



Introduction
Cattle and sheep, along with farming, were introduced to southern England c. 4000 cal BC, reaching Orkney off the north coast of Scotland c. 3500 cal BC during the final stage of the spread of Neolithic lifeways across Europe (Tresset & Vigne Reference Tresset and Vigne2011; Whittle et al. Reference Whittle, Healy and Bayliss2011; Bayliss et al. Reference Bayliss, Marshall, Richards and Whittle2017). The success of this expansion across the continent, starting in the seventh/sixth millennia BC, involved socio-economic, cultural and biological reconfigurations (Tresset & Vigne Reference Tresset and Vigne2011), including innovations in husbandry practices and physiological changes in animals. Adaptation to new environmental settings and pasture resources, and the consequences of these changes on animal feeding practices, were of particular importance.
In contrast with continental Europe, where Holocene deciduous forests provided tree-leaf fodder in times of a scarcity of grass, on Orkney, farming was introduced into a predominantly treeless landscape (Farrell et al. Reference Farrell, Bunting, Lee and Thomas2014; Bunting et al. Reference Bunting, Farrell, Bayliss, Marshall and Whittle2018). Traditionally, marine seaweed has been used as an alternative fodder in coastal North-western Europe (Amorosi et al. Reference Amorosi, Buckland, Edwards, Mainland, McGovern, Sadler and Skidmore1998). This practice still persists in the present day on Orkney, where it has reached an extreme level in the sheep farming on North Ronaldsay, the northernmost island of the archipelago. Confined to the shore by a dike (or drystone wall) constructed in 1832 around the entire island, these sheep now feed exclusively on seaweed (MacLachlan Reference MacLachlan1988)—a diet that has produced a series of metabolic adaptations (MacLachlan & Johnston Reference MacLachlan and Johnston1982; Orpin et al. Reference Orpin, Greenwood, Hall and Paterson1985; Feldmann et al. Reference Feldmann, John and Pengsprecha2000). This practice is attested by ethnographic and historical sources from medieval times onwards (Amorosi et al. Reference Amorosi, Buckland, Edwards, Mainland, McGovern, Sadler and Skidmore1998; Balasse et al. Reference Balasse, Tresset and Ambrose2006), although stable isotope analyses have revealed that sheep in Orkney switched to marine resources as early as the third millennium cal BC (Balasse et al. Reference Balasse, Tresset and Ambrose2006, Reference Balasse, Mainland and Richards2009; Balasse & Tresset Reference Balasse, Tresset and Ritchie2009; Schulting & Richards Reference Schulting, Richards and Ritchie2009; Schulting et al. Reference Schulting, Vaiglova, Crozier and Reimer2017).
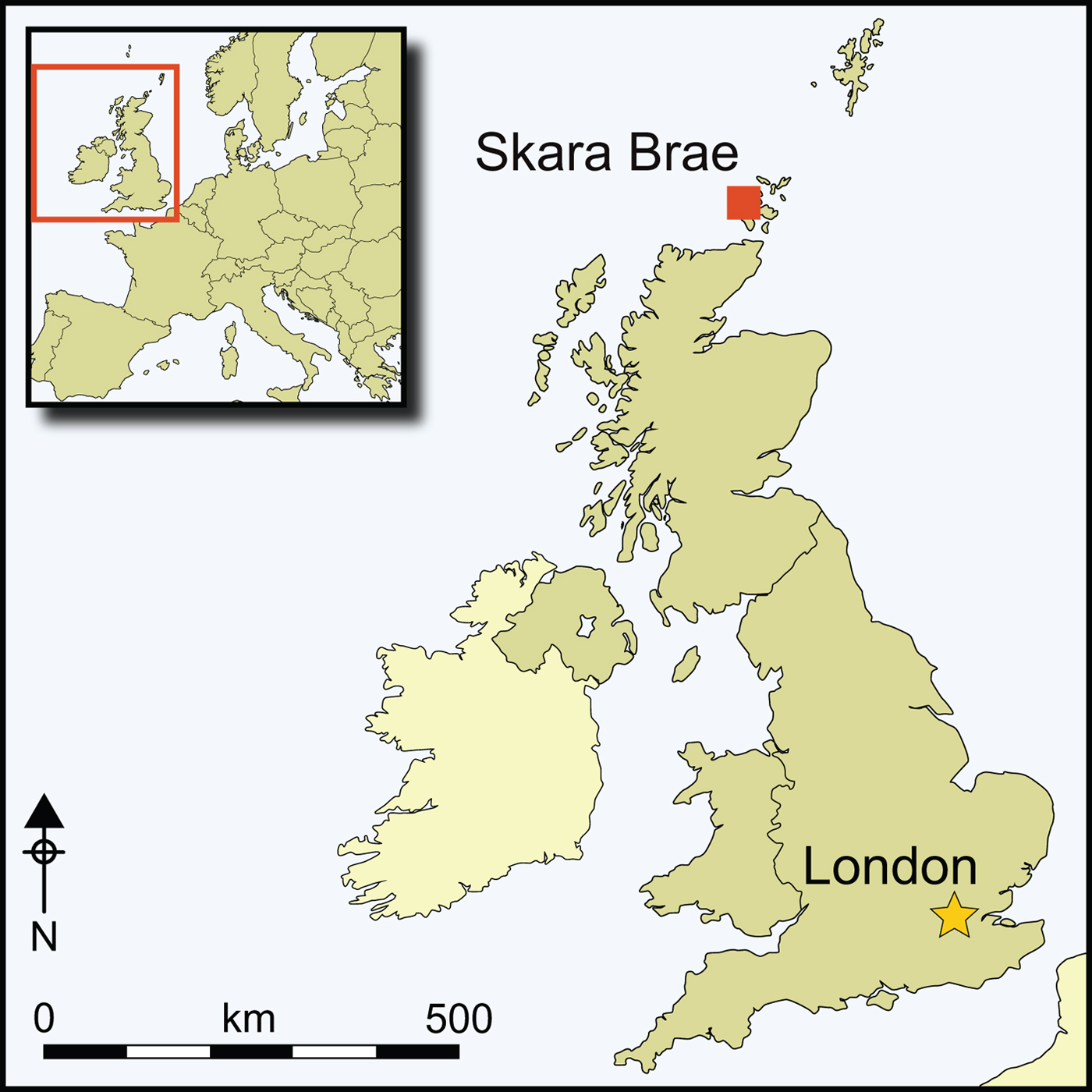
To date, seaweed consumption has not been identified at the Knap of Howar on Papa Westray, the site of the earliest introduction of sheep farming to Orkney in the fourth millennium cal BC (Figure 1). This contrasts sharply with the systematic contribution of seaweed to the winter sheep diet in the first half of the third millennium attested at the chambered cairn at Holm of Papa Westray (Figure 1) (Balasse et al. Reference Balasse, Tresset and Ambrose2006; Balasse & Tresset Reference Balasse, Tresset and Ritchie2009). The 500 years separating these two sheep populations may partly explain this difference in feeding practices. This chronological argument implies that there may have been a time lag between the introduction of the sheep to Orkney and the development of a physiological adaptation for the significant ingestion of seaweed. Alternatively—or additionally—the isolated location of Holm of Papa Westray on a small islet (Figure 1), may have involved additional environmental constraints, introducing a remoteness argument to explain seaweed consumption by sheep at this site. A few other indisputable examples of seaweed-eating sheep have been reported in Neolithic Orkney that are potentially contemporaneous with the sheep from Holm of Papa Westray: one specimen from the Point of Cott burial cairn from the third millennium cal BC (Balasse et al. Reference Balasse, Mainland and Richards2009), and two specimens from Quanterness, which date to the early to mid third millennium cal BC (Schulting et al. Reference Schulting, Vaiglova, Crozier and Reimer2017). The location of these sites on the Westray and Mainland islands (Figure 1) demonstrates that the remoteness argument alone cannot account for the introduction of seaweed into the sheep diet.

Figure 1. Location of the archaeological sites in Orkney (figure by M. Balasse).
In the present study, we investigate the role of seaweed in Neolithic husbandry at Skara Brae, in the Bay of Skaill on Mainland Orkney (Figure 1). Excavations at the settlement have yielded a large assemblage of cattle and sheep remains (Clarke Reference Clarke1976). Skara Brae was occupied from c. 3400–2500 cal BC (Bayliss et al. Reference Bayliss, Marshall, Richards and Whittle2017), contemporaneous with Knap of Howar and Holm of Papa Westray. The first aim of this study is to confirm the chronology of the appearance of seaweed consumption practices and behaviours during the fourth and third millennia BC. The second aim is to test for seaweed consumption amongst Neolithic Orkney cattle. At Knap of Howar, seaweed did not contribute significantly to the diet of cattle (Balasse et al. Reference Balasse, Tresset and Ambrose2006); this hypothesis could not be tested at Holm of Papa Westray, due to the scarcity of cattle remains. At Skara Brae, it is possible to analyse both sheep and cattle populations, thereby providing a more holistic view of Neolithic husbandry. We investigate seaweed-eating at Skara Brae using a new methodological approach, in which co-variations in carbon and oxygen isotope sequences in teeth are modelled as a means of highlighting deviation from the expected seasonal cycle in terrestrial grass δ13C values. The objective is to detect minor contributions of seaweed to animal diet.
Material and methods
Occupation of the small village of Skara Brae can be divided into three phases. The earliest human occupation, phase 0, began c. 3400 cal BC and ended c. 3200 cal BC. This was followed by a 300-year hiatus until the establishment of nucleated settlement (phase 1) c. 2900 cal BC. This first village was abandoned after a short period of use and the settlement re-occupied (phase 2) between c. 2750 and 2500 cal BC (Bayliss et al. Reference Bayliss, Marshall, Richards and Whittle2017). Sheep and cattle teeth were selected for analysis from all of these occupation phases (see the online supplementary material (OSM) Table S1).
Seaweed consumption can be investigated through the analysis of stable carbon isotope ratios. In Orkney, δ13C values varying between −32.8‰ and −29‰ have been measured in modern terrestrial grasses, while marine seaweed collected on the shore have yielded δ13C values fluctuating between −19.9‰ and −16.5‰ (Balasse et al. Reference Balasse, Mainland and Richards2009). These δ13C values are inherited along the food chain. In archaeological animal remains, this signal may be retrieved from the skeleton mineral fraction (bioapatite), whose carbon isotope composition is influenced by diet (Krueger & Sullivan Reference Krueger, Sullivan, Turnlund and Johnson1984). As enamel retains a record of the animal's dietary history over the duration of tooth formation, the changing contribution of seaweed to the diet can be investigated using sequential analysis of tooth enamel. The parallel analysis of oxygen isotope composition (δ18O) allows for the investigation of seaweed consumption on a seasonal scale (Balasse et al. Reference Balasse, Tresset, Dobney and Ambrose2005, Reference Balasse, Mainland and Richards2009).
Seasonal changes in δ13C values are expected in diets based on both terrestrial plants and seaweed. Variations of 1–2‰ have been reported in the δ13C values of C3 terrestrial plants—the prevalent type in this area of Europe—with the highest values occurring during the summer and the lowest during the winter (Smedley et al. Reference Smedley, Dawson, Comstock, Donovan, Sherill, Cook and Ehleringer1991). Seasonal variation in the relative proportion of seaweed species available for sheep grazing on the Orkney shores are also reported. Laminaria digitata and L. hyperborea, seaweed species accounting for over 75 per cent of the modern North Ronaldsay sheep diet in the summer, have higher δ13C values than Palmaria palmata, which contributes around half of their winter diet (Raven et al. Reference Raven2002; Hansen et al. Reference Hansen, Hector and Feldmann2003; Balasse et al. Reference Balasse, Mainland and Richards2009). This leads to higher values in the enamel for a summer seaweed diet compared to winter. Finally, changes in the relative contributions of seaweed and grass to the diet are expected on a seasonal scale, with reduced availability of terrestrial grass in winter coinciding with greater access to seaweed driven ashore by winter storms. This would lead to higher δ13C values for animal diet in the winter, compared with the summer. In all cases, seasonal change in diet is expected to lead to co-variations in enamel δ13C and δ18O values. This co-variation would, however, follow different patterns:
1) A diet composed exclusively of terrestrial plants throughout the year would lead to lower overall δ13C values and in phase seasonal variations in the δ13C and δ18O sequences (pattern 1).
2) A diet composed exclusively of seaweed throughout the year would lead to higher δ13C values and seasonal variations in the δ13C and δ18O sequences (pattern 2).
3) A diet with a higher proportion of seaweed during the winter would lead to in opposite phase variations in the δ13C and δ18O sequences; the higher the relative proportion of seaweed, the higher the δ13C values (pattern 3).
These hypotheses can be tested using δ13C and δ18O sequences from modern sheep with known diets as reference populations (Table S2). Pattern 1 is tested using the grazing sheep from Rousay Island (ROU, Figure 1; Balasse et al. Reference Balasse, Mainland and Richards2009). Additionally, the sample for grazing sheep is enlarged with samples from Kemenez Island (KMZ; Molène, Finistère, France). To check for consistency in δ13C and δ18O sequences between modern and preindustrial times, we also include one sheep from the Bronze Age ‘22 bis’ site on Kemenez Island (Gandois Reference Gandois2017). The references for pattern 2 are sequences measured in modern North Ronaldsay sheep (NR, Figure 1) feeding on seaweed year-round (Balasse et al. Reference Balasse, Tresset, Dobney and Ambrose2005). Recently, parts of the dike encircling North Ronaldsay have fallen into disrepair, allowing sheep increased access to terrestrial grazing. Some of these animals, which feed partly on terrestrial grass during summer (Balasse et al. Reference Balasse, Tresset, Dobney and Ambrose2005, Reference Balasse, Mainland and Richards2009), are used as references for pattern 3.
Enamel from the anterior lobe of the second molar (M2) or the middle lobe of the third molar (M3) was sequentially sampled, following Balasse et al. (Reference Balasse, Tresset, Dobney and Ambrose2005). Powdered archaeological enamel samples were pre-treated to eliminate diagenetically introduced carbonates (0.1M acetic acid for 4h, 0.1ml/mg). Bioapatite samples weighing 580–620μg were reacted with 100% phosphoric acid at 70°C in individual vessels, in an automated cryogenic distillation system (Kiel IV device), interfaced with a DeltaVAdvantage isotope ratio mass spectrometer. The analytical precision within each run—estimated from repeated analyses of our laboratory carbonate standard (Marbre LM, normalised to NBS19)—was lower than 0.06‰ for δ18O values and lower than 0.03‰ for δ13C values.
The phase shift between the δ18O and δ13C sequences was determined through a sinusoidal model (Table S3). When both signals are in phase (i.e. the highest and lowest δ18O and δ13C values are acquired at the same time in the annual cycle), the phase shift would be 0° or 360°. Opposed phased signals (the highest δ18O value is measured when the δ13C value is the lowest, and vice versa) would lead to a 180° shift.
Results
Reference grazing and seaweed-eating sheep
Results from the sequential analysis of the Kemenez sheep are shown in Figure S1. Results from the modelling of the δ13C and δ18O sequences for the Kemenez, Rousay and North Ronaldsay sheep are reported in Table S3, and the phase shift for each specimen is presented in Table S2. Figure 2A shows intra-tooth variations in enamel δ13C and δ18O values in three chosen specimens, illustrative of patterns 1, 2 and 3 respectively. Figure 2B is a plot of the phase shift between the δ13C and δ18O sequences against the highest value measured in the δ13C sequence (δ13Cmax) for all 21 reference specimens. All δ13C values measured in modern sheep were corrected (+1.5‰) for the fossil fuel effect (Francey et al. Reference Francey, Allison, Etheridge, Trudinger, Enting, Leuenberger, Langenfelds, Michel and Steele1999). The δ13Cmax value clearly discriminates between a purely grass diet in pattern 1 (−15.0‰ to −12.9‰) and a purely/predominantly seaweed diet in patterns 2 and 3 (−4.9‰ to −0.4‰).

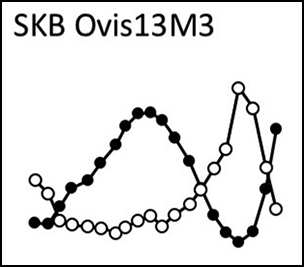
Figure 2. A) Sequential analysis of stable carbon (δ13C) and oxygen (δ18O) isotope ratios in sheep tooth enamel, illustrating patterns 1, 2 and 3. The samples are located in the tooth crown using their distance from the enamel-root junction; B) phase shift between the δ13C and δ18O sequences plotted vs the highest value measured in the δ13C sequence (δ13C max). Pattern 1 = grazing sheep; pattern 2 = seaweed-eating sheep; pattern 3 = mixed diet with higher contribution of seaweed in winter. All δ13C values measured in modern sheep corrected (+1.5‰) for the fossil fuel effect (figure by M. Balasse).
The phase shift between the δ13C and δ18O sequences clearly discriminates between a monotype diet (terrestrial only/marine only: patterns 1 and 2) and a mixed diet with changing proportions of terrestrial/marine plants (pattern 3). In patterns 1 and 2, the δ13C and δ18O sequences vary in phase, although with a slight shift (296–342°, instead of the 360° expected in-phase signals). By contrast, in pattern 3, the δ13C and δ18O sequences show opposed variations (phase shift from 179–225°). The phase shift between δ13C and δ18O sequences in the Bronze Age sheep from Kemenez (KMZ, ‘22 bis’) is similar to those observed in modern sheep from the same island, suggesting that pre-industrial-age sheep can be compared to these modern references.
Neolithic sheep and cattle
Results from the sequential analysis of the sheep and cattle from Skara Brae are shown in Figures S2–5. The range of intra-tooth variation in δ13C values is presented in Figure 3B, where it is compared to other Neolithic datasets. At Skara Brae, there is a gradient towards higher δ13C values in sheep from phases 0–2. All of the sheep from phase 0 and most from phase 1 demonstrate consistently low δ13C values (≤−11‰), comparable to those measured at Knap of Howar. One sheep from Skara Brae phase 1 and most from phase 2 (6 of 10) show higher δ13C values (>−10‰), compared with some of those measured at Holm of Papa Westray. Overall, the sequences measured at Holm of Papa Westray show higher δ13C values compared with Skara Brae phase 2 (Figure 3B). All cattle teeth from Skara Brae (phases 0–2) show consistently low δ13C values (≤−11‰), comparable to those measured in the cattle from Knap of Howar (Figure 3B).

Figure 3. A) Typical patterns of co-variation of δ13C and δ18O sequences in the sheep and cattle teeth from Skara Brae; B) range of intra-tooth variation of the δ13C values measured in Skara Brae (SKB) sheep and cattle molars, compared to values at Knap of Howar (KH) and Holm of Papa Westray (HPWN). Specimens linked by dashes belong to the same individual (SKB Ovis3 M2–M3; SKB Ovis13 M2–M3). Data for KH and HPWN from Balasse et al. (Reference Balasse, Tresset and Ambrose2006) and Balasse and Tresset (Reference Balasse, Tresset and Ritchie2009) (figure by M. Balasse).
Figure 3A shows typical patterns of co-variation in the δ13C and δ18O sequences in sheep from phases 0–2 and in cattle. Results from the modelling of the Skara Brae, Knap of Howar and Holm of Papa Westray δ13C and δ18O sequences are presented in Table S4. The model cannot be applied to a number of δ13C sequences, either due to the very low amplitude of intra-tooth variation (<1‰), or because of the absence of a suitable sinusoidal pattern. For all the remaining specimens, the phase shift between the δ13C and δ18O sequences is plotted against the δ13Cmax value in Figure 4. At Skara Brae, all the phase 0 sheep are very close in values to the reference grazing sheep (pattern 1). In phase 1, one animal follows pattern 1 and a second falls within the area defined for the winter seaweed contribution (pattern 3). In phase 2, the vast majority of sheep follow pattern 3. The cattle from Skara Brae show very limited amplitude in intra-tooth variation (≤1.3‰, Figure 3B). The phase shift between the δ13C and δ18O sequences places the cattle in pattern 1.

Figure 4. Phase shift between the δ13C and δ18O sequences plotted vs the highest value measured in the δ13C sequence: comparison of the Neolithic sheep from Skara Brae (SKB), Knap of Howar (KH) and Holm of Papa Westray (HPWN) with the reference sheep. Pattern 1 = grazing sheep; pattern 2 = seaweed-eating sheep; pattern 3 = mixed diet with higher contribution of seaweed in winter (figure by M. Balasse).
The modelling of previously published sequences from Holm of Papa Westray (Balasse et al. Reference Balasse, Tresset and Ambrose2006; Balasse & Tresset Reference Balasse, Tresset and Ritchie2009) confirms that all of the sheep from this assemblage follow pattern 3. During phase 2, the winter seaweed contribution to the sheep diet is higher at Holm of Papa Westray than at Skara Brae. None of the archaeological sheep samples following pattern 3 reach the level of seaweed contribution observed in the modern North Ronaldsay sheep.
The results for the sheep from Knap of Howar demonstrate a variety of patterns for seasonal variations in δ13C values, most of which do not follow a sinusoidal trend (Balasse et al. Reference Balasse, Tresset and Ambrose2006; Balasse & Tresset Reference Balasse, Tresset and Ritchie2009). Two δ13C sequences can be modelled. KH Ovis9M3, which shows the lowest δ13Cmax value (−12.2‰) for this assemblage, falls within pattern 1, with a phase shift of 352°. KH Ovis7M3 has a relatively low δ13Cmax value (−10.2‰—also the highest for this assemblage), but is very close to pattern 3 with a phase shift of 154° (Figure 4)—suggesting seaweed consumption during winter. Taking note of this result, three other sheep from Knap of Howar deserve special attention: KH Ovis3, KH Ovis4 and KH Ovis6 all show clear trends towards higher δ13C values in winter (Figure 5). In KH Ovis4 and KH Ovis6, the contribution of seaweed was insufficient to induce a complete reversal of δ13C values in the annual variation cycle, although the trend is clearly initiated.

Figure 5. Variation patterns of δ13C and δ18O sequences in some of the sheep from Knap of Howar (data from Balasse et al. (Reference Balasse, Tresset and Ambrose2006) and Balasse and Tresset (Reference Balasse, Tresset and Ritchie2009)). Contrasting variation of δ13C and δ18O values in some portions of the annual sequence (arrows) (figure by M. Balasse).
Discussion
Re-evaluation of the previous results from Knap of Howar (Balasse et al. Reference Balasse, Tresset and Ambrose2006; Balasse & Tresset Reference Balasse, Tresset and Ritchie2009) highlights a reversal of the expected pattern of co-variation in δ13C and δ18O values in four specimens. This emphasises the contribution of seaweed to the diet of sheep right from the start of the introduction of animal husbandry to Orkney. While seaweed has not been detected in Skara Brae phase 0, which is contemporaneous with Knap of Howar (Bayliss et al. Reference Bayliss, Marshall, Richards and Whittle2017), the small sample size from this period must be acknowledged; the contribution of seaweed to sheep diet evidenced in phase 0 at Knap of Howar is much lower than in later phases. The significant consumption of seaweed during winter is visible in the majority of the sheep from Skara Brae phase 2, which are contemporaneous with the sheep from Holm of Papa Westray (Bayliss et al. Reference Bayliss, Marshall, Richards and Whittle2017). Thus, the chronology of a well-established behaviour evidenced at Holm of Papa Westray is confirmed (Balasse et al. Reference Balasse, Tresset and Ambrose2006; Balasse & Tresset Reference Balasse, Tresset and Ritchie2009). At Skara Brae, this clear pattern is observed even earlier in one specimen from phase 1, dating to the beginning of the third millennium BC. These results confirm the progressive introduction of seaweed to the Neolithic sheep diet, reaffirming the chronological argument, which possibly reflects a progressive adaptation of the digestive physiology of sheep to seaweed.
A higher contribution of seaweed at Holm of Papa Westray
Although the pattern of seasonal seaweed contribution to the winter diet of sheep observed at Skara Brae is similar to that evidenced at Holm of Papa Westray, the relative proportion of seaweed in the diet is almost always higher at the latter site. The same is true for the seaweed-eating sheep from Point of Cott (Westray Island), the remains of which were found in the infilling of the monument's passage. This event corresponds with a separate period of activity to the primary use of the cairn for burial, and dates to between the mid third millennium and the mid second millennium BC (Bayliss et al. Reference Bayliss, Marshall, Richards and Whittle2017). Although this is later than the sheep from Holm of Papa Westray, the Point of Cott sample has yielded a δ13Cmax value of -7.9‰ (Balasse et al. Reference Balasse, Mainland and Richards2009), reflecting a lower contribution of seaweed than at the former site.
These results tend to support the remoteness argument. This is particularly relevant to the small Holm of Papa Westray islet, which may have served as a natural penning area. Here, free-ranging sheep may have had no alternative but to turn to marine resources in times of severe scarcity of grass during winter (Balasse & Tresset Reference Balasse, Tresset and Ritchie2009). Nevertheless, the significant contribution of seaweed to the diet of the sheep found at Skara Brae—a Mainland village context—suggests that the remoteness argument alone cannot explain why sheep turned to seaweed. Furthermore, by this time, seaweed may have developed into a fully established element of the husbandry system, rather than an unintentional consequence of unpenned sheep.
Differences between sheep and cattle
At Skara Brae, sheep and cattle were taken to different pasture grounds during the winter: sheep were brought to the shore, whereas cattle were kept on terrestrial pastures. Previous studies have also demonstrated the terrestrial grazing of Neolithic cattle in Orkney, as well as the Outer Hebrides (Jones & Mulville Reference Jones and Mulville2016; Schulting et al. Reference Schulting, Vaiglova, Crozier and Reimer2017). Cattle feeding on seaweed is attested on the islands of Brittany (Arzel Reference Arzel1987) and in Iceland, where it is used as a substitute for hay due to a lack or inaccessibility of grasslands (Hallson Reference Hallson, De Virville and Feldman1964). In Iceland, seaweed is observed to have been used to a lesser extent for cattle than for sheep, although the reasons for this are unclear (Hallson Reference Hallson, De Virville and Feldman1964). Schulting et al. (Reference Schulting, Vaiglova, Crozier and Reimer2017) suggest that during the Neolithic in Orkney, the greater value of cattle in relation to sheep may explain why they were given priority access to grassland resources and hay supplies.
Conclusion
Re-evaluation of the results from stable isotope analysis of faunal remains from the mid to late fourth-millennium cal BC Knap of Howar site indicates that sheep may have included moderate amounts of seaweed in their winter diet from the time of their first arrival in Orkney. Seaweed consumption is clearly detected in one sheep from phase 1 (early third millennium cal BC) at Skara Brae and found in the majority of specimens from phase 2 (c. 2750–2500 cal BC), and therefore appears to be an established practice by this time. It is not possible to establish whether Neolithic sheep turned to seaweed of their own accord or were brought to the shore by herders. In any case, these results confirm the chronology of the gradual introduction of seaweed into the Neolithic sheep diet in Orkney, which may have been the result of changes in sheep behaviour or physiology. When compared with Skara Brae, the considerably higher relative contribution of seaweed to the winter sheep diet at Holm of Papa Westray also confirms the validity of the remoteness argument, at least as far as the quantity of seaweed consumption is concerned.
The introduction of seaweed to the diet of Orkney's Neolithic sheep, however, was not the start of an unbroken tradition that endures to the present. Winter seaweed consumption by sheep has been detected through isotope analysis for a single animal from the Iron Age site of Mine Howe (Figure 1; Balasse et al. Reference Balasse, Mainland and Richards2009), although microwear analysis on the same assemblage does not confirm seaweed-eating as a common pattern in sheep at the site (Mainland et al. Reference Mainland, Towers, Ewens, Davis, Montgomery, Batey, Card and Downes2016). Further, the colonisation of Orkney by Viking settlers from Norway brought new animal husbandry practices, and the settlers may have imported their own livestock (Mainland et al. Reference Mainland, Towers, Ewens, Davis, Montgomery, Batey, Card and Downes2016). Currently, both stable isotope (Balasse et al. Reference Balasse, Mainland and Richards2009) and tooth microwear analyses (Mainland et al. Reference Mainland, Towers, Ewens, Davis, Montgomery, Batey, Card and Downes2016) suggest a year-round terrestrial diet for sheep at the Viking/Norse settlement at Earl's Bu (late 800s to mid 1100s AD). There is, therefore, little evidence for unbroken continuity in the history of Orkney husbandry between the Neolithic and the present.
Finally, none of the evidence for seaweed-eating sheep on Orkney from the Neolithic to the Viking period shows any significant contribution of seaweed to diet outside the winter period. Moreover, none of them reached the level of seaweed contribution found in modern sheep on North Ronaldsay. Here, the exclusive reliance on seaweed is a consequence of sheep being confined to the shore to prevent grazing on cultivated grounds. The associated physiological adaptations, therefore, may have arisen only following human intervention in the nineteenth century. In terms of husbandry practices and animal behaviour, however, the practice certainly has ancient roots, dating to the first arrival of farmers and sheep in Orkney.
Assessing whether this pattern was unique to Orkney or more widespread across prehistoric North-western Europe requires additional research. Today, seaweed-eating sheep are known in Shetland (latitude 60° north) and the Faroe Islands (62° north), but the archaeology of this practice has not yet been investigated in these places. At a number of sites of Neolithic to Iron Age date in the Scottish Western isles (57.5° north), bone collagen (as opposed to enamel bioapatite) stable isotope analysis has failed to demonstrate seaweed consumption by sheep (Jones & Mulville Reference Jones and Mulville2016). But, bone metabolism in sheep varies with the season—showing a peak in bone formation in summer and increased bone resorption in autumn and winter (Arens et al. Reference Arens, Sigrist, Alini, Schawalder, Schneider and Egermann2007)—winter seaweed consumption could be more difficult to identify in bone collagen. Future investigations using seasonal resolution (i.e. sequential analysis in teeth) may reveal a different picture in these locations. Exploitation of marine resources reflects not only a zootechnical innovation by Neolithic farmers to cope with local environmental constraints, but also a revolution in thinking, by exploiting the littoral margins to sustain terrestrial domestic livestock. This innovation is emblematic of a series of other changes, adopted over the course of the European Neolithic, in terms of farming practices and animal and plant physiology (Marinova & Valamoti Reference Marinova, Valamoti, Chevalier, Marinova and Peña-Chocarro2014; Orton et al. Reference Orton, Gaastra and Vander Linden2016; Ivanova et al. Reference Ivanova, De Cupere, Ethier and Marinova2018). These changes illustrate how Neolithic farmers faced the challenges of introducing domesticates and establishing sustainable agropastoral systems in the most remote areas of Europe.
Acknowledgements
This work was supported by the ‘SIANHE’ ERC Starting Grant project (GA 202881). Stable isotope analyses were performed at the Service de Spectrométrie de Masse Isotopique du MNHN, Paris. We thank Allison Sheridan and the National Museums Scotland for permission to sample the Skara Brae faunal remains, and Soizic and David Cuisnier from the Ferme insulaire de Kemenez for providing access to their property and sheep carcasses. Anneke Janzen contributed to the laboratory work as part of her PhD training. English editing was done by Louise Byrne.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.15184/aqy.2019.95






