1. Introduction
For over 30 years, following the pioneering work of Playford et al. (Reference Playford, Cockbain, Druce, Wray and Walter1976) on Devonian stromatolites from the Canning Basin of W Australia, there has been growing research interest in the possible linkage of red colouration in limestones and microbial activity. The red colour of condensed limestones, in particular, is considered by Mamet & Préat (Reference Mamet and Préat2006) as being due to the mediation of iron-fixing bacteria, and in some cases there is linkage to the problematic microbial(?) structure Frutexites, a common constituent of deep-water ‘ferroan stromatolites’ (see e.g. Playford et al. Reference Playford, Cockbain, Druce, Wray and Walter1976). Examples of red or pink limestones, where matrix pigmentation has been attributed to the activity of iron-oxidizing microbes, are now known from specific time intervals in the Silurian, Devonian and Jurassic as reviewed recently by Ferretti et al. (Reference Ferretti, Cavalazzi, Barbieri, Westall, Foucher and Todesco2012). The Lower Cretaceous Hunstanton Red Chalk Formation of eastern England (see e.g. Owen, Reference Owen1995; Mortimore, Wood & Gallois, Reference Mortimore, Wood and Gallois2001) has a number of attributes that suggest it could be included in this grouping: these red condensed and nodular chalks, reminiscent of the Ammonitico Rosso (Anderton et al. Reference Anderton, Bridges, Leeder and Sellwood1979, p. 239), are associated with underlying ironstone-ooid-bearing sandstones, and in their upper parts have surfaces that may be stromatolitic (Jeans, Reference Jeans1980). In this paper we have re-examined stromatolite-like sedimentary structures at the top of the Hunstanton Red Chalk Formation with a focus on their microstructures and their potential relationship with the prominent red colouration.
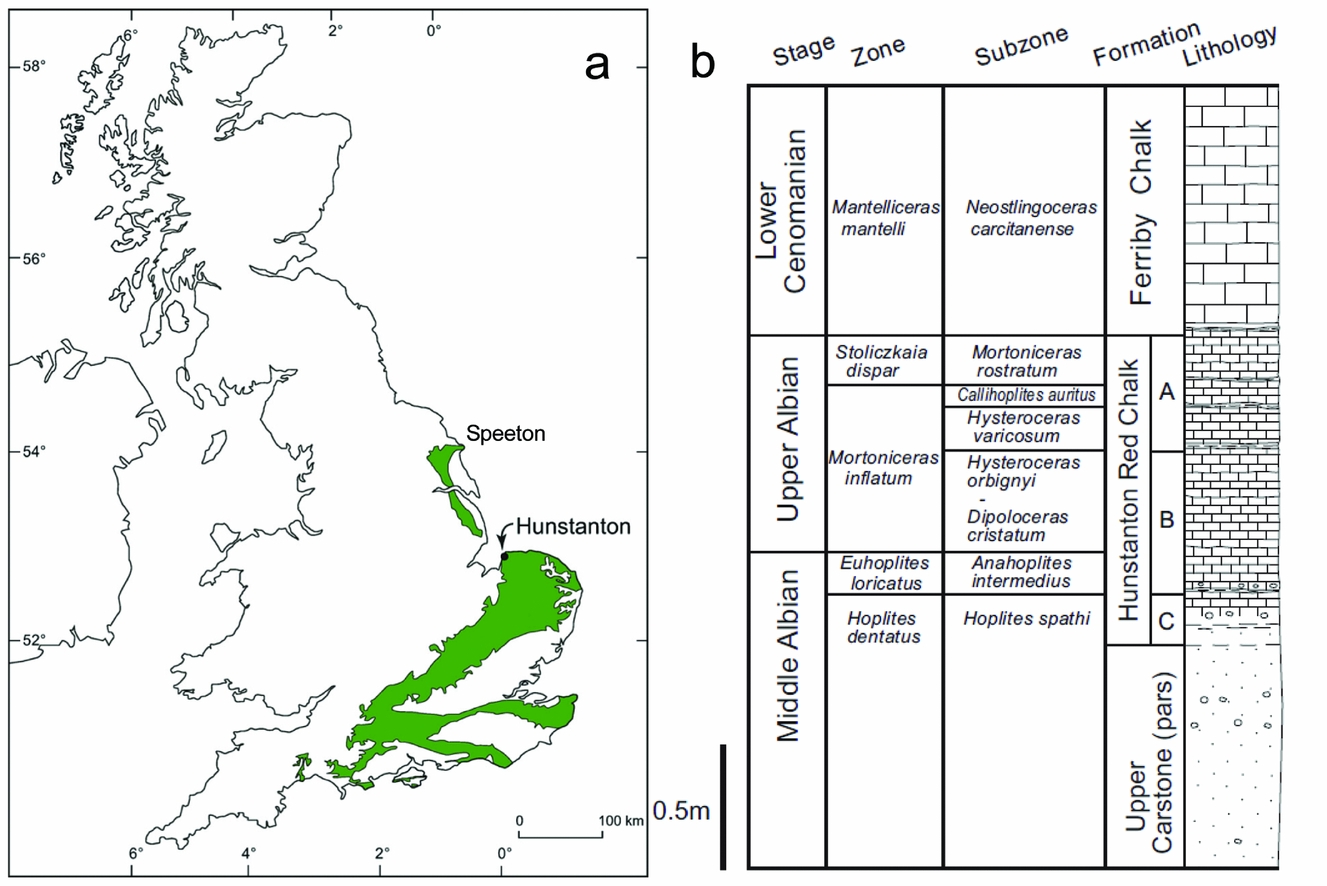
Onshore exposures of the Lower Cretaceous Hunstanton Red Chalk Formation extend over 100 km northwest from Hunstanton in Norfolk to Speeton in Yorkshire (Jeans, Reference Jeans1980; Owen, Reference Owen1995; Fig. 1). Offshore the formation and its correlatives extend across the central and southern North Sea as red marls and subordinate chalks (Rawson, Reference Rawson, Duff and Smith1992) and thus comprise a regionally significant (basin-scale) time-specific facies (Brett et al. Reference Brett, McLaughlin, Histon, Schindler and Ferretti2012). Here we concentrate on the contact between the Lower Cretaceous Hunstanton Red Chalk Formation and the overlying Upper Cretaceous Ferriby Chalk Formation (lithostratigraphic scheme of Owen, Reference Owen1995; Mortimore, Wood & Gallois, Reference Mortimore, Wood and Gallois2001) in the cliffs at the type locality, Hunstanton, NW Norfolk [TF 672 413, TF 679 424].
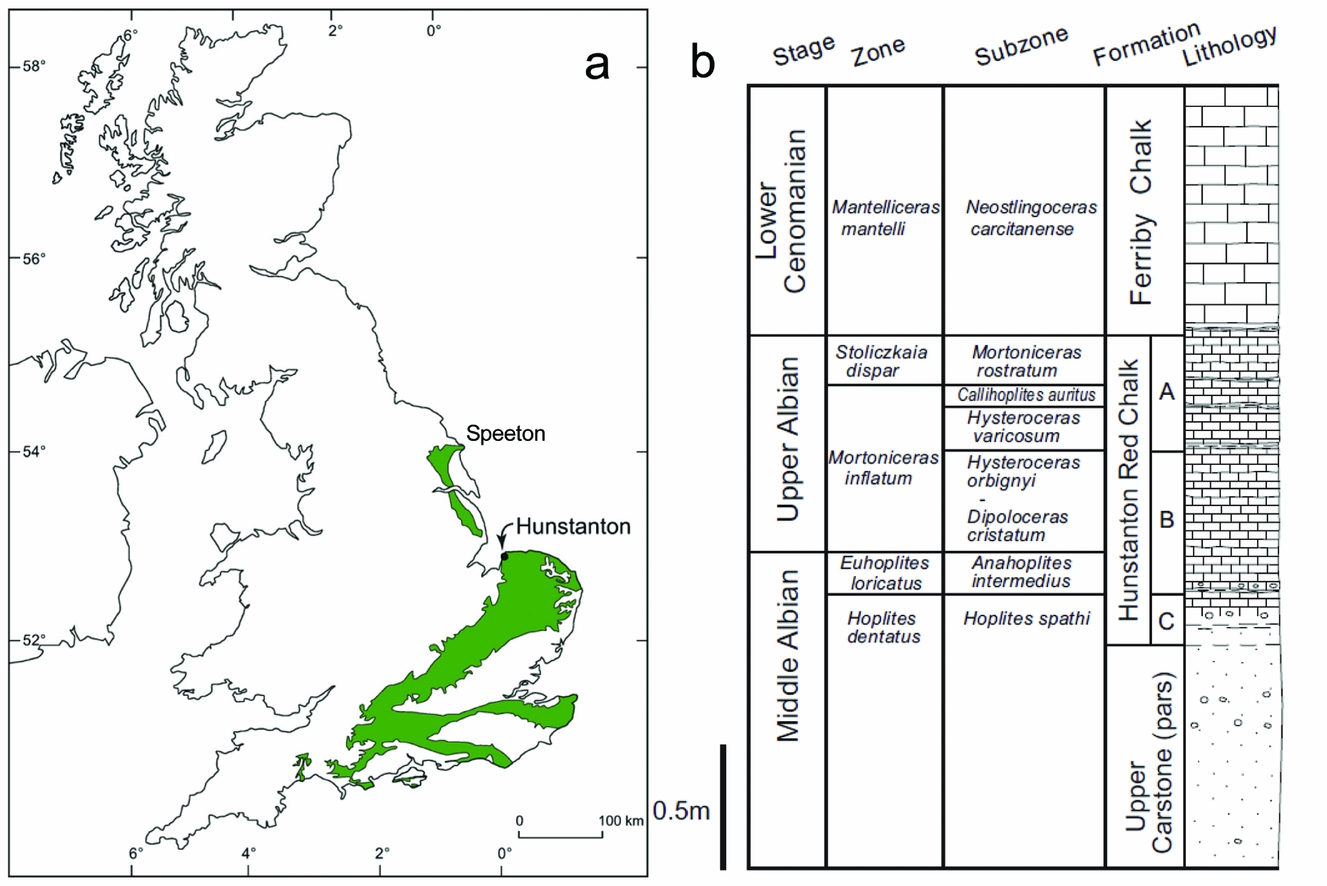
Figure 1. (a) Map showing the location of Hunstanton in the UK. The green area shows the Chalk outcrop (after Price & Harwood, Reference Price and Harwood2012). The Hunstanton Red Chalk Formation is present at the base of the Chalk outcrop from Hunstanton northwards to Speeton on the Yorkshire coast. (b) Summary stratigraphy of the cliff section at Hunstanton (after Price & Harwood, Reference Price and Harwood2012) showing the ammonite biostratigraphy of Owen (Reference Owen1995).
Viewed from a distance the boundary between the Lower and Upper Cretaceous rocks appears sharp, delineated by the marked colour change from red to white chalk. However, closer inspection reveals a complex micro-stratigraphy. The upper part of the Hunstanton Red Chalk Formation is composed of condensed nodular hardgrounds with eroded depressions and intensely bored chalk pebbles and surfaces with glauconitic staining. These are separated by thin layers and laminae of slightly more argillaceous chalks that contain closely spaced pressure-solution surfaces, along which the red colouration is concentrated. A distinctive uppermost crust is common in fallen blocks from the southern part of the cliffs, and this has been interpreted as an algal stromatolite(?) (Jeans, Reference Jeans1980). The crust is overlain by a thin, dark-red, earthy marl, which is intensely iron stained and locally may be replaced by iron oxides and hydroxides. Brick-red bulbous columnar structures, up to 5 cm high, arise from the marl and project into the basal Paradoxica Bed of the overlying Ferriby Chalk Formation (Andrews, Reference Andrews1983). These columns have been interpreted as stromatolites (Jeans, Reference Jeans1980), but also as burrows stained red by oxidized iron (Andrews, Reference Andrews1983).
2. Stratigraphy
The Chalk at Hunstanton, on the eastern shore of the Wash, represents the southernmost part of the Northern Province of the British Chalk (Mortimore, Wood & Gallois, Reference Mortimore, Wood and Gallois2001; Fig. 1). Although British Northern Province Chalks are generally considered deeper water deposits than equivalent-aged deposits to the south, most of the chalks at Hunstanton represent shallow-water chalk facies probably forming in water depths of 50–100 m (Jeans, Reference Jeans1980), being near the Anglo-Brabant massif margin.
The local Cretaceous stratigraphy at the study site (Fig. 1) begins with the Carstone Formation, a ferruginous sandstone with local conglomerates, representing shallow-marine (probably upper shoreface) sands and gravels. In places the Carstone is trough-cross-bedded and in its upper parts contains evidence of thalassinid burrows. It contains very few body fossils (Owen, Reference Owen1995), although equivalent rocks further north have yielded brachiopods, bivalves, belemnites, foraminifera and ostracods. The Upper Carstone Member is of Middle Albian age (Owen, Reference Owen1995) and has a gradational contact with the overlying Middle–Upper Albian aged Hunstanton Red Chalk Formation, the base of which is very sandy, but which becomes progressively more carbonate rich up section: the upper part is a very hard chalk. The Hunstanton Red Chalk Formation is essentially a nodular chalk in its upper part, differing from the overlying basal white chalks of the Cenomanian Ferriby Chalk Formation only by its distinctive colour. The facies progression is transgressive, representing a local response to the overall Barremian to Turonian eustatic sea-level rise (see e.g. Miller et al. Reference Miller, Kominz, Browning, Wright, Mountain, Katz, Sugarman, Cramer, Christie-Blick and Pekar2005).
The prominent red colour of the Hunstanton Red Chalk Formation is generally agreed to be from a finely divided detrital haematitic pigment, with iron mobilization and migration allowed by early diagenetic reducing conditions in the sediment (Jeans, Reference Jeans1980; Andrews, Reference Andrews1983). Bored surfaces and thalassinid-burrow systems are associated with glauconite and Liesegang structures, suggesting interface interactions between reducing sediment pore water and overlying oxidized seawater (Jeans, Reference Jeans1980). The red colour becomes generally less intense upwards as the sediment gets nodular and chalky. This may simply show that the iron is largely inherited from the underlying Carstone (Andrews, Reference Andrews1983) or that the basal sandy sediments stain more readily.
The stratigraphy of the Hunstanton Red Chalk Formation / Ferriby Chalk Formation has been debated for almost two centuries (reviewed by Owen, Reference Owen1995). The biostratigraphy is, however, much better understood now than in the early 1980s when the last two detailed studies on the sedimentology of the boundary were published (Jeans, Reference Jeans1980; Andrews, Reference Andrews1983); we outline the pertinent points here. First, it is now accepted that the upper beds of the Hunstanton Red Chalk Formation are of Upper Albian Age (i.e. to the top of unit A of Owen, Reference Owen1995; bed 5b of Gallois, Reference Gallois1994; and to the top of bed HE of Andrews, Reference Andrews1983), probably of the Callihoplites auritus ammonite Subzone (Owen, Reference Owen1995). The overlying dark-red earthy marl (Fig. 2; bed HF of Andrews, Reference Andrews1983) is typically 2.5 cm thick and has yielded the belemnite Neohibolites ultimus (d'Orbigny) and small terabratulid brachiopods that have affinity with basal Cenomanian brachiopods in Belgium (Gaunt, Fletcher & Wood, Reference Gaunt, Fletcher and Wood1992; Mortimore, Wood & Gallois, Reference Mortimore, Wood and Gallois2001). Offshore in the Central North Sea Basin this thin red seam thickens to 0.4 m of marly pink chalk with an early Cenomanian fauna (Gaunt, Fletcher & Wood, Reference Gaunt, Fletcher and Wood1992), and for this reason it is considered to be the basal bed of the Upper Cretaceous Ferriby Chalk Formation (following Owen, Reference Owen1995); the overlying Paradoxica Bed is considered to be of low (but not basal) Cenomanian age (Mortimore, Wood & Gallois, Reference Mortimore, Wood and Gallois2001). There are thus significant sedimentary gaps between the top of the Hunstanton Red Chalk Formation and the dark-red earthy marl (bed HF of Andrews, Reference Andrews1983), and between this marl and the overlying Paradoxica Bed. It is also significant that calcitic macrofossils have been found in the marl, as this discounts the otherwise tempting interpretation that this thin seam is a residue left from post-depositional groundwater dissolution of the overlying and underlying chalks.

Figure 2. Field view at the northern end of the cliff section showing the in situ contact between the Upper Albian Hunstanton Red Chalk Formation and the basal Paradoxica Bed of the overlying Cenomanian (white) Ferriby Chalk. The finger is pointing to the dark-red earthy marl (horizon F of Jeans, Reference Jeans1980, fig 12; bed HF of Andrews, Reference Andrews1983), which is considered to be of basal Cenomanian age. Note the red marl pinches and swells laterally, thickening in topographic lows. The notional position of the crust (see Fig. 3) is shown by the arrow, although it is not preserved at this location. Note the bulbous red nodules that project upwards into the base of the Paradoxica Bed.
The change from red to white chalk suggests that colouration of the Red Chalk was largely complete before deposition of the Paradoxica Bed. The change in lithology from chalk to red earthy marl, combined with the sedimentation break between deposition of the red earthy marl and the Paradoxica Bed facilitated this colour change as discussed further in Section 3b.
3. Stromatolite crusts and columns?
A prominent crust on the upper surface of the Red Chalk (Fig. 3) and centimetre-scale nodules and columns that project into the base of the Paradoxica Bed at Hunstanton (Fig. 2) have previously been interpreted as algal stromatolites(?) (Jeans, Reference Jeans1973, Reference Jeans1980), based principally on laminations seen in hand specimen and by comparison with modern algal stromatolites described elsewhere. Fine-scale laminae and cyanobacterial filaments were not observed in thin-section by Jeans (Reference Jeans1980): he attributed this to early diagenetic calcite precipitation in the sediment pores (type-2 lithification of Jeans, Reference Jeans1973, Reference Jeans1980) that was supposed to have obliterated the fine-scale laminations and presumably any traces of the microbial communities. This lithification was suggested to develop 5–10 mm below the upper surface of the microbially bound sediment, as an upward-moving alteration front, presumably keeping pace with sedimentation, and destroying microfabric as it passed through the sediment. We do not dispute here the mechanisms of early diagenetic chalk lithification proposed by Jeans (Reference Jeans1980). However, in this paper the algal stromatolite(?) interpretations are given further scrutiny, particularly as other workers have accepted these interpretations (Gallois & Morter, Reference Gallois and Morter1982; Rawson, Reference Rawson, Duff and Smith1992; Gallois, Reference Gallois1994; Mortimore, Wood & Gallois, Reference Mortimore, Wood and Gallois2001) apparently without further critical observations: some have also assumed that the stromatolite-constructing microbes were photosynthetic, implying Red Chalk deposition in the photic zone (Jeans, Reference Jeans1980; Gallois & Morter, Reference Gallois and Morter1982; Rawson, Reference Rawson, Duff and Smith1992), even in very shallow-water, possibly intertidal, conditions (Mortimore, Wood & Gallois, Reference Mortimore, Wood and Gallois2001).

Figure 3. (a) Cross-section view of the crust on the top surface of the Red Chalk (from a fallen block at the southern end of the cliff section), draping the underlying nodular topography. The base of the crust is coincident with the base of the scale bar. The crust shows indistinct lamination in the middle part of the image (arrow). Small black and white divisions on the scale bar are 1 cm2. (b) Plan view of the crust on the top surface of the Red Chalk (from a fallen block at the southern end of the cliff section) showing pustular top surface. The crust is best formed in topographic lows and pinches out against topographic highs of pale pink red chalk nodules. Camera case is 12 cm long. (c) Plan view of the crust on the top surface of the Red Chalk (from a fallen block at the southern end of the cliff section) showing irregular, polygonally arranged depressions. Small black and white divisions on the scale bar are 1 cm2.
3.a. Crust at the top of the Hunstanton Red Chalk Formation
3.a.1. Description
A prominent crust on the top surface of the Red Chalk is common on fallen blocks found at the southern 300 m of the cliff section. North of this position fallen blocks with well-preserved specimens of the crust are rare, but can be found occasionally up to ~30 m south of where the apparent dip brings the Albian–Cenomanian contact to eye level. Here the contact can be directly examined in the cliff face, although intrastratal slip, groundwater and salt weathering have altered the contact (Jeans, Reference Jeans1980) such that the crust cannot be observed in situ. Thus, along most of the northern exposure the crust is inferred to exist and probably also occurred where alteration and weathering has since obscured its existence. The crust is typically 0.5–3 cm thick, draping the underlying nodular topography (Fig. 3a): it is thickest in depressions and in places pinches out against topographic highs of bored, glauconitic, Red Chalk (Fig. 3b). In a few places oysters, belemnites, small terabratulid brachiopods and bored glauconite-coated chalk pebbles on the Red Chalk surface have been bound and entombed in the crust. The crust is composed of sandy and microfossil-rich, ocherous, micritic limestone with submillimetre-scale glauconitic laminae. Although the crust is not apparently strongly laminated at outcrop (Fig. 3a), it is clear on polished blocks that a submillimetre-scale lamination is well developed. The crust has a pustular top (Fig. 3b) with irregular, polygonally arranged, depressions (Fig. 3c). Our observations thus broadly concur with the descriptions given in Jeans (Reference Jeans1980, his fig. 12, horizon G).
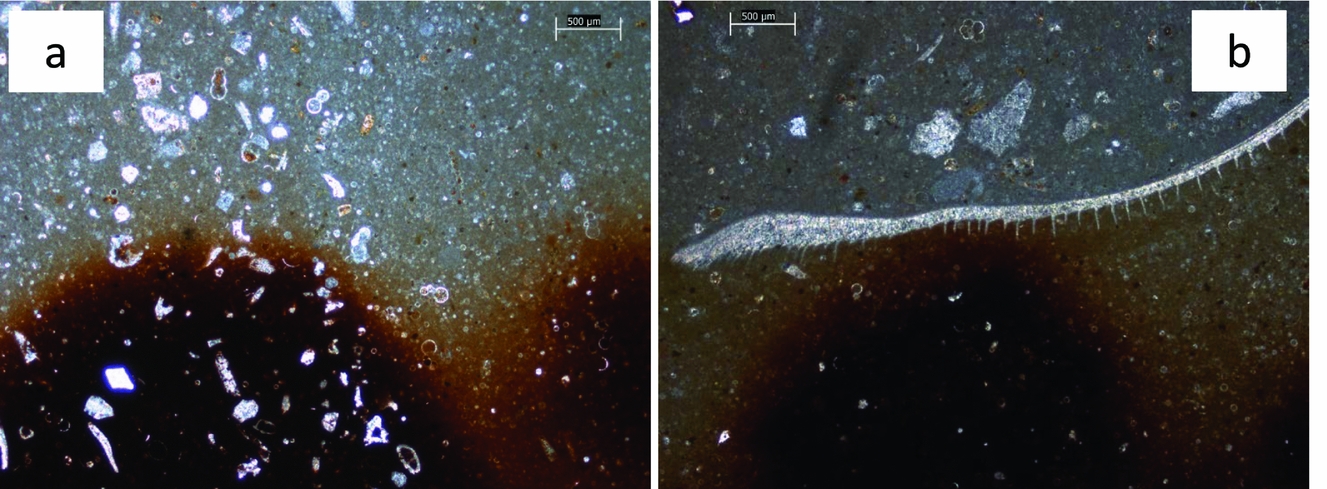
In thin-section the crust displays iron-rich microstructures (Fig. 4a), which give the crust a mottled appearance (Jeans, Reference Jeans1980, his fig. 13); two main elements are seen: (1) smoothly undulating, thin (10–20 μm), iron-rich laminae, most commonly developed at the base of the layer (Fig. 4a, b); and (2) closely spaced, essentially iron-rich vertical elements, 50–70 μm wide and 500 μm or more high, that repeatedly branch. Branches extend sideways and upwards, at angles less than 45° from the vertical elements, for distances of a few tens of micrometres (Fig. 4c). The vertical elements arise from the laminae.

Figure 4. Photomicrographs of cross-sections through the crust and the upper part of the Red Chalk substrate. The crust contains iron-rich microstructures. (a) Smoothly undulating, 10–20 μm thick iron-rich lamina are clear at the base of the crust (base indicated by white arrow), but also higher up. Vertical elements 50–70 μm wide and 500 μm or more high, that arise from the laminae, branch to form bush-like structures. Iron-rich laminae are also present in the upper part of the Red Chalk substrate. White scale bar = 1 mm. (b) Smoothly undulating, 10–20 μm thick iron-rich laminae are clear at the base of the crust. Vertical elements are heavily iron impregnated, such that bush forms are less clear than in (c). The crust base, shown by the white pecked line, drapes around micro-topography on the Red Chalk substrate. White scale bar = 500 μm. (c) Detail of the vertical elements arising from laminae, branching at angles less than 45° from the vertical elements to form bush-like structures. Some of the vertical elements, especially in the top right of the image, are mere upward projections from the laminae. White scale bar = 500 μm. (d) Detail of the vertical elements in (b), where original microstructure has been altered by the addition of iron oxides/hydroxides such that the branching vertical elements begin to merge. White scale bar = 100 μm
In some instances the vertical elements are mere upward projections from the laminae (Fig. 4c), their continued development having been terminated by a subsequent lamina. Where branching begins, the vertical elements thicken, so that the latter periodically vary in width vertically. Commonly the branching and thickening occur at the same levels in adjacent vertical elements, while occasionally the vertical elements terminate upwards and then it is the branches that appear to continue growing. These changes occur in adjacent vertical elements at the same distance above the base (Fig. 4a), which imparts the sense of layering seen in the crust: their differential development is also coincident with the irregularities of the upper surface, as illustrated by Jeans (Reference Jeans1980, his fig. 13). Typically, the original microstructure has been altered by the addition of iron oxides/hydroxides so that the branching vertical elements begin to merge together (Fig. 4d). Alternatively, in places iron mineralization is only weakly developed or has been leached out.
Spaces between these iron-rich microstructural elements are occupied by an iron-stained, micritic carbonate containing numerous foraminifera, calcispheres and quartz sand grains. This material is seemingly identical with the sediment that constitutes the uppermost Red Chalk upon which the crust lies.
The stable isotope composition of the crust, and associated Red Chalk components, were measured exactly as described in Woolhouse et al. (Reference Woolhouse, Andrews, Marca-Bell and Dennis2009); the data are shown in Table 1 and shown in context on Figure 5. The crust has bulk δ18O values of −2.6 to −3.3‰, values which are most likely indicative of chalk, slightly altered by meteoric diagenesis as discussed in the following Section.
Table 1. Stable isotope data

Repeated analyses of the laboratory standard (n = 14) gave a 2σ precision of ±0.09‰ for carbon and ±0.08‰ for oxygen. VPDB – Vienna Pee Dee Belemnite.
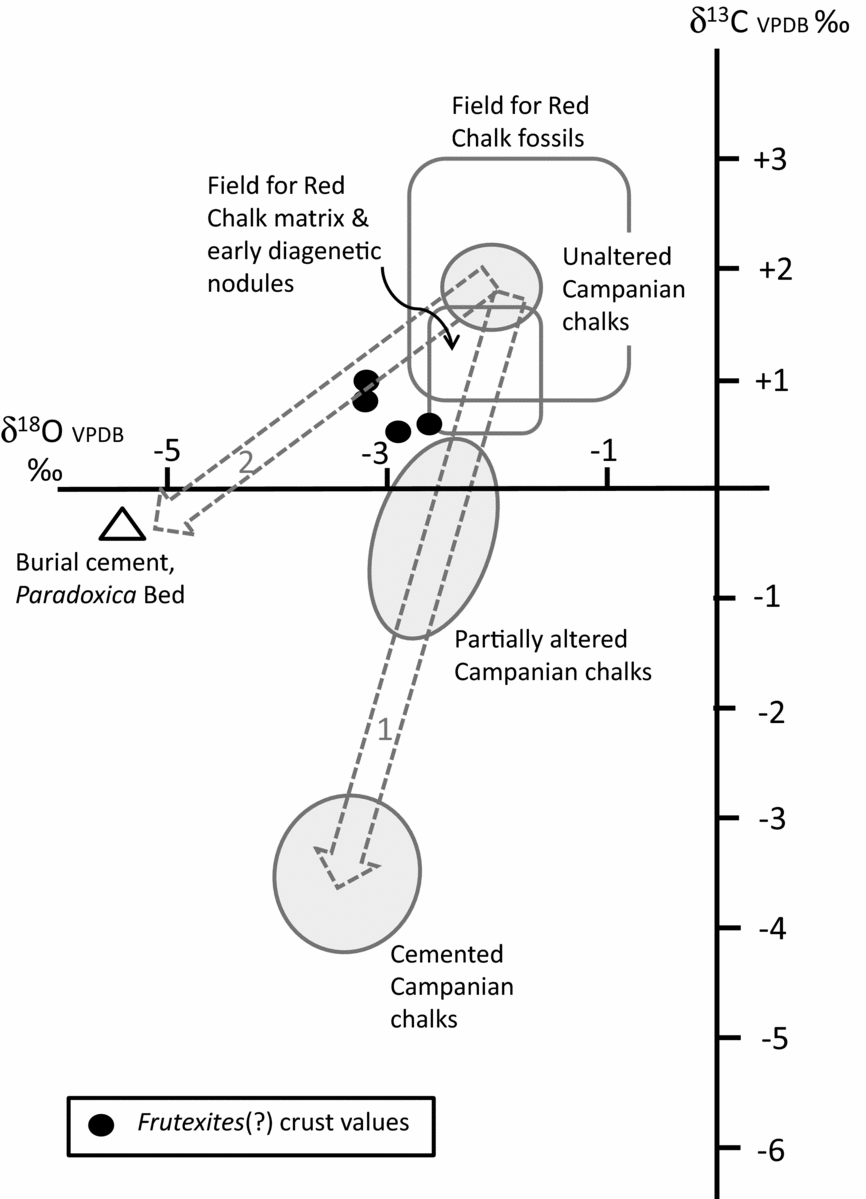
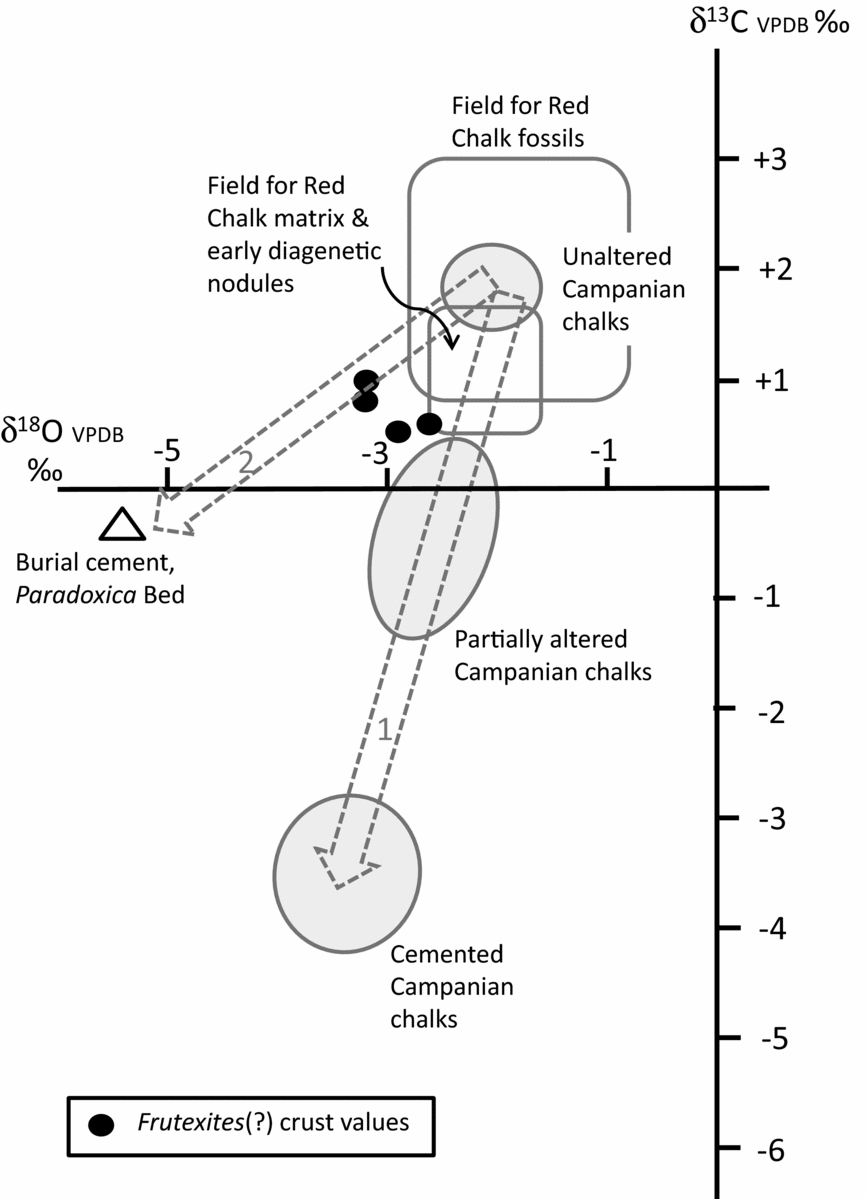
Figure 5. Cross plot of stable isotope data for the Hunstanton Red Chalk Formation and nearby Campanian Chalks in Norfolk. The Frutexites(?) crust data are shown in relation to various fields of data as follow: open boxes show fields for well-preserved Red Chalk fossils (belemnites and brachiopods) and for chalk matrix/early diagenetic nodules from data in Table 1 and that in Price & Harwood (Reference Price and Harwood2012); shaded ellipses show fields for unaltered, partially altered and cemented (altered) Campanian Norfolk chalks from Weybourne from data in Woolhouse et al. (Reference Woolhouse, Andrews, Marca-Bell and Dennis2009). The data from the Frutexites(?) crust are consistent with chalks that have an early (marine) diagenetic origin, possibly altered a little by later meteoric diagenesis. The two pecked arrows show likely pathways of alteration of isotope values: arrow 1 is based on the study of Woolhouse et al. (Reference Woolhouse, Andrews, Marca-Bell and Dennis2009) and arrow 2 is based on data in this study.
3.a.2. Interpretation
Overall the iron-rich microstructural elements show a marked resemblance to the fossil microbe(?) Frutexites as originally described by Maslov (Reference Maslov1960). However, compared with other Frutexites described and illustrated in the literature, the Red Chalk material is rather dominated by vertical elements with the branching more subdued. Frutexites elsewhere normally forms bush-like growths. Frutexites has been described from a variety of ancient shallow-marine (Myrow & Coniglio, Reference Myrow and Coniglio1991) to deep-marine (Bohm & Brachert, Reference Bohm and Brachert1993) environments. While most authors agree that Frutexites is a fossil microbe, there is considerable ambiguity about whether the microbes are dominantly bacterial or fungal and thus non-photosynthetic (Bohm & Brachert, Reference Bohm and Brachert1993) or cyanobacterial, and thus photosynthetic (Walter & Awramik, Reference Walter and Awramik1979).
Frutexites is usually described from two marine settings: (1) slowly accumulating and condensed, red nodular limestones (Playford et al. Reference Playford, Cockbain, Druce, Wray and Walter1976, Reference Playford, Mclaren, Orth, Gilmore and Goodfellow1984; Wendt, Reference Wendt1988; Hurley & Van der Voo, Reference Hurley and Van der Voo1990; Mamet & Préat, Reference Mamet and Préat2006; Reolid & Nieto, Reference Reolid and Nieto2010) like the Red Chalk, or (2) within cavities in mud mounds or reefs (e.g. Myrow & Coniglio, Reference Myrow and Coniglio1991; Reolid & Molina, Reference Reolid and Molina2010). In all cases the Frutexites growths are attributed to microbial fixation of iron in the depositional environment, typically in dysaerobic–anoxic conditions where chemotrophic bacteria might gain energy from iron oxidation (e.g. Préat et al. Reference Préat, Mamet, Ridder, Boulvain and Gillan2000). Mamet & Préat (Reference Mamet and Préat2006) in particular, attributed the red colour of condensed limestones, like those of the Red Chalk, as being due to the mediation of iron-fixing bacteria at a dysoxic–anoxic interface at the sediment–water interface, with Frutexites growths perhaps the more extreme manifestation of this.
It is possible that these Frutexites-like structures originally formed on the seafloor at the end of the Red Chalk depositional episode but as a non-iron-mineralized microbialite. This, at a later date, was then preferentially mineralized by the passage of iron-rich waters. Later mineralization would help explain the poor preservation of the material where mineralization extends beyond the original microstructures (Fig. 4d). It would also explain instances where mineralization is poorly developed or is non-existent.
The stable isotope data are helpful in determining the likely origin of the ‘Frutexites crust’. Well-preserved fossil belemnites and brachiopods from the Red Chalk at Hunstanton (Fig. 5) have δ18O values in the range −0.8 to −2.2‰ (Table 1; Price & Harwood, Reference Price and Harwood2012), while the nodular red chalk matrix and early diagenetic nodules (Fig. 5) have δ18O values in the range −2.2 to −2.5‰ (this study) and −1.5 to −2.3‰ (Price & Harwood, Reference Price and Harwood2012): the crust δ18O values are slightly more negative than the matrix/early diagenetic nodule values, suggesting a small amount of meteoric diagenesis affecting the crust values (trajectory 2, Fig. 5) and consistent with earlier work on British and Norfolk Chalk diagenesis (Scholle, Reference Scholle, Hsu and Jenkyns1974; Woolhouse et al. Reference Woolhouse, Andrews, Marca-Bell and Dennis2009). δ13C values of the crust range from +0.4 to +1.0‰, similar to data for nodular red chalks at Hunstanton (δ13C +0.6 to +1.7‰; this study; Price & Harwood, Reference Price and Harwood2012). Thus, although the crust δ18O data are somewhat altered from pristine depositional values, they are not negative enough to represent extensive meteoric water diagenesis, where δ18O values would be closer to −5 or −6‰ (based on burial diagenetic sparry calcite cement data from the Paradoxica Bed; Fig. 5). This information suggests strongly that the Frutexites crust was not a diagenetic microbial deposit associated with mineralizing diagenetic fluids, or resulting from groundwater flow along bedding surfaces. The isotope data are consistent with Albian seafloor deposition, the iron-rich microbialites being locally enhanced by subsequent mineralization or locally leached of original iron content by migrating diagenetic fluids which caused only limited alteration to δ18O values.
Overall, because a calcareous precursor cannot be totally excluded, because the Red Chalk microstructures are less bushy than Frutexites sensu stricto, and because of poor preservation, we identify these microbes as Frutexites(?).
3.b. Iron migration, columnar and nodular structures
Evidence for late (burial) diagenetic migration of iron by percolating waters is evident within both the Red Chalk below the crust and in the Paradoxica Bed of the Ferriby Chalk above the crust. Iron oxides/hydroxides are concentrated within solution seams between limestone nodules in the Red Chalk where clay minerals are preferentially concentrated, but later upward migration of iron is clearly observed above these seams as flame-like and nodular structures (Fig. 6a). In the base of the overlying Ferriby Chalk, late-stage iron migration is evident with the development of patchy iron staining and a faint pink body colour. Even today iron migration is occurring as shown by the formation of vertically oriented iron staining on surfaces of rotated Red Chalk blocks in the cliff talus.

Figure 6. Field views of iron oxide-stained chalks. (a) Iron oxides/hydroxides concentrated within seams (indicated by finger) between limestone nodules in the upper part of the Red Chalk (fallen block at the southern end of the cliff section). Later diagenetic upward migration of iron is clearly observed above these seams as flame-like and nodular structures. (b) Underside of a fallen block of the Paradoxica Bed. This lower bedding surface view shows the bulbous red nodules that project upwards into the Paradoxica Bed. Note the ‘inverted tear-drop’ shape, the apparently sharp margins to the nodules and the more diffuse pink staining of the white chalk matrix close to the nodules. Small black and white divisions on the scale bar are 1 cm2. (c) Underside of a fallen block of the Paradoxica Bed. This plan view of the lower bedding surface shows the bulbous red nodules, three of which have an initial ‘singular’ starting point that appears hollow. The nodules have sharp margins while the chalk matrix close to the nodules is stained a lighter pink colour. Finger nail is 1 cm wide. (d) Field view of a fallen block of the Paradoxica Bed. This view shows a sheet-like base of laminated red-stained chalk with bulbous red nodules projecting upwards. Thumb nail is 1.5 cm wide.
Although continued accretion of iron has occurred within the Frutexites(?) structures described above, it differs from that in the underlying and overlying chalks in being significantly more concentrated. In the adjacent chalks, migrated iron oxides/hydroxides typically have diffuse boundaries. It is also unlikely that the iron concentrations occur within a fracture fill. This is because (1) the matrix of the Frutexites(?) structures appears to be an undisturbed chalk identical to that immediately below, and (2) no other fractures in the Red Chalk or overlying white chalks possess Frutexites(?). On this basis we think the iron migration within the Frutexites(?) was syndepositional or early diagenetic.
Related to these discussions on iron migration are the enigmatic columnar and nodular structures (Fig. 2) that overlie the Frutexites(?) crust (south end of outcrop) and/or the thin, dark-red, earthy marl (middle and north end of the outcrop). The nodules project upwards (Fig. 2), into the basal Paradoxica Bed of the overlying Ferriby Chalk Formation (Andrews, Reference Andrews1983). These brick-red mini-columns and nodules may be discrete (Fig. 6b, c) or have a sheet-like base, only the latter having lamination in places (Fig. 6d). The nodules range in size from a few millimetres up to about 5 cm high: many have an inverted tear-drop shape, starting with a thin neck-like base and widening upwards to bulbous tops (Figs 2, 6b, c). Although Jeans (Reference Jeans1980, his fig. 12) seems to have illustrated these nodules with laminar fabric, there are no laminae visible, either in the field, or on polished blocks or in thin-section (Fig. 7a). The laminae in the sheet-like bases (Fig. 6d) are simply layers of heavier or lesser iron impregnation. In thin-section these red columns have identical fabric to the surrounding white chalk, containing foraminifera, calcispheres and detrital grains in a micritic matrix (Andrews, Reference Andrews1983); there is no trace of lamination and no evidence of microbial structures such as cyanobacterial filaments or Frutexites-like structures. Stable isotope values for the red nodules (mean δ18O −2.98; δ13C +0.78‰; Table 1) are not significantly different to the surrounding white chalk matrix (mean δ18O −3.12; δ13C +0.84‰; Table 1). The bulbous shape and generally diffuse boundaries of these nodules (Fig. 7a) are consistent with a migration front of soluble iron, which was stopped sharply only at physical barriers such as shell fragments (Fig. 7b). There is no evidence that these are microbial structures, rather they are patches of iron-stained chalk. It is also notable that the lower part of the Paradoxica Bed, up to a prominent solution seam, is largely stained pale pink, suggesting more widespread, although less intense, iron diffusion. Similar iron-stained structures ranging from columnar to flame shaped are also present in the largely white-coloured early diagenetic nodules of the upper 15–25 cm of the Hunstanton Red Chalk Formation (Fig. 6a).

Figure 7. Photomicrographs of the bulbous red nodules that project upwards into the Paradoxica Bed as shown in Figures 2, 6b, c. (a) Note the diffuse nodule margin where the colour fades from red-stained to white chalk. The matrix of the red-stained chalk is identical with the white chalk containing foraminifer tests, calcispheres and detrital grains. There is no evidence of lamination in the red-stained chalk. (b) Note the nodule margin is diffuse on the sides where the colour fades from red-stained to white chalk; however, the colour staining is terminated upwards by a shell which has arrested diffusion.
The reason for the positioning and initiation of these apparently randomly distributed nodules and columns is not clear. The dark-red earthy marl may have acted as an aquitard to upward flowing fluids and essentially acted like a ‘sponge’, absorbing almost all the upward migrating iron. The nodules might thus represent sites where the aquitard was disturbed or penetrated, probably during early (syndepositional) compaction, allowing iron to ‘bleed’ diffusively upwards and outwards from a discrete point source. There is no field evidence that the nodules are in any way linked to burrows as suggested by Andrews (Reference Andrews1983). The burrow networks in the Paradoxica Bed have glauconitic-stained walls (Jeans, Reference Jeans1980), but the chalk filling of the burrows is white, even where the host bed is stained red or pink. These observations suggest the burrows were open to seawater when the host chalk was stained, the back-filling of the burrows occurring largely after iron migration.
4. Summary and conclusions
A prominent stromatolitic crust found at the top of the Hunstanton Red Chalk Formation contains clear evidence of fossil microstructures attributable to the microbe(?) Frutexites. These Red Chalk microstructures are less bushy that Frutexites sensu stricto, and because of poor preservation, we identify them as Frutexites(?). Stable isotope data from the crust are consistent with Albian seafloor deposition, the iron-rich microbialites probably being locally enhanced by subsequent mineralization or locally leached of original iron content by migrating diagenetic fluids which caused limited alteration to Albian marine δ18O values. As the microbial affinities of Frutexites are not yet proven, it is prudent to simply caution that the presence of Frutexites alone is not diagnostic of either photic zone or deep-water sedimentation.
The presence of Frutexites(?) does, however, suggest that the red colour of the Hunstanton Red Chalk is due, at least in part, to the mediation of iron-fixing microbes at a dysoxic–anoxic interface at the Albian sediment–water interface. These red chalks are thus examples of regionally significant (basin-scale) time-specific facies (Brett et al. Reference Brett, McLaughlin, Histon, Schindler and Ferretti2012), with condensation caused by numerous high-frequency–low-amplitude sea-level fluctuations (Immenhauser & Scott, Reference Immenhauser and Scott1999) and associated disruption of the carbonate factory. These conditions induced prolonged sediment stability through low sedimentation rates and episodic scouring/erosion and stasis of sediment geochemical conditions suitable for early diagenetic nodular carbonate precipitation and microbial iron fixation.
Enigmatic columnar and nodular structures above the Frutexites(?) crust, that project upwards into the basal Paradoxica Bed of the overlying Ferriby Chalk Formation, are sites of localized iron staining probably formed during early (syndepositional) diagenesis. These nodules are not stromatolitic or microbial and are not evidence for deposition in shallow-water or intertidal settings.
Acknowledgements
We are very grateful to Robert Riding who advised us on the pitfalls of over interpreting the presence of Frutexites. Greg Price and Annalisa Ferretti provided very helpful reviews. Sarah Dennis, Alina Marca-Bell and Paul Dennis collectively worked on the isotope analyses reported here and Jennifer Mason allowed us to use her sparry calcite isotope data from the Paradoxica Bed. Stephen Bennett, Liz Rix and the late Angela Maher provided technical support in the laboratories. We dedicate this paper to Angela, recalling her sunny disposition while helping UEA's environmental science students and staff over many years.