INTRODUCTION
The Mediterranean countries have a continental Mediterranean climate with: (a) variable rainfall, 56–88%, which falls in the cold period, extending from October to March, and (b) extended summer drought periods (Chennafi et al., Reference Chennafi, Aidaoui, Bouzerzour and Saci2006; Nefzaoui et al., Reference Nefzaoui, Ketata, El Mourid, Young and Silvern2012). So, the water availability as well as the inappropriate grazing management cause limited pastoral resources that are the major obstacle to livestock biological and economic sustainability, particularly to extensive livestock production systems. Thus, the choice of suitable forage species establishment during the winter months represents one of the main strategies to meet the animals’ requirements through low cost sources of energy and nutrients. For this purpose, the use of grasses and legumes in the Mediterranean basin has been proposed (Abbas and Porqueddu, Reference Abbas and Porqueddu2012; Mefti et al., Reference Mefti, Bouzerzour, Chebouti, Kebour and Abdelguerfi2012). Legumes have higher nutritive value than grass species, so growing mixtures of grasses and legumes can improve forage quality compared to grass monocultures (Sleugh et al., Reference Sleugh, Moore, George and Brummer2000; Zemenchik et al., Reference Zemenchik, Albrecht and Shaver2002). More specifically, the protein and carbohydrate rates of feed originated from mixtures are balanced and have higher feeding value compared to feed from pure stands (Mariotti et al., Reference Mariotti, Masoni, Ercoli and Arduini2009; Tuna and Orak, Reference Tuna and Orak2007). Compared with pure stands, grass-legume mixtures provide higher yields with better seasonal distribution and decreased unsown presence. Traditionally, pasture seed mixtures available in southern Europe consisted of a small number of legume species and low rates of annual grasses. Nowadays it is common to find complex mixtures including up to 20 annual self-regenerating legumes in combination with perennial grasses. Encouraging results have been obtained by Porqueddu and Maltoni (Reference Porqueddu and Maltoni2005) evaluating four species mixtures.
Taking into account that there is lack of knowledge with regard to determine the optimum species composition of grass/legume mixtures in temperate grassland areas, this study was designed to provide more knowledge on this subject. More specifically, the aim of the present study was to examine the production efficiency as well as various aspects of fodder composition using various plant species of ‘high yield combining ability’ in different mixtures under temperate conditions.
MATERIALS AND METHODS
The experimental sward was established in the experimental fields of the Department of Agronomy at the Agricultural University of Athens (37° 58′ 43″N and 23° 42′ 57″E). The soil of the field could be categorized as sandy clay loam (SCL) type, was of modest fertility and no fertilizers were applied during the course of the trial.
The experimental field was divided in 54 (2 m × 3 m) sub-plots and six treatments (comprising of species seed mixtures) were applied, in a completely randomized design. Sowing was performed in October 2009, after tillaging, using the following plant species mixtures (PSM): (1) Festuca arundinacea, Lolium perenne (FL); (2) Pisum sativum, Hordeum vulgare (PH); (3) Festuca arundinacea, Dactylis glomerata, Trifolium repens (FDT); (4) Vicia sativa, Hordeum vulgare (VH); (5) Festuca arundinacea, Lolium perenne, Dactylis glomerata, Trifolium repens (FLDT); and (6) Festuca arundinacea, Lolium perenne, Trifolium repens (FLT). The species were represented by the following breeders lines: Festuca arundinacea (cv FUEGO), Lolium perenne (cv ROSALIN), Trifolium repens (cv HAIFA R1), Dactylis glomerata (cv MONTE), Pisum sativum (cv CHEYENNE-CANION), Vicia sativa (cv ΤΕMBI) and Hordeum vulgare (cv ΑTHINAIS). Furthermore, three different species ratios within each PSM were used and each combination was replicated three times (Table 1). The selection of those ratios was based on earlier studies carried out in the same field during the assessment of several mixture ratios in the productivity of aforementioned plant species (Economou et al., Reference Fales, Fritz, Barnes, Nelson, Moore and Collins2010). After seeding, the field was irrigated by sprinklers with 25 mm of water for the successful establishment of PSM. Then the field remained rainfed for rest of the growing period. The sward was allowed to establish for 12 weeks before the first harvest (87 days from seeding) and the plots were subjected to a further four cuts at 121, 148, 170 and 196 days from seeding for all the PSM. Two additional cuts were done for the FL, FLDT and FLT PSM at days 216 and 250 from seeding. The cuts were performed with a forage harvester with reciprocate blades set at the height of 5 cm above ground.
Table 1. Three different species ratios within each plant species mixtures (PSM).
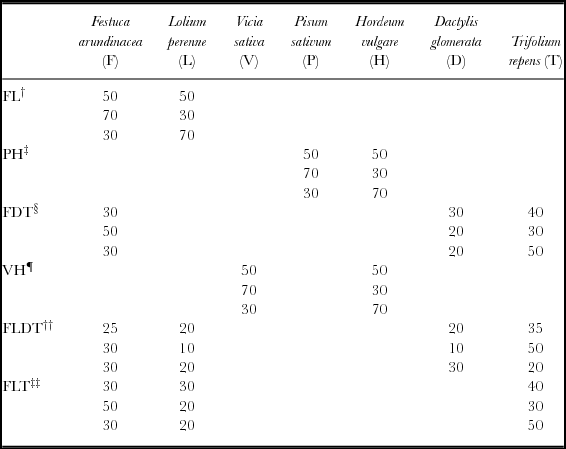
†FL: Festuca arundinacea, Lolium perenne.
‡PH: Pisum sativum, Hordeum vulgare.
§FDT: Festuca arundinacea, Dactylis glomerata, Trifolium repens.
¶VH: Vicia sativa, Hordeum vulgare.
††FLDT: Festuca arundinacea, Lolium perenne, Dactylis glomerata, Trifolium repens.
‡‡FLT: Festuca arundinace, Lolium perenne, Trifolium repens.
Herbage harvested from each plot was weighed as fresh and then sampled after thorough mixing. One sample was dried overnight at 90 °C in a fan-assisted oven, for herbage dry matter (DM) determination and a second was dried for 48 h at 55 °C and ground through a 1 mm screen for subsequent proximate composition analysis. A third sample was immediately frozen and stored for subsequent fatty acids (FA) analysis.
Chemical composition analyses
The chemical composition of the PSM was estimated using near-infrared reflectance spectroscopy (NIRS). The PSM collected samples were dried for 48h at 60 °C and ground through a 1 mm screen before proximate analysis. The samples were scanned on a NIRSystems 5000 scanning monochromator (FOSS GmbH, Rellingen, Germany), and software (ISI-version) for data collection and manipulation was supplied by Infrasoft International® (ISI, Port Matilda, PA, USA). Calibration and validation subset samples were analysed for the crude protein (CP), NDF, ADF and ether extracts (EE) according to Goering and Van Soest (Reference Goering and Van Soest1970).
Fatty acids analysis
Fresh herbage from each plot (~150 g) was sampled immediately, and pooled over the three replicates of each PSM × species ratio combination, then closed in plastic bags with dry ice before transfer to a deep freezer (–80 °C) where they were stored until FA analysis. The samples of the 1st cut were not analysed for FA profile because they were considered damaged due to a freezer. The frozen herbage sample was shred into 5–10 mm pieces with a paper cutter and thoroughly mixed. Two grams of each final sample were used for FA analysis according to the method of O’ Fallon et al. (Reference O’ Fallon, Busboom, Nelson and Gaskins2007) by gas chromatography. For the determination of FA concentration an Agilent 6890 N gas chromatograph equipped with an HP-88 capillary column (80 m × 0.25 mm i.d. with 0.20 μm film thickness; Agilent) was used. A flame ionization detector temperature was set at 260 °C and the chromatographic analysis involved a temperature programmed run starting at 120 °C and held for 1 min. This followed by two step up ramp, one of 1.25 °C min–1 to 230 °C, and another of 10 °C min–1 to 240 °C, and held for 3 min. Helium was used as the carrier gas with a linear velocity set at 30 cm s–1.
STATISTICAL ANALYSIS
Application of the quantile–quartile plots showed that all variables showed reasonably the normal distribution. Since this was a purely nested design, appropriate data analysis for DM was the application of a mixed model with fixed and random effects. The fixed effects part of the model included the effects of PSM, the effect of ratio (R) nested within PSM as well as the effect of cut (C) within PSM by R. The random effects part of the model included the effects of replicate (rep) nested within the cut by ratio by PSM and the error term (eijkl). Thus the following model was used for analysis:
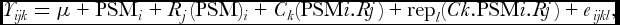 \begin{equation*}
Y_{\it ijk} = \mu + {\rm PSM}_i + R_j ({\rm PSM})_i + C_k ({\rm PSM}i{\rm .}Rj) + {\rm rep}_l (Ck.{\rm PSM}i.Rj) + e_{\it ijkl} ,
\end{equation*}
\begin{equation*}
Y_{\it ijk} = \mu + {\rm PSM}_i + R_j ({\rm PSM})_i + C_k ({\rm PSM}i{\rm .}Rj) + {\rm rep}_l (Ck.{\rm PSM}i.Rj) + e_{\it ijkl} ,
\end{equation*}where Υijkl is the dependent variable, μ the overall mean, PSMi the effect of PSM (i = 1,. . .,6), Rj the effect of ratios (j = 1,. . .,3), Ck the effect of cut (k = 1,. . .,7), replication (l = 1,. . .,3) and eijkl the residual error. The random effects (rep and e) were assumed to follow the normal distribution with mean equal to zero and variances s 2r and s 2e, respectively. Results of this analysis are presented as least squares means. Because of multiple comparisons a Bonferroni adjustment of the p-values was applied here. EE, NDF, ADF and CP were analysed by MANOVA fitting PSM, the ratio within PSM and the cut within PSM as the fixed effects. The respective analysis for FA included the effects of PSM and the ratio within PSM. All the above analyses were carried out by SAS (SAS v9.0, 2004).
RESULTS
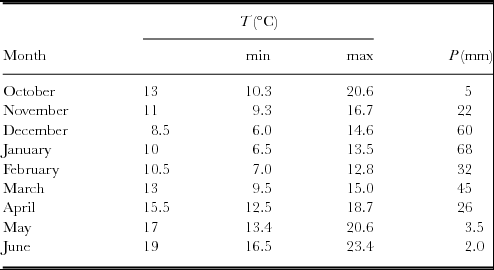
Climatic data: temperature and precipitation throughout the experimental period are presented in Table 2. As reasonably expected, highest precipitation and lowest temperatures were observed during the winter months.
Table 2. The mean temperature (T), and precipitation (P) throughout the experimental period.
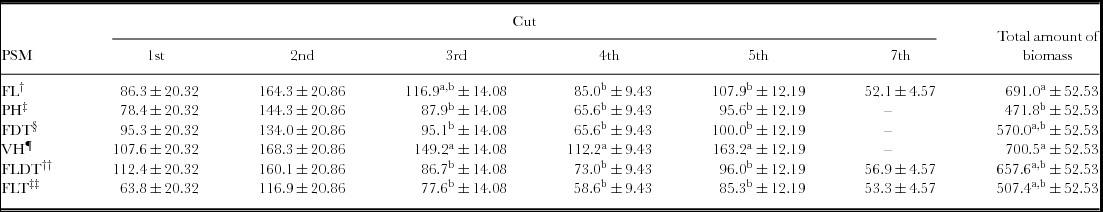
The DM of the different PSMs across the cuts is presented in Table 3. In all PSMs highest values in dry weight were attained at the 2nd cut, with values ranging from 116.94 g m–2 to 168.25 g m–2 (Table 3). The VH showed the highest value in DM across the first five cuts with values being significant at the 3rd, 4th and 5th cut (Table 3). With regard to DM, it should be pointed out that the FL, FLDT and FLT PSM gave two more cuts, with the FLDT showing higher values when contrasted to the rest of the PSMs. In addition, in the total amount of biomass produced throughout the experimental period, no differences were observed between the VH and FLDT PSM (Table 3).
Table 3. Mean DM (g m–2) of the three ratios of each plant species mixture (PSM) among the different cuts.

Note: Footnotes†, ‡, §, ¶, ††, ‡‡ as in Table 1. Sixth LSM of the 6th cut are not presented because of the infinite likelihood. Means with different superscripts (a and b) in each column (among different plant species mixtures) at the same cut differ significantly (p ≤ 0.05).
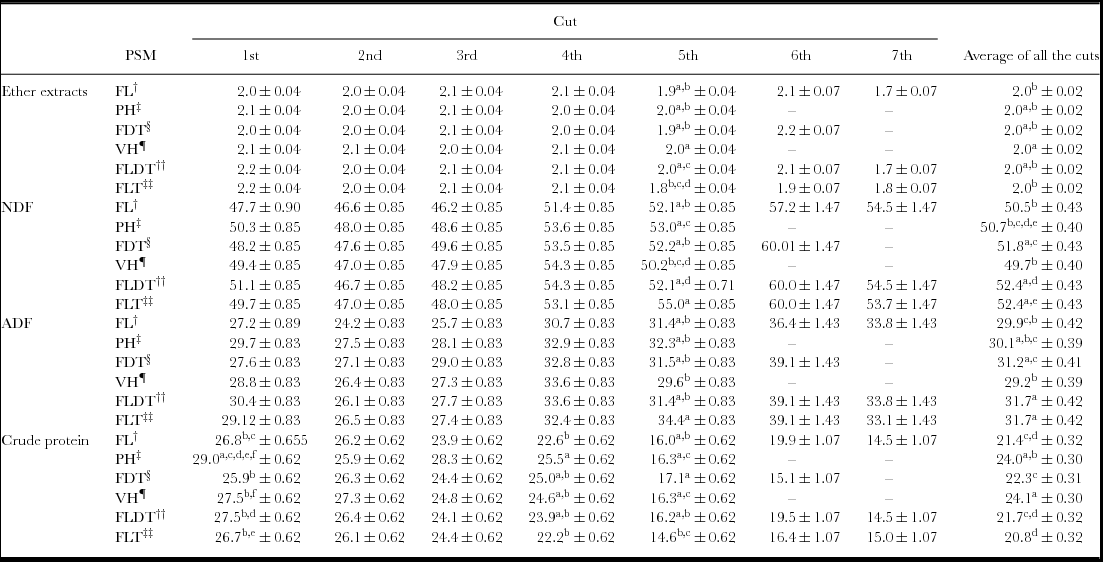
The chemical composition per cut and in total is presented in Table 4. Because the ash content varied across samples, the EE, NDF, ADF and CP are expressed as per cent organic matter (% OM). In all the PSMs, the EE and the CP content decreased while the NDF and ADF increased from cuts 1 to 5. This was also the case for the PSMs that gave a total of seven cuts. The highest CP content across the cuts was observed in the VH and PH (Table 4). Significant higher average values across the cuts in ADF and NDF were found for the FLDT when compared to VH. In addition, no difference was observed between VH and FLDT regarding the EE content (Table 4).
Table 4. Mean chemical composition (ether extracts, NDF, ADF and crude protein in % OM) of the three ratios of each plant species mixture (PSM) at the different cuts.
Note: Footnotes†, ‡, §, ¶, ††, ‡‡ as in Table 1. Means with different superscripts (a, b, c, d, e and f) for each parameter in each column (among different plant species mixtures) at the same cut differ significantly (p ≤ 0.05).
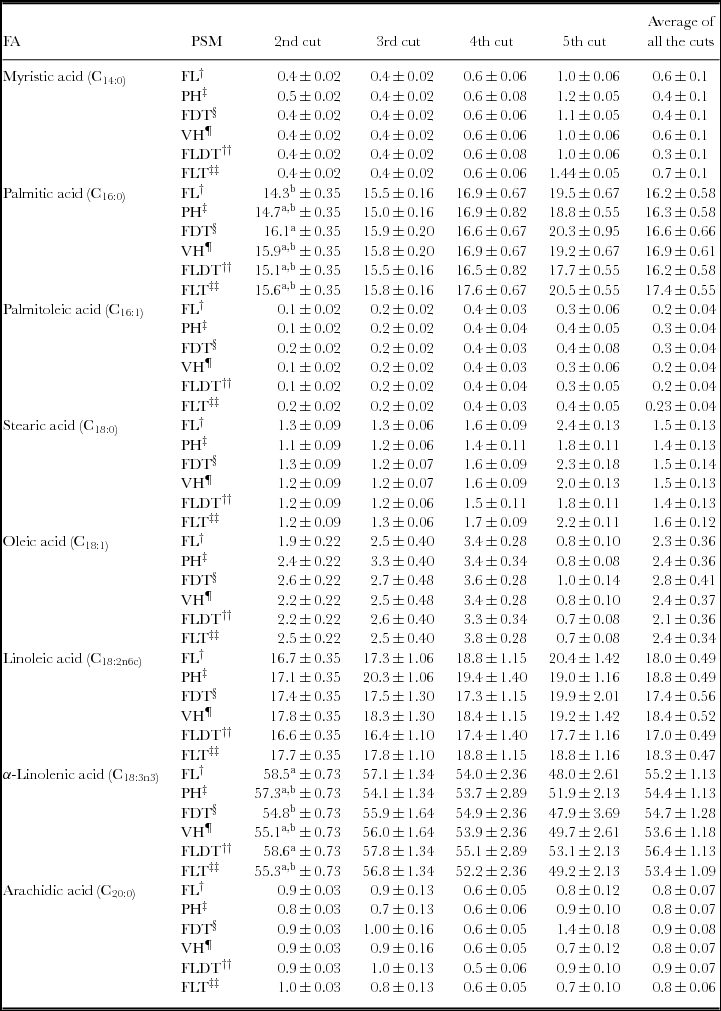
Table 5 presents the FA profile of the PSM per cut and on average. In all the PSMs, and irrespective from the cut, α-linolenic, linoleic and palmitic acid showed the highest concentration. In all PSMs, the concentrations of myristic, palmitic, stearic, oleic and linoleic acid increased significantly from cuts 2 to 5, while the opposite was observed for the α-linolenic acid content. In addition, in the 2nd cut, the concentration of α-linolenic acid was significantly higher in the FL and FLDT when compared to the rest ones. The same trend was observed for the FLDT with regard to the α-linolenic acid content per cut and in total, although without no significant differences.
Table 5. The mean of the main fatty acids (FA; % of total fatty acids) of the three ratios of each plant species mixture (PSM) at the different cuts.*
*The FA profile of the 1st cut is not presented because the samples were damaged due to a freezer failure.
Note: Footnotes†, ‡, §, ¶, ††, ‡‡ as in Table 1. Means with different superscripts (a and b) for each FA in each column (among different plant species mixtures) at the same cut differ significantly (p ≤ 0.05).
DISCUSSION
In the present study all the PSMs showed the highest productivity during the 2nd cut (25 February 2010), a period that coincides with greatest vegetative growth of the cool season plant species under the Mediterranean conditions (Tables 2 and 3). Furthermore, the climatic conditions (temperature and precipitation recorded during the negative phase of the plants: December 2009 to February 2010) proved a favourable factor for their growth (Table 2). From all the PSMs studied here, VH has proved its superiority in terms of DM in the 3rd, 4th and 5th cuts, while as its total biomass produced throughout the experimental period, it did not differ between the FL, FDT, FLDT and FLT significantly (Table 3). However, it should be emphasized that the FL, FLDT and FLT gave two more cuts during early summer months, a feature that is very important for ruminants’ feeding. In the latter cases, highest yields attained may be attributable to the rapid re-growth of Lolium perenne favoured by the temperature increase and air humidity, a finding that is in accordance with other studies (Barthram and Grant, Reference Barthram and Grant1994).
On an annual basis, the produced DM biomass obtained from these leys was at least two-fold the maximum quantity produced by natural grasslands of Greece (Zervas, Reference Zervas1998). During months October to March the nutrition of small ruminants, almost all over Greece, typically falls below requirements, particularly after lambing/kidding and during lactation (Zervas et al., Reference Zervas, Fegeros and Papadopoulos1996) due to lack of natural vegetation. Therefore, the production of valuable grass from leys and its supply to lactating small ruminants is of great importance for the production costs.
In all PSMs, CP content gradually decreased by cut (Table 4). This trend may be due to the increase of the temperature from January (1st cut) to May (5th cut) and onwards (Hatfield et al., Reference Hatfield, Boote, Fay, Hahn, Izaurralde, Kimball, Mader, Morgan, Ort, Polley, Thomson and Wolfe2008). Significantly higher values in ADF and NDF in the FLDT compared to VH were found throughout. In addition, a steady increase for NDF and ADF in all varieties was observed from the 1st to the 5th cuts (Table 4). This trend may also be due to increased temperature from January to May and onwards (Table 2). It is well documented that temperature can alter the carbohydrate status of metabolic sinks by speeding/slowing individual metabolic reactions, by changing rates of active transport across membranes, by changing the concentration of different enzymes (through modification of gene expression) and by changing enzyme activity (Fales and Fritz, Reference Goering and Van Soest2009). Deinum and Dirven (Reference Deinum and Dirven1972, Reference Deinum and Dirven1975) associated the increases in temperature from 24 to between 28 and 33 °C with greater stem weight and increased CF of both C3 and C4 forage grasses. Although the negative effect of increasing temperature is not consistent across all species and conditions, Wilson et al. (Reference Wilson, Deinum and Engels1991) demonstrated this response for C3 grasses, and Henderson and Robinson (Reference Henderson and Robinson1982) for C4 grasses. Data for forage legumes are limited. Wilson and Minson (Reference Wilson and Minson1984) reported lower digestibility of the tropical legume sirato (Macroptillium atropurpureum (DC.) Urb.) with increasing growth temperature. This decrease was associated with an increase in cell wall and lignin concentration, mainly in stem fraction. It has been suggested that temperature effects on the nutritive value of tropical legumes are often not as great as those on grasses where lignin accumulation in the cell wall was observed throughout the plant (Newman et al., Reference Newman, Sollenberger, Boote, Allen, Vu and Hall2005).
Regardless the cut, in all the PSMs, α-linolenic, linoleic and palmitic acids were the main FAs (Table 5). These results agree with those of Dewhurst et al. (Reference Dierking, Kallenbach and Roberts2001), Boufaїed et al. (Reference Boufaïed, Chouinard, Tremblay, Petit, Michaud and Belanger2003) and Clapham et al. (Reference Clapham, Foster, Neel and Fedders2005) who have demonstrated that the concentrations of α-linolenic, linoleic and palmitic acids range from 45 to 70%, 10 to 25% and 15 to 25%, respectively, of the total FAs in many forage species. In the 2nd cut, α-linolenic acid concentration was significantly higher in the FL and FLDT when compared to the rest ones. The same holds for the FLDT, in the other cuts also, with no statistical significance. This may be attributed to the participation of Lolium perenne and Festuca arundinacea in this particular PSM. Boufaїed et al. (Reference Boufaïed, Chouinard, Tremblay, Petit, Michaud and Belanger2003) observed that grasses contained more α-linolenic acid than legumes. In addition, Lough and Anderson (Reference Lough and Anderson1973) found that α-linolenic acid was the major component of esterified FA (simple and complex lipid) in mixed pasture grasses including ryegrass (such as Lolium multiflorum Lam) and meadow fescue (Festuca pratensis huds.). High concentration of α-linolenic acid (31.87%) has also been observed in Lolium perenne by Dierking et al. (Reference Economou, Theodorou, Tsiplakou, Kotoulas and Hadjigeorgiou2010).
Studies examining forages have highlighted that the FA profile of plants is modulated by environmental factors. Such factors include light (Dewhurst and King, Reference Dewhurst, Scollan, Youell, Tweed and Humphreys1998), and the season of the year (Dewhurst et al., Reference Dierking, Kallenbach and Roberts2001). Given that both factors were the same across the experiment, at least per cut, the observed significant differences in α-linolenic acid between the PSMs at the 2nd cut may be only due to the different composition of the mixtures. However, significant reductions of the α-linolenic acid content, within PSMs, could be due to different temperatures from February (2nd cut) to May (5th cut) as variation in α-linolenic acid proportions between periods seem to be related to differences in temperature (Table 2). Indeed, the proportion of α-linolenic acid has been reported to be higher during periods with lower temperatures (April and May) than during periods with higher temperatures (July and August; Witkowska et al., Reference Witkowska, Wever, Gort and Elgersma2008). Hawke (Reference Hawke, Butler and Bailey1973) and Vight et al. (Reference Vight, Maresca and Harwood1998) have reported also higher α-linolenic acid proportions at lower temperatures.
Forages provide a low-cost approach in comparison with diet supplementation strategies, such as oils and starch, which are designed to improve milk FA profile relative to their impacts on human's health. Manipulation of milk fat content and its FA profile through breeding and nutrition strategies have been important targets for the dairy industry in many parts of the world. Animal production on rangelands, as in other grazing systems, depends on the quality as well as the quantity of forage. Key quality parameters for forages include ADF, NDF, CP content and the FA profile. Taking into account the main characteristics of the PSMs, from quantity (DM) and quality (chemical composition and FA profile) point of view, it may be concluded that highest DM can be obtained at the 2nd cut of all the PSMs. This is very important finding since in February the grass of natural grassland in Mediterranean basin is very limited. This herbage of high quality and of low production cost can be used very efficiently by small ruminants for milk production. Additionally, from all the PSMs that yielded seven cuts, the FLDT gave higher DM in early summer when there is almost no available herbage for small ruminants in Mediterranean basin. In this particular PSM, the α-linolenic acid content was higher throughout, with apparent positive effect on ruminant's products quality such as the FA profile. Increasing the supply of ω-3 polyunsaturated fatty acids in animal's diet is one of the most important ways of improving the beneficial FA in ruminant's milk. It has been reported that pasture plays a key role in improving the health benefits from milk and dairy products of ruminants (Tsiplakou et al., Reference Tsiplakou, Mountzouris and Zervas2006). Taking into account all the parameters examined (DM, NDF, ADF, EE, CP and α-linolenic acid) the FLDT seems to be the superior one, throughout the cuts.
CONCLUSIONS
Results of the present study have demonstrated that the establishment of leys by high-yielding PSMs can provide sufficient quantities of grass of high quality for small ruminants’ feeding. Maximum dry weight of biomass was reached in February when there is no available grass from natural grasslands. Furthermore, the total annual biomass was at least double when compared to that of natural grasslands. Due to its high NDF, ADF and relatively high CP content, the chemical composition of the FLDT grass can meet the requirements of lactating animals while its FA profile can significantly contribute to milk fat enrichment in polyunsaturated fatty acids and conjugated linoleic acids for the consumer's health aspects.