


INTRODUCTION
Tropical moist forests (TMF) are generally known for their high species diversity (Valencia et al. Reference VALENCIA, BALSLEV and MINO1994, Wright Reference WRIGHT2002). However, in some places, they may be dominated by one species, which sometimes covers several km2 (Hart et al. Reference HART, HART and MURPHY1989, Letouzey Reference LETOUZEY1970). When 50–100% of the trees belong to one species, these forests are called monodominant forests (Connell & Lowman Reference CONNELL and LOWMAN1989). Monodominance may be the result of one or the combination of several mechanisms: creation of a large seed bank, the use of a mycorrhizal network and its effect on nitrogen cycling, monopolization of light, and adaptation to soil properties (Connell & Lowman Reference CONNELL and LOWMAN1989, Corrales et al. Reference CORRALES, MANGAN, TURNER and DALLING2016, Henkel Reference HENKEL2003, Torti et al. Reference TORTI, COLEY and KURSAR2001).
Several studies have tested whether Gilbertiodendron dewevrei (De Wild) J. Léonard-dominated forests (GDF) developed on particular soils, by comparing the physical and chemical properties of their soils to those of soils under adjacent mixed forests (AMF). Most found no significant or biologically meaningful differences in any of the soil variables studied (Conway Reference CONWAY1992, Hart et al. Reference HART, HART and MURPHY1989, Kearsley et al. Reference KEARSLEY, VERBEECK, HUFKENS, VAN DE PERRE, DOETTER, BAERT, BEECKMAN, BOECKX and HUYGENS2017, Peh et al. Reference PEH, SONKE, LLOYD, QUESADA and LEWIS2011). These results are surprising, because, as some legume (Fabaceae) species are known to acidify soils (Hinsinger et al. Reference HINSINGER, PLASSARD, TANG and JAILLARD2003), and as G. dewevrei is a legume, soils under GDF should be more acid, and be characterized by higher base cation depletion than soils under AMF. Regarding organic matter dynamics, other studies found similar annual leaf litter fall under the two types of forest but thicker litter, lower decomposition rates, lower level of available nitrogen litter (NO3-N and NH4-N), and higher C concentration in soils under GDF than in soils under AMF (Cassart et al. Reference CASSART, ANGBONGA, HUGUES, ENRIQUE and QUENTIN2017, Kearsley et al. Reference KEARSLEY, VERBEECK, HUFKENS, VAN DE PERRE, DOETTER, BAERT, BEECKMAN, BOECKX and HUYGENS2017, Torti et al. Reference TORTI, COLEY and KURSAR2001) as Corrales et al. (Reference CORRALES, MANGAN, TURNER and DALLING2016) reported for a monodominant forest in Panama. These results are consistent with those of other recent studies suggesting that species associated with ectomycorrhizal fungi – like G. dewevrei (Onguene & Kuyper Reference ONGUENE and KUYPER2001, Torti & Coley Reference TORTI and COLEY1999) – compete directly for nitrogen with free-living decomposers which degrade the litter (Averill et al. Reference AVERILL, TURNER and FINZI2014, Corrales et al. Reference CORRALES, MANGAN, TURNER and DALLING2016, Orwin et al. Reference ORWIN, KIRSCHBAUM, JOHN and DICKIE2011).
The reason why most previous studies did not obtain the same results is certainly because some (Peh et al. Reference PEH, SONKE, LLOYD, QUESADA and LEWIS2011) did not take topography into account. Topography is known to be a key factor which locally affects both the texture and the chemical properties of the soil (Lieberman et al. Reference LIEBERMAN, LIEBERMAN, HARTSHORN and PERALTA1985, Sabatier et al. Reference SABATIER, GRIMALDI, PREVOST, GUILLAUME, GODRON, DOSSO and CURMI1997, Valencia et al. Reference VALENCIA, FOSTER, VILLA, CONDIT, SVENNING, HERNANDEZ, ROMOLEROUX, LOSOS, MAGARD and BALSLEV2004). When topography is not taken into account, the variability of soil properties under the same type of forest can be high, making the null hypothesis difficult to reject.
The aim of the present study was to compare the physical and chemical properties of soils under GDF and under AMF in the Yoko forest reserve in the Democratic Republic of Congo, considering topography as an important factor. We hypothesized that (1) soils under GDF are more acid and have lower concentrations of base cations than soils under AMF; (2) soils under GDF have thicker litter, higher C concentrations, lower concentrations of total N and a higher C:N ratio than soils under AMF.
METHODS
Study site
The study site is located in north-eastern Democratic Republic of Congo (DRC), 32 km south-east of Kisangani, in a dense lowland (450–500 m) moist forest in the Yoko forest reserve, between the villages of Banango (00°21′N, 025°4′E) and Bagao (00°07′N, 025°18′E). The forest reserve covers a total area of 6975 ha in two north–south blocks separated by the River Yoko.
The reserve is characterized by a main plateau ranging from 450 to 470 m a.s.l., dissected by a drainage network. The transition between the plateaux and the streams is abrupt and, in most cases, characterized by short but steep (~50%) convex slopes. The climate is equatorial, Af type according to the Köppen classification. Based on the meteorological data collected from the Kisangani between 1956 and 2005, mean annual rainfall was about 1720 mm with two distinct rainy seasons (March–May and September–November), a dry season (January–February) with monthly precipitation < 100 mm, and a total intermediate season length of ~4 mo (June–August and December) with monthly precipitation >100 mm and <150 mm (Nshimba Reference NSHIMBA2008, Service National de Climatologie). Mean monthly temperatures ranged from 22.4°C to 29.3°C, and the mean annual temperature is 25°C. The canopy of the multi-strata terra firme Yoko forest reaches 40 m in height, with large gaps filled with Marantaceae. The higher strata are dominated by Guarea thompsonii Sprague & Hutch., Tessmannia africana Harms, Petersianthus macrocarpus (P. Beauv.) Liben, Scorodophloeus zenkeri Harms and Irvingia grandifolia (Engl.). In some places, Gilbertiodendron dewevrei (De Wild.) J. Léonard dominates in the higher strata: Gilbertiodendron dewevrei stands occur across the whole landscape, both on the plateaux and in the bottomlands. The rocks underlying the geological formation of the Kisangani area are mainly Upper Jurassic sedimentary sequences, and include argillites and sandstones (Kadima et al. Reference KADIMA, DELVAUX, SEBAGENZI, TACKW and KABEYAZ2011). The soils in the Yoko forest reserve are Xanthic Ferralsols and Dystric Arenosols (Ngongo et al. Reference NGONGO, VAN RANST, BAERT, KASONGO, VERDOODT, MIJINYA and MUKALAY2009). The Ferralsols on the plateau are of such low base status that they qualify as Geric, whereas those in the bottomlands are less leached and some may be Eutric. The Arenosols are similar, some qualifying as Eutric in the bottomlands. The plateau soils are very well drained, ranging from dark brown in colour (7.5YR 3/3) in the 0–5 cm soil layer, to strong brown (7.5YR 4/6) in the 120–200 cm layer. The texture ranges from loamy sand to sandy loam. Faunal activity is rare (a few ants and termites). The bottomland soils are poorly drained, brown in colour (7.5YR 4/3) in the 0–10 cm soil layer, to reddish yellow (7.5YR 6/6) with mottling (2.5YR 3/6) in the 100–110 cm layer, then light greenish grey (Gley1 8/10Y) in the 110–160 cm layer. The texture ranges from sandy loam to sandy clay loam. There are many coarse fragments in the 80–160 cm layer, with in situ argillite below.
Experimental design
Sixteen 0.25-ha plots (50 × 50 m, Figure 1) were set up taking two factors into account: topography, i.e. plateau vs. bottomland, and forest type, i.e. GDF vs. AMF. Four replicates were performed for each combination of the two factors (plateau, GDF), (plateau, AMF), (bottomland, GDF), (bottomland, AMF). The 16 plots were spatially distributed as eight pairs (GDF, AMF), with each pair sharing the same topographic position (plateau or bottomland) and the same environment (altitude, slope, soil type). The maximum distance between the two plots forming a pair was 150 m, while the distance between pairs was between 150 and 4000 m.

Figure 1. Location of the 16 0.25-ha plots in Yoko forest reserve in the Democratic Republic of Congo. The colours are the Digital Elevation Model values based on Shuttle Radar Topography Mission (SRTM) at a resolution of 3 arc seconds.
Six of the eight GDF plots were qualified as monodominant G. dewevrei forests as defined by Connell & Lowman (Reference CONNELL and LOWMAN1989), with the relative species basal area ≥50%. This relative basal area was slightly smaller in the two last GDF plots (43% and 46%, respectively) (Table 1). We inventoried all the trees with a diameter at breast height (dbh) ≥10 cm. Each tree was spatially located inside each plot, its dbh was measured, and it was botanically identified. In the centre of each plot, a soil pit was excavated to a depth of 150–200 cm. The thickness of the litter and the depth reached by the coarse roots were measured. Soils were sampled at five depths: 0–5 cm, 5–10 cm, 10–20 cm, 20–40 cm and 120–150 cm. The total number of samples was 80 (16 plots × 5 depths). The samples were analysed in the CIRAD, IFA-Yangambi and Kisangani University soil laboratories. To make the results of our study easier to compare with those of previous studies, only the results of soil analysis of the topsoil (0–5 cm layer) and of the deep soil (120–150 cm layer) are presented here.
Table 1. Characteristics of eight 1-ha plots of monodominant Gilbertiodendron dewevrei forest (GDF) and eight 1-ha plots of adjacent mixed forest (AMF), as a function of topography (plateau vs. bottomland), in Yoko forest reserve, Democratic Republic of Congo. Altitude was deduced from a Shuttle Radar Topography Mission (SRTM), 90 m resolution model.

Soil analysis
Soil pH was measured with a soil:deionized water ratio of 1:2.5. Particle size distribution was determined using the pipette method and five classes: clay (<2 µm), fine silt (2–20 µm), coarse silt (20–50 µm), fine sand (50–200 µm) and coarse sand (200 µm–2 mm). Total organic carbon and total nitrogen were determined by dry combustion and measured with a thermal conductive detector. Phosphorus was extracted with 0.5 M ammonium fluoride (NH4F) and 0.5 M sodium bicarbonate (NaHCO3) at pH 8.5 (Dabin Reference DABIN1967), and measured using continuous flux spectrometry. Exchangeable Ca, Mg, K and Na were extracted during a saturation step using 1M ammonium acetate (pH7) and then measured using inductively coupled plasma atomic emission spectrometry. After the saturation step, the residue was washed with ethyl alcohol, and the ammonium extracted using NaCl. Cation exchange capacity (CEC) was determined by measuring extracted ammonium using continuous flux spectrometry. Exchangeable Al and H were extracted using 1M KCl and then titrated against 0.01 M NaOH to pH7 (Pansu & Gautheyrou Reference PANSU and GAUTHEYROU2006).
Statistical analyses
Covariance analysis was used to compare the soil chemical parameters according to the topographic factor (plateau vs. bottomland), the forest type (GDF vs. AMF) and the combined clay + fine silt (<20 µm) content, as the last parameter is often correlated with most soil chemical parameters, such as soil organic carbon (Zinn et al. Reference ZINN, RATTAN and RESCK2005). Covariance analysis is appropriate when the aim is to separate the effects of topography, forest type and texture on the chemical properties of soils. Because some of the soil parameters were clearly non-normal, we transformed them with a log function before performing covariance analysis.
RESULTS
The mean tree density across the 16 plots was 372 ± 62 ha−1. Neither forest type, nor topography had any effect on this density. The density of Fabaceae trees was significantly higher in GDF stands (151 ± 54 trees ha−1, relative density: 42.0% ± 9.0%) than in AMF stands (90 ± 38 trees ha−1, relative density: 23.6% ± 10.2%) (Table 1). GDF and AMF stands had a negative exponential stand structure on both the plateaux and bottomlands (Appendix 1), which is typical of natural uneven-aged moist forests. The litter was significantly thicker in GDF plots (10.6 ± 3.6 cm) than in AMF plots (4.6 ± 2.0 cm), whereas there was no effect of topography on litter depth (Figure 2).

Figure 2. Litter thickness (a); and soil C:N ratio in the 0–5 cm soil layer (b) as a function of topography (plateau vs. bottomland) and forest type (GDF = monodominant Gilbertiodendron dewevrei forest vs. AMF = Adjacent mixed forest), in Yoko forest reserve in the Democratic Republic of Congo. The boxes represent the first and the third quartile, the whiskers the minimum and maximum, the bars the median value, and the crosses the mean values.
Soil texture varied considerably in Yoko forest reserve: clay, silt and sand contents ranged from 4–50%, 2–37% and 40–94%, respectively. Most of the soils were loamy sand, sandy loam and sandy clay loam (Figure 3). Topography was the determining factor for soil texture in this site: soils were generally sandier on the plateaux and more clayey in the bottomlands. In the bottomlands, soils also became more silty when they reached the saprolite (Figure 3). Clay + fine silt content ranged between 5% and 61% in the whole soil profile, and was positively correlated (between 0.23 and 0.92) with most of the soil chemical properties, except available P and pH (no correlation) and the C:N ratio (negative correlation). Results were similar in the 0–5-cm soil layer. Finally, the more sandy soils on the plateaux had a lower concentration of C, total N, CEC, Ca2+, Mg2+, K+, Na+, Al3+, H+ than the more clayey soils in the bottomlands. Conversely, they had a significantly higher C:N ratio, both in the 0–5-cm soil layer (14.3 vs. 9.2) and in the 120–150-cm soil layer (11.6 vs. 6.5) (Table 2, Figure 2).

Figure 3. Comparison of the proportion of clay (a); proportion of silt (b); and proportion of sand (c) found in soils at different depths sampled as a function of topography (plateau vs. bottomland) and forest type (GDF = Gilbertiodendron dewevrei forest vs. AMF = adjacent mixed forest). Plateau soils are in black and bottomland soils are in blue. The solid lines represent soils under GDF and the dotted lines soils under AMF.
Table 2. Summary of chemical properties of soils (averages ± SD) in the Yoko forest reserve as a function of topography (P = plateau; B = bottomland) and forest type (GDF = monodominant Gilbertiodendron dewevrei forest; AMF = Adjacent mixed forest) for the 0–5 cm soil layer and the 120–150 cm soil layer. P, extracted using the Olsen-Dabin method; CEC, cation exchange capacity; TEB, total exchangeable bases (= Ca2+ + Mg2+ + K+ + Na+); BS, base saturation (= TEB/CEC × 100). Al, exchangeable Al3+ concentration; H, exchangeable H+ concentration. All soil characteristics were determined in four pits.
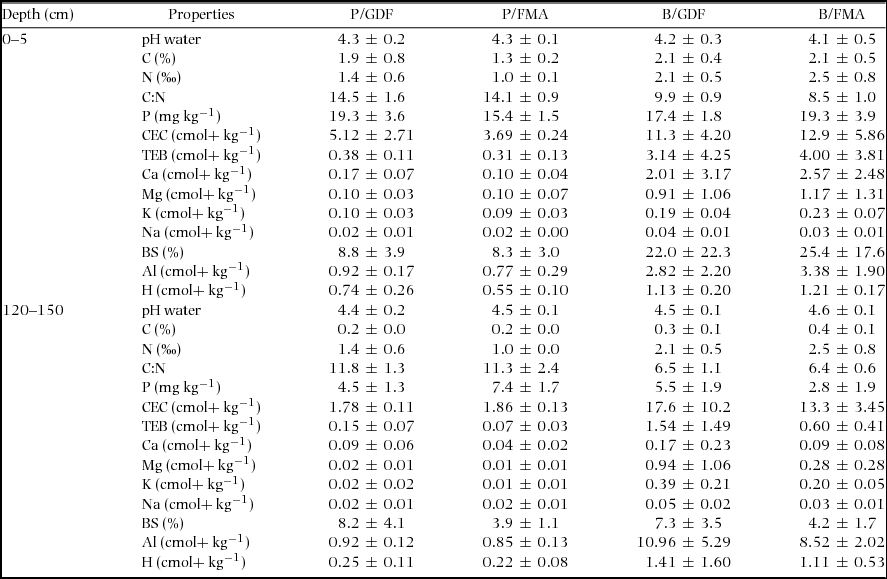
When the clay + fine silt content and topography were taken into account, covariance analysis revealed no significant difference in most of the chemical properties – pH, N, C:N, P, CEC, exchangeable Ca, Mg, K, Al, H – between the soils under GDF and the soils under AMF (Table 3). The type of forest had a significant association only with the soil C concentration in the 0–5-cm soil layer, and with exchangeable Na in the 120–150-cm soil layer. More precisely, for a given clay + fine silt content, under GDF, soils in the 0–5-cm layer had a higher C concentration (mean: 0.5%) than soils under AMF. For a clay + fine silt content > 40%, soils under GDF had a higher exchangeable Na concentration (about 0.02 cmolc kg−1) in the 120–150 cm layer than under AMF (Figure 4).
Table 3. Results of analysis of covariance of chemical properties of soils in the Yoko forest reserve, as a function of clay + fine silt, topography (plateau vs. bottomland), and forest type (monodominant Gilbertiodendron dewevrei forest vs. adjacent mixed forest) in the 0–5 cm and 120–150 cm soil layers. NS: effect not significant, or significant with (P> | t |) <5% (*), or <1% (**) or <1 ‰ (***). A few of these properties were normalized using ln function to ensure normality before the analysis of covariance. The soil properties are abbreviated as in Table 2.

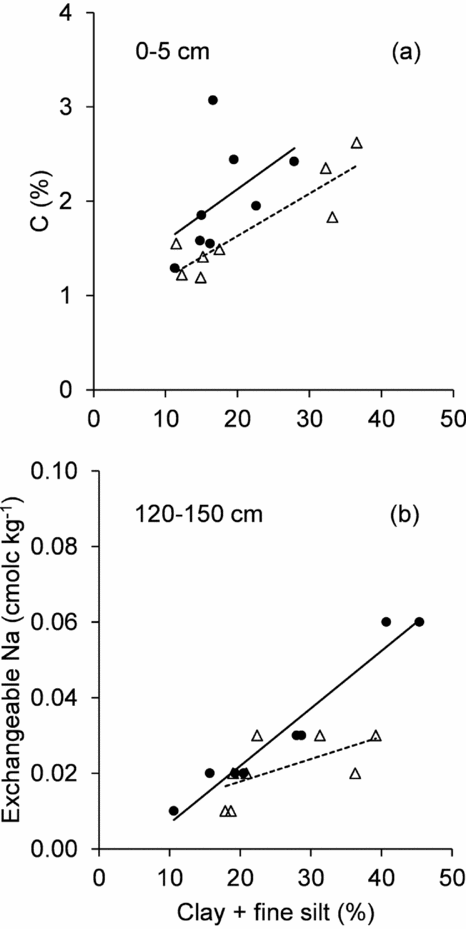
Figure 4. Correlation between clay + fine silt (< 20 µm) and total carbon concentration in the 0–5 cm soil layer (a); and exchangeable Na concentration in the 130–150 cm soil layer (b), in soils under the monodominant Gilbertodendron dewevrei forest (black dots) and under an adjacent mixed forest (empty triangles) in Yoko forest reserve in the Democratic Republic of Congo.
DISCUSSION
In the Yoko forest reserve, GDF stands can be found both on the plateaux and in the bottomlands, on soils whose texture varies between loamy sand, sandy loam and sandy clay loam. The GDF stands studied by Peh et al. (Reference PEH, SONKE, LLOYD, QUESADA and LEWIS2011) in the Dja reserve in Cameroon were on more silty soils. Hart et al. (Reference HART, HART and MURPHY1989), who cited several studies in the Democratic Republic of Congo (Uélé, Kivu, Ubangi, Yangambi), reported that GDF stands developed on soils with a variety of textures. As Gérard (Reference GÉRARD1960) and Peh et al. (Reference PEH, SONKE, LLOYD, QUESADA and LEWIS2011) also studied GDF stands growing on a wide range of soils (well drained or with a water table at a depth of 1 m; deep soils or shallow and gravelly soils), in DRC and Cameroon, respectively, all these studies suggest that G. dewevrei can adapt to different types of soil. This ability is certainly reinforced by its rooting plasticity, as reported by Gérard (Reference GÉRARD1960): the species developed taproots in deep soils, and lateral roots in shallow gravelly soils.
Our first hypothesis was that soils under GDF were more acid and had a lower base cation concentration (Ca2+, Mg2+, K+, Na+) than soils under AMF. This hypothesis was not validated, as we found a similar pH in the soils in the two stands, and similar concentrations of all the cations except exchangeable Na, for which we found a slight difference in the 120–150 cm layer between soils under GDF and soils under AMF. This result is in agreement with that obtained by Peh et al. (Reference PEH, SONKE, LLOYD, QUESADA and LEWIS2011), but not with that obtained by Hart et al. (Reference HART, HART and MURPHY1989), who found a slightly higher pH (pH = 4.2 vs. 4.0) and slightly lower concentration of exchangeable K (0.05 vs. 0.11 cmol+ kg−1) in soils under GDF than in soils under AMF at depths of 20 cm and 150 cm, respectively. Two main factors could explain these results: (1) the moist tropical climate is the main factor that acidifies soils, which would mask any minor differences in acidity in soils under GDF and under AMF; (2) as Fabaceae are also abundant in AMF stands, they could acidify soils in the same way as in GDF stands.
Our second hypothesis was that soils under GDF have thicker litter, higher C concentration, lower total N concentration and higher C:N ratio than soils under AMF. This hypothesis was partially validated: on one hand, we found that the litter layer was 2.3 times thicker under GDF than under AMF stands. This result is in agreement with the results of Torti et al. (Reference TORTI, COLEY and KURSAR2001). Furthermore, for a given clay + fine silt content, we found that the mean C concentration of the topsoil under GDF (i.e. the 0–5 cm soil layer) was 0.5% higher than that of the topsoil under AMF. This result is partly in agreement with that of Kearsley et al. (Reference KEARSLEY, VERBEECK, HUFKENS, VAN DE PERRE, DOETTER, BAERT, BEECKMAN, BOECKX and HUYGENS2017), who also found higher C concentrations in soils under GDF than in soils under AMF, but also in deeper soil layers, i.e. from the 0–30-cm to the 60–90-cm layers. Similar results to those of Kearsley et al. (Reference KEARSLEY, VERBEECK, HUFKENS, VAN DE PERRE, DOETTER, BAERT, BEECKMAN, BOECKX and HUYGENS2017) were obtained by Cassart et al. (Reference CASSART, ANGBONGA, HUGUES, ENRIQUE and QUENTIN2017) from the 0–10-cm to the 75–100-cm layers. By contrast, our result does not agree with that of Peh et al. (Reference PEH, SONKE, LLOYD, QUESADA and LEWIS2011), who found similar C concentrations in the soils under the two types of stand, even in the 0–5 cm layer. The difference between the results of the two studies could be due to the different methods of statistical analysis of data used. As a matter of fact, if we had analysed our data in the same way as Peh et al. (Reference PEH, SONKE, LLOYD, QUESADA and LEWIS2011), i.e. without taking the clay + fine silt content into account, we would not have found a significant difference in C concentration in the soils under the two types of stand. Our results can be considered as intermediate between those of Kearsley et al. (Reference KEARSLEY, VERBEECK, HUFKENS, VAN DE PERRE, DOETTER, BAERT, BEECKMAN, BOECKX and HUYGENS2017) and Cassart et al. (Reference CASSART, ANGBONGA, HUGUES, ENRIQUE and QUENTIN2017) vs. Peh et al. (2001). On the other hand, we found no significant difference in total N concentrations in soils under GDF and under AFM (Table 3), like Peh et al. (Reference PEH, SONKE, LLOYD, QUESADA and LEWIS2011) and Kearsley et al. (Reference KEARSLEY, VERBEECK, HUFKENS, VAN DE PERRE, DOETTER, BAERT, BEECKMAN, BOECKX and HUYGENS2017). Torti et al. (Reference TORTI, COLEY and KURSAR2001) found the mineral N concentration to be three times lower in soils under GDF than under AMF. The results of their study suggest that mineral N concentration is a limiting factor in soils under GDF involving competition between G. dewevrei ectomycorrhizal roots and the soil litter decomposers (Onguene & Kuyper Reference ONGUENE and KUYPER2001, Torti & Coley Reference TORTI and COLEY1999). More generally, an emergent property of the ectomycorrhizal monodominant forests could be reduced nitrogen availability (Corrales et al. Reference CORRALES, MANGAN, TURNER and DALLING2016, Orwin et al. Reference ORWIN, KIRSCHBAUM, JOHN and DICKIE2011), which can in turn influence soil organic matter (Averill et al. Reference AVERILL, TURNER and FINZI2014). Lastly, as we found a similar C:N ratio in the soils under GDF and under AMF, whereas the thickness of the layer of litter differed significantly, unlike in other studies (Springob & Kirchmann Reference SPRINGOB and KIRCHMANN2003), our study suggests that the C:N ratio is not necessarily a good proxy of litter degradation and organic matter mineralization.
CONCLUSION
Our study clearly demonstrated that, for a given topography, most of the physical and chemical properties of the soils under Gilbertiodendron dewevrei did not differ from those of the soils under an adjacent mixed forest, in particular soil acidity, base cation concentration, total N concentration, and the C:N ratio. On the other hand, we found an emergent difference between the two forest stands in terms of organic matter dynamics: the litter was thicker and, for a given clay + fine silt content, the total C concentration in the 0–5 cm soil layer was slightly higher in soils under GDF than in soils under AMF. However, one limit of our study was not to have measured soil mineral N and the activity of litter decomposers. This could have validated or generalized the results obtained by other studies (Cassart et al. Reference CASSART, ANGBONGA, HUGUES, ENRIQUE and QUENTIN2017, Kearsley et al. Reference KEARSLEY, VERBEECK, HUFKENS, VAN DE PERRE, DOETTER, BAERT, BEECKMAN, BOECKX and HUYGENS2017, Torti et al. Reference TORTI, COLEY and KURSAR2001) which reported low availability of nitrogen and a low litter decomposition rate in G. dewevrei stands. These characteristics could play an important role in maintaining the monodominant G. dewevrei stands, whose origin remains to be identified.
ACKNOWLEDGEMENTS
This study was conducted as part of M. Lokonda's PhD, and was supported by a grant from the European Community through the Re2AFor and REFORCO projects. We are grateful to the CIRAD soil laboratory for conducting some of the physical and chemical analyses. We thank all the people who helped develop this work: M. Bola, R. Aundja, A.W. Doka, L. Ndjele, P. Sabongo and C. Lomba. We thank two anonymous reviewers for their insightful comments on a former version of the manuscript.
Appendix 1.
Comparison of the stand structure on the plateaux (a); and bottomlands (b) of Gilbertiodendron dewevrei stands (green bars) and adjacent mixed forest stands (empty bars). The bars represent the standard deviation.








