



1. Introduction
The genus Protoichthyosaurus was established by Appleby (Reference Appleby1979), who recognized a unique forefin morphology. Unrecognized composite specimens and a rare co-ossification in Ichthyosaurus forefins that mimics the Protoichthyosaurus morphology led to a misunderstanding of the range of individual variation within Ichthyosaurus (Lomax et al. Reference Lomax, Massare and Mistry2017). Thus the difference between the two genera was not considered sufficient to distinguish the taxa. For these reasons, and because of a lack of information on the skull, Protoichthyosaurus was synonymized with Ichthyosaurus (Maisch & Hungerbühler, Reference Maisch and Hungerbühler1997), and this was accepted in subsequent publications (Maisch & Matzke, Reference Maisch and Matzke2000; McGowan & Motani, Reference McGowan and Motani2003). Lomax et al. (Reference Lomax, Massare and Mistry2017), however, argued that the forefin morphology was distinctly different because the forefin of Protoichthyosaurus has only three primary digits (II, III and IV), whereas Ichthyosaurus has four (Motani, Reference Motani1999a). The loss of digit V, the number of elements in the distal carpal row, and the broad contact between the intermedium and distal carpal three are among the features that distinguish the forefin of Protoichthyosaurus from that of Ichthyosaurus (Lomax et al. Reference Lomax, Massare and Mistry2017).
Many similarities exist between the two genera, so their previous synonymy is not surprising. Protoichthyosaurus and Ichthyosaurus are the only Lower Jurassic ichthyosaurs to display anterior digital bifurcations in the forefin. This results in a distinctive forefin that is much wider than that of contemporaneous genera. The two genera also have similar morphologies of the coracoid, scapula, humerus and femur (Lomax et al. Reference Lomax, Massare and Mistry2017). Recent work, however, has recognized a combination of skull characteristics that can distinguish the two genera (Lomax et al. Reference Lomax, Massare and Mistry2017, Reference Lomax, Porro and Larkin2019; Lomax & Massare, Reference Lomax and Massare2018). This work examines differences in morphology of the skull roof, as seen on previously undescribed specimens, herein assigned to Protoichthyosaurus prostaxalis (primarily LEICT G142.1991). We also discuss the variation in the exposure of the maxilla, which, in isolation, might have been interpreted as an unusual character and thus perhaps evidence of a new taxon.
Institutional abbreviations: BRLSI ─ Bath Literary and Scientific Institution, UK; BU ─ Lapworth Museum, University of Birmingham, UK; CAMSM ─ Sedgwick Museum, University of Cambridge, UK; GPIT ─ Institut und Museum für Geologie und Paläontologie, Universität Tübingen, Germany; LDUCZ ─ The Grant Museum of Zoology, University College London, UK; LEICT ─ Leicester Arts and Museum Service, New Walk Museum and Art Gallery, UK; NHMUK ─ The Natural History Museum, London, UK; NMING ─ National Museum of Ireland, Dublin; NMW ─ National Museum of Wales, Cardiff, UK; OUMNH ─ Oxford University Museum of Natural History, UK; SMNS ─ Staatliches Museum für Naturkunde, Stuttgart, Germany; UOD ─ University of Derby, UK; WARMS ─ Warwickshire Museum, Warwick, UK.
2. Materials
Four specimens described herein are previously unrecognized specimens of Protoichthyosaurus. LEICT G142.1991 and LEICT G738.1889 are partial skulls. The former is a historic specimen which has become dissociated from its original accession information. The bone colour and preservation, however, suggest that the specimen is from Barrow-upon-Soar, Leicestershire. A search of LEICT records of unlocated ichthyosaur skull specimens indicates four potential matches. All of these are from Barrow-upon-Soar and were collected by curator Montagu Browne, also known as Alexander Montagu Browne Anderson (McCann, Reference McCann1981), through the quarry owners Messrs J Ellis and Sons in either 1889 or 1891. LEICT G738.1889 is recorded as being collected in October 1889 by Browne/Anderson from Barrow-upon-Soar. WARMS G15646 and LDUCZ #X424 are proximal portions of forefins. The recognition of these four additional specimens increases the known number of specimens of Protoichthyosaurus to 32.
The most informative specimen and the focus of this study, LEICT G142.1991, is a moderately sized skull preserved in three dimensions, but missing much of the rostrum anterior to the external naris (preserved jaw length ∼38 cm). The dorsal side has well-exposed sutures that are filled with dark pyritous matrix, which in places are further revealed by superficial surface damage. The ventral side of the skull is preserved but damaged posteriorly. Parts of the pectoral girdle are also preserved, as are the proximal ends of the humeri. LEICT G738.1889 is also a partial skull in three dimensions that is missing most of the rostrum. The skull roof is exposed from the middle of the nasals to the temporal fenestra, but it is poorly preserved. The bones around the posterior and ventral orbit margin, bones of the occipital region and the entire mandible are missing. Acid preparation has produced a pitted surface on many of the bones. The matrix of each of the two skulls contains a number of poorly preserved ammonites which can be referred to Caloceras sp.
The forefin specimens cannot be assigned to a species. WARMS G15646 is an incomplete, articulated forefin that preserves the distal end of the humerus and the proximal portion of the fin. It was figured by Smith & Radley (Reference Smith and Radley2007, fig. 4A) as a specimen of Ichthyosaurus. LDUCZ #X424 similarly comprises an incomplete, articulated forefin that preserves the distal end of a humerus and the proximal portion of the fin. A second, more distal portion of a fin is stored alongside the specimen, but it might not belong with the proximal portion of the fin.
3. Systematic Palaeontology
ICHTHYOSAURIA de Blainville, Reference de Blainville1835
Ichthyosauridae Bonaparte, Reference Bonaparte1841
Protoichthyosaurus Appleby, Reference Appleby1979
Diagnosis. From emended diagnoses in Lomax et al. (Reference Lomax, Massare and Mistry2017) and Lomax & Massare (Reference Lomax and Massare2018): Distinguished from other ichthyosaurian genera by autapomorphies of the forefin: distal carpal 3 contacts the ulnare; proximal element of bifurcation in metacarpal row nearly separating distal carpal 2 from distal carpal 3; and intermedium separated from contact with distal carpal 4 by distal carpal 3. The combination of three elements in the distal carpal row and a bifurcation of distal carpal 2 producing four elements in the metacarpal row is unique for the genus.
Additionally characterized by a unique combination of characters: pineal foramen bordered posteriorly by the parietals (shared with Temnodontosaurus, Leptonectes, Stenopterygius and others); posteriorly wide nasals (shared with Ichthyosaurus and S. aaleniensis); elongated internasal foramen often present (see discussion in Lomax & Massare, Reference Lomax and Massare2018); prefrontal anterior process dorsoventrally large and separates most, if not all, of dorsal process of lacrimal from anterior orbit margin (shared with I. somersetensis, I. larkini); triradiate lacrimal; large external naris, usually somewhat triangular (much higher posteriorly than anteriorly) (similar to Temnodontosaurus, I. conybeari); strongly asymmetric maxilla with long, slender anterior process (shared with I. conybeari); prominent grooves on tooth root extend to base of crown (shared with Temnodontosaurus, I. somersetensis); coracoid with wide anterior and posterior notches (shared with Ichthyosaurus); scapula with long shaft, and slightly expanded anterior end, but without prominent acromion process (shared with Ichthyosaurus, Temnodontosaurus, Suevoleviathan); humerus nearly equal in width distally and proximally, with slight constriction in the shaft (shared with Ichthyosaurus, Macgowania); three primary digits (II, III, IV) in forefin, with digit V absent (shared with Toretocnemus, Temnodontosaurus and others, but not Ichthyosaurus); presence of three elements in distal carpal row of forefin; only distal carpal 3 in broad contact with the intermedium; bifurcations of digit II result in five digits in forefin, although fin must be preserved to the fourth phalangeal row to verify number (Ichthyosaurus has bifurcations in digit II or III; no other taxa have an anterior digital bifurcation); forefin phalanges closely packed, mostly rectangular, but rounded at distal end of fin; and femur head robust, but with narrow shaft and distal end wider than proximal.
Additional referred specimens. WARMS G15646 and LDUCZ #X424, both proximal portions of forefins.
Locality and horizon. WARMS G15646 is from the Saltford Shale Member of the Blue Lias Formation (Lower Jurassic, Hettangian; Liassicus to Angulata Chronozones) of Southam Cement Works Quarry, Warwickshire, England. No information is available for LDUCZ #X424.
Remarks. Lomax et al. (Reference Lomax, Massare and Mistry2017) included another character in the diagnosis, namely that the maxilla is excluded from the margin of the external naris by the subnarial process of the premaxilla and anterior process of the lacrimal. The specimens described herein illustrate that this character is variable (Section 6 below). In fact, on some specimens one side has the maxilla excluded from the narial margin whereas the other side has the maxilla making up a portion of the ventral margin. Whether it is the result of variation or preservation is not always clear.
Protoichthyosaurus prostaxalis Appleby, Reference Appleby1979
Diagnosis. From emended diagnosis in Lomax et al. (Reference Lomax, Massare and Mistry2017): Distinguished from P. applebyi by the following features: large, dorsoventrally tall triangular maxilla with a long anterior process that extends beyond nasals in lateral view; dorsoventrally short, but anteroposteriorly wide, almost rectangular, postorbital (anteroposteriorly narrow but dorsoventrally tall (crescentic) in P. applebyi); lacrimal dorsal process longer than anterior process (roughly equal in P. applebyi); postorbital makes up half or less of posterior orbit margin (makes up more than half in P. applebyi); rectangular squamosal with triangular process extending ventrally from the posteroventral edge (unknown in P. applebyi); robust humerus with small dorsal process that is centrally located and does not extend far down the shaft (plate-like dorsal process forms a narrow ridge in P. applebyi); and hindfin with three elements in distal tarsal (third) row (unknown in P. applebyi).
Additional referred specimens. LEICT G142.1991, a partial skull in three dimensions, missing the anterior rostrum, with pectoral girdle and associated axial elements. LEICT G738.1889, a poorly preserved, partial skull in three dimensions (see Section 2 above).
Locality and horizon. Barrow-upon-Soar, Leicestershire (see Section 2 above). Ammonites in the matrix place the specimens in the Johnstoni subzone, Planorbis Zone, lower Hettangian, Lower Jurassic. Given that the specimens are from Barrow-upon-Soar, they probably originated from either the ‘Rummels’ limestone or from a nodule within the underlying ‘Rummels Calf’ shale (Martin et al. Reference Martin, Frey and Riess1986).
4. Description of LEICT G142.1991
LEICT G142.1991 is assigned to Protoichthyosaurus because the posterior edge of the pineal is within the parietals; the large anteroventral process of the prefrontal separates the dorsal process of the lacrimal from the orbit margin; the maxilla is very asymmetric, with a long anterior process; and prominent grooves in the tooth root extend to the base of the crown. The specimen is assigned to P. prostaxalis because of a dorsoventrally short, anteroposteriorly broad, almost rectangular postorbital (postorbital is dorsoventrally long, narrow crescent in P. applebyi), a posterior jugal ramus that makes up almost half of the posterior orbit margin (posterior jugal ramus makes up at least a quarter in P. applebyi), and a triangular, dorsoventrally tall maxilla (maxilla is dorsoventrally short and bar-like in P. applebyi).
4.a. Skull
The skull is preserved best in left lateral view, although it is slightly dorsoventrally crushed and laterally skewed. The dorsal view, however, provides new information on Protoichthyosaurus prostaxalis (Fig. 1). The maxilla is dorsoventrally tall and somewhat triangular, with the anterior process extending well beyond the external naris, as in P. prostaxalis (e.g. BU 5323, BRLSI M3555) and I. conybeari, but it is not as narrow as in the latter (Massare & Lomax, Reference Massare and Lomax2016; Lomax et al. Reference Lomax, Massare and Mistry2017, Reference Lomax, Porro and Larkin2019). The posterior extent of the maxilla cannot be determined exactly, because of damage, but the maxilla is definitely asymmetric (Fig. 2). It is dorsoventrally tall and contacts the ventral margin of the external naris on both sides of the skull. The contact occurs because the anterior process of the lacrimal and subnarial process of the premaxilla do not overlap the maxilla and meet along the narial boundary, as typically occurs in Protoichthyosaurus (see Section 5 below).

Fig. 1. LEICT G142.1991, Protoichthyosaurus prostaxalis (a) skull in dorsal view and (b) interpretive drawing of the sutures of the skull roof. Scale bar equals 10 cm. Abbreviations: brk, broken; en, external naris; f, frontal; if, internasal foramen; j, jugal; l, lacrimal; mx, maxilla; n, nasal; o, orbit; p, pineal; pa, parietal; pmx, premaxilla; po, postorbital; prf, prefrontal; ptf, postfrontal; q, quadrate; qj, quadratojugal; sq, squamosal; st, supratemporal.
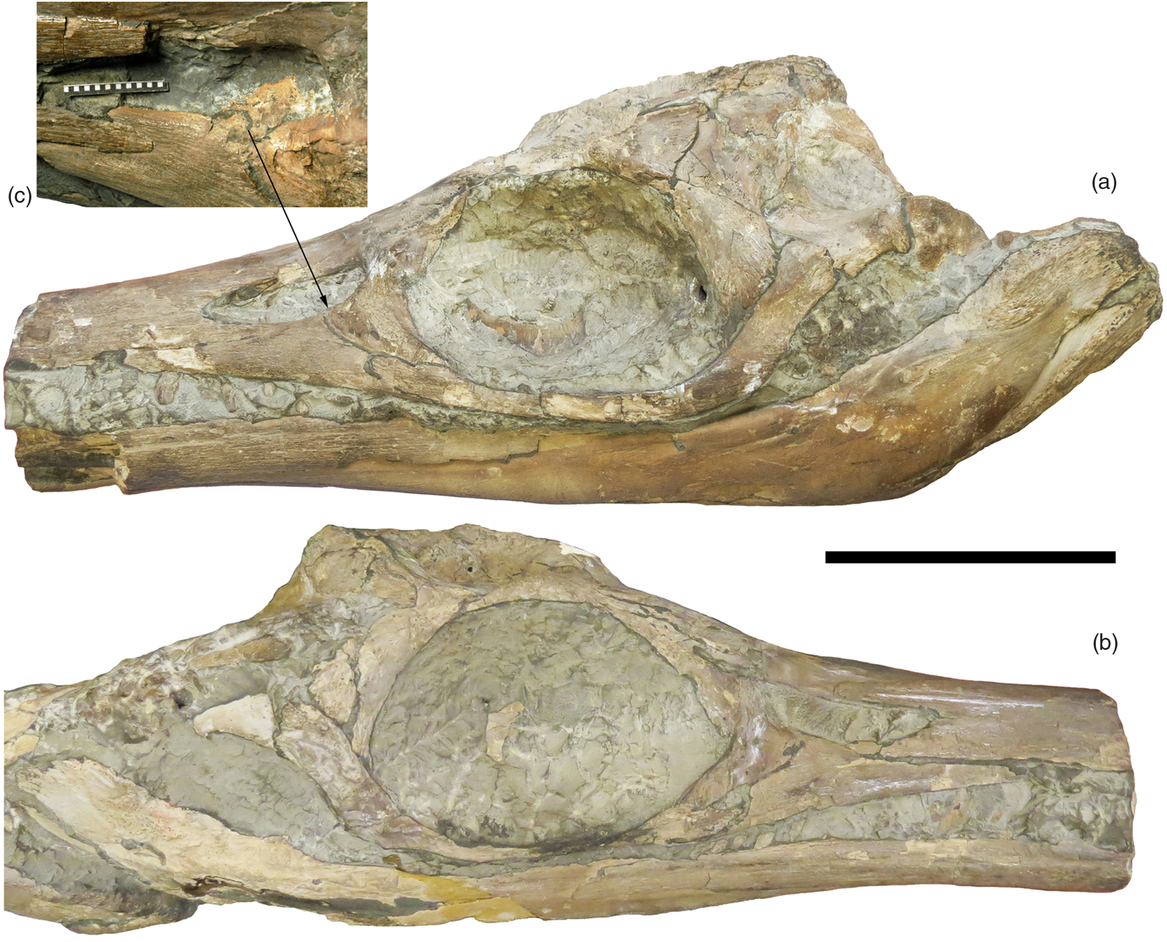
Fig. 2. LEICT G142.1991, skull of Protoichthyosaurus prostaxalis (a) in left lateral view, and (b) in right lateral view. (c) Closer view of the left external naris, after additional preparation, showing the flange that extends from the lacrimal into the naris. Also note the ‘hook’ on the nasal that makes up part of the posterior margin of the external naris. Scale bar equals 10 cm.
The external naris is large and anteroposteriorly elongated due to crushing, which is particularly apparent on the left side. Typically on undeformed skulls of Protoichthyosaurus, the large naris has a triangular shape (Lomax et al. Reference Lomax, Massare and Mistry2017; Lomax & Massare, Reference Lomax and Massare2018). The supranarial process of the premaxilla extends almost halfway across the margin of the external naris on the right side. But on the left side, it is separated from the naris margin by a sliver of the nasal for at least half of its length. On the left side, the premaxilla supranarial process is clearly broken. Most of the dorsal margin of the external naris is formed by the nasal, which has a ventral projection that makes up the posterodorsal margin of the naris (Fig. 2a, c). This can be seen clearly in at least two other specimens of Protoichthyosaurus prostaxalis(BU 5323, GPIT/RE/09728, previously GPIT 1796/1 (see Lomax & Massare, Reference Lomax and Massare2018, fig. 3C)), but the posterior margin of the external naris is frequently damaged, even on otherwise well-preserved skulls (e.g. WARMS G347, NMW 2012.23.G1). Protoichthyosaurus applebyi might share this feature, but neither known specimen is sufficiently well preserved to be certain (Lomax et al. Reference Lomax, Massare and Mistry2017; Lomax & Massare, Reference Lomax and Massare2018). We have not been able to positively identify this feature on Ichthyosaurus. Furthermore, crushing often distorts the contacts between the lacrimal and nasal at the narial margin. The nasal is widest posteriorly, making up almost the entire skull width dorsal to the mid-orbital region, as in Ichthyosaurus. It narrows abruptly anterior to the position of the external naris, being covered by the premaxilla. An elongated internasal foramen is present, along with a long depression, as seen in most specimens of Protoichthyosaurus prostaxalis (e.g. BU 5323; see remarks in Lomax & Massare, Reference Lomax and Massare2018) and rarely on a few specimens of Ichthyosaurus (e.g. OUMNH J13799; Lomax & Massare, Reference Lomax and Massare2018).
The lacrimal dorsal process is noticeably longer than the anterior process and makes up about half of the posterior margin and less than half of the ventral margin of the external naris. What appears to be a broad flange from the anterior process of the left lacrimal extends posteromedially into the narial opening (Fig. 2c). On the right side, the external naris is more filled with matrix, and a small part of what might be the flange is exposed posteriorly. The posteroventral process of the lacrimal is longer than both the dorsal and anterior processes. Several small foramina are preserved towards the ventral edge of the lacrimal.
On the left side, the anterior process of the jugal is wide and rounded, and extends slightly beyond the anterior edge of the orbit. On the right side, the jugal dorsal ramus makes up about half of the posterior orbit margin. On the left side, it appears that the dorsal ramus has a gentle bend and makes up less than half of the orbit margin, but this is due to crushing. The postorbital is poorly preserved on the right side and on the left side is dorsoventrally compressed. It is roughly rectangular and has a long contact with the jugal dorsal ramus, typical of P. prostaxalis. It is not narrow relative to its dorsoventral length, as occurs on P. applebyi (Lomax & Massare, Reference Lomax and Massare2018).
In lateral view, the prefrontal is dorsoventrally long anteriorly and narrows posteriorly, best observed on the right side (Fig. 2). It separates the dorsal process of the lacrimal from the anterior margin of the orbit, as in Protoichthyosaurus, Ichthyosaurus somersetensis and I. larkini (Lomax & Massare, Reference Lomax and Massare2017, Reference Lomax and Massare2018). It bears a noticeable longitudinal trough parallel to the long axis of the bone on the left side of the skull, less so on the right, but this might be due to deformation. We have not seen a similar trough on other specimens of Protoichthyosaurus. Posteriorly, the prefrontal contacts the postfrontal, which makes up most of the dorsal margin of the orbit.
The margins of the upper temporal fenestra are made up of the postfrontal (anterior), parietals (medial and minor contribution to the anterior) and supratemporal (lateral and posterior). The anterior end of the postfrontal is anteroposteriorly broad and extensive; and contacts the parietal posteriorly, the frontal medially, and prefrontal and nasal anteriorly (Fig. 1). The postfrontal-nasal suture interdigitates laterally. In dorsal view, the frontal is anteroposteriorly longer than wide, and broader anteriorly than posteriorly. Both parietals are complete, and their articulation forms a relatively low sagittal crest, seen best in posterior view. Anterolaterally, the parietal contacts the postfrontal. A posterior shelf (parietal ridge) is present at the posterior end of the parietals, ventral to the sagittal crest, and leads to the articulation with the supratemporals posterolaterally, although both supratemporals are damaged posteriorly.
The pineal foramen is situated between the frontals and the parietals. The posterior edge of the frontals makes up the anterior and lateral edges of the pineal foramen. Only the posterior edge and a minor part of the lateral edge of the pineal foramen is bordered by the anteriorly dividing parietals (Fig. 1). The foramen is a somewhat teardrop shape, narrowest where it is between the parietals. GPIT/RE/09728 has a teardrop-shaped foramen; however, other specimens of P. prostaxalis have an oval foramen (e.g. LEICT G720.1899, BU5323). The minor difference is likely the result of preservation.
Portions of the squamosal, quadratojugal and quadrate are preserved but are too incomplete to provide morphological information. Some elements of the braincase are also preserved, including the supraoccipital, but they are either poorly preserved or buried in matrix.
4.b. Lower jaw and dentition
The posterior portion of the jaw is best preserved on the left side. At the posterior end, the angular makes up less than half of the lower jaw in lateral view, but anteriorly its exposure decreases abruptly, and its lateral exposure ends posterior to the posterior jugal ramus. The surangular makes up most of the posterior portion of the jaw. It extends anteriorly at least as far as the middle of the external naris in lateral view and extends ventrally for c. 40 mm anterior to the naris. More posteriorly, the surangular meets the dentary at a position approximately level with the middle of the orbit.
In ventral view, the jaws are preserved to just posterior to the symphysis. Anteriorly, the angular has only a narrow exposure, overlapped laterally and medially by the surangular and the fairly robust splenial respectively. A rod-like right hyoid element is preserved in what is probably ventral view. The left articular is also preserved, but slightly damaged and buried. A few poorly preserved teeth, mainly tooth roots, are present. On one maxillary tooth, longitudinal grooves in the roots extend to the base of the crown and are continuous with the longitudinal striations of the crown, as is typical of Protoichthyosaurus.
4.c. Postcranial skeleton
Portions of both coracoids are preserved in articulation with each other and with the right clavicle, scapula and interclavicle (Fig. 3). The coracoid has wide anterior and posterior notches, producing a fairly long anteroposterior constriction between the glenoid/scapular facets and the medial portion of the coracoid, as is typical of Protoichthyosaurus (Lomax et al. Reference Lomax, Massare and Mistry2017). The interclavicle is the usual T-shape, although most of the left lateral process is missing. The medial process is somewhat shorter than the lateral process and widens distally. The scapula is only slightly longer than the mediolateral length of the coracoid. Its straight, narrow shaft widens at the anterior end but lacks an acromion process. The proximal portions of both humeri are also preserved, associated with the coracoids, but neither element is complete enough to provide any information.

Fig. 3. LEICT G142.1991, Protoichthyosaurus prostaxalis, incomplete pectoral girdle. Scale bar equals 10 cm. Abbreviations: cl, clavicle; cr, coracoid; h, humerus; ic, interclavicle; sc scapula.
5. Description of LEICT G738.1889
This specimen is much less complete than the specimen described above. It is a fragmentary, small skull, referred to Protoichthyosaurus based on the anteroventral extent of the prefrontal, the posteriorly wide nasals, and the position of the pineal foramen. It is assigned to P. prostaxalis based on the triangular, dorsoventrally tall maxilla. The preserved skull roof extends from the narial region, posterior to the posterior end of the temporal fenestra. The nasals are widest at their posterior end, and they contact the postfrontals and frontals. An elongated internasal foramen is in a depression where the medial edges of the nasals meet. The pineal foramen seems to be positioned between the frontals and the parietals, but the bone surface is damaged and sutures are unclear. In lateral view, the ventral region of the skull is badly damaged and only the anterodorsal portion of the orbit is preserved. The maxilla is dorsoventrally tall, preserved best on the right side, where it is separated from the external naris by the narrow ends of the subnarial process of the premaxilla and anterior process of the lacrimal (similar to BU 5323, P. prostaxalis, Lomax et al. Reference Lomax, Massare and Mistry2017). On the left side, the maxilla makes up a portion of the ventral edge of the external naris, similar to LEICT G142.1991, although this may be because of damage to the premaxilla and lacrimal. The prefrontal is poorly preserved, but on the left side it seems to extend between the lacrimal dorsal process and the anterior orbit margin. In ventral view, some portions of the palate and braincase are preserved, but the bones are largely damaged. However, the shape of the basisphenoid, although crushed, is similar to that of another specimen of P. prostaxalis (Lomax et al. Reference Lomax, Porro and Larkin2019, fig. 6), in having robust, oblong basipterygoid processes.
6. Discussion
6.a. Variation in exposure of the maxilla in Protoichthyosaurus
In lateral view of a complete Protoichthyosaurus skull, the maxilla forms a roughly triangular element. The extent of the posterior process varies, but the anterior process always extends beyond the anterior edge of the external naris. The dorsoventral height of the maxilla is greatest ventral to the middle or mid-posterior end of the naris. The height differs in the two species, with a much dorsoventrally taller maxilla present in P. prostaxalis than in P. applebyi (Lomax et al. Reference Lomax, Massare and Mistry2017; Lomax & Massare, Reference Lomax and Massare2018). The shape of the maxilla in lateral view has been used as a diagnostic feature of species of Ichthyosaurus and Protoichthyosaurus (Lomax & Massare, Reference Lomax and Massare2017, fig. 4C, D; Lomax et al. Reference Lomax, Massare and Mistry2017, fig. 4). However, the actual shape of the maxilla is dorsoventrally taller than what is typically exposed in lateral view because the dorsal portion of the maxilla is overlapped by the subnarial process of the premaxilla (McGowan, Reference McGowan1973). The lacrimal also overlaps the maxilla (e.g. BU 5323, NHMUK R36958). Thus the extent and preservation of the premaxilla and lacrimal will affect the exposure of the maxilla, and its height and dorsal extent in lateral view.
LEICT G142.1991 is unusual because the maxilla makes up a portion of the ventral margin of the external naris on both sides of the skull (Fig. 4a, b), unlike in any previously known specimen of Protoichthyosaurus (Lomax et al. Reference Lomax, Massare and Mistry2017; Lomax & Massare, Reference Lomax and Massare2018). This feature might suggest that the specimen is a new taxon, but we argue otherwise because the morphology of the rest of the skull is consistent with an assignment to P. prostaxalis (Section 4 above). Typically, P. prostaxalis has a dorsoventrally tall maxilla in lateral view (Lomax et al. Reference Lomax, Massare and Mistry2017). The anterior process of the lacrimal and subnarial process of the premaxilla separate it from the margin of the external naris (Fig. 4c, d). On some specimens, however, the separation is a thin sliver of bone (Fig. 4e, f; e.g. LEICT G738.1889, left side), such that the maxilla almost reaches the margin of the naris. Thus the extent of the separation of the maxilla from the external naris is variable within the species. Although LEICT G142.1991 displays an unusual morphology, we contend that it is just a more extreme variation, where the premaxilla and lacrimal are shorter than is typical, resulting in the maxilla exposure at the margin of the external naris. Stenopterygius quadriscissus shows a comparable range of morphologies, in that the maxilla usually makes up part of the ventral margin of the external naris, but in some specimens the maxilla is excluded from the external naris by the subnarial process of the premaxilla and anterior process of the lacrimal (Maisch & Matzke, Reference Maisch and Matzke2000, p. 9).

Fig. 4. Lateral exposure of the maxilla on specimens of Protoichthyosaurus prostaxalis. LEICT G142.1991 (a) left side and (b) right side. Note that the maxilla makes up a substantial part of the ventral margin of the external naris. Left sides of (c) NMW 2012.23G.1 and (d) GPIT GPIT/RE/09728, showing a wide separation between the dorsal margin of the maxilla and the ventral margin of the external naris. Right sides of (e) BU 5323 and (f) BRLSI M3555, a paratype of the species, show the thin premaxilla and lacrimal separating the maxilla from the external naris margin, a more typical morphology. On UOD D1, the maxilla makes up part of the ventral margin on (g) the left side, but (h) the right side has the typical morphology. Part of the premaxilla subnarial process has been worn away, and some of the lacrimal anterior process may also have been worn. Scale bars equal 3 cm. Abbreviations: la, lacrimal; mx, maxilla; pmx, premaxilla.
In four isolated skulls of P. prostaxalis, the left and right sides show different morphologies. In UOD D1, the right maxilla is separated from the external naris by the lacrimal and premaxilla, but the left maxilla contacts the external naris (Fig. 4g, h). Possibly the subnarial process of the premaxilla and anterior process of the lacrimal have been worn away, but it is debatable. LEICT G738.1889, NMING:F8756 and NHMUK R36958 also show a typical morphology on one side of the skull, and the maxilla contacting the external naris on the other. The difference on these skulls, however, is more likely due to wear or breakage. Nonetheless, this demonstrates the importance of a larger sample size to capture the variation among specimens of the same species.
6.b. Skull roof configuration of Protoichthyosaurus
Although hundreds of specimens of Lower Jurassic ichthyosaurs with skulls are in museum collections, relatively few show a well-preserved skull roof. Even three-dimensionally preserved skulls often lack clear sutures. In general, difficulties arise in identifying sutures and key features of the skull roof because skulls are often fractured from crushing, partially disarticulated, or flattened at an oblique angle that deforms the skull roof. Because the bones of the skull roof overlap, the underlying bones could potentially be exposed by damage, weathering, or even acid preparation, leading to misinterpretations. Moreover, the range of variation within a species is rarely known.
The skull roof is better preserved in LEICT G142.1991 than in any other specimen of Protoichthyosaurus, and allows comparison with two common Lower Jurassic taxa, Ichthyosaurus and Stenopterygius. The most detailed study of the skull roof of Ichthyosaurus and Stenopterygius was Motani (Reference Motani2005), who figured and cited specific specimens to support his reconstructions. Although his reconstruction of Stenopterygius has been supported subsequently (Maxwell et al. Reference Maxwell, Fernandez and Schoch2012), his reconstruction of Ichthyosaurus is debatable (Section 7.b below). Three characters discussed below are unambiguous on well-preserved skulls and can distinguish these three genera: (1) the position of the pineal foramen relative to the frontal and parietal, (2) the presence or absence of a dorsal exposure of the prefrontal that contacts the frontal, and (3) the position of the maximum width of the nasals. Protoichthyosaurus differs from Ichthyosaurus in the first feature, and differs from Stenopterygius in the second and third features.
6.b.1. Position of the pineal foramen relative to the frontal and parietal
In Protoichthyosaurus, the frontals do not enclose the pineal foramen; instead, the parietals make up the posterior edge of the pineal, as in Stenopterygius (Motani, Reference Motani2005, fig. 2; Maxwell et al. Reference Maxwell, Fernandez and Schoch2012, fig. 3B). The position of the pineal is clearly shown in LEICT G142.1991 (Fig. 5a) and confirms the position beyond doubt, but it is also evident in LEICT G738.1899 and LEICT G729.1889 (referred to P. prostaxalis by Lomax et al. Reference Lomax, Massare and Mistry2017). Three other specimens of Protoichthyosaurus prostaxalis (NHMUK R36958, BU 5323, GPIT/RE/09728) also suggest that the parietals make up the posterior margin of the pineal foramen, but its position is not as clear. In Ichthyosaurus, the frontals narrow posteriorly and wrap around the pineal foramen, completely enclosing it at the posterior edge of the frontals (Fig. 5b, c), a feature that is thus far unique to the genus (Massare & Lomax, Reference Massare and Lomax2017). Because the posterior end of the frontal can be a narrow bone, it is often damaged or missing, even on otherwise well-preserved specimens (e.g. CAMSM J35186). Motani (Reference Motani2005, fig. 3A, B, C) suggested that NHMUK OR2090 might be a new genus. However our assessment is that it is probably Ichthyosaurus somersetensis because of the anterior extent of the prefrontal, the shapes of the maxilla, lacrimal and postorbital, and the pineal foramen completely enclosed by the frontal (Lomax & Massare, Reference Lomax and Massare2017).

Fig. 5. Position of the pineal foramen, anterior towards the top. (a) Protoichthyosaurus prostaxalis, LEICT G142.1991, shows the frontals making up the anterior and lateral margins of the pineal and parietals making up the posterior margin. (b) Ichthyosaurus somersetensis, OUMNH J13799, and (c) Ichthyosaurus?somersetensis, NHMUK OR2090, show the pineal completely enclosed by, and at the posterior end of, the frontals. Scale bars equal 2 cm.
The position of the pineal foramen is correctly shown in the reconstructions of Ichthyosaurus in McGowan & Motani (Reference McGowan and Motani2003, fig. 69) and Motani (Reference Motani2005, fig. 1D), although we question other aspects of the reconstructions and the identification of the latter (Section 6.b below). However, reconstructions in Motani (Reference Motani1999b, fig. 3E) and Ji et al. (Reference Ji, Jiang, Motani, Rieppel, Hao and Sun2016, fig. 4G), both of which were based on Romer (Reference Romer1956, fig. 85B), incorrectly showed the pineal position with the posterior margin contacting the parietals in Ichthyosaurus. The reconstruction of Ichthyosaurus communis in Maisch & Matzke (Reference Maisch and Matzke2000, fig. 11) similarly showed the pineal foramen incorrectly positioned between the parietals and the frontals, but it was based on Sollas (Reference Sollas1916, text-fig. 5). Sollas (Reference Sollas1916) described a serially sectioned skull in which the frontals made up the anterior and lateral margin of the pineal, and the parietal made up the posterior margin. Although the specimen label indicated it was Ichthyosaurus communis, Sollas doubted that identification and dubbed it ‘I. communis var. a’, a species he thought was closely related to I. communis (Sollas, Reference Sollas1916, p. 63). It is highly probable that Sollas’ specimen was a Protoichthyosaurus because he described the pineal as clearly bordered by the parietals posteriorly. Additionally, the dorsoventrally robust prefrontal excludes the dorsal process of the lacrimal from the orbit margin (Sollas, fig. 1a). However, an unequivocal identification is not possible from the figure. Maisch & Matzke (Reference Maisch and Matzke2000) thus might have inadvertently figured Protoichthyosaurus rather than Ichthyosaurus communis.
6.b.2. Contact between frontal and prefrontal
A second important feature of Protoichthyosaurus involves the postfrontal, nasal and frontal contacts on the skull roof. Motani (Reference Motani2005, fig. 2) argued that the prefrontal extends medially onto the dorsal skull and contacts the frontal in Stenopterygius, separating the postfrontal from the frontal and usually from the nasal as well. He documented this ‘medial exposure of the prefrontal’ in six specimens of Stenopterygius. This morphology has also been reported more recently in S. aaleniensis, where the prefrontal has an isolated medial exposure (an ‘island’, using the term in Motani, Reference Motani2005) between the frontal, prefrontal and nasal (Maxwell et al. Reference Maxwell, Fernandez and Schoch2012, fig. 3B). In LEICT G142.1991, the interdigitating sutures clearly show that the postfrontal directly contacts the nasal and the frontal, with neither an exposure of the prefrontal separating them nor a contact between the frontal and prefrontal (Fig. 1). Coincidentally, Motani (Reference Motani2005, fig. 3D, E, F) illustrated NHMUK R1164, pointing out that the specimen lacks the prefrontal exposure between the postfrontal and frontal. The specimen has since been referred to Protoichthyosaurus applebyi (Lomax & Massare, Reference Lomax and Massare2018). Thus the contact between the postfrontal and frontal (i.e. the prefrontal does not separate the postfrontal and frontal) occurs on both species of Protoichthyosaurus and can be considered a characteristic of the genus that distinguishes it from Stenopterygius.
Motani (Reference Motani2005, fig. 1) also argued that Ichthyosaurus has a similar medial exposure of the prefrontal, separating the postfrontal from the frontal. Unlike most previous reconstructions, Motani (Reference Motani2005) cited specific specimens upon which his reconstruction was based, making it possible to evaluate his interpretations. He stated that the medial exposure of the prefrontal occurs unequivocally in eight specimens of Ichthyosaurus (NHMUK R8177, NHMUK R6697, NHMUK R10021, NHMUK R15943, NHMUK OR49203, NHMUK OR39492, NHMUK OR43006, OUMNH J13799). Only the latter two specimens are associated with diagnostic postcranial elements, and of those, only OUMNH J13799 has a dorsally preserved skull. In that specimen, we presume that Motani (Reference Motani2005) interpreted the small bone fragments anterolateral to the frontal on each side as the prefrontal (Fig. 5b). We interpret the bones as fragments of the frontal, which widens anteriorly. The texture of the main body of the frontal and the bone fragments is similar, and bone striations are continuous from one bone to the other on both sides. The other specimen with diagnostic postcranial features, NHMUK OR43006, the holotype of I. breviceps, shows the skull mainly in lateral view although a portion of the skull roof is displaced and exposed (pers. obs.; McGowan, Reference McGowan1974, fig. 7). Because of the deformation and skull orientation, we are hesitant to agree with Motani’s (Reference Motani2005, fig. 1E) interpretation of the morphology.
Of the other specimens, we can verify that the prefrontal is exposed medially and separates the postfrontal from the frontal only on NHMUK R15943. The specimen is an isolated skull roof that has been acid prepared. Although the pineal foramen seems to be enclosed by the frontals, the contact between the nasals and the frontals, specifically the narrow, anterior extension of the frontals between the nasals, is unusual for Ichthyosaurus. More significantly, the exposure of the prefrontal is at a markedly lower level of the skull roof than either the postfrontal or the nasal. This suggests the possibility that the growth of the nasal and/or postfrontal was not complete, for whatever reason, thus exposing the prefrontal below. Alternately, acid preparation could have removed some of the overlying bone. Additionally, the generic identification of the specimen is somewhat equivocal as it is not associated with any diagnostic postcranial material. It was collected in a loose block on the shore between Blue Anchor and Watchet, Somerset (T Smithson, pers. comm., 2019). Thus it could be as old as early Rhaetian (Late Triassic), pre-dating the Late Triassic extinction, raising the possibility that it is not Ichthyosaurus. So although we agree with Motani’s (Reference Motani2005, fig. 1D) interpretation of the morphology, this specimen is problematic.
NHMUK OR39492 is an isolated skull that is on display behind glass, mounted in left lateral view. The orientation and deformation make it difficult to interpret the skull roof. The prefrontal appears to extend onto the skull roof, but it seems to contact the postfrontal rather than the frontal. Thus, the skull roof morphology is debatable.
The four remaining isolated skulls identified as Ichthyosaurus by Motani (NHMUK R8177, NHMUK R6697, and NHMUK R10021, NHMUK OR49203) were described as having ‘exceptionally clear sutures’ (Motani, Reference Motani2005, p. 338). The first three specimens have been acid prepared and might have the same kind of issue with bone removal as NHMUK R15943. McGowan (1973) described NHMUK R8177 and NHMUK R6697 in detail in his monograph on the skull of Ichthyosaurus. NHMUK R8177 is a completely disarticulated skull. Although Motani (Reference Motani2005, fig. 1A, B) articulated the elements such that the prefrontal contacts the frontal, McGowan’s reconstruction of the skull roof showed the postfrontal contacting the frontal, presumably overlapping the prefrontal (McGowan, 1973, fig. 35, although fig. 37, a lateral view, is equivocal). The contacts in the articulated skull are not clear in the plates (McGowan, 1973, pls 7, 8), although Motani (Reference Motani2005, p. 339) indicated that he could see the prefrontal–frontal contact in another photograph of the specimen. NHMUK R6697 is a partially articulated, posterior portion of the skull, also described by McGowan (1973). The specimen also includes vertebrae, ribs, a partial fin and pectoral girdle elements. The skull does not appear to preserve the area of interest here, although it was mentioned, but not discussed, in Motani (Reference Motani2005). NHMUK R10021 is a highly fractured skull in dorsolateral view. Several skull elements have been labelled directly on the specimen. Of particular relevance, what is labelled as ‘Frontal’ on the specimen was identified as part of the prefrontal by Motani (Reference Motani2005), but it is actually part of the nasal. The surface striations on the bone fragments are continuous with those of the adjacent nasal. Thus the nasals extend posteriorly to about even with the middle of the orbit. Furthermore, Motani (Reference Motani2005) identified the element labelled ‘R. Pre-Frontal’ as part of the postfrontal. However, this element is indeed part of the prefrontal. Not only is the bone surface texture continuous with the rest of the prefrontal, the bone contacts the lacrimal and is located in the typical position of a prefrontal, too anteriorly to be a postfrontal. Our interpretation of this poorly fragmented skull is that the prefrontal clearly does not contact the frontal. As for NHMUK OR49203, the skull is dorsoventrally crushed and the portion of the skull roof of interest here is damaged and poorly preserved. We could not identify a prefrontal contacting the frontal. As with specimen NHMUK R6697, Motani (Reference Motani2005) mentions it, but does not discuss it. The main issue with all of these specimens, except for NHMUK R6697, is that they are isolated skulls.
A different approach is to examine the skulls of some well-preserved skeletons that are irrefutably Ichthyosaurus. Specifically, we looked for an exposure of the prefrontal immediately lateral to the frontal. The best specimen, OUMNH J13799, has already been discussed above (Fig. 5b). Three other specimens are preserved with a skull in dorsolateral view. In ANSP 15766, the holotype of I. somersetensis, a contact between the frontal and the postfrontal, not the prefrontal, is evident, even though the skull roof is deformed. Although damaged, TTNCM 8373, another I. somersetensis, also shows a frontal–postfrontal contact, shown most clearly on the right side of the skull roof. NHMUK OR2013* is similar to OUMNH J13799 in that a small bone fragment is adjacent to the nasal and postfrontal on each side of the skull. We again interpret them as fragments of the frontals. Motani (Reference Motani2005, p. 340) conceded that medial exposure of the prefrontal was debatable on NHMUK OR2013*. A fourth specimen, CAMSM J35186, is preserved in dorsal aspect, but the skull is badly fragmented. Again, as in the other specimens, the anterior portion of the frontal is broken into several pieces, and there is no medial exposure of the prefrontal.
At most, two of the skulls that Motani (Reference Motani2005, fig. 1A–D) described have a medial exposure of the prefrontal that contacts the frontal, but the evidence is equivocal, as discussed above. Skeletal specimens cited above more strongly support the skull roof reconstruction of Ichthyosaurus in Figure 6. But a medial exposure of the prefrontal might occasionally occur (e.g. maybe NHMUK R8177, see above; Motani, Reference Motani2005, fig. 1A, B). The bones of the skull roof overlap, and in particular, the postfrontal and nasal overlap the prefrontal (Sollas, Reference Sollas1916; McGowan, 1973). So it is conceivable that a suture might not close ‘properly’ as the skull grows, and the underlying bone, the prefrontal, could be exposed. Therefore, if this does occur, it would be the exception rather than the norm for Ichthyosaurus. Figure 6 illustrates our interpretation of the skull roof of Ichthyosaurus, in which the postfrontal contacts the frontal, without a dorsal exposure of the prefrontal contacting the frontal, and the pineal is enclosed entirely within the frontals. A medial exposure of the prefrontal is not evident on any specimen of Protoichthyosaurus, an observation that agrees with Motani (Reference Motani2005, fig. 3F, P. applebyi).

Fig. 6. Reconstruction of the skull roof of Ichthyosaurus, modified from Motani (Reference Motani1999b fig. 3E). Note the contact between the postfrontal and frontal, and the position of the pineal foramen entirely within the frontals. The latter distinguishes Ichthyosaurus from Protoichthyosaurus. Abbreviations: en, external naris; f, frontal; la, lacrimal; mx, maxilla; n, nasal; o, orbit; p, pineal foramen; pa, parietal; pmax, premaxilla; prf, prefrontal; ptf, postfrontal; st, supratemporal.
6.b.3. Position of the maximum width of the nasals
A third characteristic of the dorsal skull roof of Protoichthyosaurus that is worth noting is that the nasals are widest at their posterior end, dorsal to approximately the middle of the orbit (Fig. 1). At their posterior extent, the nasals make up most of the width of the skull, and the contact with the frontal and postfrontal is usually roughly perpendicular to the long dimension of the skull. This feature is shared with Ichthyosaurus (Motani, Reference Motani2005, fig. 3C is incorrect). In contrast, the nasals of Stenopterygius are somewhat narrower, with their widest point about level with the external naris, and they maintain that same width posteriorly to the orbits. In some specimens (e.g. SMNS 51133; Maisch & Matzke, Reference Maisch and Matzke2000, fig. 12; Motani, Reference Motani2005, fig. 2B), the nasals taper irregularly at the posterior end, and are lateral to the frontals, but in others they do not (e.g. S. aaleniensis, Maxwell et al. Reference Maxwell, Fernandez and Schoch2012, fig. 3A, B). Both morphologies are distinct from that of Protoichthyosaurus (and Ichthyosaurus), and are thus a useful distinguishing feature.
6.b.4. Other differences
Other, more subtle differences also occur. In many Protoichthyosaurus and some Ichthyosaurus specimens, an elongated internasal foramen occurs in a depression along the midline of the nasals (Lomax & Massare, Reference Lomax and Massare2018). The depression is often filled with sediment so the presence of the foramen can be difficult to confirm. In LEICT G729.1889, the cut and polished transverse section through the nasals shows that there is no suture between the elements, resulting in a 2 mm gap. However, this skull is sheared laterally so there could have been separation of a butt joint. Neither the depression nor the foramen has been reported in Stenopterygius.
The premaxillary teeth of Protoichthyosaurus have prominent, deeply grooved roots that have a circular cross-section. The grooves extend to the base of the crown and merge with the striations of the fairly robust crown. They are most similar to the teeth of I. somersetensis, but the grooves in the roots usually do not extend to the base of the crown in this species (Lomax & Massare, Reference Lomax and Massare2018). Stenopterygius and other species of Ichthyosaurus have more slender crowns and less prominent grooves in the roots, if any. Tooth morphology is not a good basis for identification, however, because it varies ontogenetically (Dick et al. Reference Dick, Schweigert and Maxwell2016), and many taxa share similar morphologies (Massare, Reference Massare1987; Foffa et al. Reference Foffa, Young, Stubbs, Dexter and Brusatte2018).
The three genera also have minor differences in skull shape, but these are difficult to quantify and preservation can alter it considerably. Protoichthyosaurus tends to have a lower crowned skull and a more robust rostrum than Stenopterygius. Species of Ichthyosaurus usually have a higher crowned skull than Protoichthyosaurus, although this is strongly affected by crushing. The relative rostrum length is variable within Ichthyosaurus. The relatively long, robust rostrum of Protoichthyosaurus is most similar to that of I. anningae and I. larkini. I. conybeari has a long rostrum, but it is much more slender. These three species have high-crowned skulls, which differs from Protoichthyosaurus. As with tooth morphology, skull shape alone is not particularly useful in distinguishing these genera.
7. Conclusion
The newly recognized specimens of Protoichthyosaurus prostaxalis clarify the morphology of the skull roof. The pineal foramen is between the frontals and the parietals, the main feature that distinguishes the skull roof of Protoichthyosaurus from Ichthyosaurus. The pineal position is shared with Stenopterygius. However, Protoichthyosaurus can be distinguished from that genus in skull roof morphology by a contact between the postfrontals and frontals (lacking in Stenopterygius; Motani, Reference Motani2005; Maxwell et al. Reference Maxwell, Fernandez and Schoch2012) and nasals that are widest at their posterior end. However, preservation of Lower Jurassic ichthyosaurs is most often in lateral view, and thus it is difficult to assess individual variation in morphology of the skull roof because so few well-preserved specimens are known.
The variation in the exposure of the maxilla described here among specimens of Protoichthyosaurus prostaxalis emphasizes the need to examine multiple specimens to evaluate intraspecific variation. The specimen described herein represents an extreme variation, where the maxilla makes up part of the ventral border of the external naris. In isolation, this might have suggested that the skull represented a new taxon. However, upon examination of other specimens, it is clear that this is just variation. Interestingly, as discussed above, a similar range of variation occurs in Stenopterygius.
Protoichthyosaurus shares many features of the skull roof with Ichthyosaurus, not unexpected given that the two genera are closely related (Lomax et al. Reference Lomax, Massare and Mistry2017, fig. 7). Many reconstructions of Ichthyosaurus exist in the literature (e.g. Sollas, Reference Sollas1916; Romer, Reference Romer1956; Motani, Reference Motani1999b, Reference Motani2005; Maisch & Matzke, Reference Maisch and Matzke2000; McGowan & Motani, Reference McGowan and Motani2003; Ji et al. Reference Ji, Jiang, Motani, Rieppel, Hao and Sun2016), but none of them is entirely correct due to a combination of misidentifications of specimens, difficulty in recognizing cranial sutures and a lack of well-preserved dorsal skulls. Contrary to the most recent study of the skull roof of Ichthyosaurus (Motani, Reference Motani2005), the prefrontal is not exposed medially on the skull roof and does not contact the frontal, except possibly in rare cases. Instead, the postfrontal and nasal overlap the prefrontal and contact the frontal, as in Protoichthyosaurus. The new reconstruction of the skull roof of Ichthyosaurus (Fig. 6) is similar to that of Motani (Reference Motani1999b, fig. 3E), with the correction that the pineal foramen is entirely enclosed by the frontals and positioned at the posterior end. This new interpretation might affect relationships among Early Jurassic parvipelvians in phylogenetic analyses, although that is beyond the scope of the present study.
Acknowledgements
We thank S Chapman, P Barrett, and L Steel (NHMUK), N Monaghan and M Parkes (NMING), P Joy (UOD), C Howells (NMW), J Clatworthy and K Riddington (BU), V Gausa (GPIT), E Maxwell (SMNS), M Riley and S Finney (CAMSM), E Howlett and H Ketchum (OUMNH), T Davidson (LDUCZ) and J Radley (WARMS) for help with and access to specimens at their institutions. Thanks also to B Gibson for providing photos of the specimen at UOD, T Smithson at the University of Cambridge for discussing the history and discovery of NHMUK R15943, and A Roberts for reviewing earlier versions of this paper. D.R.L.’s travel was partly covered by a PGR, Dean’s Doctoral Scholarship Award from the University of Manchester.
Conflict of interest
None






