hybrid seeds; there were no matromorphic or apomictic seeds. Since all the interspecific hybrids are susceptible, as they have no resistant parent and the resistance is dominant (Issaka Magha et al., Reference Issaka Magha, Guerche, Bregeon and Renard1993; Darmency et al., Reference Darmency, Lefol and Fleury1998; Darmency and Fleury, Reference Darmency and Fleury2000), this also indicates that we have identified them all. Since no interspecific hybrids were observed among the 3400 CMS seeds from the cage with S. arvensis, these seeds were further used to represent the CMS × CRS type, and data on S. arvensis were not included in the study.
Immediately after the interspecific hybrids had been identified on 20 May 2005, all the seedlings were transplanted into Jiffy-7 peat pellets and grown for 2 weeks in the greenhouse. In May, CRS plants derived from large-sized seeds were planted in a sandy-clay soil field at the Institute in Dijon. There were six rows 0.3 m apart; and plants in the two external rows were planted every 0.3 m, and those in the four internal rows were alternately planted every 0.6 m, thus allowing zigzag places for transplanting the various seed-size categories of parents and hybrids between the two external rows. Up to 30 plants per plant type and category were transplanted on 3 June 2005 in the empty places. Each of these plants was at a distance of 0.3 m to CRS crop plants in each of the four directions, and there was no direct contact to other studied plants, ensuring a homogeneous competition environment. The precise location of each plant was randomly selected, and each plant was labelled. The whole design was divided into seven lanes, each separated by a 1-m path, with a total length of 140 m. Every plant was watered the day that it was transplanted and three times by sprinkler throughout the whole field from early July to mid-August. The experiment was kept weed-free by hand weeding. Flowering dates were recorded. The plant height, rosette diameter and total dried aboveground biomass were measured at harvest in September 2005, a date when the few seeds that escaped from Meligethes (pollen beetle) predation became mature. No reproductive traits were measured, since the experiment suffered a strong and continuous attack by pollen beetles, which destroyed nearly all the buds in spite of numerous insecticide sprays.
Experiment with direct seed sowing
As interspecific hybrids between CMS and B. rapa were expected to be abundant, a similar design was set up on 30 April 2005 in the same field as the above experiment, but with direct seeding in the soil. Large-sized CRS seeds were sown in six rows 0.3 m apart at a density of 120 seeds m− 2. Just after sowing, 30 precise positions were assigned at random to each seed-size category of B. rapa, CMS × CRS and CRS, and 60 positions were also assigned randomly to CMS from the cage with B. rapa containing the potential interspecific hybrids, herein called CMS × B. rapa. Labelled positions (450) were distributed 0.45 m apart within the three inter-rows. Different numbers of seeds were sown at each position according to the number of seeds available and/or expected germination rate: two for M seed-size of B. rapa, three for B. rapa L and CMS × B. rapa L–M, four for B. rapa S, CMS × CRS L–M–S, CRS L–M and CMS × B. rapa S, and five for CRS S. Therefore, from 60 to 240 seeds were sown per plant type and seed category. Emerged seedlings were recorded on 26 May 2005, and randomly thinned to one plant per specific position after identifying the interspecific hybrids. Flow cytometry was used as described by Akinerdem (Reference Akinerdem1991) to confirm the hybrids between CMS and B. rapa, because morphology alone was less reliable in the field than in the greenhouse. Daily mean temperatures roughly increased linearly from 10 to 25°C from sowing time to the end of June, with an average difference of c. 12°C between night-time and day. There was rain on 3 May (10.5 mm), 8 May (3 mm), 16 May (3.5 mm), 22 May (20.5 mm), 6 June (5 mm) and 15 June (7.5 mm), thus allowing germination and seedling emergence, and then watering was performed three times by sprinkler from early July to mid-August. The measurements were the same as for the transplant experiment.
Statistics
Trait analyses included all individual plant values and compared differences among seed sizes and plant types, using a two-way ANOVA procedure in SYSTAT software version 10 (SPSS Inc., Chicago, Illinois, USA) with pairwise comparisons of Fisher's least significant differences (LSD) test. In cases where one-way ANOVA was carried out, contrast tests were applied to differentiate among seed-size categories or plant types. In no instance was data transformation needed (Leven test). We calculated the correlations between the performance of hybrids in the transplant experiment and the percentage of small seeds or the average seed weight of their respective parents (Bartlett chi-square test). A linear regression of seedling emergence in the direct seeding experiment was calculated in terms of seed weight of each seed-size category.
Results
Seed characteristics
Small seeds < 1.2 mm dominated seed production for all the species and 100% for B. nigra and H. incana (Table 1). The weight of 1000 seeds decreased with the seed size. On average, B. juncea, R. raphanistrum and B. napus CRS had the heaviest seeds, while B. nigra and H. incana had the lightest. Germination in the greenhouse was lower for the small seed size, except for R. raphanistrum which had low germination for the three seed categories.
Table 1 Size category distribution, weight and percent germination (±95% confidence limits) of the seeds harvested from various plants grown in the cages for use in subsequent experiments. Seed-size categories were large (L, >1.6 mm), medium (M, between 1.6 and 1.2 mm), and small (S, <1.2 mm). No hybrid seeds were produced with S. arvensis as a parent

CRS, sulphonylurea herbicide-resistant spring oilseed rape, Brassica napus cv. Brutor. CMS, cytoplasmic male sterile spring oilseed rape cv. Drakkar.
aAverage of 6 times 1000 seeds (dehulled seeds for R. raphanistrum), not estimated for B. rapa because of seed shortage.
bMost of the seeds had a CRS male parent.
Interspecific hybrids were found in the seedlings of CMS plants from the cage, with B. rapa at a high percentage (66.2, 65.2 and 62.1% for L, M and S categories, respectively). In CMS plants from the cages with the four other species, interspecific hybrids were only observed in the small seed-size category: 11.9% for R. raphanistrum, 4.4% for B. nigra, 0.9% for H. incana and 0.8% for B. juncea. The actual hybridization rate per number of seeds produced in the cages would be lower when considering all seed-size categories together.
Transplant experiment
Very few plants died after transplanting. Most of the plants flowered, except for small-seeded B. rapa (48% flowering, Table 2). From flowering to maturity, pollen beetle attack resulted in so much damage that no seed production data could be scored. When data of the three different seed-size categories were available (i.e. for CRS, B. juncea, B. rapa and R. raphanistrum parents, and CMS × CRS and CMS × B. rapa hybrids), there was no significant seed category effect, and no plant type × seed-size interaction was found in the two-way ANOVA. For instance, the plant biomass of large-, medium- and small-seeded plants was 6.23 ± 0.77, 6.13 ± 0.74 and 5.98 ± 0.73 g, respectively. Plant type was always a highly significant factor (F 5,451 = 202, 15, 234 and 62 for height, diameter, flowering and biomass, respectively, P < 0.001).
Table 2 Plant characteristics from the transplant experiment for the small (S) seed-size category of the five hybrid types and their male parent species (±95% confidence limits)

DAS, days after sowing; CRS, sulphonylurea herbicide-resistant spring oilseed rape, Brassica napus cv. Brutor; CMS, cytoplasmic male sterile spring oilseed rape cv. Drakkar.
Different letters within a column indicate significantly different values at P < 0.05.
Comparing hybrids to their respective male parents was possible for small seeds only because interspecific hybrids were found in a majority (79.5% in B. rapa) or in totality (for the other species) in that seed category. The small seed category was also the most important fraction of the seeds produced in the cages for the wild species. Differences were found between hybrids and their corresponding parents in 10 cases out of 24 possible by-pair comparisons (Table 2, one-way ANOVA F 11,235 = 51, 10, 49 and 15 for height, diameter, flowering and biomass, respectively, P < 0.001). There was no significant correlation between the mean score of hybrid types and the mean seed weight or the small seed proportion of their respective male parents: the best correlation, between seed weight of male parents and biomass of hybrids, had a probability of P = 0.11.
Experiment with direct seed sowing
Frequencies of seedling emergence ranged from 0.23 ± 0.05 to 0.69 ± 0.07 (mean ± 95% CL), with apparent differences among both plant types and seed-size categories (Fig. 1). The three seed categories of CSR showed the highest values, but there was no variation among them. In contrast, the other plant types showed a significant effect of seed size, with the emergence rate of large seeds higher than that of small seeds. The percentage of seedling emergence was regressed against the seed weight of CMS × CRS, CMS × B. rapa and B. rapa ( y = 10.08x+12.68, y in % and x in g for 1000 seeds; F 1,8 = 28 significant at P < 0.001, R 2 = 0.80) to illustrate this relationship (Fig. 1).
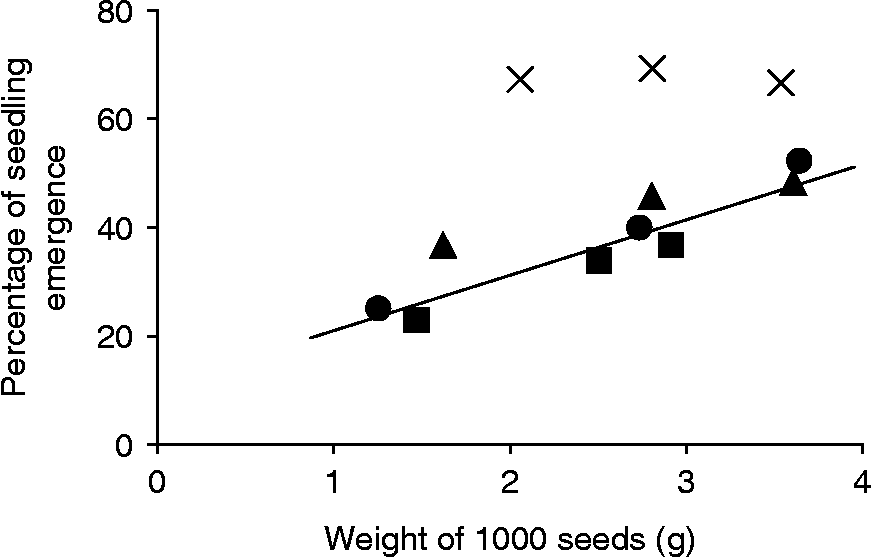
Figure 1 Percentage of seedling emergence of CRS ( × ), CMS × CRS (▲), CMS × B. rapa (■) and B. rapa (●) seeds according to the 1000 seed weight of each seed-size category in the direct sowing experiment. The regression equation does not include CRS data (see text).
Seedling mortality was observed, especially for the small seed category of the interspecific hybrid CMS × B. rapa and the wild parent B. rapa (47 and 25%, respectively). Seedling mortality was less than 11% for the other seed-size categories and plant types. Several plants had not initiated flowering at the time of harvest (Table 3). The lowest percentage of flowering plants was observed for the small seed size of CMS × B. rapa and B. rapa, but there was no difference among seed categories for CRS and CMS × CRS (Table 3). Combining the performance for seedling emergence and survival and percentage of flowering plants can provide an estimate of the likelihood of seed categories participating in reproduction: c. 45% of the large seeds of CMS × CRS, CMS × B. rapa and B. rapa, while only 36 and 20% of the medium and small seeds, respectively, produced adult plants.
Table 3 Plant characteristics from the direct seeding experiment for the four plant types of the cross between oilseed rape and B. rapa, and the three seed categories: height, diameter, flowering time and biomass (±95% confidence limits)

DAS, days after sowing; L, large; M, medium; S, small.
Different letters within a column indicate significantly different values at P < 0.05.
There were significant differences among the four plant types for all four characters analysed by two-way ANOVA: height (F 3,265 = 9.6, P < 0.001), plant diameter (F 3,265 = 3.8, P = 0.011), flowering time (F 3,265 = 18.2, P < 0.001), and biomass (F 3,265 = 3.5, P = 0.016). The B. rapa parent was the smallest, but the interspecific hybrid CMS × B. rapa had the same height as B. napus plants. CMS × B. rapa had the largest plant diameter and biomass, but it flowered later than the other plant types. No difference was found between different seed-size categories for plant height and diameter, while flowering time was longer with small seeds (F 2,265 = 3.6, P = 0.029), and biomass was lower for small seeds but bordered on significance (F 2,265 = 2.9, P = 0.057). Small seeds resulted in lower biomass, especially in B. rapa, and the longest time to flower, especially for CMS × CRS and B. rapa. No interaction was observed between seed category and plant type.
Discussion
Seed size of hybrids
The small seed category represented the largest fraction of the seeds produced in the cages in terms of number of seeds, 74.3% on average. It was not so different from previously harvested field trials (Chèvre et al., Reference Chèvre, Eber, Darmency, Fleury, Picault, Letanneur and Renard2000), where small seeds represented 5% of the weight but 66% of the seed number. As expected from data published earlier (Eber et al., Reference Eber, Chèvre, Baranger, Vallée, Tanguy and Renard1994; Jørgensen and Andersen, Reference Jørgensen and Andersen1994; Bing et al., Reference Bing, Downey and Rakow1996; Chadoeuf et al., Reference Chadoeuf, Darmency, Maillet and Renard1998), in the case of B. napus as the female parent, a higher frequency of interspecific hybrids was found among the seeds produced by the male sterile CMS plants in the cages with B. rapa, while the frequency was much lower in the cages with the other species. Hybrids were found only in the small seed-size category, thus confirming previous studies on B. juncea, R. raphanistrum and H. incana (Eber et al., Reference Eber, Chèvre, Baranger, Vallée, Tanguy and Renard1994; Bing et al., Reference Bing, Downey and Rakow1996; Chadoeuf et al., Reference Chadoeuf, Darmency, Maillet and Renard1998; Chèvre et al., Reference Chèvre, Eber, Darmency, Fleury, Picault, Letanneur and Renard2000). This was also the case for hybrids with B. nigra. No data have been reported to date on seed size of hybrids when the female parent was the wild species, but it is thought that in most cases with Brassica, hybrid seeds could be distinguished from those produced by self-pollination because of their smaller size (Heyn, Reference Heyn1977).
Disturbed development of Brassica hybrid seeds is frequent when parents have different numbers of chromosomes (Howard, Reference Howard1939; Håkansson, Reference Håkansson1956). Paternal genomes for B. nigra (2n = 16, BB), B. rapa (2n = 20, AA), H. incana (2n = 14, AdAd), and R. raphanistrum (2n = 18, RrRr) are smaller than the maternal genome (2n = 38, AACC), and therefore could result in maternal genomic excess and problems with seed development. However, hybrids were found in all three size categories in the hybridization between B. napus and B. rapa, including the large size, which was slightly at variance with the observations of Jørgensen and Andersen (Reference Jørgensen and Andersen1994). In addition, B. juncea (2n = 36, AABB) has a similar chromosome number to that of B. napus and produced only small hybrid seeds, which casts serious doubt on the hypothesis of maternal genomic excess. However, the genetic proximity of B. napus and B. rapa, which share the A genome and highest homology between A and C genomes (Truco et al., Reference Truco, Hu, Sadowski and Quiros1996), might explain the lack of hybridization effect on seed size.
From a practical point of view, the production of small-seeded hybrids is important because small seeds are generally sieved out by the combine harvester and fall onto the soil, so that they represent the major risk of transgene escape in the fields. The relationship between seed size and persistence in the soil of arable fields is complex, but it is commonly assumed that the survival of small-sized seeds of weeds is higher in conventional tillage systems (Ghersa and Martinez-Ghersa, Reference Ghersa and Martinez-Ghersa2000). Interspecific hybrid seeds survived in the soil as well as seeds of oilseed rape (Chadoeuf et al., Reference Chadoeuf, Darmency, Maillet and Renard1998).
Growth of plants from various seed sizes
Greenhouse germination of small seeds was lower in four out of the five studied cases. A similar trend was observed in the direct sowing experiment, where seedling survival was also reduced when the seed size was small. In addition, plants coming from small seeds in the direct seeding experiment flowered later and were lighter at harvest (especially for B. rapa). These effects fit the assumption that seedlings from large-seeded species have higher rates of survival than seedlings from small-seeded species, which could reduce plant fitness (Westoby et al., Reference Westoby, Falster, Moles, Vesk and Wright2002; Moles and Westoby, Reference Moles and Westoby2004). There already exist a few reports of similar seed-size effect on seedling performance in Brassica (Ahmed and Zuberi, Reference Ahmed and Zuberi1973; Major, Reference Major1977; Kumar et al., Reference Kumar, Tomer, Kumar and Chaudhary2005).
No seed-size effect was observed in the transplant experiment with the same plant material and similar growth conditions. Optimum germination and emergence conditions probably cancelled negative seed-size effects in the greenhouse, compared to what was observed in the field. It is noteworthy that the germination protocol in vermiculite, followed by transfer of the young seedlings to Jiffy-7 pellets, was designed especially to prevent any harsh treatment to the seedlings, and there was no plant competition among plots. This was close to what Stanton (Reference Stanton1984) had observed with seed-size differences in R. raphanistrum. She found a significant effect of seed size on final plant size in a field experiment, but not in the greenhouse-growth studies. She also pointed out the impact of the presence or absence of competitive inequities among neighbours within the experimental design. Indeed, spatial heterogeneity and direct competition between plants of different seed categories could greatly enhance growth differences. In our study, we chose to mimic an oilseed rape field in order to provide homogeneous aboveground growth conditions, so that the direct neighbours of every studied plant were CRS oilseed rape, thus establishing similar competition pressure for all the studied plants. This design reduced the variances of the measured traits, although they were still very high, but probably avoided enhancement of plant differences. Assuming that the homogenizing effect of greenhouse germination and transplanting could be generalized to the other plant material of the transplant experiment, this could explain why very few seed-size effects on developmental traits were recorded in our work. Such a difference between growth cabinet and field conditions confirms the need for genuine experimental field conditions as major elements of fitness estimates (Guéritaine et al., Reference Guéritaine, Bazot and Darmency2003; Hauser et al., Reference Hauser, Damgaard and Jørgensen2003). In contrast, growth cabinet/greenhouse experiments can be used to further examine the effects of stress conditions on such variables, allowing a more in-depth understanding of the factors explaining the differences observed in the field, where some factors cannot be controlled.
Paternal effects affecting hybrids
Although the genetic contribution of the same maternal CMS parent made the hybrids less diverse than their wild parents, not all hybrids could be grouped in a single general class. The interspecific hybrids CMS × B. rapa, CMS × B. juncea and CMS × R. raphanistrum appeared to be intermediate between their respective parents, while CMS × B. nigra was closer to B. napus, and CMS × H. incana was closer to its wild parent. These results bear out those of previous studies (Lefol et al., Reference Lefol, Danielou, Darmency, Boucher, Maillet and Renard1995; Linder and Schmitt, Reference Linder and Schmitt1995; Guéritaine et al., Reference Guéritaine, Bazot and Darmency2003; Ammitzbøll et al., Reference Ammitzbøll, Mikkelsen and Jørgensen2005). Therefore, the contribution of the paternal parent could account for differential survival of hybrids in the fields. In particular, in the case of B. rapa, the unique occurrence of large hybrid seeds matches with the only case of natural hybrids reported with B. napus in floras, namely B. × harmsiana O.E. Schulz. (Stace, Reference Stace1975). No other botanists have reported such hybrid taxa in the case of the other species. Beyond the fact that weed control operations in subsequent crops are putting a severe constraint on the development of hybrid populations, the small seed size of most hybrids is probably the cause of lower hybrid establishment in the fields, which could explain the lack of, or extremely low occurrence of, such hybrids in the wild. Independent of the hybridization rate, the extreme rarity of successful hybrids reaching the adult stage hampers the first step of gene flow. The low fertility of hybrids will add a second barrier to the evolution of their progeny, even when they bear beneficial transgenes.
Conclusion
The results for B. rapa showed that the optimum conditions of germination and juvenile growth had masked the differential growth potential of the different seed sizes as observed in heterogeneous field conditions. The difference between the two types of experiment, transplant versus direct seeding, could affect the assessment of the risk of transgene escape by ignoring the very initial stage of hybrid development and overestimating the frequency of hybrid occurrence in the field. Most published studies have estimated the fitness of already established hybrids, thus dismissing the importance of field emergence. Although the small seed-size category falls well within the range of natural variation for each species, this seed class contributes weakly to the adult cohort in the population when co-occurring with another seed class. In most cases, small-seeded hybrids make interspecific hybrid establishment in the field very unlikely, which could hamper the gene flow to some extent. Agricultural practices, such as hindering the escape of small seeds by sorting them with an adequate device on the combine harvester, or the prevention of hybrid seedling establishment by growing an early competitive oilseed rape cultivar, could further reduce the risk of gene flow. However, in the case of B. rapa, when a genetic factor explaining the different seed sizes of hybrids exists, the higher emergence and survival of large-seeded hybrids could be inherited through gene hitchhiking in the progeny, thus modifying the trait in the subsequent generations, which is an additional risk towards establishing ‘super-weed’ populations.
Acknowledgements
The authors thank A. Fleury for competent help in preparing the experiments, and the China Scholarship Council and the Burgundy Region Council for grants. The first author is partially supported by a project of National Natural Science Foundation of China (grant no. 30670316). The INRA experiments were conducted in Dijon, France.




