



Fishing can be complicated. So is the archaeology of prehistoric fisheries. To fish well, one must know the specific vulnerabilities of the prey, conceive of a strategy that exploits these vulnerabilities, and design the appropriate equipment that makes it possible to catch them. The gear is diverse and sometimes species specific, and most ichthyofaunal assemblages lack direct evidence for the manner or method used to harvest them. When direct evidence for fishing technology is lacking, the techniques and equipment associated with fishing must be inferred from the structure of archaeological fish assemblages.
Numerous models have been deployed to reconstruct the gear and the associated social behaviors that can generate accumulations of fish in the archaeological record (Balme Reference Balme, Grigson and Clutton-Brock1983; Butler Reference Butler1996; Noe-Nygaard Reference Noe-Nygaard, Grigson and Clutton-Brock1983; Owen and Merrick Reference Owen and Merrick1994). The obvious assumption underlying these models is that certain kinds of fishing gear capture particular species or size ranges of fish and that this selectivity is reflected in the population structure of ichthyofaunal assemblages. The reconstructed population structure then can be used to address a variety of research topics of anthropological interest. Economic and evolutionary approaches focus on the energetic costs associated with fishing as well as the causes and consequences of subsistence change (Losey et al. Reference Losey, Nomokonova and Goriunova2008; Luff Reference Luff and Bailey2000; Van Neer Reference Van Neer2004). Ecological studies consider land and aquatic use strategies, and the status of past and present fish populations (Broughton et al. Reference Broughton, Martin, McEneaney, Wake and Simons2015; Hawkins and Caley Reference Hawkins and Caley2012; Leach and Davidson Reference Leach and Davidson2000). Taphonomic research considers issues of measurement, recovery bias, and the origins of fish assemblages (Balme Reference Balme, Grigson and Clutton-Brock1983; Butler Reference Butler1996; Greenspan Reference Greenspan1998; Noe-Nygaard Reference Noe-Nygaard, Grigson and Clutton-Brock1983; Owen and Merrick Reference Owen and Merrick1994).
Most of the fishing technologies considered up to this point entail some form of mass harvesting. This presents interpretive challenges for archaeologists attempting to relate the energetics of fishing to general models of foraging efficiency. Most of these models rely on assumptions that are particular to the survival of an individual forager, but the mass harvesting of fish is commonly a group-oriented or communal activity. The nature of the water habitat and seasonal changes in fish behaviors also condition foraging decisions, and they can complicate the return rates of foraging for fish. Moreover, fish, unlike most small-bodied animals that are gathered in mass, can be captured in so many different contexts and with such a wide variety of techniques that they can occupy a variety of positions in prey choice rankings (Lindström Reference Lindström and Plew1996).
Some scholars have attempted to model return rates of mass harvested prey relative to investments in different technologies (Ugan et al. Reference Ugan, Bright and Rogers2003) and the degree to which hunters would invest labor in enterprises, such as the production of large nets, with delayed rather than immediate returns (Satterthwait Reference Satterthwait1987). These are largely theoretical, however, and they have not been applied to archaeological datasets. Gear selectivity models that have been employed to identify the technologies associated with fishing in archaeological assemblages similarly fail to consider issues of labor investment and the organization of fishing in different aquatic environments. Therefore, as compelling as these models might seem, such shortcomings invite a host of middle range questions, which seek to unite the observed behaviors of people who fish with the static remains found in archaeological assemblages (Kelly Reference Kelly and Plew1996; Thomas Reference Thomas, Janetski and Madsen1990).
This article seeks to address some of these shortcomings in a comparative analysis of tui chub (Siphateles bicolor) assemblages from two archaeological sites, one in the Warner Valley of southeastern Oregon and the other in the Harney Basin of east-central Oregon (Figure 1). The former is situated in a series of shallow marshy lakes connected by streams and sloughs, and the latter is located in an open-water environment. Based on ethnographic and experimental data, a statistical model is presented that relates fish size to two types of mass harvesting, high-cost, high-return gillnet fishing in open water as compared to low-cost, low-return basketry or dipnet fishing in the marshes.
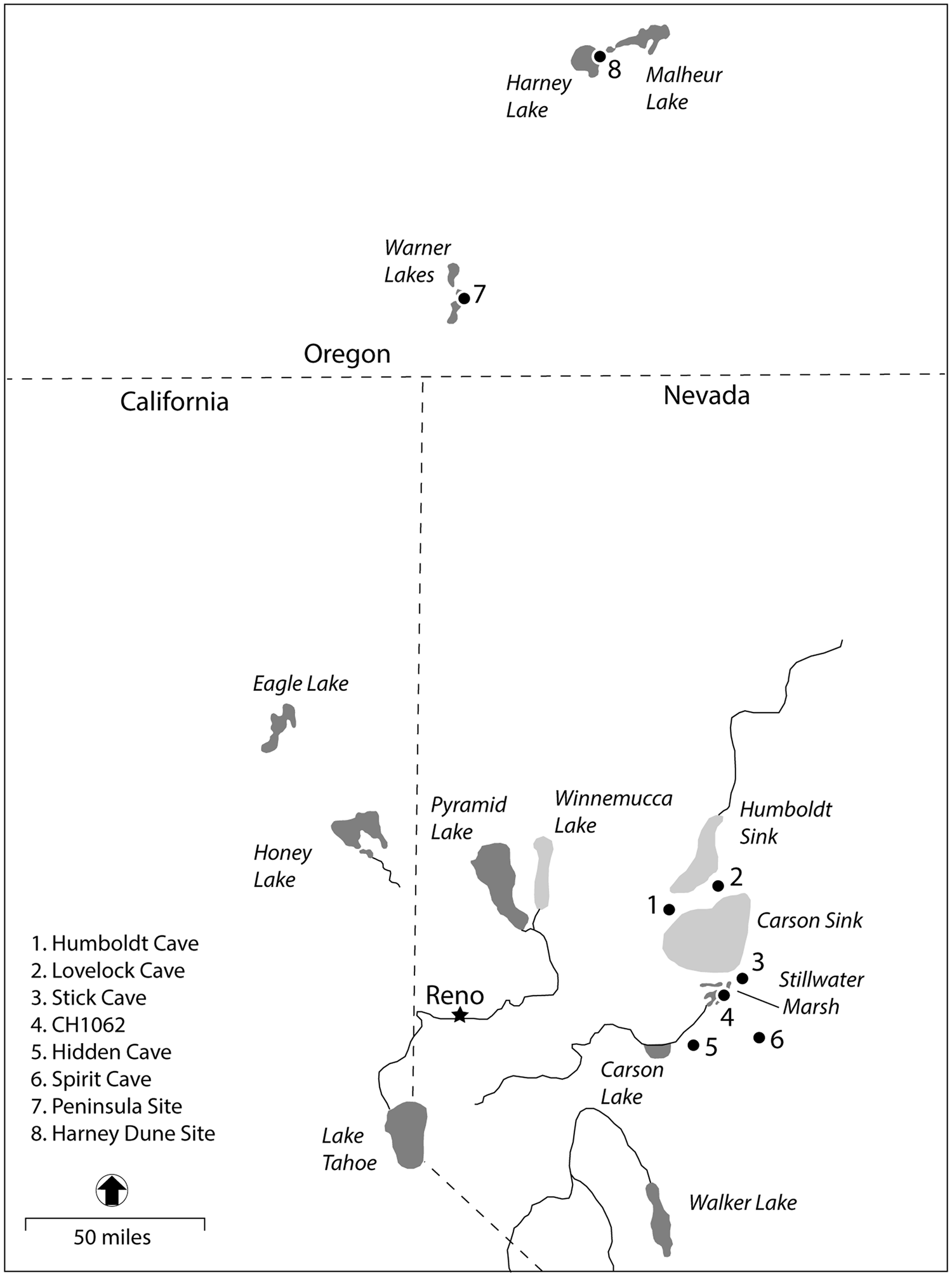
Figure 1. Locations of archaeological sites and landscape features mentioned in the text.
The fishing practices of the Great Basin Northern Paiute (Numa), as the most closely affiliated descendant community, provide an excellent opportunity to explore some of these issues. The Northern Paiute people exploited seasonally aggregated tui chub minnows that, when gathered in mass, constituted some of the most highly ranked resources in the region (Lindström Reference Lindström and Plew1996; Raymond and Sobel Reference Raymond and Sobel1990). Both men and women targeted tui chub in different water habitats, throughout multiple seasons of the year, and they used a variety of techniques that had different levels of investment and operating costs. Although environmental and cultural conditions have changed, and continue to do so, traditional Paiute fishing practices were probably similar in the precontact past.
I begin by reviewing the behavior and biology of tui chub populations in large and small bodies of water, and I relate this information to ethnographic accounts of Northern Paiute fishing, particularly those activities that were used to target tui chub in open-water and marshland settings. The different types of Paiute fishing gear used to capture tui chub demonstrates that investments in fishing were conditioned in part by water habitat and the sizes of tui chub in each setting. Modern gillnetting experiments conducted at Eagle Lake (California) and the Stillwater Marsh (Nevada) in the 1980s and 1990s provide data that link fish sizes and fishing returns to nets with different mesh specifications, but we currently lack information on basketry scooping, which was conducted primarily by Paiute women. In order to remedy this, I present my own experimental data, which I collected at Eagle Lake and Stillwater Marsh using simulated burden baskets and winnowing trays. These fishing simulations provided the experimental data that are needed to develop a regression formula to predict the sizes of archaeologically recovered tui chub as well as to evaluate the mean sizes and variances of tui chub captured with different gear types. This investigation, in turn, allows me to both identify the fishing techniques that were used at each of the two archaeological sites and compare these findings to several other Great Basin ichthyofaunal assemblages.
Finally, I present a technology investment model that analyzes the compositions of the two archaeological assemblages in relation to seasonality and relative commitments to fishing in different environmental settings. This analysis and the associated methodology are particularly useful for diagnosing the conditions under which net rather than basketry fishing may have occurred and identifying the social composition and organization of fishing parties most appropriate to either task. In addition to introducing a statistical methodology for reconstructing gear that typically does not preserve, this study provides new insights regarding traditional practices of Northern Great Basin fishing that specifically addresses the inferential gap between what we see on the ground and what we can learn from the data.
The Great Basin Tui Chub
Tui chubs are a group of western minnows (Cyprinidae) that are widely distributed in isolated aquatic systems across the Great Basin in western North America (Williams and Williams Reference Williams and Williams1981). They can be found in a variety of water environments, where their densities and sizes are conditioned by the scale and stability of the water system, and they are better adapted than other fishes to fluctuations in water temperature and salinity. Tui chub also are the first to invade dry shallow basins after they have flooded.
The life history of the tui chub is known primarily from studies conducted in large lake systems, where their abundances can reach into the millions. Using a combination of census methods, Heredia (Reference Heredia2014:56) calculated that standing stock in Eagle Lake, California, was 42.7 million, comprising up to 3,481 metric tons. A single tui chub female may produce as many as 68,900 eggs per year, but juvenile mortality is high (Kimsey Reference Kimsey1954; Kucera Reference Kucera1978). This, in combination with a short spawning season, results in a distinctly multimodal population size profile dominated by differently aged cohorts (Craig and Oertel Reference Craig and Oertel1966; Sheldon Reference Sheldon1965). Adult fish typically reach 25 cm standard length (SL) and can grow as large as 40 cm SL (Cooper Reference Cooper1985). Spawning begins in late April and culminates by mid- to late summer, when thousands of fish gather along the shoreline in large schools.
Less is known about the habits of tui chub occupying the smaller basin and marsh systems. Tui chub in these environments rarely exceed 12 cm SL, and they have shorter lifespans by at least three years and considerably smaller population densities (Bills and Bond Reference Bills and Bond1980; Bisson and Bond Reference Bisson and Bond1971; Snyder Reference Snyder1908; U.S. Fish and Wildlife Service 1998). Fish reach sexual maturity within one to two years at 8 cm in length (Bird Reference Bird1975), and although spawning may not involve massing at the scale seen in large lakes, tui chub densities increase substantially as the summer progresses (Raymond and Sobel Reference Raymond and Sobel1990).
The Organization of Northern Paiute Fishing
Northern Paiute fishing practices are similarly conditioned by aquatic habitat and the massing behaviors of tui chub in large and small bodies of water. Fishing in deep or open-water lakes is organized primarily through the cooperative efforts of men who targeted large-bodied species, including lake-dwelling tui chub, during the summer spawning season. Speth (Reference Speth1969:225) describes turn-of-the-last-century Paiute “fishing cliques” in the Walker River and Walker Lake systems in western Nevada as small, multimembered, non-kin-based groups of individuals who bonded as a more or less permanent and exclusive unit to achieve specific social and economic goals related to fishing. The men of the lake group consisted of two subgroups that worked as a unit in the manufacture, placement, and removal of several jointly owned gillnets. About three months of cooperative work were required to make a net, and group members divided the catch equally (Speth Reference Speth1969:234).
George Knerim, one of Willard Park's consultants from Walker River in the 1930s, describes a slightly different scenario involving about 20 men, “one from each family” who camped near the gillnet at night and divided the resulting catch in the morning (Fowler Reference Fowler1989:33). Despite these differences in the way that the catch was distributed, it is clear that net fishing was organized at the level of the clique or band in large lake settings where the potential of fishing returns was high and adult men were the primary participants in the catch.
Marshes and streams present a different series of environmental preconditions that involve smaller bodies of slow-moving and murky water. Fish populations are smaller—even tiny—and occur in much lower densities. Paiute fishing in these water systems focused on the capture of smaller tui chub. The men employed dipnets and the women used willow roasting baskets in the shallows of small streams and ditches (Fowler Reference Fowler1992:62; Kelly Reference Kelly1932:95; Stone Reference Stone and Mackedon1988:42). Dipnets were occasionally operated like a gillnet or a seine (Fowler Reference Fowler1989:32).
Fishing in the marshes was conducted as communal fish drives that involved the entire household or extended family. Reporting from the Carson Valley (Stillwater) marshes in June of 1859, Captain James H. Simpson noted, “A number of Pi-utes [sic], some two dozen, live near our camp, and I notice they have piles of fish lying about drying, principally chubs and mullet. They catch them with a seine”(Simpson Reference Simpson1876:85). The Toidikadi band in Stillwater Marsh employed waders to drive small tui chub into gillnets set across narrow channels or into square dipnets operated by the men on foot or in tule balsas (Fowler Reference Fowler1992:60; Stone Reference Stone and Mackedon1988:42).
Historical accounts indicate that although marsh fishing was seasonal, it focused on mass harvesting of tui chub to augment winter stores with dried fish in some areas. Writing about her tribe's concerns over the threat of white immigrants passing through the marshlands in the 1850s, Sarah Winnemucca Hopkins conveyed the following directive from her father, Chief Winnemucca (Wobitsawahkah): “If the emigrants don't come too early, we can take a run down and fish for a month, and lay up dried fish. I know we can dry a great many in a month. . . . In that way we can live in the mountains all summer and all winter too” (Hopkins Reference Hopkins1883:15). Helen Bowser Stone of Stillwater, Nevada, recalled that small fish were dried and “kept for the winter months” (Stone Reference Stone and Mackedon1988:44). Once captured, the responsibility of fish processing and storage fell to the women (Fowler Reference Fowler1989, Reference Fowler1992; Fowler and Bath Reference Fowler and Bath1981). Small fish were dried and stored whole, or they were eaten fresh and whole (Fowler Reference Fowler1992).
Northern Paiute Fishing Gear
Each of the two water environments—open lakes and shallow marshes—also relied on different types of fishing equipment with contrasting production and operating costs. Using data derived from Washo and Northern Paiute fisheries, Lindström (Reference Lindström and Plew1996:137–140) found that a gillnet with a 2.5 cm mesh size, measuring 30 m long × 1 m wide, requires over 1,900 hours to construct (63 hours per linear meter). Mesh openings as small as 1.3 cm require up to 2,220 hours to construct (73 hours per linear meter). By contrast, a 2 × 1 m dip- or liftnet can be made in 99 to 565 hours, depending on the size of the net and mesh openings. Baskets, generally used by women, provided the cheapest technological alternative because these could be constructed in 12 hours or less, and they could be used for a variety of tasks (not just fishing).
Lindström's data indicate that labor investments in fishing were substantial in open-water settings, where gillnets were needed to capture large and medium tui chub. Greater investments in gillnet production implies a similar commitment to fishing by men during the summer spawning season, when fishing could produce high returns with little effort once the net was set. The opposite was true in the marshes, where per-capita human-labor investment in communal drives to catch the fish was offset by the use of equipment, such as winnowing trays, with production costs that were next to nothing when compared with those of a gillnet. Similar contrasts pertain to the harvest. Larger fish, which are caught during the summer in open water, have to be eaten fresh or filletted and dried. Smaller fish captured with dipnets and scoops can be dried and stored with little to no processing costs.
These contrasts are well represented in ethnographic and archaeological nets with different mesh specifications. Table 1 lists the dimensions of 10 dip- and gillnets collected from Northern Paiute fisherfolk at Truckee, Walker, and Carson Rivers during the early to mid-twentieth century (Fowler Reference Fowler1992; Raymond and Sobel Reference Raymond and Sobel1990:6). The three dipnets that are attributed to tui chub all have openings that are 2 cm or less. These dipnets could be produced in approximately 500 hours, according to Lindström's data. The single gillnet has a 3 cm mesh opening that would require 1,900 hours or more and that was likely used to capture larger lake-dwelling chub.
Table 1. Dimensions of Ten Dipnets and Gillnets Collected from Northern Paiute Fisherfolk at Truckee, Walker, and Carson Rivers during the Early to Mid-Twentieth Century.

Sources: Fowler Reference Fowler1992; Raymond and Sobel Reference Raymond and Sobel1990:6, as attributed to data collected by Catherine Fowler.
Note: Specimen ID Categories: Milwaukee Public Museum (MPM), Lowie Museum (L), Museum of the American Indian (H).
Nets and net fragments recovered from dry caves in the western Great Basin are similar to ethnographic gill- and dipnets. Lovelock Cave yielded four gillnets and two possible dipnets. The gillnets measure 10–13 m in length and have mesh openings of 4 cm, whereas the dipnets have small mesh openings in the range of 1.5 cm (Loud and Harrington Reference Loud and Harrington1929:87, 89). Dipnets found in the vicinity of Hidden Cave have similar mesh openings between 0.5 and 1.6 cm (Ambro Reference Ambro1966:108), as do several net fragments with mesh openings between 0.5 and 3.3 cm (Goodman Reference Goodman and Thomas1985:281).
Low investments and processing costs associated with dipnetting or scooping are further implied by several ichthyofaunal assemblages recovered from dry caves in western Nevada. Stick Cave, on the northeast side of Winnemucca Lake, yielded numerous caches of small tui chub with an average length of 9–10 cm (Raymond and Sobel Reference Raymond and Sobel1990:8). The fish were found wrapped in moss and buried in thick layers of grass (Orr Reference Orr1952). Sun-dried caches of small, whole tui chub were similarly recovered from Humboldt and Lovelock Caves in western Nevada (Heizer and Krieger Reference Heizer and Krieger1956:93; Loud and Harrington Reference Loud and Harrington1929:11). Lovelock and Spirit Caves also yielded numerous human coprolites with small and tiny tui chub (Eiselt Reference Eiselt1997), and at Hidden Cave, digestive etching on tui chub elements further indicate that small fish were eaten whole (Smith Reference Smith and Thomas1985:173–176).
Tui Chub Fishing Experiments
Although it is well known that the Northern Paiute tailored their gear to capture tui chub of different sizes in the marshes and open-water systems, we still lack good information on how choice in gear type affects the size structure of captured fish, and consequently, the returns associated with fishing. Modern tui chub fishing experiments fill this important gap. Supplemental Tables 1 and 2 list the results of 13 gillnet sets conducted by Raymond and Sobel (Reference Raymond and Sobel1990) at Stillwater Marsh and Carson Sink in Nevada, and by Larry Davis (Reference Davis1986) at Eagle Lake in California. Experiments were conducted over the course of several months in 1986, 1987, and 1989, using gillnets with mesh openings that ranged between 1.27 cm and 3.8 cm. Raymond and Sobel's study took place in a marsh and shallow lake system, and the Davis study took place from the shoreline of a deep lake. Both studies targeted small, medium, and large tui chub, the results of which can be used to model the investment and return rates for gillnetting and establish the selectivity of gear with different mesh sizes.
Less is known about the selectivity and energetics involved in scooping techniques even though scooping and dipping were the methods most commonly utilized by women in ethnographic marshland settings. Unlike gillnetting (an unattended technique), pursuit involves active searching, dropping, and lifting using rigid equipment operated in shallow waters. In order to investigate distributions of fish sizes and the returns produced by this method, I devised a study using simulated burden baskets and winnowing trays at the Eagle Lake watershed in California and at Stillwater Marsh in western Nevada (Figure 1). Equipment was constructed to mimic the size and configuration of ethnographically known basketry. This included two burden baskets and two winnowing trays, each of equal size, with 0.64 cm (0.25 in.) and 1.27 cm (0.5 in.) mesh hardware cloth, respectively. The study targeted three habitat types (meadow stream, lakeshore, and marsh) at five separate locations (Figure 2). Table 2 lists the habitat types and the counts of fish species encountered over the course of each 15-minute interval of fishing at each site.

Figure 2. Basketry fishing experiment locales showing water habitat types: (a) Bly Tunnel, Willow Creek; (b) Eagle Lake shoreline; (c) Murrer's Meadow, Willow Creek; (d) Cattail Lake Canal, Stillwater Wildlife Refuge. (Photographs by B. Sunday Eiselt.)
Table 2. Counts of Fish Species Captured in the Five Water Habitats Sampled as Part of the Basketry Fishing Experiment.

Note: All field work conducted on 8/6/1997 and 8/7/1997.
Table 3. Summary Returns from Basketry Fishing Experiments Using Simulated Burden Baskets and Winnowing Trays with 6.35 mm and 12.7 mm Mesh Operated over 15-Minute Intervals in Five Water Habitats.

Robert Bear, a wildlife biologist, operated the equipment in order to maintain consistent results. Techniques involved searching for small schools of fish by wading upstream through a section of the exploitation zone, scooping, and immediately transferring the catch into a water-filled bucket. At the end of each timed interval of fishing, we transported the bucket to the shore, where we measured total length, standard length, and mass in grams for each fish identified to species prior to its release.
Although our efforts resulted in the capture of multiple native and non-native species of tiny fish, which generally reflected the compositions of fish populations in each water habitat, the majority of the captured fish (39%) were tui chub, and 97% of the total sample (305 of 313 individuals) were cyprinid species that had similar caloric returns, body part ratios, and swimming abilities (tui chub = 39%, mosquito fish = 26%, speckled dace = 18%, and Lahontan redside = 15%). Given that the primary goal of the fishing experiment was to sample different water habitats and obtain data on the lengths and mass of small fish captured with simulated baskets, the effects of species diversity on the study are immaterial to links between technology and fish size or encounter rates.
Results of the study indicate that return rates for basketry-harvested fish are conditioned by equipment type and water habitat (Table 3; see also Supplemental Table 3 for raw data). Burden baskets returned twice as many individuals and three times the mass in fish as compared to winnowing trays. The large and deep capture area of conical baskets (relative to the shallow and smaller surface area of winnowing trays) increases the likelihood that a fish will be retained upon encounter. In both cases, half-inch mesh was ineffective at capturing tiny fish in the shallows because tiny fish passed through the mesh and larger (faster) fish were able to avoid capture. Constricted and shallow water channels, such as the Bly Tunnel and the Stillwater Cattail canals, yielded the highest returns where fish had fewer options for escape and the equipment was easier to manipulate in shallow water. The difficulties associated with operating scoops in deeper water (in excess of 80 cm deep) reduced the encounter rates for any fish larger than 7 cm because these fish were faster than their smaller counterparts.
Gear Selectivity in the Mass Harvesting of Tui Chub
The fishing experiments yield data that can be used to compare fishing technique and mesh size for reconstructed fish lengths from archaeological assemblages. Gillnets and dipnets should produce distinctively different fish size distribution profiles (Hamley Reference Hamley1975; Jensen Reference Jensen1986, Reference Jensen1990; Rollefsen Reference Rollefsen1953). Gillnets capture a narrow range of fish sizes because mesh openings entangle fish by the gills. Smaller fish escape and larger fish back away from the mesh openings. Gillnetted populations, therefore, should have a narrow and normal distribution in most cases, whereas dipnets exhibit unimodal profiles that are skewed in favor of smaller fish.
Figure 3 illustrates the relationship between tui chub standard length (SL) and mesh size for the gillnet and basketry experiments. Mesh openings that range from 0.64 cm to 1.0 cm capture tiny fish measuring from 1 to 7 cm SL, whereas those with mesh openings from 1.27 to 1.9 cm capture small fish measuring from 8 to 13 cm SL. Medium fish measuring 14–19 cm SL are captured with mesh sizes from 1.9 to 3.8 cm, and large fish greater than 19 cm SL are vulnerable to mesh sizes greater than 3.8 cm.
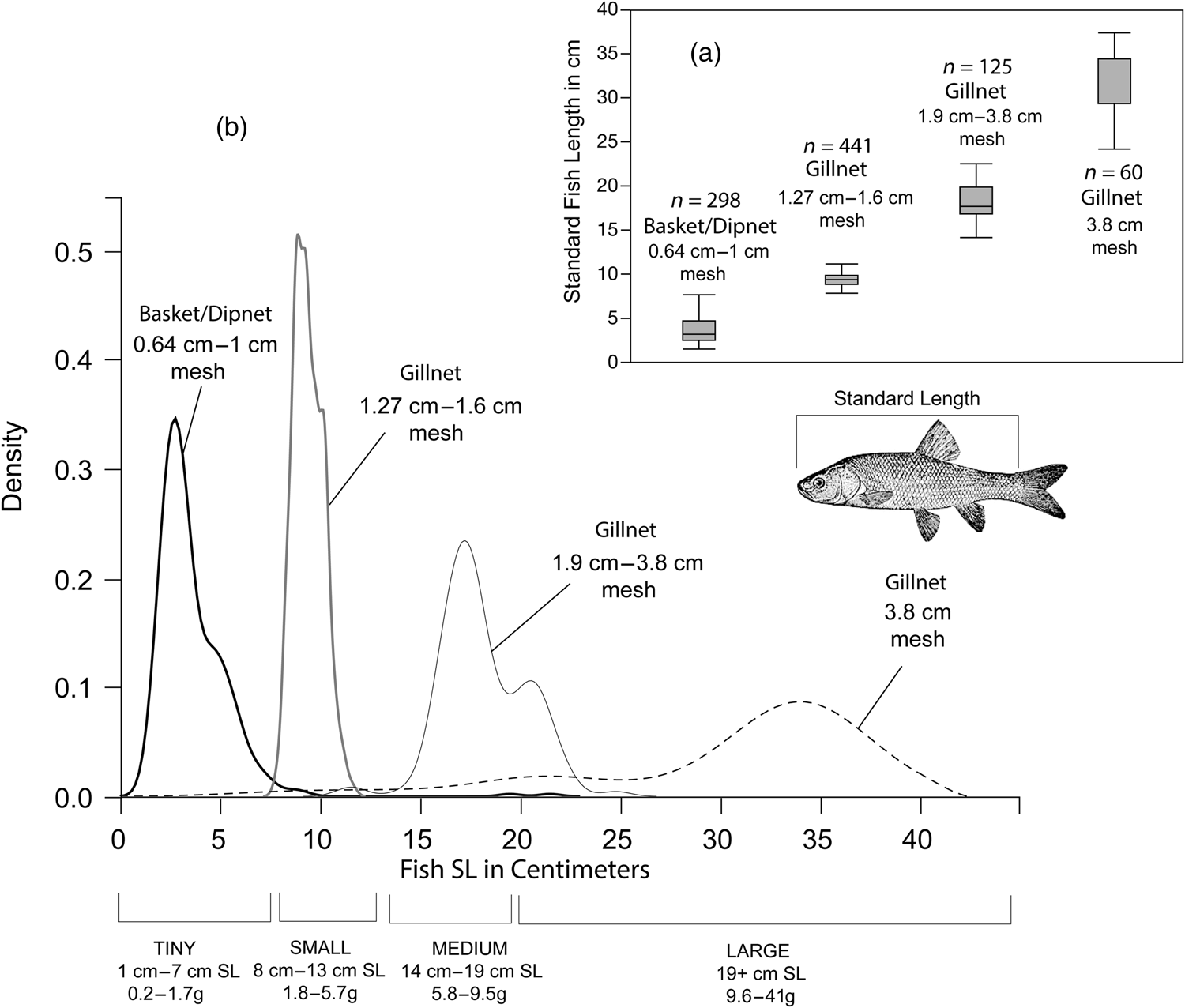
Figure 3. Relationships between tui chub standard length (SL) and mesh size for basketry compiled as part of this study, as well as gillnet data generated by Raymond and Sobel (Reference Raymond and Sobel1990) and by Davis (Reference Davis1986): (a) box and whisker plots showing means, quartiles, and ranges for fish SL in centimeters, ordered from smallest to largest and labeled with the corresponding mesh size and count of individual measured fish (n = 999 total fish); (b) smoothed density estimate curves for the same data. Fish captured with different mesh sizes can be segregated into tiny, small, medium, and large tui chub with corresponding size ranges and mass listed below the x-axis of the smoothed density plot.
Experimental data also can be used to evaluate the selectivity of different gear types by comparing mean fish size with the dispersion of fish lengths around the mean using the coefficient of variation (CV). The CV is the sample standard deviation divided by the sample mean, multiplied by 100, to arrive at a standardized measure of dispersion, expressed as a percentage value (Eerkens and Bettinger Reference Eerkens and Bettinger2001:495; VanPool and Leonard Reference VanPool and Leonard2011:80). The main benefit of the CV is that it allows metric data among samples of variable absolute sizes to be compared. One common application of the CV in archaeology is to investigate standardized production, with the assumption that small coefficients of variation indicate greater standardization of products, and consequently, specialized production. Larger coefficients of variation typically are assumed to reflect generalized production at the household level (Blackman et al. Reference Blackman, Stein and Vandiver1993; Roux Reference Roux2001; Stark Reference Stark2003:217). In the case of fish nets, lower CVs should indicate relatively selective gear types, such as gillnets, that target specific fish sizes. Generalized gear, such as dipnets or basketry scoops, should be reflected in higher CVs.
Figure 4 is a bivariate plot of mean tui chub SL in centimeters and the CV values for 999 tui chub caught with the experimental basketry and gillnets (See Supplemental Table 4 for summary values). Ellipses are labeled according to gear type. Plotted data points within ellipses represent mean and CV values for fish captured during 20 separate fishing events in three different water environments (meadow/stream, lake/shoreline, and marsh). The results indicate that gillnets tend to capture relatively large fish between 14 and 30 cm SL and exhibit relatively low CV values on the order of 7%–15% in most cases. One exception is Net D, which captured small, medium, and large fish and yielded CV values that varied between 14% and 30% over four sets in Eagle Lake. These variable results are probably due to the combined construction of this net, which included three linear sections of 1.3 cm, 1.9 cm, and 3.8 cm mesh openings, respectively. The nine other gillnetting events all produced CV values at or below 18%. Also notable is the CV value for Net F, a linear beach seine net, which returned grossly inflated CV values (137%) because of two large (27+ cm) fish that were trapped in the net. When these individuals are removed, the CV value (11.6%) is consistent with expectations using a gillnet. From this, it may be inferred that assemblages returning inflated CV values similar to Net F may indicate the use of a seine that occasionally traps large fish (hook-and-line or spearing techniques might also produce a similar result). Basketry scooping, on the other hand, tends to capture tiny fish between 1 cm and 7 cm SL, yielding CV values between 20% and 30%. Dipnets should produce CV values similar to baskets, but with small to medium fish sizes corresponding to the size of the mesh.
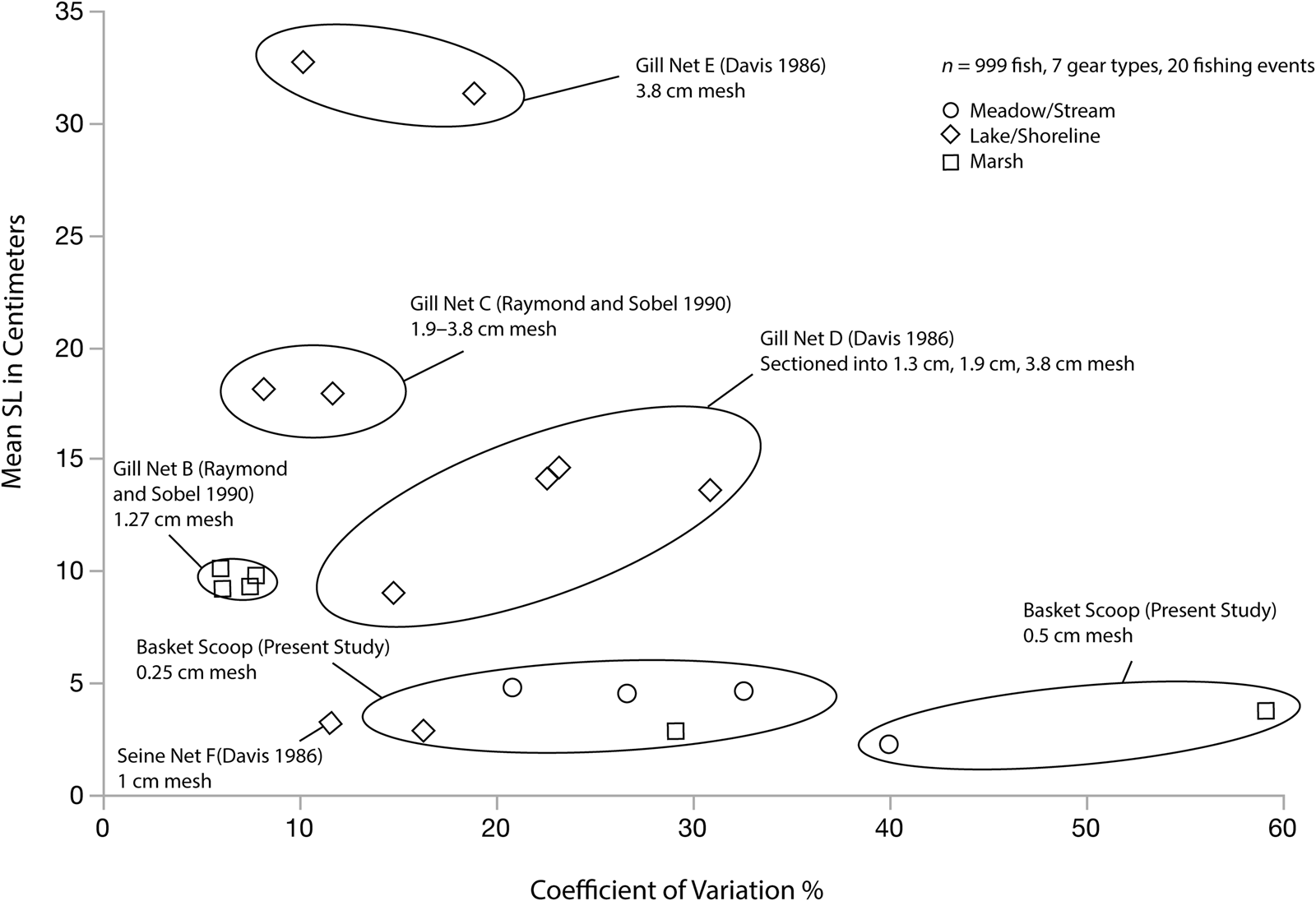
Figure 4. Bivariate plot of mean tui chub SL in centimeters and the CV values for 999 individuals caught with experimental basketry scoops and gillnets. Ellipses are labeled according to gear type (n = 7). Plotted data points within ellipses represent mean and CV values for fish captured during 20 separate fishing events in three different water environments (meadow/stream, lake/shoreline, and marsh). Net D appears to have been constructed in sections of 1.3 cm, 1.9 cm, and 3.8 cm mesh.
Fishing experiments demonstrate that mean fish SL and CV values can be plotted to reveal selective versus general harvesting techniques. Based on these patterns, it is possible to conclude that archaeological assemblages of larger-bodied fish assemblages with low CV values were most likely produced by gillnets, whereas assemblages of smaller-bodied fish yielding higher CV values were the result of fishing with technologically less costly baskets or dipnets. Each of these two contrasting techniques for mass harvesting is associated with a particular fishing environment: gillnet fishing is most appropriate for deeper or more open lake contexts yielding larger fish, whereas fishing with a dipnet or basket for small fish is the most effective method in shallow marshes. These practices have not only a determining effect on the size of fish appearing in tui chub assemblages but also implications for understanding the economic trade-offs of ancestral Paiute fishing in open lakes and shallow marshes. Nets are expensive to produce and maintain, and in the pursuit of smaller fish, the production of smaller mesh sizes requires exponentially more work that ultimately results in diminishing returns. Basket scoops and dipnets are low cost and, although some physical effort may be required to achieve a successful harvest, they are all that is needed to capture tiny minnows in large quantity with little to no post-processing of the catch.
Comparative Analysis of Archaeological Fish Remains
Tui chub assemblages are ubiquitous at many Great Basin sites, but these assemblages are rarely employed to investigate the organization of fishing and investments in different gear types. This may be attributed, all or in part, to the fact that fishing equipment rarely preserves in association with mass-harvested ichthyofaunal assemblages, When it does, it is either highly fragmentary (nets, for example), or it is preserved in dry caves, which are well removed from the behavioral context in which the gear was used. Consequently, based on the archaeology alone, we understand very little about the conditions under which net and basketry fishing may have occurred.
Tui chub size distribution profiles from two archaeological sites in southeastern Oregon are considered here. The Harney Dune site is interpreted as an example of gillnet fishing, whereas the Peninsula site is believed to be an example of fishing using dipnets and basketry scoops (Figure 5). Following brief summary descriptions, a regression analysis is used to reconstruct fish size and fish population profiles represented in faunal assemblages recovered from each site. Based on the previous review of ethnographic and experimental data, SL and CV values are calculated to infer the type of gear that was used and, by association, the social composition of the fishing parties.
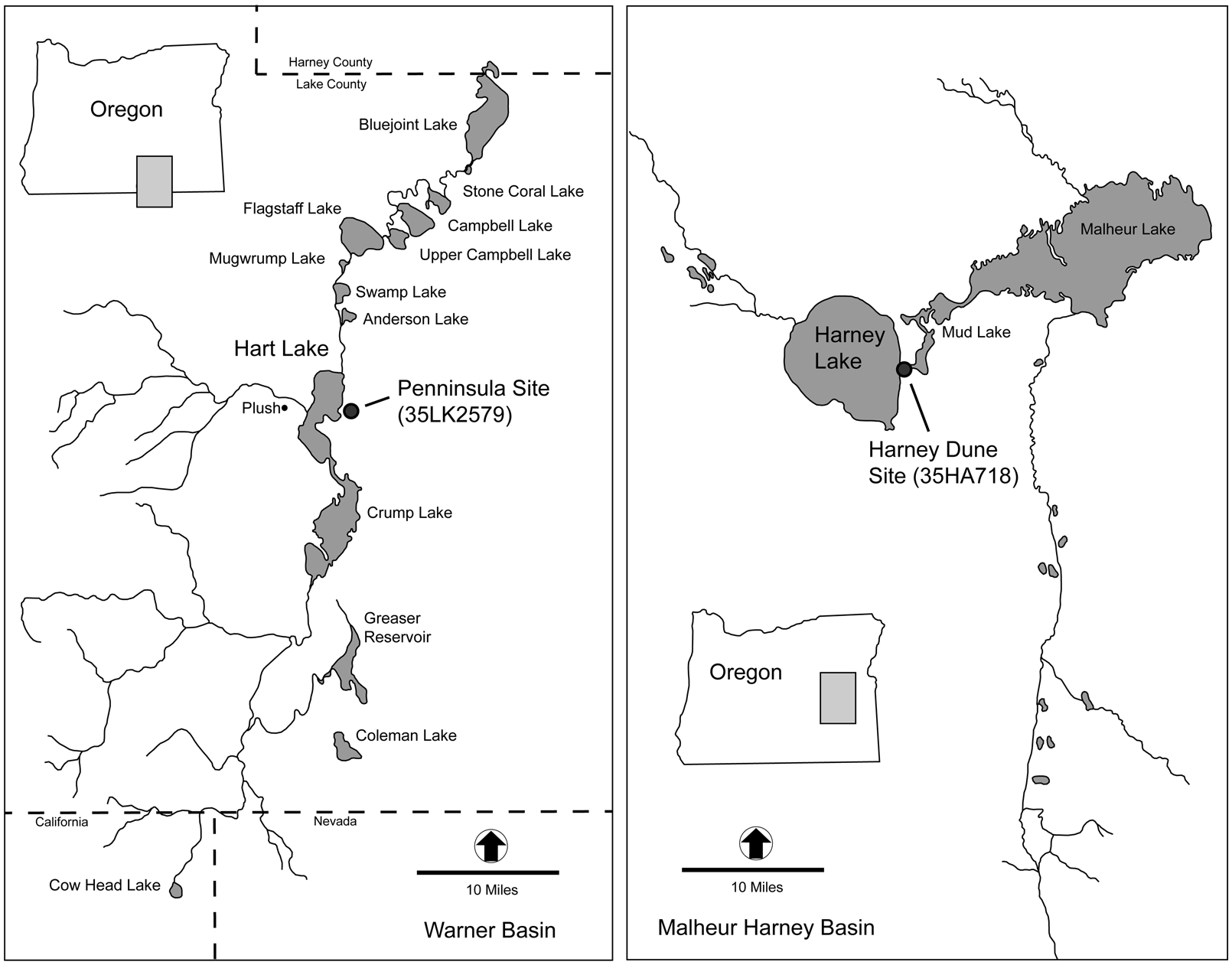
Figure 5. Locations of archaeological sites included in this study and associated water environments.
The Harney Dune Site
The Harney Dune site (35HA718) is an extensive artifact scatter situated on the north shore of Harney Lake (Raymond Reference Raymond1994). Currently, the lake covers 106 km2 (26,400 acres) with a maximum depth of 2 m. The artifact assemblage consists of chipped and ground stone tools, debitage, fire-cracked rock, stone net weights, bone barbs, and thousands of burned and unburned fish remains. Several features containing groups of notched stones found clustered together are interpreted as abandoned nets, which—following decomposition—left only the net weights behind. Greenspan (Reference Greenspan1998) reconstructed the size distribution profile of tui chub specimens to test the hypothesis that deliberate and size-selective fishing occurred at the site. She concluded that foragers mass-captured tui chubs using gillnets based on the fish size distribution profiles from three different features: a shallow pit (Feature 68) that dates from the early AD 1500s to 1600s and two shallow middens. Feature 89 dates from the mid to late AD 1800s, and Feature 66 dates from the early AD 1700s to 1800s (Greenspan Reference Greenspan1998:979–980). Feature contents were removed from the field in bulk, and they were screened in the laboratory through nested sieves of 3 mm and 1 mm mesh.
The Peninsula Site
The Peninsula site (35LK2579) is located on the eastern shore of Hart Lake in the Warner Basin. Hart Lake presently covers 29.64 km2 (7,324 acres) with an average depth of 1.5 m, and it exists as one of a series of small lakes connected from north to south by marsh-lined sloughs and canals. The site represents an intensively occupied winter-to-spring residential camp with numerous pithouse features. Excavations produced late-period projectile points, bone tools, and a substantial ichthyofaunal assemblage (Eiselt Reference Eiselt1998). Dated features revealed at least two occupations. Structure 1 (Feature 304) probably dates from the mid to late AD 1600s, Structure 2 (Feature 632) from the early to late AD 1400s, and Structure 3 (Feature 4) also from the early to late AD 1400s (Eiselt Reference Eiselt1998:21, 37, 41; see also Supplemental Figure 1 for uncalibrated and calibrated radiocarbon dates). A dense midden deposit (Feature 106) is undated, but it yielded a rich ichthyofaunal assemblage. All fish elements were recovered from well-defined cultural contexts (house floors, house fill, and middens) using 3.2 mm screens.
Fish Size Reconstructions
Fish size is perhaps the most direct indicator of the gear used in mass capture. In order to investigate this, I secured bone measurements from 20 modern tui chub specimens to develop baseline data that could predict fish length based on bone size for archaeological assemblages. Specimens included tui chub captured during the basketry experiment and skeletonized tui chub from the University of Michigan, Museum of Zoology in Ann Arbor (Figure 6a; Supplemental Tables 5 and 6). I took seven measurements on six skeletal elements. One measurement of the cleithrum was selected for analysis because this element was the most abundant and best preserved at the Peninsula site, and it is highly correlated with fish length (r2 = 0.98). I then measured a total of 158 individuals from the four feature assemblages at the Peninsula site and applied a regression formula to reconstruct fish sizes based on the reference bone data. Fish size data for the Harney Dune site were derived from Greenspan (Reference Greenspan1998), who used the hyomandibular and basioccipital elements to reconstruct the sizes of 196 individuals from Features 66, 68, and 89 (Figure 6b; Supplemental Table 7).

Figure 6. Measurements utilized to reconstruct fish sizes from (a) Peninsula site and (b) Harney Dune tui chub assemblages.
Additional calibrations were necessary in order to normalize the data from different datasets. Greenspan (Reference Greenspan1998) used total fish length (TL) for the Harney Dune site, and Raymond and Sobel (Reference Raymond and Sobel1990) also recorded TL in their fishing experiment for tui chubs at Stillwater Marsh. The present study recorded SL and TL for all modern tui chub reference specimens. This enabled me to calculate a regression formula to derive standard lengths from total lengths (SL = 0.31 + 0.83 × TL) for all comparative specimens. The resulting reconstructed fish sizes are grouped into tiny, small, medium, and large categories. These categories are correlated with fish mass and the mesh size required to capture each size class.
Mesh Size and Gear Choice
Figure 7 provides smoothed density estimate curves for reconstructed fish sizes from each of the seven features at the two archaeological sites relative to capture data generated by Raymond and Sobel (Reference Raymond and Sobel1990), Davis (Reference Davis1986), and the basketry experiment presented here (Supplemental Tables 1, 2, 3, 7). As expected, fish sizes for each feature from the Peninsula site overlap with the 0.64–1.27 cm mesh sizes that are common for dipnets and basketry scoops, whereas fish size ranges at the Harney Dune site overlap with the 1.9–3.8 cm gillnet mesh sizes. Fish from the Harney Dune site are larger than those at the Peninsula site, and these differences are statistically significant (t[352] = 1.96; p = 0.05). It is likely that recovery bias is present in the Peninsula site assemblage due to the use of 3.2 mm mesh screens and the potential loss of tiny bone specimens (Butler and Schroeder Reference Butler and Schroeder1998), but even if these elements were recovered and added to the measured assemblage, the conclusions would be the same. The Peninsula site assemblage is dominated by tiny and small tui chub.
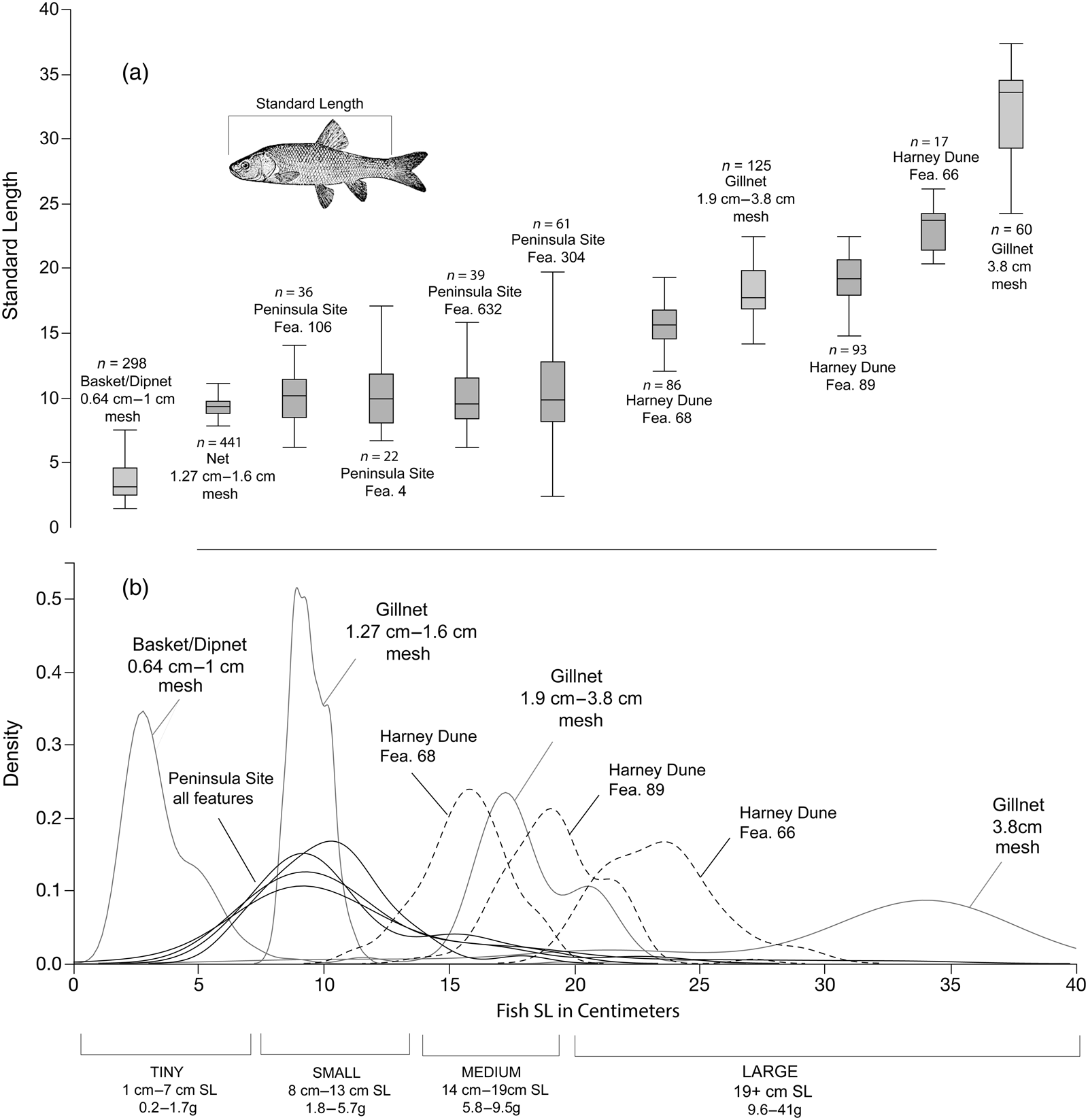
Figure 7. Box and whisker plots and smoothed density estimate curves for reconstructed fish sizes from each of the seven archaeological features at the Peninsula and Harney Dune sites relative to capture data as referenced in Figures 3a and 3b. Figure 7a displays box and whisker plots showing means, quartiles, and ranges for fish SL in centimeters, ordered from smallest to largest and labeled with the corresponding feature or gear type. Figure 7b displays smoothed density estimate curves for the same data, showing overlaps between reconstructed fish sizes recovered from archaeological features and the sizes of modern fish captured with different mesh openings.
Additional support for the use of gillnets at the Harney Dune site is evident in the CV values for the fish assemblages from each of the three features (Figure 8; Supplemental Table 4). All CV values are less than 11%, and they are consistent with the low CV values associated with experimental gillnets. The abundance of net sinkers at the Harney Dune site (n = 50+) further suggests a fishing strategy that utilized gillnets to target medium to large tui chub. If this is the case, then men were the likely foragers. This pattern is consistent with Walker Lake Paiute techniques that involved cooperation organized at the level of the fishing clique or band (Speth Reference Speth1969).

Figure 8. Bivariate plot of mean tui chub SL in centimeters and the CV values for fish recovered from different archaeological sites. Open ellipses are labeled according to gear type as referenced in Figure 4. Plotted data points represent mean and CV values for archaeological fish assemblages coded by site. Shaded ellipses encompass fish assemblages from each of the seven features from the Peninsula and Harney Dunes sites.
The Peninsula site assemblages show the opposite pattern. CV values are greater than 23%, and they are consistent with the higher CV values associated with experimental basket scooping and, by extension, dipnetting. The general lack of net sinkers in excavated contexts at the site (n = 1) also suggests a focus on dipping as the preferred procurement strategy to target small and tiny tui chub. This pattern is consistent with ethnographic information from the marsh-dwelling Toidikadi, who occupied the Carson Sink area. Male and female foragers probably utilized dipnets or basketry scoops and employed fish drives that were organized at the level of the nuclear or extended family (Fowler Reference Fowler1992).
Figure 8 also contains comparative data for six additional Great Basin sites to better represent regional variation in fishing practices. Of these sites, only the Stick Cave cache of dried and wrapped tui chub returned CV values that are consistent with selective linear net fishing (CV = 8.1%). The tui chub in this assemblage also are extremely small (x̅ = 8.6 cm SL). This pattern is similar to the experimental results obtained by Davis (Reference Davis1986) for Net F, a 10 m long net that was operated in July as a beach seine. The net was set at a depth of 0.5–1.0 m at Lederer Marsh, Eagle Lake, where tiny, foraging tui chub were massing in large numbers. The use of a dipnet as a seine would have the effect of producing assemblages of small fish with low CV values.
The tui chub assemblage from Humboldt Cave also represents a cache of dried tui chub, but fish sizes and CV values are more consistent with the Harney Dune site, where gillnets with 1.9–3.8 cm mesh openings captured medium to large fish. The Goshen Island assemblages from Utah Lake (Utah's largest freshwater lake) yielded similarly large sizes of tui chub, but with high CV values, possibly representing a palimpsest of fishing events involving different set depths or gillnets with variable mesh sizes (as in Davis's Net D; Nauta Reference Nauta, Madsen and Metcalf2000).
Assemblages from Lovelock Cave and 26CH2062 (Stillwater Marsh) are more ambiguous, but both have higher CV values in the range of 20%–34% and tiny to small fish sizes in the range of 4–14 cm. These fishbone assemblages are similar to those produced by basket scooping or dipnetting. The results are particularly compelling in the case of Lovelock Cave, given that two dipnets with mesh openings of 1.5–4.0 cm were found in association with tui chub remains identified in coprolites (Follett Reference Follett1967). Although CV values are not available for the fish recovered from the intestinal remains of either the Spirit Cave individual or the Hidden Cave coprolites, reconstructed mean lengths for these assemblages are between 3 cm and 6 cm at Spirit Cave, and they are between 10 cm and 12 cm at Hidden Cave (Eiselt Reference Eiselt1997; Smith Reference Smith and Thomas1985), which are similarly suggestive of scooping or dipping techniques.
Technological Investments in Gillnetting and Scooping
Regional variation in ichthyofaunal assemblages suggest that ancestral Paiute fisherfolk adapted their fishing strategies to fit different environmental contexts. Gillnets provide the best solution for targeting larger fish in open water, whereas low-cost dipnets and scoops used to catch small and tiny fish are the best solution in murky marshes and streams. It also is apparent that the the two strategies constituted different levels of investment in fishing based on anticipated returns relative to other pursuits. Gillnets have greater manufacturing costs, but they produce greater returns in the summer with little to no effort once the net is set. Baskets and dipnets are cheaper to make and maintain, but they require substantial labor in the form of family members who drive the fish and those on the receiving end who have to “stoop and scoop,” with potentially lower energetic returns overall. The question then becomes, when and under what conditions do greater investments in fishing pay off (Ugan et al. Reference Ugan, Bright and Rogers2003)?
This can be assessed by a simple point-estimate model of technological investment. Technology investment models predict the amount of time a cheaper technology should be used before a more costly alternative produces returns that are equal to or greater than the cheaper alternative per unit of time invested in manufacture. Bettinger (Reference Bettinger2009:65) refers to this as the “procurement time switching point” between two technologies.
Figure 9 presents the procurement return rates (r), manufacturing times (m), and procurement switching times (s 1↔2) for the small and medium mesh nets presented in Figure 4. Returns in kcals per hour for each fishing event are plotted against manufacture costs for each piece of equipment used during that event (Lindström Reference Lindström and Plew1996). In order to explore patterns in the assemblage of tiny to small tui chub represented at the Peninsula site, manufacturing costs and return rates for burden baskets and small-mesh dipnets are combined in this analysis and compared to Net B—a small-mesh, 10.9 m long gillnet. Manufacture costs for different nets and returns are compared to each other in order to explore patterns in the medium fish assemblage represented at the Harney Dune site.
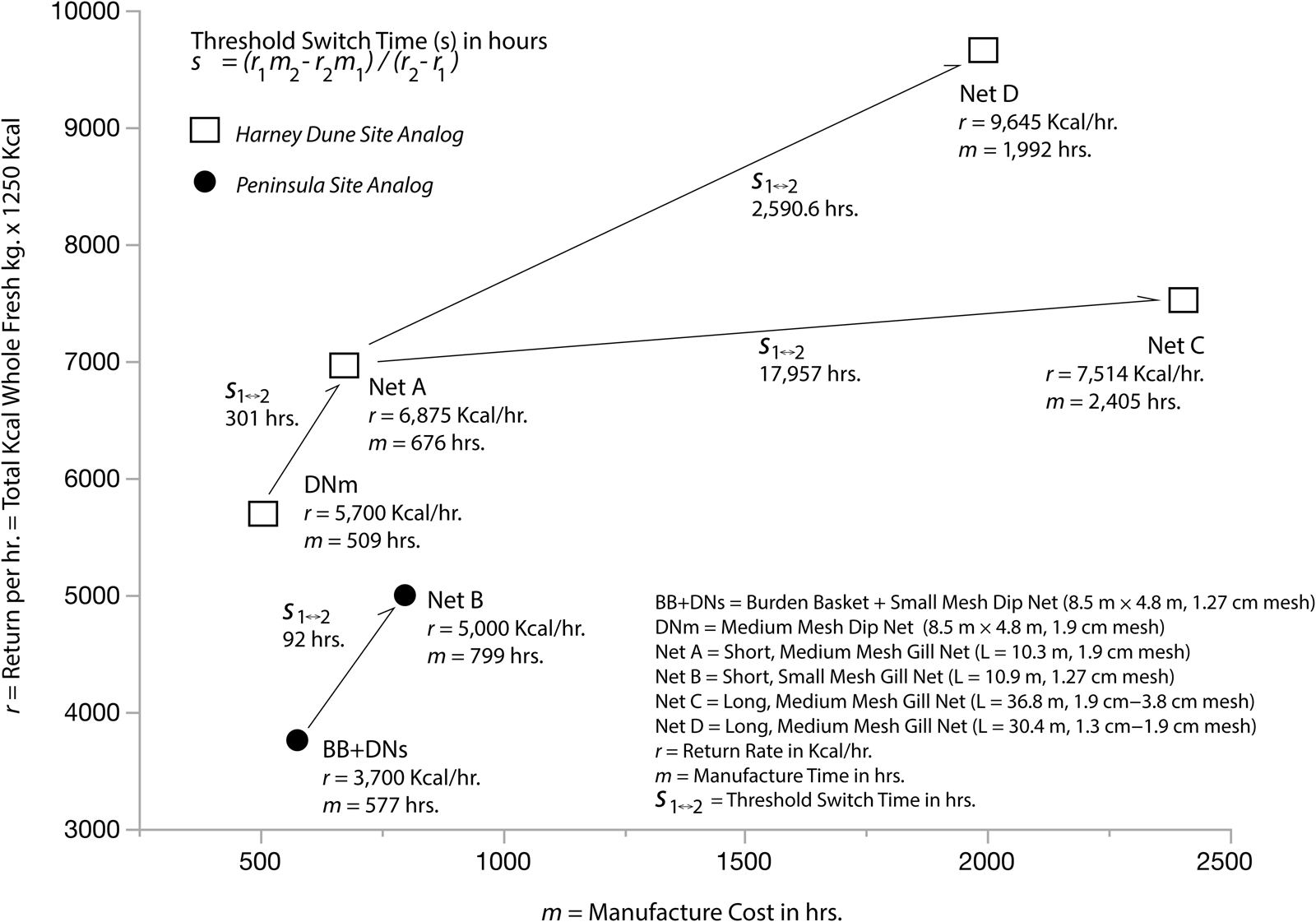
Figure 9. Procurement return rates (r), manufacturing times (m), and procurement switching times (S 1↔2 ) for the small and medium mesh nets in Figure 4 (see Supplementary Table 4).
As Figure 9 shows, the switching point between scooping or dipnetting and Net B is only 92 hours. In other words, the gillnet should be a better investment when a forager expects to spend more than 92 hours using it. At the Peninsula site—a marsh environment—higher CV values indicate that the assemblage consists of tiny and small fish captured in baskets or dipnets. This implies that fish densities (and sizes) were not high enough to warrant the additional investment in a gillnet, consistent with conditions that would have prevailed during the late winter or spring when tui chub densities were low. It also implies that foraging opportunities were greater elsewhere in the surrounding environment during the summer season.
The opposite pattern is suggested by the lake-derived assemblage at the Harney Dune site. The switch time between a medium-mesh dipnet and Net A (a 10.3 m long, medium-mesh gillnet) is 301 hours. Although a dipnet might seem to be the least costly option, the CV values on reconstructed fish sizes indicate that these fish were captured with a gillnet. Fish densities must have been sufficiently high to warrant the additional investment in gillnet manufacture. This implies a higher commitment to fishing at the Harney Dune site and a fishing pattern that is consistent with summer spawning procurement. It also indicates that foraging opportunities were greater during the summer at Harney Lake than elsewhere in the surrounding environment.
Figure 9 further suggests that the experimental returns on long nets with the highest manufacture costs have threshold switch times in the thousands of hours relative to the shorter Net A. The returns for each of these longer nets would need to be doubled in order to bring down the switch times and make them worth the investment. In the case of Net D, a forager would have to anticipate capturing a total of 288 medium-sized tui chub—or 15 kg of fish per hour—conditions that might only prevail on rare occasions or in larger bodies of water. Consequently, the fish represented at the Harney Dune site were probably captured with a relatively short net.
Final Observations
This study demonstrates that mass harvesting of tui chub not only varied by water environment but was also heavily conditioned by gear choice and the social actors who participated in the catch. A methodology is provided that relates the size structure of ichthyofaunal assemblages to specific fishing techniques. Mean fish size relative to CV values for assemblages makes it possible to differentiate mass-harvested fish remains that were captured with a basket or dipnet from those that were caught in a gillnet or seine. Moreover, excessively high CV values may indicate assemblages that represent a combination of fishing techniques, a merging of multiple mass-harvesting events, or other factors such as a naturally occurring die-off of fish.
For the majority of cases examined here, assemblages tended to be either scooped or gillnetted, which, in turn, may be related directly to the organization of fishing in the past and the underlying conditions that guided investments in fishing. This can be summarized in terms of male fishing cliques that used gillnets that were expensive to make but cheaper to operate in open water environments, as opposed to family-oriented fishing with scoops and dipnets that were cheaper to make but more expensive to operate in the marshes. Use of baskets as scoops also would have been an activity that was generally confined to women, who would have been most involved in their upkeep and repair.
As Lindström (Reference Lindström and Plew1996) suggests, ranking of fish on a scale of prey types may be based on size, but as this study shows, these differences can also be attributed to regional adaptations and the seasonal use of water environments. For example, male social roles expressed through their participation in fishing cliques and communal net production almost certainly would have taken place in the summer, when the returns from fishing were high. This is in contrast to scooping or dipping, which could be conducted year-round but at potentially lower levels in some marshes. Peninsula site data and ethnographic accounts (Kelly Reference Kelly1932) indicate that women were heavily involved in these activities during the spring and that investments in fishing were relatively low in Warner Valley; however, in areas with extensive marshes (such as Stillwater Marsh), summer fishing was critical to augment winter stores with dried fish. Ethnographic evidence and the CV values obtained for small or tiny tui chub assemblages at the Peninsula site and elsewhere further suggest that dipping or scooping technologies may have been more common in some portions of the Great Basin where extensive marshlands once existed.
In summary, this exercise in middle-range analysis examines the behaviors associated with fishing in different water environments, and it employs the ethnographic record of descendant communities and experimental research to interpret archaeological site data. Mathematical models make it possible to accurately reconstruct fish sizes and distributions in archaeological assemblages as well as the choices and social constraints that may have pertained to different lacustrine environments. This information, in turn, can be placed within a global context of research on past fishing strategies that focuses on social, evolutionary, economic, or ecological interpretations (Butler Reference Butler1994; Kirch and Dye Reference Kirch and Dye1979; O'Connor et al. Reference O'Connor, Ono and Clarkson2011; Van Neer Reference Van Neer2004). Such studies do more than simply address issues concerning human adaptations in dryland lakes and marshes and the role of traditional ecological knowledge in fishing. They also fill a persistent evidentiary gap in the development of broader theoretical constructs that seek to understand forager economy and subsistence based, in part, on gear that does not preserve and the people we cannot see.
Acknowledgments
Special thanks go to Andrew Darling, Catherine Fowler, Don Fowler, Bill Cannon, Ruth Greenspan, Lawrence Davis, Anan Raymond, and Frank Bayham for their advice, suggestions, and fieldwork results. Thanks also go to three anonymous reviewers who provided thoughtful insights and suggestions. Robert Bear was instrumental in carrying out the fishing experiments and assisting with initial statistical analysis.
Data Availability Statement
The data generated for this analysis are available in the text and as supplemental materials.
Supplemental Materials
For supplemental material accompanying this article, visit https://doi.org/10.1017/aaq.2020.37.
Supplemental Table 1. Raymond and Sobel capture data.
Supplemental Table 2. Davis capture data.
Supplemental Table 3. Results of simulated basketry-scooping experiments.
Supplemental Table 4. Mean lengths, standard deviations, and CV values for modern fishing experiments and archaeological fish assemblages.
Supplemental Table 5. Measurements on elements from comparative tui chub specimens.
Supplemental Table 6. Information on the University of Michigan tui chub modern specimens used in this study.
Supplemental Table 7. Reconstructed archaeological fish lengths from the Harney Dune and Peninsula sites.
Supplemental Figure 1. Peninsula site (35LK2579), Lake County, Oregon, radiocarbon dates.














