


1. Introduction: From active perception to social cognition and beyond
Like many today, I view perception as inherently active (Hurley Reference Hurley1998; Hurley & Noë Reference Hurley and Noë2003, O'Regan & Noë Reference O'Regan and Noë2001a; Reference O'Regan and Noë2001b; Noë Reference Noë2004) and cognition as embodied and situated. How does cognition relate to active perception? This article shows how subpersonal resources for social cognition can be built on those for active perception. Its central issue is the following: How is it possible to perceive events as instrumentally structured intentional actions, and to learn new instrumental actions by means of such perceptions of actions? The article shows how the structures and mechanisms for perceiving action and for situated social cognition can be built on those for active perception. It extends my previous contributions to a view of perception as inherently active and of cognition as embodied and situated (Hurley Reference Hurley1998; Hurley & Noë Reference Hurley and Noë2003).
The classical sandwich conception of the mind – widespread across philosophy and empirical sciences of the mind #x2013; regards perception as input from world to mind, action as output from mind to world, and cognition as sandwiched between. I have argued that the mind isn't necessarily structured in this vertically modular way (Brooks Reference Brooks1999; Hurley Reference Hurley1998). Moreover, there is growing evidence that it is not actually so structured in specific domains, where perception and action share dynamic information-processing resources as embodied agents interact with their environments, rather than functioning as separate buffers around domain-general central cognition.
Rather, cognitive resources and structure can emerge, layer by layer, from informational dynamics, enabling both perception and action. Such a horizontally modular structure can do significant parts (I don't claim all) of the work the classical sandwich conception assigned to central cognition. Here I show how this promise can be fulfilled for the perception of action and associated social cognition, as embodied agents interact with their social environments.
I first review recent work on social cognition, focusing on imitation (Hurley Reference Hurley2005b; Hurley & Chater Reference Hurley and Chater2005a; Reference Hurley, Hurley and Chater2005b). Imitation is still popularly regarded as cognitively undemanding. However, Thorndike (Reference Thorndike1898) showed that many animals can learn through individual trial and error but not imitatively; scientists regard the later as more cognitively demanding. Imitative ability is rare across animal species and linked to characteristically human capacities: for language, culture, and understanding other minds (Arbib et al. (Reference Arbib, Billard, Iacoboni and Oztop2000); Arbib Reference Arbib2005; Arbib & Rizzolatti Reference Arbib and Rizzolatti1997; Barkley Reference Barkley2001; Frith & Wolpert Reference Frith and Wolpert2004; Gallese Reference Gallese2000; Reference Gallese2001; Reference Gallese, Hurley and Chater2005; Gallese & Goldman Reference Gallese and Goldman1998; Gallese et al. Reference Gallese, Keysers and Rizzolatti2004; Gordon Reference Gordon1995b; Iacoboni Reference Iacoboni, Hurley and Chater2005; Meltzoff Reference Meltzoff, Hurley and Chater2005; Rizzolatti & Arbib Reference Rizzolatti and Arbib1998; Reference Rizzolatti and Arbib1999; Stamenov & Gallese Reference Stamenov and Gallese2002; Tomasello 1999; Whiten et al. Reference Whiten, Horner, Marshall-Pescini, Hurley and Chater2005b; Williams et al. Reference Williams, Whiten, Suddendorf and Perrett2001). Imitation is important in adult human sociality, as well as human development, in ways we're just beginning to understand.
Part 1 of this article reviews recent research on imitation, under the headings of behavior, subpersonal mechanisms, and functions. Part 2 presents a functional architecture that shows how subpersonal mechanisms of control, mirroring, and simulation can enable distinctively human skills of imitation, deliberation, and action understanding. The shared circuits model (SCM) draws together many threads of work from Part 1. It includes elements suggested by various researchers, contributes further elements, and unifies these in a distinctive framework. SCM aims to show how the following are possible:
1 Building subpersonal informational resources for situated social cognition on those for active perception, while
2 Uniting a large body of evidence and theorizing in a common framework;
3 Avoiding the “classical sandwich”; and
4 Respecting the personal/subpersonal distinction and avoiding interlevel isomorphism assumptions.
Philosophers distinguish descriptions of contentful actions and mental states of persons from subpersonal (informational or neural) descriptions (Bermudez 2000; 2003; Dennett Reference Dennett1969; Reference Dennett1991; Elton Reference Elton2000; Hornsby Reference Hornsby2000; McDowell Reference McDowell1994). At the subpersonal level of description, information is processed and the cycling of causes and effects knits actively embodied nervous systems into environments they interact with.Footnote 1 But these processes are not correctly attributed to persons. Persons see trees, make friends, look through microscopes, vote, want to be millionaires. Subpersonal informational and causal theories explain how personal-level phenomena become possible – are enabled – but need not share structure with personal-level descriptions of processes as rational or conscious.Footnote 2 I distinguish three levels of description: the personal level, the informational/functional subpersonal level, and the neural subpersonal level.
Two questions arise about personal/subpersonal relations: (1) How are specific personal-level capacities actually enabled by subpersonal processes? (2) What kinds of subpersonal processes could possibly do the enabling work? For example, must there be isomorphism between levels? Views about question 2 can influence answers to question 1.
SCM addresses question 2 for social cognition by using subpersonal resources from an active perception approach. SCM is cast at the subpersonal functional level, not the personal or the neural levels, though it aims both to show how certain personal-level capacities can be informationally enabled and to raise empirical questions about neural implementation. Since SCM addresses the “how possibly?” rather than the “how actually?” question, it provides a higher-order theoretical model. But it also provides generic heuristic resources for framing specific first-order hypotheses and predictions about specific ontogenetic or phylogenetic stages. Its five layers, detailed below, can be re-ordered in formulating specific first-order hypotheses.
SCM's central hypothesis is that associations underwriting predictive simulation of effects of an agent's own movement, for instrumental control functions, can also yield mirroring and “reverse” simulation of similar perceived movements by others. Mirroring allows ends/means associations with instrumental control functions to be accessed for simulative functions bilaterally, so causes of observed movements can be simulated, as well as effects of intended acts. Such bilaterally accessible simulations of instrumental structure can provide enabling information for deliberation, imitation, and understanding the instrumental acts of others. Shared dynamics for action and perception can provide the foundations of shared dynamics for self and other, and of the self/other and actual/possible distinctions characteristic of human cognition.
Simulation has a generic sense throughout, including, but broader than, that in simulationist theories of mindreading (Gallese Reference Gallese2003; see Goldman Reference Goldman1989; Reference Goldman1992 on process-driven simulation). Simulation uses certain processes to generate related information, rather than theorizing about them in separate meta-processes. Effects or causes can be simulated, online or offline. Simulation can be subpersonal or personal; in SCM it is subpersonal. Subpersonal processes that predict results of movement online can also generate information about results of possible movements offline. Subpersonal mirroring that enables copying can also generate information covertly about observed movements or their goals, without overt copying (cf. Barkley [2001] on executive functions as covert behavior).
2. Part 1. Review
I begin in Part 1 by reviewing recent work, strands of which are knitted together by SCM in Part 2.
2.1. Behavior
Imitative learning is a sophisticated form of social cognition. It requires copying in a generic sense: Perception of behavior causes similar behavior by an observer, and the similarity plays a role – not necessarily consciously – in generating the observer's behavior. True imitation, restrictively understood, requires novel action learned by observing another do it, plus instrumental or means/ends structure: the other's means of achieving her goal is copied, not just her goal or just her movements. The concept of true imitation is contested, given the different aims and methodologies of imitation researchers (Byrne Reference Byrne, Hurley and Chater2005; Heyes Reference Heyes, Heyes and Galef1996; Reference Heyes2001; Rizzolatti Reference Rizzolatti, Hurley and Chater2005).
Other forms of social learning can seem similar to imitation, but should be distinguished. In stimulus enhancement, another's action draws your attention to a stimulus, which triggers an innate or previously learned response; but a novel action isn't learned directly from observation. Bird A's pecking may draw bird B's attention to a food, which evokes pecking in bird B. In goal emulation,Footnote 3 you observe another achieving a goal by certain means, find that goal attractive, and try to achieve it yourself. Monkey A may use a tool in a certain way to obtain an attractive object, leading monkey B to acquire the goal of obtaining a similar object. Through his own trials and errors, monkey B may arrive at the same type of tool use to obtain the object. Emulation is found in macaques, who have not shown imitative learning. In movement priming, bodily movements are copied, but not as learned means to a goal. Primed movements can be innate, as in contagious yawning.
Goal emulation and movement priming provide the ends-and-means components of full-fledged imitation. Ends and means can be relatively distal or proximal; the distinction is relative, not absolute. Misunderstandings can result concerning whether ends, means, or both are copied and hence whether imitation or emulation is present (Voelkl & Huber Reference Voelkl and Huber2000, pp. 196, 201). A movement can be the proximal means to a bodily posture, which is regarded as the proximal end of the movement (Graziano et al. Reference Graziano, Taylor, Moore and Cooke2002, pp. 354–55), but posture can also be a means to more distal ends – effects on objects or others in social groups. Complex imitation can involve structured sequences or hierarchies of ends–means relations: acquiring a goal, learning to achieve it via subgoals, and so on.
How are these forms of copying distributed across animals, children, and adults? Stimulus enhancement, goal emulation, and movement priming are certainly found in nonhuman animals. But careful experiments are needed to distinguish these from imitation proper and obtain evidence of the latter. The two-action paradigm has been the tool of choice. Suppose two demonstrators obtain an attractive result by two different means: One group of animals observes one demonstrator, and the other observes the other demonstrator. Will the observer animals tend differentially to copy the specific method they have seen demonstrated? If not – if the animals' choices of method do not reflect the specific method they have observed, say, because both groups converge on one method – they may be displaying stimulus enhancement or goal emulation plus trial-and-error learning, but not imitative learning. Even if they do differentially display the behavior demonstrated, this may be merely movement priming if the behavior is already in their repertoire. But if the behavior is differentially used in a new way to achieve a result, it expresses imitative learning (Call & Tomasello Reference Call and Tomasello1994; Nagell et al. Reference Nagell, Olguin and Tomasello1993; Voelkl & Huber Reference Voelkl and Huber2000).
The difference between copying ends and copying means is important for theorizing the phylogeny of imitation and action understanding. (Action understanding is short for understanding observed behavior as goal-directed action.) Is action understanding phylogenetically prior to imitation? This view seems to face an objection: Some animals copy movements (schooling fish), though we don't think they understand the others' actions. A response to this objection distinguishes movement copying from mirroring of goals (Rizzolatti Reference Rizzolatti, Hurley and Chater2005) and both from imitation. Movement copying may precede action understanding, whereas action understanding may require goal mirroring, but precede imitation. True imitation involves something phylogenetically rare: the flexible interplay of copying ends and copying means; a given movement can be used for different ends and a given end pursued by various means (Barkley Reference Barkley2001, p. 8; Tomasello 1999). This is something humans are distinctively good at.
It is difficult to find evidence of true imitation in nonhuman animals (Byrne Reference Byrne1995; Galef Reference Galef, Zentall and Galef1988; Reference Galef, Sabourin, Craik and Roberts1998; Reference Galef, Hurley and Chater2005; Heyes & Galef Reference Heyes and Galef1996; Tomasello Reference Tomasello, Heyes and Galef1996; Tomasello & Call Reference Tomasello and Call1997; Voelkl & Huber Reference Voelkl and Huber2000; Zentall Reference Zentall2001). Early work with chimps seems to reveal imitation, but critics have challenged this interpretation effectively; the results of subsequent experiments were negative for chimp imitation. Skeptics about nonhuman imitation long had the upper hand; for example, Tomasello et al. (Reference Tomasello, Kruger and Ratner1993) found no convincing evidence of nonhuman imitative learning. They proposed that understanding behavior as intentional distinguishes human from nonhuman social learning. On this view, humans can imitate observed means or choose other means to emulate observed goals. Other animals don't distinguish means and goals this way; rather, they copy movements without understanding their relevance to goals, or learn about the affordances of objects by observing action on them. In neither case, it was claimed, do other animals learn about the intentional, means/end structure of observed action.
Many skeptics have now been won over by work on imitation in great apes and monkeys (Voelkl & Huber Reference Voelkl and Huber2000; Whiten et al. Reference Whiten, Horner, Marshall-Pescini, Hurley and Chater2005b), dolphins (Herman Reference Herman, Dautenhahn and Nehaniv2002), and birds (Akins & Zentall Reference Akins and Zentall1996; Reference Akins and Zentall1998; Akins et al. Reference Akins, Klein and Zentall2002; Hunt & Gray Reference Hunt and Gray2003; Pepperberg Reference Pepperberg1999; Reference Pepperberg, Dautenhahn and Nehaniv2002; Reference Pepperberg, Hurley and Chater2005; Weir et al. Reference Weir, Chappell and Kacelnik2002). Continuities are described along a spectrum from the capacities of other social animals to human socio-cognitive capacities (Arbib Reference Arbib2005; Tomasello 1999). For example, innovative experiments extend the two-action method by using “artificial fruits” that can be opened in different ways to obtain a treat: Chimps tend to imitate for one aspect of a demonstrated task and emulate for another aspect, whereas children tend to imitate both aspects, even when the method imitated is inefficient. These and other experiments suggest that chimps imitate more selectively than children (Whiten Reference Whiten, Dautenhahn and Nehaniv2002; Whiten et al. Reference Whiten, Custance, Gomez, Teixidor and Bard1996; Reference Whiten, Horner, Marshall-Pescini, Hurley and Chater2005b; see also Call & Tomasello Reference Call and Tomasello1994; Galef Reference Galef, Hurley and Chater2005; Harris & Want Reference Harris, Want, Hurley and Chater2005; Heyes Reference Heyes1998; Nagell et al. Reference Nagell, Olguin and Tomasello1993; Tomasello & Carpenter Reference Tomasello, Carpenter, Hurley and Chater2005).
Children have been called “imitation machines” (Tomasello 1999, p. 159). They don't always imitate unselectively and sometimes emulate goals instead (Gergely et al. Reference Gergely, Bekkering and Király2002), but they have a greater tendency than chimps to imitate rather than emulate when the method demonstrated is transparently inefficient or futile (Tomasello 1999, pp. 29–30). After seeing a demonstrator use a rake inefficiently, prongs down, to pull in a treat, 2-year-old children do the same; they almost never turn the rake over to use more efficiently, edge down. By contrast, chimps given a parallel demonstration, tend to try turning the rake over (Nagell et al. Reference Nagell, Olguin and Tomasello1993).Footnote 4 The differential tendency of children and chimps to imitate suggests an interplay of biological and cultural influences, with a role for innate endowment enabling human imitation (perhaps a matter of articulated relations among multiple mirror subsystems, enabling recombinant structure in social learning, rather than the presence versus absence of a mirror system at all; see discussion of mechanisms in sect. 2.2).
Imitative and related behaviors appear throughout human development (Meltzoff Reference Meltzoff1988a; Reference Meltzoff, Cicchetti and Beeghly1990; Reference Meltzoff1995; Reference Meltzoff, Heyes and Galef1996; Reference Meltzoff, Meltzoff and Prinz2002a; Reference Meltzoff and Goswami2002b; Reference Meltzoff, Hurley and Chater2005; Meltzoff & Moore Reference Meltzoff and Moore1977; 1983a; 1983b; 1989; 1997; 1999; 2000). Infants younger than 1 month of age appear to copy facial gestures. By 14 months, infants imitate a novel act a week later: They turn on a light by touching a touch-sensitive panel with their forehead instead of their hand, differentially copying the novel means demonstrated, as well as the result (Meltzoff Reference Meltzoff1988a; Reference Meltzoff, Hurley and Chater2005; cf. Gergely et al. Reference Gergely, Bekkering and Király2002). They don't turn the light on in this odd way unless they have seen it demonstrated. By 15 to 18 months, infants recognize the underlying goal of an unsuccessful act they observe, and produce it: After seeing an adult try but fail to pull a dumbbell apart in her hands, they succeed in pulling it apart using knees and hands. But they don't pick up goals from failed “attempts” involving similar movements by inanimate devices, thus apparently discriminating agents from non-agents (Meltzoff Reference Meltzoff1988a; Reference Meltzoff1995; Reference Meltzoff, Heyes and Galef1996; Reference Meltzoff, Hurley and Chater2005; Meltzoff & Moore Reference Meltzoff and Moore1977; 1999; Tomasello & Carpenter Reference Tomasello, Carpenter, Hurley and Chater2005). Children's perception of behavior tends to be enacted automatically in similar behavior, unless actively inhibited; but frontal inhibitory functions are not well developed in young children (Barkley Reference Barkley2001, pp. 5, 22; Kinsbourne Reference Kinsbourne, Hurley and Chater2005; Preston & de Waal Reference Preston and de Waal2002, p. 5).
Adult “imitation syndrome” patients with frontal brain lesions also imitate uninhibitedly (Barkley Reference Barkley2001, p. 15; Frith Reference Frith1992, pp. 85–86; Lhermitte Reference Lhermitte1983; Reference Lhermitte1986; Lhermitte et al. Reference Lhermitte, Pillon and Serdaru1986, p. 330). They persistently copy the experimenter's gestures, though not asked to, even when these are socially unacceptable or odd, such as putting on eyeglasses when already wearing glasses.
But the human copying tendency isn't confined to the young or brain damaged! Normal adults can usually inhibit overt imitation selectively, which is evidently adaptive, but their underlying tendency to copy is readily revealed or released. Overt imitation is the disinhibited tip of the iceberg of continual covert imitation (Barkley Reference Barkley2001; Dijksterhuis Reference Dijksterhuis, Hurley and Chater2005). Experiments show how action is modulated or induced by perception of similar action (Brass et al. Reference Brass, Bekkering and Prinz2001; Prinz Reference Prinz, Meltzoff and Prinz2002). Imitative tasks have shorter reaction times than nonimitative tasks; gestures are faster when participants are primed by perceiving similar gestures or their results or goals – even when primes are logically irrelevant to the task (W. Prinz Reference Prinz, Hurley and Chater2005). Similarity between stimulus and response also affects which response is made. Normal adults, instructed to point to their nose when they hear “Nose!” and to a lamp when they hear “Lamp,” performed perfectly while watching the experimenter demonstrate the required performance, but made mistakes when watching the experimenter doing something else: they tended to copy what they saw done rather than to follow instructions (Eidelberg Reference Eidelberg1929; Prinz Reference Prinz, Neumann and Prinz1990). Movements can be induced by actions you actually perceive or by actions you would like to perceive – as when moviegoers or sports fans in their seats make movements they would like to see (W. Prinz Reference Prinz, Hurley and Chater2005). Visually or verbally represented, as well as observed, actions can induce similar actions.
It is helpful to distinguish copying of specific behaviors from chameleon effects, where complex patterns of behavior are induced – a relevant kind of copying, if not strict imitation. In an experiment involving specific behaviors, when normal adults interact in an unrelated task with someone rubbing her foot, they rub their own feet significantly more. Transferred to another partner who touches his face, they touch their own faces instead. Demonstrations of chameleon effects show that exposure to traits and stereotypes automatically elicits general patterns of behavior and attitude and influences how behavior is performed (Bargh Reference Bargh1999; Reference Bargh, Hassin, Uleman and Bargh2005; Bargh & Chartrand Reference Bargh and Chartrand1999; Bargh et al. Reference Bargh, Chen and Burrows1996; Reference Bargh, Gollwitzer, Lee-Chai, Barndollar and Trötschel2001; Chartrand & Bargh Reference Chartrand and Bargh1999; Chartrand et al. Reference Chartrand, Maddux, Lakin, Hassin, Uleman and Bargh2005; Dijksterhuis & Bargh Reference Dijksterhuis and Bargh2001). Normal adults primed with stimuli associated with traits (e.g., hostility, rudeness, politeness) or stereotypes (e.g., elderly persons, college professors, soccer hooligans) tend to behave in accordance with the primed traits or stereotypes. For example, hostility-primed participants deliver more intense “shocks” than control participants in subsequent, ostensibly unrelated experiments based on Milgram's (1963) classic shock experiments. Priming can also affect intellectual performance: College professor–primed participants perform better and soccer hooligan–primed participants perform worse than controls on a subsequent, ostensibly unrelated general knowledge test (Dijksterhuis Reference Dijksterhuis, Hurley and Chater2005; Dijksterhuis & van Knippenberg 1998).
Such priming results are robust across a wide range of verbal and visual primes and induced behavior, using dozens of stereotypes and general traits, and various priming methods, including primes perceived consciously and subliminally. Whether subjects perceive primes consciously or not, they are unaware of any influence or correlation between primes and their behavior. These influences are rapid, automatic, and unconscious, apply both to goals and means, and don't depend on subjects' volition or having independent goals that would rationalize their primed behavior.
Copying, at various levels of generality, is thus a default social behavior for normal human adults; it requires specific overriding or inhibition (Barkley Reference Barkley2001, p. 22; Dijksterhuis Reference Dijksterhuis, Hurley and Chater2005; Preston & de Waal Reference Preston and de Waal2002). Just thinking about or perceiving action automatically increases, in ways participants are unaware of, the likelihood that they will perform similar actions themselves. Nevertheless, these influences are often inhibited, as when goals make conflicting demands: elderly–primed participants tend to walk more slowly, but not if they have independent reasons to hurry.
2.2. Mechanisms
Copying perceived behavior seems to pose a correspondence problem (Nehaniv & Dautenhahn Reference Nehaniv, Dautenhahn, Dautenhahn and Nehaniv2002): How is another's observed action translated into the observer's similar performance? When I copy your hand movements, I can see my own hands, though my visual perspectives on the two movements are different. But when I copy your facial expressions, I cannot see my own face. What information and mechanisms are needed to map perception to similar behavior?
Evidence that newborns copy facial gestures, though they cannot see their own faces, suggests innate supramodal correspondences between action and perception of similar action (Meltzoff Reference Meltzoff1988a; Reference Meltzoff, Cicchetti and Beeghly1990; Reference Meltzoff1995; Reference Meltzoff, Heyes and Galef1996; Reference Meltzoff, Meltzoff and Prinz2002a; Reference Meltzoff and Goswami2002b; Reference Meltzoff, Hurley and Chater2005; Meltzoff & Moore Reference Meltzoff and Moore1977; 1983a; 1983b; 1989; 1997; 1999; 2000). Although further correspondences could be acquired as imitative abilities develop, skeptics about newborn copying can also be skeptical about the need to postulate any innate correspondences (Anisfeld Reference Anisfeld1979; Reference Anisfeld1984; Reference Anisfeld1991; Reference Anisfeld1996; Reference Anisfeld, Hurley and Chater2005; Anisfeld et al. Reference Anisfeld, Turkewitz, Rose, Rosenberg, Sheiber, Couturier-Fagan, Ger and Sommer2001; Heyes Reference Heyes, Hurley and Chater2005).
Heyes, who is one such skeptic, argues that sensorimotor associations subserving copying can be acquired through general-purpose associative learning mechanisms whereby neurons that fire together wire together. Direct sensorimotor associations between motor output and sensory feedback could result from watching one's own hand gestures. An indirect route is needed when the agent cannot perceive her own actions, as in facial expressions: The sensorimotor association could be mediated by environmental items such as mirrors, action words, or stimuli that evoke similar behavior in the actor and in other agents the actor observes. Moreover, adults commonly copy infants, performing the associative function of a mirror. When baby smiles and father smiles back, baby's motor output is associated with sensory input from father's smile (Heyes Reference Heyes, Hurley and Chater2005, p. 161; Preston & de Waal Reference Preston and de Waal2002, p. 8). Imitation can thus develop from interactions between organisms with associative learning mechanisms and certain cultural environments (see Heyes Reference Heyes2001; Reference Heyes, Hurley and Chater2005; see also SCM, Layer 3 in sect. 3.3 here).
Common coding for perception and action has been postulated to explain human copying tendencies. On this view, perception and action share subpersonal processes carrying information about (“coding for”) what is perceived or intended, in which perception and action are not distinguished. The differentiation between perception and action is overlaid on those shared resources, so that they are informationally interdependent at a basic level. If capacities X and Y share an information space, their commonality is informationally prior to their differentiation.
Meltzoff and Moore (1997) postulate common coding of perception and action in explaining infant imitation: proprioceptive feedback is compared to an observed target act, where these are coded in common, supramodal terms. Innate common coding could initially be for relations among, say, lips and tongue; more dynamic, complex, and abstract common coding could develop with experience of body babbling. Common coding might also be acquired as in Heyes's (2005) model.
Wolfgang Prinz (Reference Prinz, Prinz and Sanders1984; Reference Prinz, Hurley and Chater2005; cf. Bekkering & Wohlschläger (Reference Bekkering, Wohlschläger, Prinz and Hommel2002); Preston & de Waal Reference Preston and de Waal2002, pp. 4ff., 9–10) appeals to common coding of perception and action to explain the normal adult tendency to imitate and the reaction-time advantage of imitative tasks. Common coding facilitates imitation, avoiding the correspondence problem and any need for translation between unrelated input and output codes to solve it. Prinz associates common coding with what William James called ideomotor theory, on which every representation of movement awakes in some degree the movement it represents (Brass Reference Brass1999; Prinz Reference Prinz, Heuer and Sanders1987). Perceiving another's observed movement tends inherently to produce similar movement by the observer, and primes similar movement even when it doesn't break through overtly. Regular concurrence of action with perceived effects allows prediction of an action's effects and selection of action, given an intention to produce certain effects (Greenwald Reference Greenwald1970; Reference Greenwald1972). Thus, representation of an action's regular result, whether proximal or distal, can evoke similar action, in the absence of inhibition.
Other sources also support the view that perception and action share processing resources. Observing an action primes the very muscles needed to perform the same action (Craighero et al. Reference Craighero, Buccino and Rizzolatti2002; Fadiga et al. Reference Fadiga, Fogassi, Pavesi and Rizzolatti1995; Reference Fadiga, Craighero, Buccino and Rizzolatti2002). Watching an action sequence speeds the observer's performance of that sequence; merely imagining a skilled performance, in sport or music, improves performance – is a way of practicing – as many athletes and musicians know (Jeannerod Reference Jeannerod1997, pp. 117, 119–22; Pascual-Leone Reference Pascual-Leone2001). Similar points concern perception and experience of emotion: Gordon argues that a special containing mechanism, which isn't fail-safe, is needed to keep emotion recognition from producing emotional contagion. On his simulationist theory, only a thin line separates one's own mental life from one's representation of another's; offline representations of others tend inherently to go online (Gordon Reference Gordon1995b; cf. Adolphs Reference Adolphs2002; Preston & de Waal Reference Preston and de Waal2002).
Common coding theories characterize subpersonal architectures for copying functionally. What neural processes might implement such functional architectures?
Certain neurons directly link perception and action: their firing correlates with specific perceptions and specific actions. Canonical neurons (Gallese Reference Gallese, Hurley and Chater2005; Rizzolatti Reference Rizzolatti, Hurley and Chater2005) reflect affordances (Iacoboni Reference Iacoboni, Hurley and Chater2005; Miall Reference Miall2003): They fire when an animal perceives an object that affords a certain type of action and when the animal performs the afforded action. Mirror neurons fire when an animal perceives another agent performing a type of action, and also when the animal performs that type of action itself; they don't distinguish own action from others' similar actions (see SCM, Layer 3 in sect. 3.3). Some fire, for example, when a monkey sees the experimenter bring food to her own mouth with her hand or when the monkey brings food to his own mouth with his hand (even in the dark, so the monkey cannot see his hand). Specificity of tuning varies.
How mirror neurons relate to imitation is of much current interest (e.g., see Frith & Wolpert Reference Frith and Wolpert2004; Rizzolatti et al. Reference Rizzolatti, Fadiga, Fogassi, Gallese, Meltzoff and Prinz2002; Williams et al. Reference Williams, Whiten, Suddendorf and Perrett2001). It may be tempting to think they avoid correspondence problems, thus facilitating imitation: If the same neurons code for perceived action and similar performance, no translation is needed. But things are not so simple. Rizzolatti, one of the discoverers of mirror neurons, holds that imitation requires both the ability to understand another's action and the ability to replicate it. On his view, recall, action understanding precedes imitation phylogenetically; action understanding is subserved by mirror systems, which might be necessary, but are not sufficient, for imitation. Rizzolatti (Reference Rizzolatti, Hurley and Chater2005) suggests that the motor resonance set up by mirror neurons makes action observation meaningful by linking it to the observer's own potential actions.
Mirror neurons were discovered by single-cell recording in macaques (Di Pellegrino et al. Reference Di Pellegrino, Fadiga, Fogassi, Gallese and Rizzolatti1992; Rizzolatti et al. Reference Rizzolatti, Camarda, Fogassi, Gentilucci, Luppino and Matelli1988; Reference Rizzolatti, Fadiga, Gallese and Fogassi1995), which can emulate but have not been shown to imitate in a strict sense (cf. Voelkl & Huber [2000] on marmoset imitation). Evidence for human mirror systems (Craighero et al. Reference Craighero, Buccino and Rizzolatti2002; Decety & Chaminade Reference Decety, Chaminade, Hurley and Chater2005; Decety et al. Reference Decety, Grèzes, Costes, Perani, Jeannerod, Procyk, Grassi and Fazio1997; Fadiga et al. Reference Fadiga, Craighero, Buccino and Rizzolatti2002; Hari et al. Reference Hari, Forss, Avikainen, Kirveskari, Salenius and Rizzolatti1998; Iacoboni et al. Reference Iacoboni, Molnar-Szakacs, Gallese, Buccino, Mazziotta and Rizzolatti2005; Rizzolatti et al. Reference Rizzolatti, Fadiga, Matelli, Bettinardi, Paulesu, Perani and Fazio1996; Ruby & Decety Reference Ruby and Decety2001) includes brain imaging and demonstrations that observing another person move primes the muscles needed to move in a similar manner (whether or not movements are goal directed; Fadiga et al. Reference Fadiga, Fogassi, Pavesi and Rizzolatti1995).
Rizzolatti (Reference Rizzolatti, Hurley and Chater2005) describes mirror neurons in monkey frontal brain area F5 as part of a circuit including parietal area PF and visual area STS (superior temporal sulcus). He regards a similar human brain circuit as a control system: Sensory results associated with certain movements are compared in PF to observed target movements, enabling imitative learning (cf. Iacoboni Reference Iacoboni, Hurley and Chater2005, with regard to locating the comparator in STS). Differently structured mirror systems may explain different copying capacities across species. In monkeys, mirror neurons appear to code for the goals or results of performed or observed actions.Footnote 5 By contrast, human mirror systems include specific movements that can be means to achieving goals (Fadiga et al. Reference Fadiga, Fogassi, Pavesi and Rizzolatti1995). Recall how the difference between mirroring ends versus means of action matters for the view that action understanding precedes imitation phylogenetically. If seeing someone reach for an apple produces motor activation associated with the same goal in the observer (though not necessarily with the same movements in the observer), that could provide information about the observed action's goal directness. But it wouldn't provide information about how to achieve the goal by means of the observed movements, as in imitation.
Brain imaging suggests a division of labor in the human mirror system: Its frontal regions tend to code for goals of action, whereas its parietal regions tend to code for means (i.e., movements; Iacoboni Reference Iacoboni, Hurley and Chater2005). (Monkey parietal mirror neurons seem to be goal–related; Fogassi et al. Reference Fogassi, Ferrari, Gesierich, Rozzi, Chersi and Rizzolatti2005; Nakahara & Miyashita Reference Nakahara and Miyashita2005.) One theory of how this division of labor enables imitation relates signals generated by these brain regions to comparator circuits for instrumental motor control combining inverse and forward models. Inverse models estimate the motor plan needed to achieve a goal in a given context. They can be adjusted by comparison with real motor feedback, but this is slow. It is often more efficient to use real feedback to train forward models, which anticipate the sensory effects of motor plans, associating action with its perceived results (as do mirror neurons). Forward models combine with inverse models to control goal-directed behavior more efficiently. Forward models can predict the results of imitative motor plans for comparison to observed action, and motor plans can be adjusted until a match obtains (Flanagan et al. Reference Flanagan, Vetter, Johansson and Wolpert2003; Iacoboni Reference Iacoboni, Hurley and Chater2005; Miall Reference Miall2003; Wolpert et al. Reference Wolpert, Doya and Kawato2003).
Thus, mirror neurons are arguably part of the neural basis for true imitation, though not sufficient for it. Monkey mirror neurons code for ends rather than means. Human mirror systems, by contrast, have articulated structure: some regions code for goals, whereas others code for specific movements that are means to goals. It has been suggested that human mirror systems enable imitation (not just emulation) because they code for means as well as ends (unlike the macaque's system), and that mirror neurons contribute predictive forward models to subpersonal comparator control circuits.
2.3. Functions
Human brains differ most from chimp brains in expanded areas around the Sylvian fissure which subserve imitation, language, and action understanding – where many mirror neurons are found (Iacoboni Reference Iacoboni, Hurley and Chater2005). Can mirror systems illuminate the functions of imitation in relation to distinctively human capacities – for language, or for identifying with others and understanding the mental states motivating their actions? The relationships among capacities for imitation, language, and mindreading are important for understanding phylogeny and human development. Does development of either language or mindreading depend on imitation? If so, at what levels of description and in what senses of “depend”? Or does dependence run the other way? Or both ways, dynamically? Answers may differ for language and for mindreading. Issues about relations between imitation and mindreading entwine with issues about whether mindreading is best understood as theorizing about other minds or as simulating them.
I shall survey some hypothesized functions of imitation in language, cultural evolution, cooperation, and mindreading. The first three topics, discussed briefly, provide context for SCM and illustrate its broader relevance to understanding what is distinctive about human minds. Mindreading is directly related to SCM and so is discussed more fully.
2.3.1. Language
It has been suggested that “mirror neurons could … be an important neural stepping stone … to spoken language” (Miall Reference Miall2003, p. 1). Mirror systems for action goals include Broca's area,Footnote 6 a main language area of human brains, which is active during imitative tasks. Moreover, transient virtual “lesions” to Broca's created by transcranial magnetic stimulation interfere with imitative tasks (Heiser et al. Reference Heiser, Iacoboni, Maeda, Marcus and Mazziotta2003; Iacoboni Reference Iacoboni, Hurley and Chater2005). Nativism about language might view Broca's as the best candidate for an innate language module (M. Iacoboni, in discussion). But discovery that Broca's subserves mirror systems and has some role in enabling imitation has generated new arguments about how language acquisition could build on capacities for action understanding and imitation, in either evolutionary or developmental time frames, exploiting imitative learning rather than, or in addition to, innate linguistic knowledge (Arbib Reference Arbib2005; Arbib & Rizzolatti Reference Arbib and Rizzolatti1997; Iacoboni Reference Iacoboni, Hurley and Chater2005; Rizzolatti & Arbib Reference Rizzolatti and Arbib1998; Reference Rizzolatti and Arbib1999; Stamenov & Gallese Reference Stamenov and Gallese2002). (On language and social learning, see Baldwin Reference Baldwin, Moore and Dunham1995; Barkley Reference Barkley2001; Christiansen Reference Christiansen1994; Reference Christiansen, Hurley and Chater2005; Christiansen & Kirby Reference Christiansen and Kirby2003; Deacon Reference Deacon1997; on establishing shared reference to objects through joint attention, via gaze following and role-reversal imitation, see Tomasello 1999.)
What features of imitation and human mirror systems might language build on? First, I suggest, flexible articulated relations between means and ends in imitative learning could be an evolutionary precursor of arbitrary relations between symbols and referents. Decoupling a particular bodily movement from a given result and treating it as a potential means to various possible results in varying circumstances (see SCM, Layers 2 plus 4, in sects. 3.2 and 3.4) may be a step toward treating it as lacking an intrinsic function and so available for an arbitrarily or conventionally assigned communicative function.
Second, mirror systems provide a common code for actions of self and other, and thus for language production and perception; by enabling intersubjective action understanding, mirror systems may be the basis for the intersubjective parity, or sharing of meaning, essential to language (Arbib Reference Arbib2005; Iacoboni Reference Iacoboni, Hurley and Chater2005).
Third, the flexible recombinant structure of ends and means in imitation may be a precursor of recombinant grammatical structure in language (Arbib Reference Arbib2005). The latter may result when creatures with recombinant imitative skills learn to pursue their goals by recombinant manipulation of external symbols.
Fourth, finding recombinant units of action in streams of bodily movement has parallels with finding linguistic units (e.g., words) in continuous acoustic streams of speech (Byrne Reference Byrne, Hurley and Chater2005). The modular structure of skilled action facilitates flexible recombination. Patterns of action organization could be learned in program-level imitation,Footnote 7 despite variation in implementational details, by using mirror mechanisms plus mechanisms for parsing behavior modules. Behavior parsing and the recombinant structure of program-level imitation may be precursors of human capacities to perceive underlying structures of intentions or causes in the surface flux of experience – and perhaps of syntactic parsing and the recombinant structure of language.
2.3.2. Cultural evolution
A more fundamental question is: Why might evolution favor neural structures that enable various forms of copying to begin with? Suppose individuals vary in behavioral traits that are not genetically heritable, so some reproduce more successfully than others. Their offspring may benefit by acquiring behaviors from their successful parents by copying, as well as genetically. By copying reproductively successful parents, offspring can acquire nonheritable behaviors associated with appropriate environmental conditions. If individual learning is costly, copying may contribute more to genetic fitness.
If true imitation requires mirror circuits for means and ends to be linked in ways that give social learning recombinant flexibility, it should be harder to evolve than movement priming or emulation. And, indeed, it is found in fewer species. But wouldn't this rare development from other forms of copying to imitation be maladaptive? Recall the short-term disadvantage of children compared to chimps in two-action paradigms: Children have a greater tendency to imitate even inefficient models, whereas chimps have a greater tendency to emulate and find more efficient means to attractive goals (Nagell et al. 2003; Whiten et al. Reference Whiten, Horner, Marshall-Pescini, Hurley and Chater2005b). Despite this, could the stronger imitative tendency be adaptive long-term?
Yes: via the ratchet effect (Tomasello 1999). Gifted or lucky individuals may discover efficient new means to goals – means that are not readily rediscoverable by independent trial-and-error learning. These would be lost without recombinant imitative learning, which preserves and disseminates valuable instrumental innovations, providing a platform for further innovation. Once imitation evolves genetically, it provides a mechanism of cultural and technological transmission, accumulation, and evolution. The effects of imitative copying and selection intertwine with those of genetic copying and selection; culture and life coevolve (see and cf. Baldwin Reference Baldwin1896; Barkley Reference Barkley2001, p. 21; Blackmore Reference Blackmore1999; Reference Blackmore and Aunger2000; Reference Blackmore2001; Boyd & Richerson Reference Boyd and Richerson1982; Reference Boyd and Richerson1985; Dawkins Reference Dawkins1976/1989; Deacon Reference Deacon1997; Dennett Reference Dennett1995; Gil-White Reference Gil-White, Hurley and Chater2005; Henrich & Boyd Reference Henrich and Boyd1998; Henrich & Gil-White Reference Henrich and Gil-White2001; Hurley & Chater Reference Hurley and Chater2005a, part 4).
The capacity for selective imitation may have an important role in underwriting the ratchet effect (Harris & Want Reference Harris, Want, Hurley and Chater2005). Imitation with selective inhibition has the advantages of theft over honest toil: Instead of letting hypotheses die in his stead, a selective imitator lets others die in his stead, reaping the benefits of success without unusual native wit while avoiding the costs of trial and error. Imitative social environments may in turn generate pressure to prevent successful techniques being appropriated cost free by competitors, resulting in capacities for covert or simulated action, shielded from potential imitative theft (Barkley Reference Barkley2001, pp. 9, 18–21).
2.3.3. Cooperation
As well as being subject to automatic copying influences, humans often deliberately select a behavior pattern to imitate because it is associated with certain traits or stereotypes, even if they themselves don't exemplify these traits or stereotypes. This can be benign and contribute to moral development (J. Prinz Reference Prinz, Hurley and Chater2005); perhaps I can become virtuous, as Aristotle suggested, by behaving like a virtuous person.
But, like automatic copying, deliberate selective imitation does not always operate benignly. Selective imitation can provide “Machiavellian” social advantages (Byrne & Whiten Reference Byrne and Whiten1988; Whiten & Byrne Reference Whiten and Byrne1997). It can steal not only instrumental successes but also cooperative benefits from competitors. Suppose information about the mental states of others is not transparently available. A cooperative group can share certain behaviors by which members identify one another, obtain cooperative benefits, and exclude free-riding noncooperators. Cooperators may copy such identifying behaviors from other cooperators.
Noncooperators could invade such a cooperative group by selectively copying its identifying behaviors. They could thus induce cooperation from group members while failing to cooperate in return, deceptively obtaining cooperative benefits without paying the costs. Free riding via deceptive copying partially appropriates cooperative benefits based on in-group behavioral copying. While greenbeard genes could produce genetically determined analogues of such free riding (Dawkins Reference Dawkins1982, p. 149), selective copying provides the evolutionary advantages of flexible free riding, which is not dependent on genes for specific behaviors.
How can cooperative benefits be defended against free riding through deceptive copying? An arms race between behavioral signaling and deceptive copying in cooperative games arguably produces pressure for imitative and mindreading abilities. As a result, certain solutions to cooperative games, which require mindreading rather than mere behavior prediction, may become available. Mindreading can be based on behavioral evidence yet still have functional advantages over behavior prediction (Hurley Reference Hurley2005a).
To elaborate: To counter invasion by increasingly sophisticated deceptive mimics, mutual recognition processes among cooperators would move progressively further from copying and detecting superficial behaviors and toward more subtle and covert imitation and detection of underlying mental causes of behavior. Mere behavior reading would move toward ever-smarter reading of behavioral evidence for intentions. Mere copying would in turn become more creative and flexible, with means/ends structure: imitation. This arms race could produce capacities for mindreading and intersubjective identification via covert mirroring, albeit based on subtle behavioral perceptions (cf. Krebs & Dawkins Reference Krebs, Dawkins, Krebs and Davies1984).
The advance from cooperation plus deceptive copying via imitation to mindreading is significant for enabling cooperation and obtaining its benefits. Certain solutions to collective action problems effectively require recognizing and identify with others' mental states. A simple self-referential mirror heuristic Footnote 8 for non-iterated Prisoners' Dilemmas (PDs) says: cooperate only with any others you meet who act on this same rule (Howard's mirror strategy [Howard 1988]; Danielson's self-same cooperation [Danielson Reference Danielson and Vallentyne1991; 1992Footnote 9). When another player doesn't share your mirror heuristic, you don't cooperate with him. Famously, Tit-for-Tat can outperform Defection in iterated PDs, where given players meet repeatedly; but mirror heuristics outperform Defection even in non-iterated PDs, where given players don't meet again.Footnote 10
Mirror heuristics effectively require mindreading: discovering another player's intention, not simply predicting his behavior (Danielson Reference Danielson1992, pp. 75–82; Schmitt & Grammar 1997).Footnote 11 They are conditional metaheuristics: they explicitly condition cooperation on the other's operative heuristic itself, not on his predicted behavior. (Tit-for-Tat requires not mindreading but memory of a given player's behavior in past games.) Employing a mirror heuristic requires discerning, more or less reliably, whether others are operating on a mirror heuristic – a general intention or rule of choice. Which choices mirrorers should make are not determined by predicting what others will do; mirrorers need to know whether others have the intentions of a mirrorer before they can determine what to do. Participants in mirror-based cooperation must not only be mindreaders, but also be able to identify, more or less reliably, other mindreaders.
In non-iterated games, the mindreader has not previously played with the player she is mindreading, so cannot refer to memories of their past play. In informationally clouded social environments, mindreading is based on evidence from observing behavior, which may be subtle and/or deceptive. But mindreading need not be foolproof to provide mirror-based cooperative benefits to individual mirrorers and groups of them; the benefits would vary with the accuracy of mindreading (cf. Danielson Reference Danielson1992, pp. 157ff.).
What is the difference between genuine mindreading and mere smart behavior reading? Many social problems that animals face can be solved via behavior-circumstance correlations and behavioral predictions, without postulating mediating mental states. What problem-solving pressures are addressed by additionally attributing mental states to explain observed behavior? (Cf. Call & Tomasello Reference Call and Tomasello1999; Hare et al. Reference Hare, Call, Agnetta and Tomasello2000; Reference Hare, Call and Tomasello2001; Heyes Reference Heyes1998; Heyes & Dickinson Reference Heyes, Dickinson, Davies and Humphreys1993; Hurley Reference Hurley, Hurley and Nudds2006b; Povinelli Reference Povinelli, Carruthers and Smith1996; Povinelli & Vonk Reference Povinelli, Vonk, Hurley and Nudds2006; Sterelny Reference Sterelny2003, pp. 67ff.; Tomasello & Call Reference Tomasello, Call, Hurley and Nudds2006; Whiten Reference Whiten, Carruthers and Smith1996; Reference Whiten, Whiten and Byrne1997.)
Mental state attributions may support more flexible behavior prediction in novel conditions. But mirror metaheuristics show that mindreading's function in enabling cooperation goes beyond providing better predictions of behavior. As explained, mirror metaheuristics do not require predicting other players' behavior per se, but rather, ascertaining the heuristic they use. Such mindreading may be done by observing others' behavior, but that does not mean that its function is only behavior prediction, or that it has the same functions as behavior prediction. Mindreading can function to enable cooperation in a way that merely predicting behavior cannot (Danielson Reference Danielson1992, p. 82), even if mindreading is based on behavioral evidence. This is why the emergence of mindreading, via imitation, from an arms race between cooperative and deceptive copying is significant for enabling cooperation.
2.3.4. Mindreading
What more can be said about the possible functions of imitation in relation to mindreading?
Human mirror systems may be part of the mechanisms for understanding observed actions and intersubjective empathy. Observing another act primes your motor system to copy, even if overt copying is inhibited. Covert copying is a kind of process-driven simulation, which uses offline the processes that would be used actually to copy the observed action, but it inhibits motor output. This direct resonance with another's action provides a fundamental similarity between yourself and other agents that enables the understanding of another's actions as instrumentally structured. Mirror systems also provide a plausible neural basis for emotional empathy and understanding (see Adolphs Reference Adolphs2002; Decety & Chaminade Reference Decety and Chaminade2003; Reference Decety, Chaminade, Hurley and Chater2005; Gallese Reference Gallese2001; Reference Gallese, Hurley and Chater2005; Gallese & Goldman Reference Gallese and Goldman1998; Goldman Reference Goldman, Hurley and Chater2005; Gordon Reference Gordon, Davies and Stone1995a; Reference Gordon1995b; Iacoboni Reference Iacoboni, Hurley and Chater2005; Iacoboni et al. Reference Iacoboni, Molnar-Szakacs, Gallese, Buccino, Mazziotta and Rizzolatti2005; Meltzoff Reference Meltzoff, Hurley and Chater2005; Preston & de Waal Reference Preston and de Waal2002; Rizzolatti Reference Rizzolatti, Hurley and Chater2005; Williams et al. Reference Williams, Whiten, Suddendorf and Perrett2001).
Within this broad perspective, I shall compare the views of Gallese, Meltzoff, Gordon, and Tomasello on simulation theory versus theory theory and on relations between imitation and mindreading. And I will preview how SCM reconciles opposed views on both topics. An outline follows.
2.3.4.1. Simulation theory (ST) versus theory theory (TT)
Gallese views mirror systems as enabling broad interpersonal empathy by implementing primitive intersubjective information, prior to differentiation of self from other.
Meltzoff views early imitation as foundational for the ability to understand other agents: In imitation my acts are directly, noninferentially identified with others' acts; I then associate my acts with my mental states and infer a similar association in others.
TT accounts of mindreading invoke laws and inferences about mental states and behavior, whereas on ST accounts mindreaders use their own psychological processes offline to attribute similar mental states or actions to others.
Underived similarity between one's own and others' acts is shared ground between Meltzoff's TT account of mindreading based on early imitation and ST accounts based on inhibited copying.
Gordon criticizes first-person- to third-person-inference accounts of early imitation's role in mindreading, for taking the self/other distinction for granted.
In Gordon's ST view of how mindreading involves offline imitation, “constitutive mirroring” “multiplies the first person” by reference to a shared scheme of reasons.
First reconciliation of ST and TT: Foundations of intersubjectivity and the self/other distinction can be provided by simulative mirroring (SCM's layers 3 and 4), although richer self/other and other/other distinctions depend on interpretation, theorizing, and inference (layer 5 and beyond).
2.3.4.2. Relations between imitation and action understanding
Tomasello and Carpenter's view that imitation depends on action understanding contrasts with views of action understanding as depending on imitation.
Second reconciliation, concerning relations between imitation and action understanding: Simple mirroring, of goals or movements, can express a fundamental intersubjectivity, enabling simple forms of action understanding and providing elements of more complex imitative mirroring with flexible instrumental structure, which in turn contributes to more articulated, instrumentally structured understanding of other agents and their minds (layers 3 and 4).
2.3.5. Simulation theory (ST) versus theory theory (TT)
On Gallese's (2001; 2005) shared manifold hypothesis, mirror systems enable various aspects of interpersonal understanding and empathy. Mirror systems develop from the way biological control systems model interactions between organisms and their environments. They provide the neural basis of a primitive intersubjective information space or “shared manifold” that is prior to a self/other distinction both phylogenetically and ontogenetically, yet preserved in human adults (SCM's layer 3 incorporates this feature). The shared manifold underwrites automatic intersubjective identifications across different perceptual modalities and action, but also for sensations and emotions: There is evidence of mirror mechanisms for pain and disgust, and hearing anger expressed increases the activation of muscles used to express anger. Empathy involves a common scheme of reasons under which persons, self and others, are intelligible, rather than recognition that others' bodies also have minds.
Meltzoff (Reference Meltzoff, Hurley and Chater2005) argues that early imitation and its enabling mechanisms begot understanding of other agents, not vice versa. In his view, ability to understand other minds has innate foundations, but develops in stages, in which imitation plays a critical role. Infants have a primitive ability to recognize being imitated and to imitate, and hence to distinguish people from other things and recognize equivalences between acts of self and other. This initial bridge between self and other provides privileged access to people that we don't have in relation to other things; it develops in three stages. First, own acts are linked to others' similar acts supramodally, as evidenced by newborns' imitation of others' facial gestures. Second, own acts of certain kinds are linked bidirectionally to own mental states of certain kinds, through learning. Third, others' similar acts are linked to others' similar mental states. This early process is conceived not as formal reasoning, but as processing the other as “like me.” It gets mindreading started on understanding agency and closely associated mental states: for example, intentions, emotions, and desires. Meltzoff emphasizes that mindreading isn't all or nothing. (Tomasello [1999] makes similar claims for nonhuman animals.) Understanding mental states further from action, like false beliefs, comes in later development.
Meltzoff's view of mindreading is usually put in the theory theory (TT) rather than simulation theory (ST) category, but it has ST as well as TT aspects. TT regards commonsense psychology as a proto-scientific theory. It represents knowledge as laws about mental states and behavior that can be known innately or discovered by testing hypotheses against evidence. Specific mental states and behaviors are inferred from other mental states and behaviors by means of such laws; the process does not depend on copying. On ST, mindreading starts with taking another's perspective and generating “pretend” mental states or behavior that match the other's. These offline states are not objects of theoretical inference. Rather, they are entered into the simulator's own psychological and decision-making processes, which are held offline to produce further simulated mental states and behavior that are then assigned to the other. Further behavior by the other can be predicted, or mental states attributed that explain his observed behavior. Such simulation is an extension of practical abilities rather than a theoretical exercise: it copies the other's states and uses the copies in the simulator's decision-making equipment, instead of using laws to infer the other's states (Davies & Stone Reference Davies and Stone1995a; Reference Davies and Stone1995b).
Meltzoff's three-stage process can be restated in explicitly TT terms. First, innate equivalence between my own and others' acts (exploited by early imitation and recognition of being imitated) provides a fundamental, underived similarity between some acts (by myself) and other acts (by another). Second, first-person experience provides laws linking one's own acts and own mental states. Third, it is inferred that another's acts and mental states are lawfully linked in the same ways as my similar acts and mental states are linked. Proceeding through stages 2 and 3, we find inferences from first-person mind-behavior links to similar third-person links as in traditional arguments from analogy.Footnote 12 “The crux of the ‘like-me hypothesis’ is that infants may use their own intentional actions as a framework for interpreting the intentional actions of others” (Meltzoff Reference Meltzoff, Hurley and Chater2005, p. 75). For example, 12-month-old infants follow a model's “gaze” significantly less when the model's eyes are closed rather than open, but only similarly refrain from following the “gaze” of a blindfolded model after they are given first-person experience with blindfolds.
However, as Meltzoff points out (personal communication), there is no first-person to third-person inference at stage 1. The initial bidirectional self-other linkage, expressed in early imitation and recognition of being imitated, is via a supramodal common code for observed and observer's acts that's direct and noninferential (Meltzoff & Moore 1997). Stage 1 of Meltzoff's view thus has important common ground with ST: In covert offline copying, direct noninferential resonance with another's action with inhibited motor output enables understanding of the other's action. But such direct noninferential resonance can also occur in overt copying, as Meltzoff postulates; copying can provide information for understanding another's actions, even when not inhibited and serving other functions. Mindreading's foundation at Meltzoff's first stage is noninferentially direct, not theoretically derived. His view shares this nontheoretical basis, at its first, online copying stage, with ST views of mindreading as based on offline copying, though they diverge on how mindreading develops further. If mindreading develops in stages, theoretical inference can enter later, increasing with development.
While Meltzoff's theory theory involves first-person to third-person inference, Gordon's “radical” simulation theory (see Gordon Reference Gordon1986; 1995; Reference Gordon, Davies and Stone1995a; Reference Gordon, Carruthers and Smith1996; Reference Gordon2002; Reference Gordon, Hurley and Chater2005) explicitly rejects it, and provides a different view of the relations between imitation and mindreading. In constitutive mirroring, a copied motor pattern is part of the perception of another's action, though overt movement may be inhibited. Gordon finds constitutive mirroring in Gallese's primitive intersubjective “we”-space, the basis of empathy that implicitly expresses similarity of self and other rather than their distinctness. When constitutive mirroring imposes first-person phenomena, a process of analysis by synthesis occurs whereby another's observed behavior and the self's matching response – part of the very perception of the other's behavior – become intelligible together, in the same process. When I see you reach to pick up the ringing phone, your act and my matching response are made sense of together within a scheme of reasons that is fundamentally common to persons. As Gordon (Reference Gordon, Hurley and Chater2005) puts it, I don't infer from the first to the third person, but rather multiply the first person. To understand what I or another believes, perceives, or intends, I look out at the world and the reasons it provides, though in the case of others I imaginatively recenter to the other's perspective (Gordon Reference Gordon, Davies and Stone1995a).
Gordon criticizes first-person to third-person inference in Meltzoff's account not because it attributes similarity to one's own and others' acts or experiences, but because it requires that they be identified and distinguished. In Meltzoff's stage 1, there is innate equivalence between acts of self and other; this stage may involve constitutive mirroring, as in Gallese's primitive shared manifold. But later stages of Meltzoff's account, where analogical inference occurs, require that self and other also be distinguished: If this kind of act by me is linked to my mental states of a certain kind, then a similar (as per stage 1) kind of act by another is also linked to her mental states of a similar kind. Gordon explains that I cannot infer analogically from a to b unless I can distinguish a from b. He is skeptical that infants have this capacity, though mature imitative mirroring may involve such inference (Gordon Reference Gordon, Hurley and Chater2005).
Pure ST views of mindreading are standardly criticized for lacking resources to explain how mature mindreaders distinguish and identify people and track which actions and mental states are whose. Gordon suggests that multiple first persons are distinguished and tracked in the personal-level process of making others intelligible, avoiding incoherence under the common scheme of reasons (see also Hurley Reference Hurley1998, part 1; 1989). Mental states that don't make sense together are assigned to different persons. Can this be done in pure simulation mode, without theorizing? Simulation supposedly uses practical abilities rather than theorizing about actions. How does interpreting an action to make sense of it differ from theorizing about it? When I use practical reason offline in interpretative mindreading, I don't formulate normative laws from which I make inferences; rather, I activate my normative and deliberative dispositions. As Millikan might say (Millikan Reference Millikan, Hurley and Chater2005), my thought about another's action isn't wholly separate from my entertaining that action.
SCM will suggest a reconciliation between ST and TT. The fundamental similarity between self and other is understood in terms not of theorizing but mirroring (as in Gallese's shared manifold, Gordon's constitutive mirroring, Meltzoff's innate self-other equivalence, and SCM's layer 3). Such primitive intersubjectivity persists into adulthood, providing a basis for mature empathy and mindreading, as Gallese holds. The informational origin of the self/other distinction is understood in terms of monitoring whether mirroring is inhibited (layer 4). As mindreading develops, it also employs a richer self/other distinction, as when children come to distinguish imitating from being imitated (see Decety & Chaminade Reference Decety, Chaminade, Hurley and Chater2005), or to attribute beliefs different from their own to others. Mature personal-level mindreading requires abilities to distinguish, identify, and track multiple other persons, to assign acts and mental states to them in an interpretative process, and to entertain multiple possible acts by multiple other persons (layer 5). If decentering from me-here-now creates a trail to others and other possible actions, mature mindreading creates multiple branching and interacting trails. Negotiating these, by using the full range of distinctions and identifications required by mature mindreading, probably demands theoretical resources, even though the subpersonal enabling foundations of intersubjectivity are found in mirroring, and of the self/other distinction in monitored simulation. SCM explains how mirroring and simulation can provide foundations for mindreading on which theorizing builds.
2.3.6. Relations between imitation and action understanding
How are imitation and action understanding related to each other? On imitation-first views, imitation underwrites early mindreading abilities. Gallese, Meltzoff, Gordon, and Goldman stress the contribution of imitation to understanding other agents. By contrast, understanding-first views emphasize the way imitative learning depends on action understanding and intention reading (Carpenter et al. Reference Carpenter, Akhtar and Tomasello1998; Rizzolatti Reference Rizzolatti, Hurley and Chater2005; Tomasello & Carpenter Reference Tomasello, Carpenter, Hurley and Chater2005). Recent paradigms with children where the demonstrated action is unsuccessful or accidental (Meltzoff Reference Meltzoff1995) distinguish imitation from other forms of social learning more clearly than the two-action method does. If the observer copies what was intended even though it wasn't achieved, as opposed to copying only the observed movements or the observed though unintended result, that suggests the observer understands the intentional structure of the observed action. Tomasello and Carpenter argue that intention reading is needed to explain what is copied by imitators when the modeled behavior is the same across conditions while the modeled intention varies. In their view, results from various paradigms are most parsimoniously explained by holding that children use their understanding of intentions to imitate.
Imitation-first and understanding-first views are not necessarily opposed; each may tell only part of the story. SCM provides a framework for their reconciliation, accommodating both views at different points in its layered architecture. Different types of copying, and covert forms of each that enable corresponding types of understanding, can dovetail over evolution and development, building on one another reciprocally, with increasing instrumental structure in both action and understanding over successive stages. A simpler form of copying can precede a simpler form of understanding, which precedes a more complex form of copying, which precedes a more complex form of understanding (the ordering can be interpreted phylogenetically or ontogenetically). Start, say, with goal mirroring and emulation. Covert goal mirroring can then enable understanding the goals of observed action. Such goal understanding, along with mirroring of movements, may be needed for instrumentally articulated imitation (understanding first). But richer instrumental understanding, of how observed means contribute to observed ends, may involve covert imitation (imitation first). In SCM, self-other similarities expressed by mirroring, whether more or less structured, are informationally prior to the self/other distinction required for understanding action as another's.
3. Part 2. The shared circuits model
The shared circuits model (SCM) shows how subpersonal resources for control, mirroring, and simulation can enable the distinctively human sociocognitive skills of imitation, deliberation, and mindreading. The model has intertwined empirical and philosophical aims. One aim is to provide a unified framework for the various strands of empirical evidence and theorizing surveyed thus far. Another is to illustrate the philosophical view that embodied cognition can emerge from active perception, avoiding the “classical sandwich” architecture, which insulates central cognition from the world between twin buffers of perceptual input and behavioral output (Hurley Reference Hurley1998; Reference Hurley2001). It does this, recall, by addressing a higher-order theoretical question, about how it is possible for subpersonal processes to enable certain personal-level abilities: in particular, how it is possible to build subpersonal resources for sociocognitive skills on those for active perception.
SCM thus provides a generic heuristic framework for specific first-order hypotheses, about how particular sociocognitive capacities map onto specific layers of the model or develop in phylogenetic or ontogenetic time. SCM itself does not articulate specific first-order hypotheses, but does make some general predictions: for example, of neural systems for mirroring based on those for instrumental prediction, and of the priority of online over offline mirroring. Nor is SCM exclusive; important work in enabling persons' cognitive capacities is done by other processes, including linguistic processes. The point is to illustrate how it is possible for important cognitive resources to emerge from active perception. Although SCM is therefore somewhat abstract, in accord with its higher-order theoretical aim, I suggest in this target article how it lends itself to more specific empirical predictions, and hope the commentaries will offer suggestions as well. Details follow, layer by layer.
3.1. Layer 1: Basic adaptive feedback control
SCM begins with specific comparator feedback control systems. A comparator system generates outputs that are means to a target, by establishing an instrumental association between outputs and their results. For example, a thermostat compares a target signal with an input signal. If they don't match, system output is adjusted and the resulting change in input signal or feedback is tracked. Input continues to be recompared with target and output readjusted, to minimize mismatch to target. The elements of such control are (Fig. 1):

Figure 1. Layer 1: Basic adaptive feedback control.
1 A target or reference signal (e.g., target room temperature for a thermostat).
2 An input signal (e.g., actual room temperature), the joint result of elements 3 and 5.
3 Exogenous environmental events (e.g., nightfall).
4 A comparator, which determines whether target and input signals match and the direction and degree of any mismatch or error (e.g., the room is still five degrees below target temperature).
5 The output of the control system (e.g., the level of heat output), regulated by comparison between target and input signals (e.g., heat output is increased if measured room temperature is below target).
6 A feedback loop by which output has effects on succeeding input signals (e.g., measured room temperature rises when heat output increases).
Feedback control is adaptive; output is adjusted to compensate for changing exogenous influences, keeping sensed input close to target. Under different exogenous influences, feedback calls for differing outputs to achieve the target; when the weather changes, a thermostat adjusts heat output to maintain the target temperature. Feedback at layer 1 operates in real space and time, and therefore can be slow (e.g., a room takes time to warm up after the heat is turned up). A control system implements a mapping from the target, in the context of actual input, to output, thus specifying the means for approaching the target in given circumstances. Inverse model is engineering terminology for this instrumental mapping.
Net sensed input results from the system's output plus independent environmental influences. In organisms, reafferent feedback carries input resulting from the organism's own activity, whereas exafferent input results from exogenous events. Reafference includes visual and proprioceptive inputs resulting from movements of one's hands, movement through space, manipulation of objects, and so on. Exafference includes visual inputs resulting from environmental events, such as movements by others in a social group. However, at layer 1, information distinguishing reafference from exafference is not available.
Feedback control is a cyclical and dynamic process, with no nonarbitrary start, finish, or discrete steps; input is as much an effect as a cause of output (Marken Reference Marken2002; Powers Reference Powers1973). Control depends on dynamic relations among inputs and outputs. Information about inputs is not segregated from information about outputs; this blending of information is preserved and extended in the informational dynamics of further layers. Perception and action arise from and share this fundamental informational dynamics (Hurley Reference Hurley1998; Reference Hurley2001).
Specific means/ends associations or instrumental mappings can be chained (output A is the means to controlled result B, while B in turn is the means to controlled result C, and so on) or organized into hierarchies. There are independently determined evolutionary, developmental, and individual differences in the grain and complexity of the possible control sequences and hierarchies of different creatures.
3.2. Layer 2: Simulative prediction of effects for improved control
Real-time feedback can be slow and produce overshooting. Control functions can be speeded and smoothed by adding simulative predictions to a comparator system (Grush Reference Grush2004; Miall Reference Miall2003): Instrumental output-result associations can then be activated predictively, simulating the effects of specific outputs for informational purposes. Over time, associations are established between output and subsequent input in certain contexts, so that copies of motor-output signals can evoke associated input signals. An inner loop maps copies of output signals directly onto “expected” input signals – means to results (Fig. 2; new aspects italicized). Forward model is engineering terminology for this mapping; copies of output signals in organisms are called efference copy. This subpersonal process simulates feedback – predicts the results of output on input. Prediction can occur during actual action, to smooth a behavioral trajectory by anticipating feedback, or prior to action, to provide information about alternative possible actions (layer 4).

Figure 2. Layer 2: Simulative prediction of effects for improved control.
A general improvement in instrumental control results. A control system with predictive simulation no longer need await actual feedback. A thermostat can turn the heat down before the target is reached, avoiding overshooting; hand movement can be initiated in accord with predictions of retinal signals based on eye movements. When real and simulated results don't match, a local switch can default back to actual feedback control while predictive simulations are fine-tuned to improve subsequent predictions.Footnote 13 For purposes of online instrumental control, the system need not monitor continuously or access globally whether it is using actual or simulated feedback.
Comparisons can now be made not just between targets and actual results of action, but between the latter and anticipated results. This permits reafference to be distinguished from exafference: Information about the organism's goal-directed behavior can be distinguished from information about environmental events. Consider the familiar ambiguity: When my train pulls out of the station, I register movement relative to the train on the next platform, but this does not itself provide information about whether my train or the neighboring train is moving. Comparison of predicted feedback from action (efference copy) with actual feedback provides resources to resolve an analogous subpersonal ambiguity (between reafference and exafference), and hence to distinguish the self 's activity from environmental events.Footnote 14 This subpersonal information can contribute to enabling the personal-level distinction between action of the self and perception of the world. The perception/action distinction emerges from subpersonal informational dynamics between world-to-animal inputs and animal-to-world outputs. However, perception and action don't map, respectively, onto input and output (Hurley Reference Hurley1998). Rather, layer 2 inherits unsegregated information about inputs and outputs from layer 1, and uses this blended information in enabling the perception/action distinction. Perception and action share these basic informational dynamics and processing resources. SCM thus provides a dynamic process version of a common coding view of perception and action (sect. 2.2).
However, the system does not yet provide information about similarities between the agent's actions and actions by others, nor information distinguishing the agent's actions from similar actions by other agents (as opposed to distinguishing the agent's actions from environmental events in general). This suggests that a basic distinction between action by self and perception of world, associated with instrumental control functions, can be available to creatures still lacking intersubjective information, a self/other distinction, or mindreading abilities. There are more and less fundamental layers of information about self (self in SCM is neutral between persons and other animals).
Some general predictions derive from layer 2. First, neural mechanisms that implement sensorimotor affordance associations (such as canonical neurons [sect. 2.2]) are predicted. Suppose an animal typically acts in ways afforded by certain kinds of object: for example, eating a particular food in a specific way. Copies of motor signals for eating movements will be associated with a multimodal class of exafferent and reafferent inputs deriving from such objects and the agent's eating of them. Cells mediating this sensorimotor association could thus have both sensory and motor fields and carry information about objects' affordances. Second, deficits in predictive simulation functions should be associated with deficits in distinguishing self from world and action from perception.Footnote 15 A specific first-order prediction relating to layer 2 might be that capacities requiring information that distinguishes self from world are phylogenetically prior to capacities for social learning and action understanding.
3.3. Layer 3: Mirroring for priming, emulation, and imitation
SCM next postulates that instrumental output-result associations can be activated bilaterally, from effect to cause, as well as from cause to effect. Not only do copies of motor signals predictively simulate input signals (layer 2), but input signals can evoke corresponding motor signals.Footnote 16 To put it more technically, not only does efference copy produce simulated reafferent input in forward models, but input signals can evoke mirroring efference or motor output. Mirroring in effect runs the predictive simulations of forward models in reverse (Fig. 3).
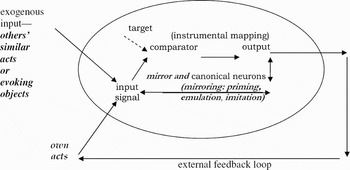
Figure 3. Layer 3: Mirroring for priming, emulation, and imitation.
Observed actions are thus mirrored in the observer; if mirroring is sufficiently strong and not inhibited, overt copying results. (Mirroring is here a functional subpersonal rather than neural description of behavioral priming produced by observing action, and may be implemented in neural mirror systems.) Mirroring within specific control structures of differing grain and structure would enable different copying capacities, observed across various social species: mirroring of basic movements in priming (Rizzolatti's “low-level resonance”), mirroring of goal-directed action or emulation (Rizzolatti's “high-level resonance”), and even full-fledged imitation (if mirrored elements of control structures are sufficiently articulated and flexibly linked to provide the information needed for social learning of novel means to a goal [see further on in this section]).
Note the intimate relationship between the sharing of circuits for self and other and for action and perception: Layer 3's shared informational dynamics for intersubjectivity presupposes layer 2's shared informational dynamics for perception and action, which builds on layer 1's generic informational dynamics for sensorimotor control. SCM explicitly builds shared resources for self and other on those for action and perception. It thus integrates W. Prinz's shared information for perception and action (though in terms of functional dynamics rather than coding) with Gallese's primitive intersubjective “we”-centric information. By bringing together information about motor causes of one's own and others' similar observed actions, mirroring enables simulation of means/ends associations from either direction: Observed action retrodicts motor activation in the observer via mirroring of causes, which are associated with further results via simulative prediction of effects. But although mirroring makes information about action's instrumental structure accessible bilaterally (from acting and from observing others act), mirroring does not yet distinguish own action from observed action. At layer 3, self and other share informational resources: intersubjective information is subpersonally prior to the self/other distinction (as Gallese holds [sect. 2.3]).
Instrumental mapping and mirroring both map input to output; SCM distinguishes them functionally (neural implementations may overlap). Instrumental mappings have control functions: given certain inputs, they select motor outputs that in turn produce inputs matching a target. Although mirroring exploits instrumental control structures and also produces motor outputs, given certain inputs, it does not itself select outputs as means to inputs that match observed action – or any other target (cf. Peterson & Trapold Reference Peterson and Trapold1982). Rather, SCM postulates, mirroring is a by-product (via reversal) of predictive simulations, which do have instrumental control functions. However, the resulting automatic copying tendency has evolutionary functions, and copying can be exapted for cognitive functions associated with imitation, action understanding, or signaling; these in turn can enable advanced social (“Machiavellian”) forms of instrumental control (see sects. 2.1 and 2.3). Whether specific mirroring capacities are adaptive depends on their potential functions for different social species under different evolutionary pressures.
How could mirroring arise? Consider first movements that produce visual reafference. When creature A sees her own hand movements, associations form between copies of motor signals for these movements and visual reafference from these movements. Cells mediating this association can acquire congruent sensory and motor fields. These cells would also fire if creature A receives similar visual inputs from creature B's similar hand movements; the cells wouldn't distinguish observer's action from observed action producing similar inputs. So, like mirror neurons, they would fire both when A acts and when she observes such similar action by B. They mediate associations between copies of motor signals and a class of inputs including both characteristic reafference from the agent's movement and similar exafference from others' similar movements.
How could mirroring arise for movements not seen by their agents? This requires a correspondence between one's own and others' similar acts, without reafferent feedback from own acts in the same modality as observations of others' acts. Facial movements normally produce proprioceptive rather than visual feedback: How can one's own facial movements be associated with visual information about similar observed facial movements, to enable copying of facial expressions?
Several answers are possible (sect. 2.2), all compatible with SCM. Some supramodal correspondences may be innate (as in newborn copying; Meltzoff Reference Meltzoff, Hurley and Chater2005; Meltzoff & Moore 1997). Some may be acquired through experience with mirrors, or with being imitated (Heyes Reference Heyes, Hurley and Chater2005). Some could be established via stimulus enhancement, as follows. Suppose a creature repeatedly sees conspecifics act a certain way; their actions draw its attention to the typical objects of their actions, which evoke in the observer an innate or previously acquired response. An indirect association results between visual observations of others' actions and one's own similar action. This isn't copying initially; the object independently evokes others' and own similar acts. But the indirect, object-mediated association between own and others' similar acts may become direct with repetition. Cells mediating it could thus acquire similar sensory and motor fields, so that observing another's act primes similar action by the observer. Therefore, stimulus enhancement could develop into copying, and an indirect link via an enhanced stimulus into a direct sensorimotor mirroring link.Footnote 17 SCM is neutral about whether such correspondences are innate, acquired, or both.
SCM does not describe one all-inclusive structure, but has multiple instances for specific movements and results, at various points along different means/ends chains (cf. Fogassi et al. Reference Fogassi, Ferrari, Gesierich, Rozzi, Chersi and Rizzolatti2005; Wolpert et al. Reference Wolpert, Doya and Kawato2003). The ends/means distinction is relative and applies along spectra of means/ends links in which basic movements are means to proximal results that are means to more distal results. Such control spectra can vary in grain. SCM could apply at successive points along a control spectrum, or between spectra (with the right neural connections); one circuit's target could be the means to next circuit's target. A network of control spectra could support hierarchical control and flexible recombination of means and ends. With mirroring added, it could enable movement priming, emulation, or imitation.
SCM predicts that capacities for specific kinds of copying and social learning will vary across species and development with
(A) the grain and complexity of instrumental control capacities, and
(B) which of these have associated mirroring functions and how richly and flexibly they are linked (see discussion following; Csibra Reference Csibra2005).
How do these two influences operate?
(A) Different animals are equipped to different degrees with capacities for instrumental control and associated predictive simulation. This variation reflects potential means/ends chains of differing grains and lengths and with differing degrees of lateral connectivity to enable novel combination of means and ends as instrumentally appropriate. Animals suitably equipped with control functions by evolution and development could form chains of simulative predictions, resulting in information such as: this tail movement has that effect on weight over legs, facilitating this movement trajectory in relation to that gazelle, and so on – with eating the prey later in the chain. A similar chain could lead from knee movement to winning a slalom race.
Mirroring operates such instrumental associations in reverse. Mirroring the cause of another's movement, or resulting relationship to an object, could enable movement priming, goal emulation, or even full-fledged imitation (if instrumental control and associated mirroring functions are sufficiently articulated and flexible). Combined with further information distinguishing self from other (layer 4), simulative mirroring can provide information to enable understanding of others' observed movements as instrumental actions with intentional, means/end structure. Simpler control and predictive simulation capacities, with shorter, coarser, means/ends chains, limit correspondingly an animal's potential for mirroring and related functions.
Whether observed movement primes copying or is recognized as goal-directed depends on, inter alia, the instrumental capacity potentially available for mirroring and simulative functions. Mirroring and simulation might provide information about the goals of certain observed movements, given fine-grained, complex means/ends associations, but not given coarser control capacities. No doubt a monkey can move her hand to grasp a piece of sushi and move it to her mouth to eat it. But I can move my hand to operate chopsticks to pick up sushi to dip it in soy sauce and then move it to my mouth to eat it, in order to impress my boss; given associated simulative mirroring functions, I may start to resent you for eating the last piece of sushi as soon as you reach for your chopsticks.
(B) While mirroring of instrumental associations can provide information about the instrumental structure of observed movements, mirroring does not itself determine which instrumental associations, or predictive simulations thereof, are in place and hence potentially exploitable by mirroring. Control processes can be neurally distributed, with components of an articulated mean/ends chain processed in different brain areas – control of fine movements versus gaze versus posture versus whole body movement versus external objects. Some such neural areas may have mirroring as well as predictive simulation capacities, whereas others do not. Whether mirroring is associated with particular control structures will vary across species and development, as a result of evolutionary and developmental processes, yielding different capacities for copying and for generating information about goals of observed actions.
We can understand the enabling of movement priming, goal emulation, or imitation in terms of mirroring that exploits different control structures. Mirroring associated with more basic means/ends links (Rizzolatti's ‘low-level resonance’) predicts priming of basic movements; mirroring associated with less basic means/ends links predicts priming of less basic movements or results of more basic movements. Finger movements are means to chopstick deployment; when I can control chopsticks, chopstick deployment is a means to sushi eating, which could be a means to some further social result. Suppose I eat sushi by deploying chopsticks by moving my fingers a certain way. You watch. If seeing me move my fingers generates mirroring motor activation, then you are primed to move your fingers similarly. Such movement priming predicts interference if you watch me doing X while you are doing Y.
If you can already control chopsticks, less basic mirroring and prime chopstick deployment can occur. Such priming could be goal-mediated: Your chopstick deployment could mirror mine when sushi-eating results (even if no sushi is visible), but not when the results are unrelated to sushi (cf. Umiltà et al. Reference Umiltà, Kohler, Gallese, Fogassi, Fadiga, Keysers and Rizzolatti2001). Goal emulation could be enabled by mirroring midway along a control spectrum of proximal results of movements (rather than basic movements themselves) that are in turn means to more distal results (cf. Rizzolatti's “high-level resonance”). Such midway mirroring would generate motor activation associated with a corresponding midway goal for the observer (rather than with similar basic movements in the observer). As well as enabling emulation, this would contribute information for understanding observed action as goal directed (layer 4).
The phylogenetically rare capacity for imitative learning requires flexible means/ends associations; priming and emulation, respectively, provide its ends and means components (sect. 2.1). Articulation within and linkages between mirror circuits determine whether mirroring enables imitative learning. An animal with mirror circuits along a chain of means/ends associations may never have used a certain means to a given goal. But if an observed novel means to that goal is mirrored, and neural links permit specific mirroring activations to be flexibly combined with targets, that goal and those mirrored means may be newly associated, capturing information about novel instrumental structure in observed action and enabling imitative learning. You might learn to use chopsticks by watching me; emulation plus trial and error would be bettered by mirroring that primes your finger movements towards those you see me make that are associated with the target of chopstick deployment.
Children's greater imitative tendencies, compared with chimps, may depend not on the presence versus absence of a mirror system, but on articulated relationships among multiple mirror circuits, permitting recombinant structure in social learning (sect. 2.1; cf. Barkley [2001] on recombinant structure in executive functions and their relation to imitation). SCM predicts correctly that imitation should be found in fewer species than is movement priming or emulation separately, because imitation additionally requires linkages supporting flexible instrumental associations between mirrored means and ends at a relatively fine grain. Flexibly linked mirror circuits could also generate behavioral building blocks combined in program-level imitation of sequential or hierarchical structure. And they could allow infants to form three-way associations among observed behavior by parents (who have survived to reproduce, so may have adaptive behaviors, not all of which are heritable), observed circumstances of such parental behavior, and infants' own similar behavior, enabling contextual imitation: act like that, when the environment is like this. A network of linked mirror circuits could also permit mirroring activation to spread and generalize automatically, as in chameleon effects (see sects. 2.1 and 2.3).
Some general predictions derive from layer 3. First, neural systems are predicted that implement mirroring functions and don't distinguish own and others' actions. Mirror neurons need not individually implement mirroring and simulation functions. Specific hypotheses might concern how mirror neurons with varying sensorimotor congruence contribute to the functions of mirror systems (cf. Csibra Reference Csibra2005). Second, functional associations are predicted between deficits in mirroring and in predictive simulation for control, which reflects shared circuits for these functions. Third, implementational associations are predicted between neural mirror systems and neural systems for comparator control and predictive simulation (sect. 2.2). Fourth, automatic behavioral priming and copying tendencies are predicted at varying grains, reflecting the articulation and complexity of control functions across species and development (as in movement priming, emulation, human infant copying, human perceptual induction effects, imitative interference and reaction-time effects, and chameleon effects). Overt automatic copying tendencies should be greater where inhibition is weaker, that is, in young children or imitation syndrome patients (sect. 2.1). Fifth, the phylogenetic rarity of imitation as opposed to movement priming and emulation is predicted (sect. 2.1).
Another thought: Recall that canonical neurons mediate sensorimotor affordance associations; for example, between copies of motor signals for eating and a class of inputs associated with objects that afford eating and the eating of them. As a result of such associations, observing an object may prime action it affords and produce a tendency to automatic action on affordances where inhibitory function is reduced – in young children, utilization syndrome patients, or subjects with attention deficit hyperactivity disorder (Barkley Reference Barkley2001, p. 15; Lhermitte Reference Lhermitte1983; Reference Lhermitte1986; Rietveld, in preparation). Whether inhibitory functions are specific to imitation or action on affordances is a further question (Brass et al. Reference Brass, Derrfuss, Matthew-von Cramon and von Cramon2003).
3.4. Layer 4: Monitored output inhibition combined with simulative prediction and/or simulative mirroring
Information about the instrumental structure of observed action provided by a flexibly articulated network of mirror circuits can not only enable imitative learning, but also contribute to enabling the understanding of another's actions as instrumentally structured, including in novel or complex ways. SCM's layer 4 introduces the capacity to inhibit actual output and monitor this inhibition while instrumental associations are activated. This capacity for monitored inhibition could combine with layer 3's mirroring to enable action understanding. Or, it could combine with layer 2's online predictive simulations to enable offline instrumental deliberation. Or both.
Take instrumental deliberation first. Layers 2 and 4 could combine functionally to distinguish actual from possible actions. At layer 2, simulative predictions improve online control of ongoing action. This online function does not require the control system to monitor whether it is currently using actual or simulated feedback, as long as it can switch between them as needed to achieve the target. However, simulative predictions of results could also function offline, with actual motor output inhibited. Multiple simulative predictions could provide information about results of alternative possible actions, rather than anticipating results for ongoing action. Simulated results of alternative possible actions could be compared for the closest match to a target prior to actual action. Layers 2 plus 4 could thus provide information for “trials and errors in the head” (Millikan Reference Millikan, Hurley and Nudds2006) prior to actual trials and possibly fatal errors, allowing simulations to die in a chooser's stead. They could thus enable counterfactual instrumental deliberation and choice among alternative possible actions.
Enabling these capacities requires more than comparing simulated results of different acts with a target. It also requires monitoring whether motor output is inhibited, to track the distinction between actual and possible actions. The predicted results of actual actions will call for a very different response to the predicted results of possible actions. A creature unable to distinguish these two types of predictions wouldn't be long for this world. (Barkley Reference Barkley2001). Layer 4's monitored inhibition provides a basis for this distinction: Simulated results with output inhibition provide information about possible actions, whereas simulated results without output inhibition provide information about actual actions. Multiple predictive simulations provide information about the consequences of various actions by the agent, whereas monitoring of output inhibition provides information that these are possible actions, not actual ones.
Whether such a subpersonal informational structure corresponds directly to the personal-level sense of ability to do otherwise – of having alternative possible actions open to choice – is a further question. The point here is to explain how information for an actual/possible distinction and counterfactual practical reasoning emerges in SCM. Although this information concerns agency, it may provide a basis, when combined with language, for counterfactual reasoning in theoretical contexts.
Now consider action understanding. Layers 3 and 4 could combine functionally to distinguish self from other – more precisely, to distinguish one's own action from another's. At layer 3, observing another's action primes similar action by the observer, through mirroring. At layer 4, the observer's similar action is inhibited; observed behavior isn't actually copied. Copying behavior can be beneficial (especially for young organisms), but unselective overt copying would often have disastrous results for copiers; a prey that chases predators won't survive for long. The capacity to inhibit copying is adaptive and should be expected to evolve (Barkley Reference Barkley2001). Offline mirroring simulates in the observer the causes of observed action, reversing the direction of simulative prediction: Instead of simulating feedback that would result from motor activations, mirroring simulates motor activation that would produce results similar to those observed, with actual motor output inhibited. Simulative mirroring can thus provide information for understanding the instrumental structure of observed actions. In effect, offline copying enables action understanding (Fig. 4).

Figure 4. Layer 4: Simulative mirroring (or prediction) combined with monitored output inhibition, enabling action understanding (or instrumental deliberation).
Mirroring of means/ends associations for observed actions isn't enough to enable understanding action as another's. This also requires monitoring as to whether motor output from mirroring is inhibited, to separate information about others' actions from information about one's own. Layer 3 doesn't do this job. But use of information about actions to understand others has different consequences, and makes different demands on subsequent behavior, than does use of information about actions actually to copy them. Therefore, it would be adaptive to track the distinction between own and others' actions. Layer 4's monitored inhibition provides an informational basis for this distinction, which overlays the shared informational dynamics for own and others' actions at layer 3: simulative mirroring with monitored inhibition provides information about another's action, not one's own. Thus, simulative mirroring can provide information about the causes and instrumental structure of observed action, while monitoring of output inhibition can provide information that such actions are another's, not one's own. This is how information for a self/other distinction emerges in SCM.
The length and grain of chains of means/ends associations and the flexibility of linkages between them should affect not just the types of copying enabled by mirroring (as explained at layer 3), but also the types of action understanding enabled by simulative mirroring. Goal-mediated simulative mirroring would provide information about goals of others' movements, enabling an early stage in understanding others as acting on intentions (hence, in mindreading). If mirror circuits are sufficiently articulated and flexibly recombinant to enable imitative learning, then monitored inhibition of imitative mirroring would capture the means/end structure of novel observed action more fully and flexibly, enabling more sophisticated mindreading. Depending on the articulation of control structures with associated mirroring, adding layer 4 to layer 3 could enable different mindreading abilities.
Recall the discussion (sect. 2.3) of imitation-first versus understanding-first views. SCM reconciles them, by showing how different types of copying and action understanding can be built up, enabled by differently articulated mirroring structures combined with monitored inhibition. Mirroring can enable goal emulation without a capacity for imitation; with monitored inhibition, it can enable understanding the goals of another's action. Instrumentally articulated imitation may require understanding of goals plus mirroring of movements; covert imitation can then enable fuller understanding of the instrumental structure of observed actions.
Whether such informational structures correspond directly to personal-level empathic understanding of another's actions or to knowledge of other minds are further questions. Layer 3's “first person plural” is informationally prior to layer 4's self/other distinction. Subpersonally, the problem of “knowledge” of other minds is reconfigured: it is neither one of starting from information about self and constructing a bridge to information about others, nor one of starting from information about others and from these resources generating information about self. Rather, the similarity of own and others' acts comes first, with mirroring. Monitored inhibition then distinguishes subpersonally between instrumentally structured action “centered” on self versus “decentered” onto another; self-centering and other-centering of agency arrive together. SCM gives concrete subpersonal form to the interdependence and parity of information about self and other intentional agents. The subpersonal job that remains is not to bridge a gap between self and others, but to track distinctions among them, especially when multiple other agents are in play.
Is the subpersonal priority of intersubjective information reflected at the personal level, and if so, how? Is the personal-level problem of knowledge of other minds similarly reconfigured, to avoid both first-person to third-person and third-person to first-person inferences, that is, both the argument from analogy and from behaviorism? Not necessarily. Further questions arise, about phylogeny, development, the structure of mature capacities to understand others, and the epistemology of such understanding. The role of subpersonal information in the epistemology of other minds raises general philosophical issues about the roles of reliable information and justification in knowledge: for instance, can reliable subpersonal information support knowledge of other minds? SCM does not in itself answer these questions. Rather, it provides generic, adaptable tools for framing specific hypotheses. Care is needed: We shouldn't assume an isomorphic projection from the subpersonal level to the personal level. But we should allow that understanding subpersonal processes can contribute to understanding the personal level; this doesn't require interlevel isomorphism.
Gordon's simulation theory (sect. 2.3) suggests responses to such questions. It has affinities with SCM: both build mindreading on resources for perception and action. But there are also differences: In SCM, the priority of “first-person plural” information over the self/other distinction is subpersonal, whereas Gordon's account of how constitutive mirroring “multiplies the first person” links subpersonal mirroring directly to personal-level understanding of other minds: Observed behavior and mirroring are made sense of together, under a common scheme of reasons, and incoherent mental states assigned to different persons (Gordon Reference Gordon, Davies and Stone1995a, pp. 56–58, 68; 2002; 2005; cf. Hurley Reference Hurley1989; Reference Hurley1998).
Actual/possible and self/other distinctions are necessary for much explicit theorizing and for aspects of the normativity and intersubjectivity of the personal level. SCM thus helps to understand how features of the personal level could be informationally enabled by subpersonal processes. It suggests that these distinctions share an informational basis in monitoring of motor inhibition: theoretical informational resources arise from practical ones. But SCM does not specify a phylogenetic or developmental priority between the subpersonal actual/possible and self/other distinctions, which is left to specific first-order hypotheses: Layers 2 and 4 could combine even if layers 3 and 4 do not or if layer 3 is missing altogether; and layers 3 and 4 could combine even if layers 2 and 4 do not. Hence, in different species or stages of development, we might find creatures with action understanding but not instrumental deliberation, or vice versa.
Some general predictions derive from layer 4. First, there could be deficits in inhibitory capacities although capacities to copy, or to act on affordances, are intact (as in Lhermitte's imitation and utilization syndrome patients). Second, the priority of mirroring to inhibition of copying predicts that people with intact inhibitory capacities should nevertheless retain underlying default tendencies to copy (as in perceptual induction, imitation interference effects, priming, and chameleon effects; see sect. 2.1). In addition, SCM provides generic, adaptable tools for framing specific hypotheses. A specific first-order hypothesis might be that capacities for instrumental deliberation and for understanding others' actions are enabled by shared inhibitory resources at layer 4, predicting that deficits in these capacities should be associated (Frith Reference Frith1992, pp. 81–83, 93; cf. Barkley Reference Barkley2001). Another first-order hypothesis might predict pathological dissociations of capacities supported by layers 2 plus 4 from those supported by layers 3 plus 4. Other specific first-order predictions might concern the interleaving of various copying abilities with various types of understanding of others' actions, in an ontogenetic or phylogenetic progression leading to imitative learning and mindreading capacities. Copying without inhibition should come earlier in such progressions; capacities to inhibit copying and to distinguish others' intentional actions from one's own should develop together. Yet another hypothesis might be that adding layer 4's monitored inhibition of output to layer 3's mirroring enables a transition from behavior copying to mindreading that enables effective use of mirror heuristics by cooperators (sect. 2.3).
3.5. Layer 5: Counterfactual input simulation
Finally, the system can be taken offline for input as well as output (Fig. 5). Counterfactual inputs can simulate different possible acts by others and their results. Monitored simulation of inputs to control circuits with simulative prediction and mirroring functions can provide information distinguishing between others' actual and possible acts. This social extension of counterfactual information, combined with simulation of different possible acts by self and their results, provides information about how possible acts by self could result in possible acts by others with further results, and vice versa. These combined functions provide enabling information for strategic game-theoretic deliberation, coordination, and cooperation. To distinguish possible from actual acts by others, information is needed about whether inputs are simulated; so simulation of inputs must be monitored. Simulative informational resources for strategic in addition to instrumental deliberation enrich the practical foundations for more general capacities to manipulate counterfactual information and theorize counterfactually.

Figure 5. Fifth layer: Counterfactual input simulation enabling strategic deliberation.
Recall the discussion of TT versus ST in section 2.3. Again, SCM shows how they can be reconciled. Differentiating and tracking interacting means/ends relations for multiple possible acts by self and by multiple other agents make acute informational demands. Despite their foundational role, simulation mechanisms may be insufficient to provide information for such multi-agent, multi-possibility tracking with ramifying paths of decentering. Meeting these demands, and further demands in differentiating the epistemic states of multiple others, probably requires SCM's practical simulative informational basis for understanding other agents to be supplemented by language-dependent functions and theorizing. Mindreading, like social learning and instrumental control, is a graded phenomenon, not all or nothing (Tomasello 1999). Language can build on SCM's foundational actual/possible and self/other distinctions to enable interpretative understanding of multiple others with multiple alternatives and varying beliefs. SCM hypothesizes that mindreading has practical foundations, in simulative mirroring of means/ends relations, but allows that mature mindreading with all the bells and whistles (including understanding false beliefs) requires both simulation and language-based theorizing.
Specific predictions deriving from level 5 could concern ontogeny or phylogeny. First, since understanding goals and action is foundational for mindreading, it should be prior to understanding others' epistemic attitudes (on phylogeny, see Tomasello & Call Reference Tomasello and Call1997; on ontogeny, see Rakoczy et al. Reference Rakoczy, Warneken and Tomasello2007). Second, understanding the instrumental structure of observed action can precede, and may be a phylogenetic precursor of, understanding linguistic structure. The instrumental recombinant structure of imitative learning may combine with learned manipulation of external symbols to support the richer recombinant structure of language. Third, the fundamental transition from agents “playing against nature,” instrumentally, to agents playing against one another, strategically, can be supported by simulative mechanisms. Nevertheless, simulative mechanisms without linguistic capacities may not enable advanced mindreading, such as multi-person strategic deliberation or false-belief attribution.
4. Conclusion
SCM describes a functional subpersonal architecture, at a level above that of neural implementation but below that of the conscious and/or normative contents of persons' mental states. I now step back from the details and review the functional relations among SMC's layers:
Layer 1: SCM's starting point is dynamic online motor control, whereby an organism is closely attuned to its embedding environment through sensorimotor feedback.
Layer 2: Next, add online predictions of sensory feedback from ongoing motor output. Online predictive simulation improves instrumental control and provides information distinguishing action by the agent from perception of the world (Table 1).
Layers 2+4: Combining predictions of feedback with layer 4's capacity for monitored inhibition of output has further benefits. Offline predictive simulation distinguishes actual from possible acts and provides information about results of alternative possible acts, for offline/counterfactual instrumental deliberation.
Layer 3: Mirroring reverses layer 2's predictive associations, so that observing movements generates motor signals in the observer that tend to cause similar movements. Various mirroring structures can enable various forms of copying, with various functions. If mirroring preserves novel means/ends structure of observed actions, it can enable imitative learning. But mirroring provides intersubjective information in a subpersonal “first-person plural,” without distinction or inference between own and others' similar acts.
Layers 3+4: Combining mirroring with monitored inhibition of overt copying does distinguish own from others' acts. When the goals or means/ends structure of observed actions are mirrored, this combination provides information for various levels of understanding of another's action.;>Note that layer 4's monitored inhibition can combine independently with prediction of effects or mirroring of causes (or both), providing information for various capacities: see Table 1.
Layer 5: SCM's last layer adds the function of monitored simulation of input specifying possible observed actions. This extends counterfactual information about actions socially, providing information about possible acts by others. This function combined with inhibited mirroring (layers 3 plus 4) of possible actions can generate information about possible (as opposed to actual) actions by others (as opposed to self), and possible causes and effects of such possible actions. Linguistic and theoretical resources can be added to simulative foundations, thereby enabling deft manipulation of combined actual/possible and self/other distinctions and tracking of interacting means/ends relations among multiple possible acts by self and multiple others. Strategic social intelligence is thus enabled, whereby agents can play against one another rather than merely “against nature,” a nonagent.
Table 1. Online/offline by prediction/mirroring

Specific instances of SCM layers could combine into sequences, hierarchies, or networks permitting flexible decomposition and recombination of particular links. The numbering of layers is largely heuristic and does not necessarily represent the order of evolution or development. First-order hypotheses can map the layers onto specific phylogenetic or ontogenetic progressions, and the combination of layers can vary across particular empirical applications. For example, in the absence of layer 3's mirroring function, layers 2 and 4 could combine to provide information about results of different possible actions. Nonnegotiable features of SCM are its explanation of mirroring as an exaptive reversal of online prediction, and the way the actual/possible and self/other distinctions arise as online processes are overlain by monitored inhibition (on impulsiveness as default, cf. Barkley Reference Barkley2001, pp. 5, 22).
SCM shows how information for important cognitive capacities of persons can have a foundation in the dynamic co-enabling of perception and action. Its layered build-up of functions illustrates a horizontally modular architecture, in which rich cognitive resources emerge without a classical sandwich. Specifically, materials for active perception can generate cognitively significant resources: the action/perception, self/world, actual/possible, and self/other distinctions, intersubjective information enabling social learning and mindreading, and counterfactual information enabling instrumental and strategic deliberation.
4.1. Levels versus layers
Unlike some of the work surveyed earlier, SCM distinguishes neural, functional subpersonal, and personal levels of description. Each of SCM's functionally described layers raises questions at the level of neural implementation, as well as providing information enabling personal level capacities (Table 2). Clarity and progress are served by distinguishing levels and framing issues explicitly at a given level, or as concerning interlevel relations. Sliding between levels on a priori assumptions of isomorphism is unjustified. Nevertheless, one level can shed light on another. We can look “down” a level, seeking neural implementations of aspects of SCM's functional architecture, or “up” a level, considering what SCM's functional architecture would enable persons to do.
Table 2. The shared circuits model: Layers and levels

*JK&AC: Why the question mark? We think that Susan meant to indicate that the search for the neural basis of layer 5 might begin with mechanisms that underpin imagery, but that this was just the beginning. Just how the brain might go about simulating inputs was one of the many open empirical questions that SCM raises, which Susan hoped might be taken up in the commentaries or in future work developing SCM.
4.1.1. Looking down
SCM's implementation is distributed across neural processes and embodied activity in environments, especially social environments. It predicts neural systems mediating affordance and mirroring functions, and has a heuristic role in generating specific first-order hypotheses concerning the following:
The location of postulated comparators and simulators.
The division of neural labor in mirroring ends and means and in inhibition.
The role of mirror neurons in Broca's area and their relation to linguistic capacities.
How the compositionality of imitation relates to the compositionality of language.
While SCM is described cybernetically, dynamic systems theory could represent interactions of its implementing neural processes and embodied activity over time as evolution of a phase space, and investigate its attractor structure.
4.1.2. Looking up
SCM explains how distinctive features of the personal level can be informationally enabled. It provides theoretical resources for addressing further questions concerning the following:
Relations among distinctively human capacities for imitation, deliberation, mindreading, and language;
Relations between personal-level action/perception, self/other, and actual/possible distinctions, and whether they reflect subpersonal structure.
How SCM's layers map onto evolutionary or developmental stages.
Relations between subpersonal simulation of possible actions and the personal-level sense of being able to do otherwise.
Whether knowledge of other minds requires first-person to third-person inference or can bottom out in reliable subpersonal information at layer 3.
Whether the subpersonal priority of intersubjective information in SCM is reflected in personal-level epistemology of other minds.
Whether social cognition is related to aspects of consciousness, given the roles of comparator and simulation structures in accounts of consciousness (Frith et al. Reference Frith, Blakemore and Wolpert2000a; Gray 2004; Hesslow Reference Hesslow2002; Jeannerod Reference Jeannerod1997; Reference Jeannerod2001; Milner & Goodale Reference Milner and Goodale1995, p. 64).
Whether SCM can contribute to distinguishing conscious from unconscious processes.
Whether SCM can extend from instrumental to expressive action, including facial expressions of emotion and emotional mirroring (cf. Adolphs Reference Adolphs2002; Decety & Chaminade Reference Decety, Chaminade, Hurley and Chater2005; Gallese Reference Gallese, Hurley and Chater2005; Iacoboni Reference Iacoboni, Hurley and Chater2005; Preston & de Waal 2002Footnote 18; Rizzolatti Reference Rizzolatti, Hurley and Chater2005).
Whether extending SCM to expressive action can illuminate relations between social cognition and language or consciousness.
SCM has a cybernetic rather than conceptual structure, yet can provide information for cognitive skills – for example, mindreading; deliberation – with personal-level conceptual structure. It thus probes the kind of intelligibility to be found in explanations of how subpersonal resources enable personal-level capacities. By showing how subpersonal resources for cognition can build on those for active perception, SCM illustrates that informationally enabling subpersonal structure need not recapitulate personal-level conceptual structure in any explicitly isomorphic way. It is an empirical question, to be decided case by case, whether enabling subpersonal structure corresponds isomorphically to personal-level structure, conceptual or otherwise. Interlevel isomorphism should not be required or denied a priori. Personal-level content can remain distinctively conceptual and normative, while it nevertheless becomes intelligible how the minds of persons can arise from interactions of embodied brains with environments, including social environments.
In explanations of some systems' dynamical behavior, higher-level structure corresponds to lower-level structure. But the dynamical behavior of other, complex systems cannot be so explained: System behavior can result deterministically from nonlinear relationships among lower-level factors although its structure does not correspond to lower-level structure. Brain-body-environment systems are sufficiently complex and nonlinear that emergent structure without isomorphic lower-level structure should not surprise us.
I end by highlighting some noteworthy aspects of this model:
1 SCM avoids the traditional conception of cognition as sandwiched between separate perception and action systems. Rather, it understands perception and action as enabled by shared subpersonal dynamics, and builds subpersonal resources enabling cognition on shared resources for perception and action.
2 Shared processing of actions by self and by others in social cognition is a special aspect of shared processing of action and perception in dynamic control. I perceive your action by means that engage my capacity for similar action, enabling me to copy or understand your action.
3 These shared resources are prior to self/other and actual/possible distinctions that provide information for action understanding and instrumental deliberation. The shared processing of action and perception in dynamic control is preserved when an actual/possible distinction is overlaid via inhibition of overt action. Similarly, the shared processes of action and perceiving others' action is preserved when a self/other distinction is overlaid via inhibition of overt copying.
4 The subpersonal basis of counterfactual deliberation and mindreading is simulation of instrumentally structured agency. Linguistic and theoretical resources can built on practical simulative foundations to enable more advanced counterfactual reasoning and mindreading.
ACKNOWLEDGMENTS
For helpful comments and discussions, I thank Mark Ashton Smith, John Bargh, Paul Bloom, Jeremy Butterfield, Nancy Cartwright, Nick Chater, John Cummins, Chris Frith, Vittorio Gallese, Philip Gerrans, Alvin Goldman, Robert Gordon, Jeffrey Gray, Rick Grush, Cecilia Heyes, Marco Iacoboni, Dorothee Legrand, Andrew Meltzoff, Alva Noë, Hanna Pickard, Joëlle Proust, Nicholas Rawlins, Simon Saunders, Marleen Schippers, John Schureman, Nicholas Shea, Evan Thompson, members of various audiences, and anonymous referees.







