



Introduction
Growth is a characteristic of life on Earth and also perhaps of life elsewhere (Schulze-Makuch & Irwin, Reference Schulze-Makuch and Irwin2008). Growth rate can be quantified using cardinal temperatures such as T min and T max, but this usage necessarily imposes a binary condition, that of growth or non-growth. A more productive approach is to consider growth as a continuous quantity. While agreeing that defining life remains problematic (Cleland & Chyba, Reference Cleland and Chyba2002), growth rate pragmatically represents a measurable, accessible and quantitative property of the volatility of life. By growth rate we mean the rate at which individuals reproduce, populations increase, or cells divide. We show in this paper that the concept of growth rate has considerable potential in astrobiology to understand the nature of life. Processes involved in life on Earth can occur on a variety of scales from milliseconds for enzyme catalysed reactions (Stockbridge et al. Reference Stockbridge, Lewis, Yuan and Wolfenden2010) to more than 3.5 billion years for life to reach the current level of adaptive evolution (McMeekin et al. Reference McMeekin, Olley, Ratkowsky, Corkrey and Ross2013). In this study, we are concerned with the rates at which individual organisms replace themselves. Growth, in the form of reproduction, a necessary condition for Darwinian evolution, occurs over temperature ranges narrower than that for survival (Clarke, Reference Clarke2014). In this paper, we discuss the possible distribution of growth rates, and whether the growth rate of life has a maximum limit, leaving aside consideration of a possible lower limit to a later time.
Single strain growth rates
Growth rate is usually quantified by specific or intrinsic growth rates (Birch, Reference Birch1948; McMeekin et al. Reference McMeekin, Olley, Ross and Ratkowsky1993). An example of a typical growth rate curve is shown in Fig. 1 for a single microbial strain. The lowest and highest temperatures at which growth is observed or is theoretically possible are usually termed T min and T max. The temperature at which the maximum specific growth rate is reached is T opt after which growth declines. Approximately halfway between T min and T opt is T mes, at which the cell's maintenance requirements are minimized. The T mes is estimated using a thermodynamic model for growth, described below.

Fig. 1. Example of a growth curve for a single microbial strain. The black circles (•) are the observed temperature-dependent growth rates for Methanococcoides burtonii. Also shown are four cardinal temperatures, T
min, T
mes, T
opt and T
max using blue diamonds (![]() ) and a fitted curve obtained from a mechanistic thermodynamic model (see text.) The T
min and T
max are observed limits in this case.
) and a fitted curve obtained from a mechanistic thermodynamic model (see text.) The T
min and T
max are observed limits in this case.
Thermodynamic model
The fitted curve shown in Fig. 1 is obtained from a mechanistic model that is based on the thermodynamic properties of protein folding (Corkrey et al. Reference Corkrey, Olley, Ratkowsky, McMeekin and Ross2012). This model assumes that the temperature-dependent growth rate of the organism is limited by a single rate-limiting Master Reaction System (MRS). While the cell machinery affecting cell growth may be extremely complex, the model's assumption is that there is a single process that is slower than the rest and which acts to throttle cell activity and this allows the growth rate of the whole cell to be more simply modelled.
Since the model has been described in detail elsewhere (Ross, Reference Ross1993, Reference Ross, Dodd and Gianinazzi-Pearson1997; Ratkowsky et al. Reference Ratkowsky, Olley and Ross2005; Corkrey et al. Reference Corkrey, Olley, Ratkowsky, McMeekin and Ross2012, Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2014) we only give a brief summary here. The fundamental equation of the model is shown in equation (1). In the equation, the quantity F is the predicted rate given the temperature and the values of the parameters. The numerator is essentially an Arrhenius model that describes the rate of the putative enzyme-catalysed rate-controlling reaction as a function of temperature, while the denominator models the change in expected rate due to the effects of temperature on the conformation and, hence, catalytic activity of the putative enzyme catalysing that reaction.
 $$F = \displaystyle{{T\exp \left( {c - \displaystyle{{\Delta H_{\rm A}^{\rm \ddag}} \over {RT}}} \right)} \over {1 + \exp \left( { - n\displaystyle{{\Delta H^{\rm \star} - T\Delta S^{\rm \star} + \Delta C_{\rm P} \left( {T - T_{\rm H}^{\rm \star} - T\ln \left( {\displaystyle{T \over {T_{\rm S}^{\rm \star}}}} \right)} \right)} \over {RT}}} \right)}}.$$
$$F = \displaystyle{{T\exp \left( {c - \displaystyle{{\Delta H_{\rm A}^{\rm \ddag}} \over {RT}}} \right)} \over {1 + \exp \left( { - n\displaystyle{{\Delta H^{\rm \star} - T\Delta S^{\rm \star} + \Delta C_{\rm P} \left( {T - T_{\rm H}^{\rm \star} - T\ln \left( {\displaystyle{T \over {T_{\rm S}^{\rm \star}}}} \right)} \right)} \over {RT}}} \right)}}.$$
In equation (1), ln is the natural logarithm function, R is the gas constant [8.314 J (K mol)−1]; c is a scaling constant that also incorporates the Boltzmann and Planck constants;
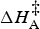 $\Delta H_{\rm A}^{\rm \ddag} $
is the enthalpy of activation (J mol−1); T is the temperature in degrees Kelvin; ΔC
P is the heat capacity change [J per (K mol amino acid residue)] upon denaturation of the rate-controlling reaction; n is the number of amino acid residues;
$\Delta H_{\rm A}^{\rm \ddag} $
is the enthalpy of activation (J mol−1); T is the temperature in degrees Kelvin; ΔC
P is the heat capacity change [J per (K mol amino acid residue)] upon denaturation of the rate-controlling reaction; n is the number of amino acid residues;
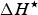 $\Delta H^{\rm \star} $
is the enthalpy change (J mol−1 amino acid residue) at
$\Delta H^{\rm \star} $
is the enthalpy change (J mol−1 amino acid residue) at
 $T_{\rm H}^{\rm \star} $
, the convergence temperature for enthalpy (K) of protein unfolding;
$T_{\rm H}^{\rm \star} $
, the convergence temperature for enthalpy (K) of protein unfolding;
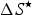 $\Delta S^{\rm \star} $
is the entropy change (J K−1 mol amino acid residue) at
$\Delta S^{\rm \star} $
is the entropy change (J K−1 mol amino acid residue) at
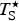 $T_{\rm S}^{\rm \star} $
, the convergence temperature for entropy (K) of protein unfolding. The starred notation follows that of Murphy et al. (Reference Murphy, Privalov and Gill1990) except that
$T_{\rm S}^{\rm \star} $
, the convergence temperature for entropy (K) of protein unfolding. The starred notation follows that of Murphy et al. (Reference Murphy, Privalov and Gill1990) except that
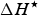 $\Delta H^{\rm \star} $
,
$\Delta H^{\rm \star} $
,
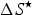 $\Delta S^{\rm \star} $
and ΔC
P are expressed per mole of amino acid residue and we introduce the n parameter (Ratkowsky et al.
Reference Ratkowsky, Olley and Ross2005). The parameters,
$\Delta S^{\rm \star} $
and ΔC
P are expressed per mole of amino acid residue and we introduce the n parameter (Ratkowsky et al.
Reference Ratkowsky, Olley and Ross2005). The parameters,
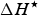 $\Delta H^{\rm \star} $
and
$\Delta H^{\rm \star} $
and
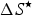 $\Delta S^{\rm \star} $
, which are discussed elsewhere with a change in notation (i.e. ΔH
u and ΔS
u) (Makhatadze & Privalov, Reference Makhatadze and Privalov1993; Privalov & Makhatadze, Reference Privalov and Makhatadze1993), arise in the context of enthalpy–entropy compensation (Privalov & Gill, Reference Privalov and Gill1988). We follow Baldwin (Reference Baldwin1986) in assuming
$\Delta S^{\rm \star} $
, which are discussed elsewhere with a change in notation (i.e. ΔH
u and ΔS
u) (Makhatadze & Privalov, Reference Makhatadze and Privalov1993; Privalov & Makhatadze, Reference Privalov and Makhatadze1993), arise in the context of enthalpy–entropy compensation (Privalov & Gill, Reference Privalov and Gill1988). We follow Baldwin (Reference Baldwin1986) in assuming
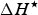 $\Delta H^{\rm \star} $
and
$\Delta H^{\rm \star} $
and
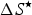 $\Delta S^{\rm \star} $
to be constants for hydrocarbons and also Privalov & Gill (Reference Privalov and Gill1988) in assuming
$\Delta S^{\rm \star} $
to be constants for hydrocarbons and also Privalov & Gill (Reference Privalov and Gill1988) in assuming
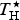 $T_{\rm H}^{\rm \star} $
and
$T_{\rm H}^{\rm \star} $
and
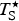 $T_{\rm S}^{\rm \star} $
are universal properties of proteins, although they use a different notation.
$T_{\rm S}^{\rm \star} $
are universal properties of proteins, although they use a different notation.
The thermodynamic model has successfully and accurately modelled the relative growth rates of strains in all three Domains of life (Bacteria, Archaea and Eukarya) ranging from psychrophilic to hyperthermophilic up to 122°C (Corkrey et al. Reference Corkrey, Olley, Ratkowsky, McMeekin and Ross2012). It also well-described relative growth rates for multicellular poikilothermic organisms and obtained values of thermodynamic parameters and relationships between them that, for the most part, were consistent with expectations from biochemistry (Fields, Reference Fields2001; Corkrey et al. Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2014). It provided excellent fits to absolute rates of 694 strains of unicellular and multicellular organisms (Corkrey et al. Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2014).
The observation that the thermodynamic model successfully fitted strains from all three Domains, and that the model parameter estimates appeared plausible, provided strong support for the MRS. This implies that the MRS, if real, must have been strongly conserved since the time of the last universal common ancestor (LUCA) to all three domains (Becerra et al. Reference Becerra, Delaye, Islas and Lazcano2007). A corollary is that, as a consequence of evolutionary contingencies, life that arose elsewhere or evolved separately would not necessarily be expected to possess the same MRS.
The biokinetic spectrum for temperature (BKST)
As is well known, life on Earth spans a range of temperatures and exhibits biological growth rates that are temperature-dependent. However, we have recently shown that the statistical distribution of specific growth rates for life on Earth is a function of temperature (Corkrey et al. Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2016). The plot of this distribution, shown in Fig. 2, contained a wide range of growth rates from all three domains of life, Bacteria, Archaea and Eukarya, and spanned the entire temperature range over which life is known to grow. These growth rates form the BKST. The BKST was assembled from data that were generated by ourselves and colleagues, or obtained by searching the scholarly literature. The BKST is described in more detail elsewhere (Corkrey et al. Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2016) and we only give a brief summary here.

Fig. 2. The BKST. Shown are a total of 11 130 observations of 1654 strains/species collated from 849 literature sources and spanning 45 phyla. This figure is based on Fig. 1 of Corkrey et al. (Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2016).
We use the word strain, rather than species or taxa, because some datasets are of a single species grown under different conditions, or the same species grown by different researchers. The data were intended to represent the full range of growth rates possible for life on Earth and accordingly we did not restrict which strains or species were incorporated. The data were not intended to represent a random sample of strains, but to include as wide a range of strains as possible. The strains were all culturable and often of economic, veterinary, agricultural or medical importance. Usually, strains were cultured in isolation, which meant that any synergistic effect resulting from ecological interactions were not included. However, the upper boundary of the BKST was very well defined suggesting that it did represent a limiting rate that evolution has not been able to breach. It seemed unlikely that a combination of multiple strains, such as are found in biofilms, could exceed this maximum limit. The distinctiveness of the BKST and the sharpness of its upper boundary also suggested that we were not likely missing many fast-growing strains. Therefore, we consider that the locus of the points forming the upper boundary represents the maximum limit to the rate of growth of life on Earth.
The upper boundary of the BKST exhibits a rapid curvilinear rise starting from a lower limit that begins below 0°C and reaches a surprisingly sharp peak at about 42°C. We previously defined a new cardinal temperature to refer to the maximum observed growth rate called T sup, where ‘sup’ was short for supremum (Fig. 3). T sup represents the temperature at which the highest known growth rate occurs. At temperatures above T sup the maximum limit declines to eventually approach zero above 120°C. We termed the boundary for the maximum rates at temperatures below T sup the ‘ascending curve’, and the boundary above it the ‘descending curve’. The ascending curve has some similarity to the Eppley curve (Eppley, Reference Eppley1972), which describes the maximum rate of growth of marine plankton but the whole of the Eppley curve is contained within the BKST. At about 50°C there appeared to be a gap that interrupts the descending curve, which we termed the Mesophile–Thermophile gap (MTG) since it lay between the two thermal groups. We previously presented evidence (Corkrey et al. Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2016) that the MTG was a real phenomenon and not the result of a sampling insufficiency. Should that prove to be the case then the BKST has two peaks, one at T sup and a secondary peak at about 65°C.

Fig. 3. Features of the BKST. The principal features are the mesophile–thermophile gap, the temperature of the maximum growth rate, T sup, and a possible secondary peak at about 65°C. We also show the maximum rates limits using dashed lines that are drawn ‘by eye’. See the text for details. This figure is based on Fig. 1 of Corkrey et al. (Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2016).
In our previous work, we did not attempt to estimate the maximum limit of the BKST. The subject of this paper is to estimate that limit, which is the maximum possible growth rate that can be expected for any form of life on this planet. We then discuss applications of the BKST to astrobiology.
Describing the BKST
We used Bayesian quantile regression to quantitatively describe the BKST. A quantile is a quantity below which a specific proportion of data fall. The quantile regression required fitting the function, repeated here from Corkrey et al. (Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2016) in equation (2), except that we change notation slightly by replacing T sup by T s. The function we used had four parameters, a, b, c, d, to which we assigned simple non-informative Gaussian priors. The r(T|a, b, c, d) is the predicted growth rate at temperature T given the four parameters. There was a separate set of the parameters for each quantile fitted. Additional details on the fitting methods for the quantile regression are given in Corkrey et al. (Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2016). The upper quantiles represent the highest growth rates at each temperature, below which are strains that grow at varying, but slower, rates. We refer to the fitted lines as quantile curves.
 $$\eqalign{r(T \vert a,b,c,d) & = \left\{ {\matrix{ {\exp (a + b \times T)} & {T \le T_{\rm s} {\rm,} \,b \gt 0,} \cr {\exp (c + d \times T)} & {T \gt T_{\rm s} {\rm,} \,d \lt 0,} \cr}} \right. \cr & {\rm where} \,T_{\rm s} = \displaystyle{{a - c} \over {d - b}}.}$$
$$\eqalign{r(T \vert a,b,c,d) & = \left\{ {\matrix{ {\exp (a + b \times T)} & {T \le T_{\rm s} {\rm,} \,b \gt 0,} \cr {\exp (c + d \times T)} & {T \gt T_{\rm s} {\rm,} \,d \lt 0,} \cr}} \right. \cr & {\rm where} \,T_{\rm s} = \displaystyle{{a - c} \over {d - b}}.}$$
Linking the BKST and the MRS
Our aim in this paper is not to define the lower and upper temperature limits, but to find the maximum rate of growth of life on Earth. We also consider if the maximum rate of growth should also be expected for life that evolved elsewhere. While the rate of growth may not determine the limiting temperatures it may help to define those limits. In other words, what is the fastest possible growth rate we can expect at any particular temperature? We show that this is obtainable from the BKST.
Since the mechanistic thermodynamic model described the temperature-dependent growth rate for individual species it was hypothesized that the BKST might be derivable from it. We have previously shown that this was possible (Corkrey et al. Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2016) and argued that the BKST could be interpretable as arising from an evolutionary trade-off between protein activity and stability with changing temperature (Fields, Reference Fields2001; Dick et al. Reference Dick, Weiergräber, Classen, Bisterfeld, Bramski, Gohlke and Pietruszka2016). In order to link the thermodynamic model with the BKST we calculated growth curves for those strains with sufficient data and used non-linear Bayesian quantile regression to construct a series of quantiles. The methods used are described in detail in Corkrey et al. (Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2016) and we only give a brief summary here. The fitted growth curves and quantile curves are shown together in Fig. 11 of Corkrey et al. (Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2016). We calculated the means of the thermodynamic parameter values for those strains that had growth curves that exceeded a given quantile curve. The trends displayed by these mean estimates against temperature were consistent with observations on biomolecules (Fields, Reference Fields2001). By assuming these estimates within the thermodynamic model we could recover the BKST, as shown in Fig. 13 of Corkrey et al. (Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2016). This meant that the thermodynamic model was at least consistent with the BKST.
This led to the following argument: the thermodynamic model successfully fitted strains from all three domains and obtained parameter estimates consistent with biochemical expectations, which provided strong support for the MRS to have been conserved since the LUCA. The BKST could be recovered based on trends in the thermodynamic parameters that were consistent with biochemical expectations. This implied the BKST was at least consistent with the MRS. It also implied that if life did not share the same evolutionary history as life on Earth and, as a result, not possess the same MRS, it might not display the same BKST. In other words, life that evolved independently from life on Earth might be distinguishable from it by means of the BKST. It may be that another mechanism is actually responsible for the BKST, but since all three domains of life appear to be similarly bounded the putative mechanism would still have to have been strongly conserved since the LUCA.
Below we extend the analysis of the BKST to estimate the maximum possible growth rate, and then discuss applications to astrobiology.
Methods
We used the same data as in Corkrey et al. (Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2016) along with a small amount of additional data (n = 9): Larkin & Stokes (Reference Larkin and Stokes1968); Breezee et al. (Reference Breezee, Cady and Staley2004); Mykytczuk et al. (Reference Mykytczuk, Foote, Omelon, Southam, Greer and Whyte2013). We used non-linear Bayesian quantile regression to calculate quantile curves to represent the quantiles 50, 51, ..., 99% using two alternative functions. We refer to a model that uses the function given by equation (2) as the single peak model; this was the same model used in Corkrey et al. (Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2016). The second is referred to herein as the double peak model and it uses the alternative function shown in equation (3). The single peak model did not take the MTG into account, while the double peak model did so by fitting a pair of exponential back-to-back functions. One set of functions corresponded to the peak at T sup and the others to the secondary peak, which is located at about 65°C. The fitting methods used for both were the same as in Corkrey et al. (Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2016) except that we relaxed the assumption of non-informative priors, as described further below. As described in Corkrey et al. (Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2016), inference for both models was obtained in the form of posterior means and variances using Markov Chain Monte Carlo (MCMC) simulation (Brooks, Reference Brooks1998) and which was implemented with Fortran 95.
 $$s(T) = {\rm max}\left[ {r(T \vert a_1, b_1, c_1, d_1 ),r(T \vert a_2, b_2, c_2, d_2 )} \right].$$
$$s(T) = {\rm max}\left[ {r(T \vert a_1, b_1, c_1, d_1 ),r(T \vert a_2, b_2, c_2, d_2 )} \right].$$
In equation (3), r is the function defined in equation (2). a 1, b 1, c 1, d 1, a 2, b 2, c 2 and d 2 are the parameters to be estimated. Since the r function appears twice there are two occurrences of T s, one of which corresponds to the peak at T sup and the other to the secondary peak shown in Fig. 3.
While quantile regression allows the distribution of the data to be modelled, the quantiles are always confined between 0 and 100% and they do not attain these limits. This means that quantile regression cannot directly allow the estimation of the maximum limit to growth. Examination of the BKST shown in Fig. 3 indicated that a maximum limit both existed and had a particular form. We approximated these maximum rates by visual inspection, shown by dashed lines in the figure. However, these are subjective estimates that are drawn ‘by eye’. To obtain more objective fits we developed the following method. We noted after some exploratory data analysis, that the (a, b, c, d) parameters of equation (2) displayed quadratic trends, particularly for the lower quantiles. This suggested that a quadratic polynomial curve could be extrapolated to estimate the values the a, b, c and d parameters could attain at quantile 100%. To do this we replaced the simple non-informative priors by those shown in equation (4), so that the (a, b, c, d) parameters now became functions of the quantile q. The α0, α1, … , δ2 were assigned non-informative Gaussian priors each with a zero mean and with the variance set to 1000. Any risk due to extrapolating beyond the range of observed data was moderated by the use of a quadratic function rather than a higher degree polynomial or spline that would have been more likely to have behaved erratically. The arcsine transformation [equation (5)] was used since the quantile value remained bound between 0 and 100%. The posterior estimates of the α0, α1, … , δ2 could then be used to estimate the values of a, b, c, d at any quantile and to extrapolate the quadratic curve to estimate the values that the parameters would attain at the 100% quantile.
 $$\matrix{ {Pr(a)\,{\rm \sim}\, N{\rm (\alpha} _{\rm 0} + {\rm \alpha} _1 \times {\rm \theta} + {\rm \alpha} _2 \times {\rm \theta} ^2, {\rm \tau} _{\rm a} {\rm ),}} \hfill \cr {Pr(b)\,{\rm \sim}\, N{\rm (\beta} _0 + {\rm \beta} _1 \times {\rm \theta} + {\rm \beta} _2 \times {\rm \theta} ^2, {\rm \tau} _{\rm b} {\rm ),}} \hfill \cr {Pr(c)\,{\rm \sim}\, N{\rm (\gamma} _0 + {\rm \gamma} _1 \times {\rm \theta} + {\rm \gamma} _2 \times {\rm \theta} ^2, {\rm \tau} _{\rm c} {\rm ),}} \hfill \cr {Pr(d)\,{\rm \sim}\, N{\rm (\delta} _0 + {\rm \delta} _1 \times {\rm \theta} + {\rm \delta} _2 \times {\rm \theta} ^2, {\rm \tau} _{\rm d} {\rm ),}} \hfill \cr} $$
$$\matrix{ {Pr(a)\,{\rm \sim}\, N{\rm (\alpha} _{\rm 0} + {\rm \alpha} _1 \times {\rm \theta} + {\rm \alpha} _2 \times {\rm \theta} ^2, {\rm \tau} _{\rm a} {\rm ),}} \hfill \cr {Pr(b)\,{\rm \sim}\, N{\rm (\beta} _0 + {\rm \beta} _1 \times {\rm \theta} + {\rm \beta} _2 \times {\rm \theta} ^2, {\rm \tau} _{\rm b} {\rm ),}} \hfill \cr {Pr(c)\,{\rm \sim}\, N{\rm (\gamma} _0 + {\rm \gamma} _1 \times {\rm \theta} + {\rm \gamma} _2 \times {\rm \theta} ^2, {\rm \tau} _{\rm c} {\rm ),}} \hfill \cr {Pr(d)\,{\rm \sim}\, N{\rm (\delta} _0 + {\rm \delta} _1 \times {\rm \theta} + {\rm \delta} _2 \times {\rm \theta} ^2, {\rm \tau} _{\rm d} {\rm ),}} \hfill \cr} $$
where
 $${\rm \theta} = {\rm arcsin}\left( {\sqrt q} \right).$$
$${\rm \theta} = {\rm arcsin}\left( {\sqrt q} \right).$$
We calculated the predicted means for the (a, b, c, d) parameters using equation (4) by setting θ = π/2. We then calculated the maximum rate of growth by setting the corresponding parameters within equations (2) and (3) to the predicted means.
As we describe later, while the BKST was well behaved between 0 and 100°C it probably overestimated below or above this range when compared with the existing data. For temperatures below 0°C we conjectured that growth rates would be attenuated below the predicted rate proportionally with decrease in temperature until reaching zero growth at a particular temperature. We borrowed this conjecture from a model for the dependence of growth on water activity for temperatures above 0°C (McMeekin et al. Reference McMeekin, Chandler, Doe, Garland, Olley, Putro and Ratkowsky1987). Water activity is defined as the ratio of vapour pressure of water to that of ice (Reid & Fennema, Reference Reid, Fennema, Damodaran, Parkin and Fennema2007) and can be interpreted as the availability of water for reactions. Since water activity for pure super-cooled water reduces with temperature below 0°C (Reid & Fennema, Reference Reid, Fennema, Damodaran, Parkin and Fennema2007) we adjusted the maximum rate of growth by multiplying by a factor a W − W, where a W was the water activity at a given temperature and W was set at 0.755, the lowest known water activity common to the three domains of life (Stevenson et al. Reference Stevenson2015). For very high temperatures (above 95°C) we calculated the reduction in the predicted maximum rate of growth under the assumption that rates would be reduced relative to the rate at 95°C in proportion to the in vitro half-life of ATP (Daniel et al. Reference Daniel, van Eckert, Holden, Truter, Crowan, Wilcock, Delong, Kelley, Baross and Cary2004) or Ala-tRNA (Stepanov & Nyborg, Reference Stepanov and Nyborg2002). The adjusted rates assume that the cell was unable to compensate for the hydrolysis and the choice of 95°C was arbitrary. The rates at these very high temperatures adjusted for hydrolysis are shown later for illustrative purposes, not as predictions of actual rates.
To compare subgroups we calculated the peak heights and the areas under the 95% quantile curves and the maximum limit curves. The subgroups considered were aerobes, anaerobes, heterotrophs and autotrophs as well as combinations of them. Allocation to subgroups was based on either the source literature from which the data were obtained or from original species descriptions. Since the maximum growth curves were unstable for the small sample sizes that arose in some cases we only considered further the areas under the 95% quantile curves. Where the observed data clearly displayed only one peak we fitted the single peak model, and where this was not clear we chose the model that resulted in the smaller areas below the curves, as calculated by numerically integrating the formulas (2) and (3) using the posterior estimates of the parameters. We contrasted the various subgroups, such as autotrophs and heterotrophs, by calculating the posterior probability that the maximum growth rate curve for one encompassed an area less than the other. It is to be expected that the limiting rates of some subgroups will have lower rates than the fastest growing strains.
Results
The predicted maximum growth rate
In Fig. 4, we show the predicted values for the (a, b, c, d) parameters calculated from the posterior estimates of the α0, α1, … , δ2. The fitted curves terminated at values very close to those we estimated ‘by eye’ in Fig. 3. This suggested that our estimation procedure for the maximum possible growth rate was plausible. Using the estimated (a, b, c, d) parameters for the 100% quantile we could calculate the maximum growth rates.

Fig. 4. Predicted quantile curve parameters (a, b, c, d) versus quantile. Shown are the quantile curve parameters for varying quantiles estimated from the posteriors of α0, …, δ2. Also shown as green triangles (![]() ) are the estimated values of the parameters obtained by the dashed curves drawn ‘by eye’ in Fig. 3.
) are the estimated values of the parameters obtained by the dashed curves drawn ‘by eye’ in Fig. 3.
We show the maximum growth rates for the single- and double peak models in Fig. 5. The parameter values for curves are given in Table 1. In Table 2, we show predicted minimum generation times between −20°C, a proposed lower limit for growth (Clarke et al. Reference Clarke, Morris, Fonseca, Murray, Acton and Price2013), and 130°C, the highest known temperature at which cells remain viable (Takai et al. Reference Takai, Nakamura, Toki, Tsunogai, Miyazaki, Miyazaki, Hirayama, Nakagawa, Nunoura and Horikoshi2008). We found that using the single peak model the minimum generation time (or maximum growth rate) occurred at T sup = 45.8 (95% CI 43.2, 49.2) which appeared slightly to the right of the visual peak (Fig. 3). In the case of the double peak model, which allows for the MTG, the minimum generation time occurred at almost the same temperature of T sup = 45.6 (95% CI 38.7, 51.6). The double peak model obtained wider credible bands than the single peak model. The double peak model had a second peak that occurred at 64.2 (95% CI 56.0, 72.2) and a distinct gap was also apparent in the fitted curve. The secondary peak had a surprisingly high growth rate, but high growth rates were observed at about 80–100°C, which render this plausible. Table 2 also shows the predicted generation times after adjustment for hydrolysis and water activity.

Fig. 5. Predicted maximum rate of growth for the single- and double peak models. Shown are the observed growth rates as dots, the predicted maximum growth rate (solid line) with 95% CI (shaded area), and adjusted maximum rates for water activity (left red solid line) and in vitro hydrolysis of ATP (right red solid line), and in vitro hydrolysis of Ala-tRNA (right red dashed line). Left: single peak model. Right: double peak model.
Table 1. Growth curve parameters of the maximum predicted growth rates

Shown are the posterior means and credible intervals for the four parameters for the single peak model and the two sets of four parameters for the double peak model.
Table 2. Estimated minimum generation times at selected temperatures

Units for generation times are minutes except when adjusted for water activity or ATP hydrolysis when they are in days. Results are shown for the single- and double peak models that allow for the MTG. We show both unadjusted and adjusted generation times for temperatures below 0°C. Adjustment is made for water activity below 0°C and ATP hydrolysis above 95°C.
a Adjusted generation times in days extrapolated using water activity of pure water.
b Lowest temperature in our data.
c T sup.
d Highest temperature in our data.
e Adjusted generation times in days extrapolated using ATP hydrolysis.
f Middle of the MTG.
g Secondary peak.
In Fig. 6, we show the 95% quantile growth rate curves for the subgroups aerobes, anaerobes, heterotrophs and autotrophs, as well as combinations of them. Overall, anaerobes obtained a larger area under their 95% quantile growth rate curve than that of aerobes (posterior probability = 1.00), and heterotrophs obtained a larger area than autotrophs (posterior probability = 1.00). Comparing the areas under the 95% quantile growth rate curves for subgroups, anaerobic heterotrophs obtained a larger area than aerobic heterotrophs (posterior probability = 1.00), anaerobic autotrophs obtained a larger area than aerobic autotrophs (posterior probability = 1.00), aerobic heterotrophs obtained a larger area than aerobic autotrophs (posterior probability = 0.98), and anaerobic heterotrophs obtained a larger area than anaerobic autotrophs (posterior probability = 0.98).

Fig. 6. 95% quantile curves by aerobic and trophic status. Shown are 95% quantile growth curves calculated separately for subgroups of strains based on their aerobic and trophic statuses. The 99% credible limits are shown as dashed lines. Where it was unclear whether to fit the one-peak or the two-peak model, the model that obtained the smallest area was retained and shown in the figure. (a) aerobic heterotrophs; (b) aerobic autotrophs; (c) all aerobes; (d) anaerobic heterotrophs; (e) anaerobic autotrophs; (f) all anaerobes; (g) all heterotrophs; (h) all autotrophs; (i) all strains.
In Table 3, we compare the maximum rate for various subgroups. Overall, these comparisons revealed that anaerobes obtained a higher peak rate than aerobes, and heterotrophs obtained a higher peak rate than autotrophs. The same tendencies are apparent in the individual cells of the table.
Table 3. Areas and maximum rates under the 95% quantile curve for subgroups

Shown are areas (degree/minute) under the curves and the peak rates for each, as shown in Fig. 6. The 99% credible intervals are shown in parentheses.
Discussion
Consideration has often been given in astrobiology to the physicochemical limits within which life is known to exist, including temperature, pH, salinity and so on (Space Studies Board, 2007). In the case of temperature, these are usually the cardinal temperatures T min, T opt and T max for individual species along with the minimum and maximum survival and growth limits for life. For example, Harrison et al. (Reference Harrison, Gheeraert, Tsigelnitskiy and Cockell2013) described the ‘parametric volume’ defined by the limits of stressors. We suggest that the use of cardinal temperatures ignores the considerable information represented by growth rates. Growth is thought to be a universal characteristic of life (Schulze-Makuch & Irwin, Reference Schulze-Makuch and Irwin2008) and growth rates are an obvious metric of growth. For these reasons, we concern ourselves here with growth rates and the uses that may be made of them in astrobiology.
Before proceeding we need to distinguish between rate limits and temperature limits. Discussions on limits of growth usually concern temperature extremes, but in this paper we deal with both types of limits, although we are more concerned with the maximum rate limits. But since these are related to temperature limits we describe these by investigating how temperature limits can be related to growth rates at the extremes of temperatures over which life on Earth grows.
Our estimates of maximum rates can serve to provide an envelope for what is possible. The maximum rate limits, or equivalently, the minimum generation times, that we describe in this paper, are obtained under favourable conditions. Actual rates achievable by life may be expected to be less, and generation times longer, particularly where extremes of temperatures are combined with other stressor conditions. For example, microbes in some permafrost soils from University Valley, Antarctica, are not just limited by extreme cold, but also by aridity and oligotrophy, which combine to severely constrain the evolution of functional cold-adapted organisms (Goordial et al. Reference Goordial, Davila, Lacelle, Pollard, Marinova, Greer, DiRuggiero, McKay and Whyte2016), and presumably the habitability of other similar environments.
Linking the MRS and the BKST
The BKST represents the distribution of growth rates that are possible for life on Earth. It bears a distinctive shape (Fig. 3) that requires explanation. We have previously argued (Corkrey et al. Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2016) that it arises from an evolutionary trade-off between stability and activity of the putative protein (or biomolecule) that composes the MRS. This assumption allowed us to recover the shape of the BKST by using trends in the thermodynamic parameters of the MRS that were interpretable in terms of protein biochemistry. We do not claim that the BKST necessarily arises from the thermodynamic model, but that the form of the spectrum is not inconsistent with it, and that it is potentially understandable on the basis of protein denaturation. While the sharpness of the upper edge of the BKST provides strong evidence in favour of the MRS, the actual mechanism that determines growth rate may be more complex than a single process; for example, it may arise from an optimization of metabolic processes (Lewis et al. Reference Lewis2010). But, if so, it would be difficult to understand why there should be a maximum rate that is bounded by a pair of simple exponential curves.
By using the posterior estimates obtained from the quadratic relationship applied to the priors of the quantile curve parameters, we were able to extrapolate the trends of the quantile curves in order to estimate the maximum rate of growth. This required us to extrapolate beyond the observed data range, but the form of the BKST suggests that there is a distinct maximum limit that should, in principle, be estimable. This was particularly clear for temperatures between 0°C and T sup in which the ascending curve was sharply defined. The descending curve was less well defined than the ascending curve, however, it was obvious that the maximum rate at, say, 60°C was greater than at 70°C, and the rate at 70°C greater than at 80°C and so on. The agreement of the trends of the quantile curve parameters with our visual approximation of the maximum rate curve also supported the feasibility of our estimation.
The predicted maximum growth rate
The posterior maximum rate curve tracked the fastest growing organisms (Fig. 5). There were only a few points noticeably above the mean curve but these were all included within the 99% credible bands for both models. The predicted maximum rate curve remained noticeably above the observed data at temperatures above 100°C, although the lower credible band came close to the observed data, and the credible band for the double peak model included them. The rate of decline for the ascending curve had a much narrower credible band than the descending curve, which indicated a greater level of support by the data for the former. For both models the ascending curve rose above all the observed data at temperatures below 0°C. We discuss these points further below.
We reported predicted minimum generation times at T sup and at very low or high temperatures (Table 2). Observed generation times for specific species have been reported for the fastest growth rates, or for growth at extreme temperatures, but we are unaware of previous attempts that have examined systematically the maximum growth rate limit. We found that the predicted minimum generation time was 5.16 min for the single peak model. This appears plausible in comparison with the fastest growing organism known, Clostridium perfringens which has a generation time of 6.3 min. Another very fast-growing organism, Vibrio natriegens, may also have a generation time below 7 min (Maida et al. Reference Maida, Bosi, Perrin, Papaleo, Orlandini, Fondi, Fani, Wiegel, Bianconi and Canganella2013), although they did not report a temperature.
The double peak model better reflected the shapes of the BKST, but its credible bands were much wider. However, the posterior mean predicted curves for the two models were almost coincident for the ascending curves. They deviated at T sup with the double peak model peak being slightly lower. The descending curves for the two models converged at temperatures higher than about 80°C. In summary, apart from temperatures within the MTG and the secondary peak the two models produced similar predictions.
It appears quite plausible that there may be a link between the maximum rate limit and cell size (Iyer-Biswas et al. Reference Iyer-Biswas, Wright, Henry, Lo, Burov, Lin, Crooks, Crosson, Dinner and Scherer2014), as well as resource transport through the cell membrane (Hoehler, Reference Hoehler2004). Fast growing Kluyveromyces marxianus cells have been reported to be larger in size than slower-growing cells by becoming more elongate (Groeneveld et al. Reference Groeneveld, Stouthamer and Westerhoff2009), while cell volumes of planktonic microbes (Chrzanowski et al. Reference Chrzanowski, Crotty and Hubbard1988; Atkinson et al. Reference Atkinson, Ciotti and Montagnes2003), protozoa (James & Read, Reference James and Read1957) and diatoms (Montagnes & Franklin, Reference Montagnes and Franklin2001) have been reported to decrease, sometimes with cells becoming more elongate (Sjöstedt et al. Reference Sjöstedt, Hagström and Zweifel2012), with increasing temperature. In these cases, the increase in surface area to volume ratio implies that growth rates are modulated by membrane transport. However, these studies on relationships between cell division rate and cell size considered exponential growth (e.g. Iyer-Biswas et al. Reference Iyer-Biswas, Wright, Henry, Lo, Burov, Lin, Crooks, Crosson, Dinner and Scherer2014), whereas in this study the descending curve would appear to be more likely related to supraoptimal effects. Fast-growing cells presumably plausibly require an optimized transcription apparatus. Thermophilic bacteria have smaller genomes than non-thermophilic bacteria (Sabath et al. Reference Sabath, Ferrada, Barve and Wagner2013), although generation time does not correlate with genome size (Mira et al. Reference Mira, Ochman and Moran2001), and fast growth within species has been associated with reduced codon usage bias (Karlin et al. Reference Karlin, Mrázek, Campbell and Kaiser2001; Vieira-Silva & Rocha, Reference Vieira-Silva and Rocha2010). Such mechanisms may be correlated with fast growth, but they do not explain why the BKST has its double-exponential profile. When viewed from the broader perspective of the whole biokinetic temperature range, we suggest a plausible explanation lies in the proposed trade-off between activity and stability of enzymes involved in a rate-limiting MRS within the cell.
Subzero growth rates
It is not clear what the limiting mechanisms for growth below 0°C may be, but the challenges presented by low temperatures include osmotic stress, reduced membrane fluidity (Bakermans, Reference Bakermans and Anitoris2012; Clarke, Reference Clarke2014), reduced enzyme catalysis and the availability of liquid water (Bakermans, Reference Bakermans and Anitoris2012). These changes place increasing demands on cell maintenance as the temperature becomes more extreme (Fields, Reference Fields2001), increasing protein turnover to avoid disruption of proteostasis (Bednarska et al. Reference Bednarska, Schymkowitz, Rousseau and Van Eldere2013). For proteins in psychrophiles compared with mesophiles there are fewer ionic interactions, intramolecular hydrogen bonds are less numerous, and the numbers of polar or charged groups increase (Fields, Reference Fields2001).
The lowest temperature limits for growth or survival for life on Earth are not known although growth and activity at subzero temperatures have been reported. For growth, generation times for Bacillus species cultured in solutions containing glycerol or ethylene glycol have been reported of 4 days at −2°C, 7 days at −4.5°C, 9–11 days −5°C to −7°C (Larkin & Stokes, Reference Larkin and Stokes1968), for Psychromonas ingrahamii 10 days at −12°C (Breezee et al. Reference Breezee, Cady and Staley2004), and for Planococcus halocryophilus Or1 50 days at −15°C (Mykytczuk et al. Reference Mykytczuk, Foote, Omelon, Southam, Greer and Whyte2013). Food spoilage due to psychrotropic yeasts was reported at −18°C (Collins & Buick, Reference Collins and Buick1989). A generation time of 3.6 years at −22°C has been suggested following extrapolation of metabolic observations (Bakermans et al. Reference Bakermans, Tsapin, Souza-Egipsy, Gilichinsky and Nealson2003). DNA and protein synthesis continue at −17°C (Carpenter et al. Reference Carpenter, Lin and Capone2000), while metabolic activity has been detected at −20 and −25°C, respectively (Rivkina et al. Reference Rivkina, Friedmann, McKay and Gilichinsky2000; Mykytczuk et al. Reference Mykytczuk, Foote, Omelon, Southam, Greer and Whyte2013). Respiration continues in permafrost soils at −39°C (Panikov et al. Reference Panikov, Flanagan, Oechel, Mastepanov and Christensen2006) although this may have resulted from abiogenic processes (Bakermans, Reference Bakermans and Anitoris2012).
From such reports we should expect long generation times at subzero temperatures. However, the estimated generation times obtained by extrapolating the ascending curve appears too short to be plausible (Table 2): the single- and double peak models predict generation times at −20°C of 14.5 and 16.7 h, respectively. Clearly the model predictions are much shorter than the values reported in the scientific literature. Close examination of Fig. 5 indicated that the ascending curves in both models were above the observed data at subzero temperatures. If it is actually the case that growth rates drop drastically at 0°C when compared with the extrapolated ascending curve then this requires explanation.
Our results could arise from insufficient data or a model inadequacy. There were 66 data points at subzero temperatures obtained from 27 sources, which would appear a reasonable number. There may be organisms that grow faster than those of which we are aware. The data we have may reflect the difficulty of culturing at low temperatures since obtaining growth rates at such temperatures is very challenging, requires considerable patience due to the lengthy generation times involved, and it may be difficult to obtain optimal conditions. Another explanation is that abiotic stresses with corresponding biotic mechanisms become operative below 0°C. The estimated curve is largely informed by the bulk of the data that occurs above 0°C and the few data at negative temperatures would have little statistical influence on the curve. In this case, if other mechanisms operate at very low temperatures then the model assumption of a single exponential relationship with temperature will be incorrect.
In the studies by Larkin & Stokes (Reference Larkin and Stokes1968) and Breezee et al. (Reference Breezee, Cady and Staley2004) solutes such as glycerol, ethylene glycol and NaCl ensured the culture remained liquid. Both the freezing and melting temperatures of water can be depressed by increasing concentrations of solutes, such as NaCl. The precise identity of the solute is not important as long as the hydrogen bond networks between water molecules are similarly affected (Koop et al. Reference Koop, Luo, Tsias and Peter2000). At subzero temperatures liquid water can persist as thin films, such as around crystals, the thickness of which can constrain growth (Rivkina et al. Reference Rivkina, Friedmann, McKay and Gilichinsky2000). Such films are likely to contain fewer solutes. Pure supercooled water will freeze at 235 K (−38°C) and melt at 273 K (0°C) (Koop et al. Reference Koop, Luo, Tsias and Peter2000). Below 0°C the water activity of supercooled pure water declines monotonically (Reid & Fennema, Reference Reid, Fennema, Damodaran, Parkin and Fennema2007). We suggest that the mechanism that reduces growth rates below 0°C involves the bulk properties of water, such as water activity, that change progressively as the temperature drops. There are related factors that may influence growth rate within extreme environments such as those in Martian brines, including ionic strength, chaotropes and kosmotropes (Fox-Powell et al. Reference Fox-Powell, Hallsworth, Cousins and Cockell2016), but these are not considered further here. It is well known that water activity can be lowered by the addition of salts and other solutes (Chirife & Resnik, Reference Chirife and Resnik1984; Resnik & Chirife, Reference Resnik and Chirife1988) and has a strong effect on biological growth rates. Empirical modelling of growth rates above 0°C (McMeekin et al. Reference McMeekin, Chandler, Doe, Garland, Olley, Putro and Ratkowsky1987) has shown that growth rates decline linearly in proportion to a W − W, in which a W is the water activity and W is a species characteristic. According to this model a W can be considered as the water activity below which growth ceases. Growth has only been observed at a W ≥ 0.61 for fungi (Pitt & Christian, Reference Pitt and Christian1968) or a W ≥ 0.755 for Bacteria and Archaea (Stevenson et al. Reference Stevenson2015). We chose to use this model to adjust the predicted ascending curve to compensate for water activity below 0°C. To calculate adjusted growth rates we chose a W = 0.755 since this value is common to all domains, although it might underestimate the maximum possible rate possible for psychrophilic fungi.
As ice forms externally a cell will lose water osmotically until the vitrification temperature is reached. Vitrification occurs when liquids, such as water, are cooled below the melting temperature without crystallization, so that the molecules are retained in a disordered glass (Roos, Reference Roos2010). Between −10 and −25°C water usually vitrifies within cells unless they are cold-adapted (Clarke et al. Reference Clarke, Morris, Fonseca, Murray, Acton and Price2013). Below the vitrification temperature diffusion of oxygen and metabolites are inhibited. This may represent the lower limiting condition for cellular activity (Clarke et al. Reference Clarke, Morris, Fonseca, Murray, Acton and Price2013). Biological activity may continue to very low temperatures because enzyme activity slows but does not stop with decreasing temperature (Bragger et al. Reference Bragger, Dunn and Daniel2000) although it may be limited by diffusion (More et al. Reference More, Daniel and Petach1995).
In summary, we propose that, as the temperature decreases the maximum rate of growth reduces according to the BKST until 0°C is reached. At temperatures below 0°C growth may continue, but the bulk properties of water, such as water activity, attenuate growth rate until the vitrification temperature is reached. Below the vitrification temperature water enters a glassy state in which diffusion rates are too slow to support biological growth, but metabolic activity may persist until the freezing temperature is reached. Below the freezing temperature enzymes may continue to function, albeit slowly, but metabolic activity effectively ceases.
While we corrected the growth rate at temperatures below 0°C for water activity, we do not claim that this necessarily explains reduced subzero growth rates. The actual mechanism may be a different bulk property, such as reduced diffusion. Diffusion of molecules reduces as the temperature decreases to the vitrification temperature below which diffusion then decreases dramatically (Karel et al. Reference Karel, Anglea, Buera, Karmas, Levi and Roos1994; Clarke et al. Reference Clarke, Morris, Fonseca, Murray, Acton and Price2013). However, if the reduction in growth rates below 0°C results from decreasing water activity, then we suggest that halophiles will be progressively favoured at lower temperatures. Non-halophiles may grow below 0°C (Harrison et al. Reference Harrison, Dobinson, Freeman, McKenzie, Wyllie, Nixon and Cockell2015), but at sufficiently low temperatures we suggest that halophiles would grow more rapidly. Permafrost isolated bacteria have been shown to be able to survive low water activities (Ponder et al. Reference Ponder, Gilmour, Bergholz, Mindock, Hollingsworth, Thomashow and Tiedje2005).
Growth rates above 100°C
At high temperatures the single peak model has growth rates that decline more slowly than the double peak model. However, in either case the likely generation times at about 130°C of 1.7–2.7 h are quite short. In the case of the single peak model, the fitted curve for the maximum rate in Fig. 5 appears to descend too slowly compared with the observed data, although the wider 99% credible band of the double peak model continues to overlap with the data. We have suggested that the BKST arises from an evolutionary trade-off of activity and stability of biomolecules (Corkrey et al. Reference Corkrey, McMeekin, Bowman, Ratkowsky, Olley and Ross2014). If so, then the discrepancy between the fitted curve and the observed data suggest a different mechanism is operative at the highest temperatures.
Between T sup and up to high temperatures, say 100°C, some of the cell's resources are allocated to growth and the rest to non-growth activities, particularly maintenance. If we consider growth rates at one temperature in this range, we may assume that as the rate of growth approaches the maximum rate the proportion of resources needed for non-growth rises faster than the proportion needed for growth, so that eventually, the latter is exhausted. A fast-growing cell maximizes its production of ribosomes to compensate for replacement of denatured proteins (Maitra & Dill, Reference Maitra and Dill2015), and other diffusion-limited proteins such as elongation factor (Klumpp et al. Reference Klumpp, Scott, Pedersen and Hwa2013), which we can assume increases with temperature. Therefore, we can conclude from the descending curve of the BKST that there is an upper limit to the production rate of ribosomes and related proteins, which we suggest may be related to the MRS.
If a cell has lower maintenance requirements, for whatever reason, then the same number of ribosomes would suffice to obtain the same growth rate. We note that the portion of the BKST above about 80°C is dominated by Archaea rather than Bacteria, which, based on the above argument, indicates that Archaea have comparatively lower maintenance requirements. Archaea, and some of the more thermophilic Bacteria, make use of stable ether lipids within cell membranes (Daniel & Cowan, Reference Daniel and Cowan2000), although not all hyperthermophiles make use of such lipids (Koga, Reference Koga2012). This may be consistent with the suggestion that Archaea are competitive with Bacteria in low-energy environments (Valentine, Reference Valentine2007) and have low permeability membranes (van de Vossenberg et al. Reference van de Vossenberg, Driessen and Konings1998; Koga, Reference Koga2012), and thus may require less energy to maintain cytoplasmic homeostasis.
High-temperature environments are very demanding, potentially reducing the maximum growth rate. Adaptation may take the form of increased biomolecular stability, greater stability of RNA and DNA, and more robust cell membranes. The greater stability of thermophile proteins compared with mesophile proteins may be obtained by decreased numbers of uncharged polar residues, increased number of charged residues, increased residue volume (Fields, Reference Fields2001), greater residue hydrophobicity (Mukaiyama & Takano, Reference Mukaiyama and Takano2009), greater use of salt bridges (Kumar & Nussinov, Reference Kumar and Nussinov2001) and compatible solutes (Sterner & Liebl, Reference Sterner and Liebl2001), and an increase in enthalpic forces (Kumar & Nussinov, Reference Kumar and Nussinov2001). Thermostable proteins also unfold more slowly (Luke et al. Reference Luke, Higgins and Wittung-Stafshede2007) and are more highly expressed (Cherry, Reference Cherry2010). Cells also need to manage deamination, depurination and hydrolysis of DNA with increasing temperature (Jaenicke & Sterner, Reference Jaenicke, Sterner, Dworkin, Falkow, Rosenberg, Schleifer and Stackebrandt2006). The stability of DNA and RNA can be improved by increasing the GC content (Daniel et al. Reference Daniel, Dines and Petach1996), although this may be more important in ribosomes (Galtier & Lobry, Reference Galtier and Lobry1997).
The continuation of the BKST descending curve to fairly high temperatures, say 90°C, does not suggest an abrupt termination, such as may be expected from a loss of protein or DNA conformation, or from membrane instability. If the MRS consists of a single, rate-limiting, enzyme-catalysed reaction, then it will depend on chaperones that are responsible for de novo folding and refolding. Such enzymes need to be sufficiently active to function, which would be reduced by too much stability. However, they should only be as stable as is necessary for their environment (Wang et al. Reference Wang, Minasov and Shoichet2002; Bloom et al. Reference Bloom, Labthavikul, Otey and Arnold2006) and may denature at temperatures not far above T opt (Daniel et al. Reference Daniel, Dines and Petach1996; Daniel, Reference Daniel2003). However, evolution seems capable of optimizing protein stability at very high temperatures (Daniel, Reference Daniel1996). Even so, there is presumably a limit beyond which this is infeasible, as suggested by the tendency of the BKST maximum limit to overshoot the observed data at very high temperatures, suggesting another mechanism becomes operative at this point. It seems likely that the maximum limit at such temperatures depends on the loss of primary structure due to changes in covalent bonding (Ahern & Klibanov, Reference Ahern and Klibanov1985). In other words, the maximum temperature limit for life may result from mechanisms such as the stability of amino acids or ATP. In summary, we suggest that the maximum limit of the BKST is reduced at very high temperatures by a failure hierarchy, such as ribosome conformational stability followed by DNA stability, lipid membrane stability (Koga, Reference Koga2012), cell wall integrity (Hansen et al. Reference Hansen, Criddle and Battley2009) and eventually the stability of smaller molecules such as ATP, upon which many cellular mechanisms depend (Kim et al. Reference Kim, Hipp, Bracher, Hayer-Hartl and Hartl2013).
The stabilities of these molecules have been assessed in vitro. The in vitro half-life of ATP is 48 min at 95°C, 18 min at 105°C, 7 min at 115°C, 3 min at 125°C (Daniel et al. Reference Daniel, van Eckert, Holden, Truter, Crowan, Wilcock, Delong, Kelley, Baross and Cary2004) and several minutes at 127°C (Leibrock et al. Reference Leibrock, Bayer and Lüdemann1995). Even at the highest of these temperatures the half-life of ATP is still much longer than the turnover half-life (Daniel et al. Reference Daniel, van Eckert, Holden, Truter, Crowan, Wilcock, Delong, Kelley, Baross and Cary2004). At very high temperatures it may be beneficial for cells to make use of ADP or AMP that are more thermostable than ATP (Daniel & Cowan, Reference Daniel and Cowan2000). Wolfenden & Snider (Reference Wolfenden and Snider2001) reported that the half-life for hydrolysis events per bond for RNase A bonds at 25°C was 4 years, whereas at 100°C it was 7 h. Similarly, the half-lives per bond cleavage for polysaccharides decreased from 50 years to 12 h, for RNA from 20 days to 3 h and for DNA from 1 month to 2 h. The half-life of ribose is 73 min at 100°C (Larralde et al. Reference Larralde, Robertson and Miller1995), while the half-lives of amino acids at 250°C range from 30 min for serine to 80 min for valine (Bernhardt et al. Reference Bernhardt, Lüdemann, Jaenicke, König and Stetter1984). Strand separation in DNA increases with temperature, but may be stabilized by salts and polyamines (Daniel & Cowan, Reference Daniel and Cowan2000), and efficient DNA repair mechanisms (Grogan, Reference Grogan1998), while transfer and ribosomal RNA may be stabilized by post-transcription modification (Daniel & Cowan, Reference Daniel and Cowan2000). Finally, amino acids can remain stable to 150°C (Daniel et al. Reference Daniel, Dines and Petach1996; Bains et al. Reference Bains, Xiao and Yu2015).
While field observations have been made of microbial colonies surviving to at least 125°C (Holden & Daniel, Reference Holden, Daniel, Wilcock, Delong, Kelley, Baross and Cary2004), we do not know of organisms growing above 122°C. This may be because such environments are too unstable, that organisms from high-temperature environments have not yet been cultured or are unculturable, or that it is the maximum limit above which life on Earth cannot grow. However, the half-lives listed above are quite lengthy. They also imply that at high temperatures protective mechanisms become vital and that the cost of repair at sufficiently high temperatures may become unsustainable (Jaenicke & Sterner, Reference Jaenicke, Sterner, Dworkin, Falkow, Rosenberg, Schleifer and Stackebrandt2006). For comparison, we show in Fig. 5 the adjusted maximum growth rates based on the assumption that growth rate is reduced in proportion to the degree of in vitro hydrolysis of ATP or Ala-tRNA. The adjusted rates after hydrolysis of ATP or Ala-tRNA were meant for illustrative purposes rather than actual predictions, but showed that the rates would decline strongly if the cell were unable to compensate by replacing or repairing such molecular species. Less steep slopes might be expected if the data had been collected in vivo rather than in vitro. While we do not claim that such effects will produce the upper temperature limits illustrated, the similarity of the rate of decline to the observed data is intriguing.
Applications to astrobiology
In astrobiology, cardinal limits for temperature, such as T min and T max, arise in the context of defining habitability. They are used when defining the feasible range for temperature for familiar life. The actual meanings of T min and T max can vary, sometimes referring to theoretical, extrapolated or observation limits. For example, T min may be used to mean the intrinsic biological minimum temperature for growth and be a lower temperature than the observed minimum temperature for growth, known as MINt; similarly, the biological maximum temperature for growth, T max, may be a higher temperature than the observed maximum temperature for growth, MAXt (McMeekin et al. Reference McMeekin, Olley, Ratkowsky, Corkrey and Ross2013). This distinction matters where the biological and observed limits differ greatly, particularly for extremophiles that are likely to be difficult to culture. Further, estimates of the upper and lower limits of data are intrinsically more variable than their central measure. For example, we may expect that estimates of T min and T max will be less certain than T opt or T mes. Analogous limits are used for other physicochemical conditions such as salinity, pH and pressure, and similar considerations may be expected for those cases. However, descriptive statistics, such as these, have been shown to be very useful in astrobiology. We suggest that growth rate data can play a complementary role to these cardinal limits. The combination of the two approaches may be expected to provide further insights in astrobiological research.
There are very few environments on Earth nor any particular physicochemical extreme where life does not exist (Space Studies Board, 2007). The conditions in which life in the most general sense may exist are referred to as habitability. One application of the stressors, such as temperature, pH, and so on, is in obtaining a metric for habitability. An example of such a metric is the maximal volume of the parameter space enclosed by multidimensional stressor limits (Dartnell, Reference Dartnell2011; Harrison et al. Reference Harrison, Gheeraert, Tsigelnitskiy and Cockell2013, Reference Harrison, Dobinson, Freeman, McKenzie, Wyllie, Nixon and Cockell2015; Cockell et al. Reference Cockell2016). Determination of these stressors depends on the detection of growth and no-growth. These are likely to be subject to error if the generation time of an organism is very long or the organism more difficult to culture near the limits, such as for extremophiles. This will introduce variance additional to that already discussed for the cardinal limits. As a novel alternative, we can calculate the areas under the maximum rate curves and quantile curves, with larger areas indicating either faster growth rates, wider temperature limits or both.
Consideration of the limits represented by extremophiles may be interesting, but we suggest that it is also necessary to investigate how well life does in such extreme environments. This may take the form of phase diagrams used to assess habitable space (Jones & Lineweaver, Reference Jones and Lineweaver2012) that are enhanced by allowing for growth rates under multiple stressors. Habitability may then be better defined in terms of survival rather than reproduction (Cockell et al. Reference Cockell2016) thus obtaining a larger parametric volume, but nevertheless, we suggest that interest should be centred on where life can do well, viz. where exponential growth is possible. Once we obtain an estimate of the range of best possible growth rate we can assess long-term habitability. If we define the feasible range as those conditions, say temperature, in which familiar life exhibits growth, then the BKST indicates that the temperature range in which life grows more quickly is a narrower range than the feasible range. While conditions within an environment may be habitable, they may not remain so. If they drift too far then the maximum growth rates will decline and although the environment remains technically habitable, the range and complexity of life would be likely to be reduced. In a marginally habitable environment where only very slow growing life is possible then the chance of extinction would increase, much in the same manner as local extinction occurs in island biogeography models (Hanski & Gilpin, Reference Hanski and Gilpin1991). Thus, even if life survives the hypothetical Gaian emergence bottleneck (Chopra & Lineweaver, Reference Chopra and Lineweaver2016), it may still succumb in marginally habitable environments. In summary, the BKST indicates viability within the limits of habitability.
As discussed extensively in Cockell et al. (Reference Cockell2016), factors affecting the habitability of a planetary environment include its position relative to the habitable zone, which will vary in time as the planet's star luminosity changes. The zone is expected to depend on a number of factors such as the planet's water inventory, necessary to maintain a greenhouse effect, and tectonic activity to maintain a carbonate–silicate cycle. Within such abiotic determinants cardinal temperature limits are of use in determining the habitable zone extent, while growth rates can provide some measure of viability of the zone. In this sense, the two measures should be viewed as complementary rather than alternatives. It would be useful to further consider the interaction of cardinal limits and growth rates with the duration and extent that a planetary environment resides within the habitable zone. While a habitable zone, as determined by the use of cardinal limits, may include a planet for some portion of its history, the BKST may inform consideration of growth rates that are possible in that interval, and hence the complexity of evolutionary trajectories that may arise. In other words, the complementary use of cardinal limits and the BKST may allow estimation of the biological richness possible during a planet's history.
While the current study only concerns the effect of temperature on growth rates, Harrison et al. (Reference Harrison, Dobinson, Freeman, McKenzie, Wyllie, Nixon and Cockell2015) discuss the physiology of adaptation to multiple extremes by examining the limits to growth arising from extremes in temperature, NaCl and pH, and conclude that the parametric volume occupied by aerobes exceeds that of non-aerobes. When comparing aerobes and two anaerobic groups they found that one of the anaerobic groups displayed narrower temperature and pH ranges, while aerobes had a broader salinity range. They attribute this to the greater energy available for aerobes compared with anaerobes. While greater energy availability might increase the tolerable range for salinity, it is not clear why the tolerable temperature range should increase. We find that aerobes (Fig. 6 and Table 3) have a smaller area under the 95% quantile curve and smaller peak rate than anaerobes. We also find that heterotrophs had a larger area and higher peak rate than autotrophs. These contrasts are also reflected in the combinations of the aerobic and trophic statuses. While the metabolisms of aerobes may be able to access a larger amount of energy than anaerobes, they display a more limited area and peak rate, suggesting that the differences between aerobes and anaerobes reflects the degree to which they allocate energy to growth. A similar possibility may arise for heterotrophs and autotrophs.
Harrison et al. (Reference Harrison, Dobinson, Freeman, McKenzie, Wyllie, Nixon and Cockell2015) found that the tolerable range for salinity and pH reduces above 40°C. They find that the mean T min, T opt and T max are all greater for anaerobes than aerobes, and that the temperature range tolerated differs between anaerobes and aerobes. As seen from the BKST this probably arises from the preponderance of anaerobes with higher temperature optima (Fig. 2) and that these tend to be Archaea. Such phenomena may be expected if the resources of organisms are utilized to cope primarily with one type of environmental stress, such as temperature, leaving less available to deal with other types of environmental stressors, perhaps salinity (Bowers et al. Reference Bowers, Mesbah and Wiegel2009). As we noted above, Archaea have comparatively lower maintenance requirements. This may be consistent with the suggestion that Archaea are competitive with Bacteria in low-energy environments (Valentine, Reference Valentine2007).
We suggest that the parametric volume approach developed in Harrison et al. (Reference Harrison, Gheeraert, Tsigelnitskiy and Cockell2013) and Harrison et al. (Reference Harrison, Dobinson, Freeman, McKenzie, Wyllie, Nixon and Cockell2015) is of considerable interest. That method made use of pH and NaCl concentration. While the use of cardinal limits alone ignores the considerable information within the BKST a combination of the parametric volume and BKST approaches may be fruitful. In its current form, the BKST includes variation of growth rates that arise from experiments that were conducted under different conditions, such as pH and water activity, that may be suboptimal for particular organisms. We explicitly opted not to introduce corrections that may have introduced artefacts, but to accept the data as they were, since the range of possible growth rates was more pertinent to describing the form of the BKST than were the estimation of mean tendencies. While it may be suggested that the data should be corrected to allow for these variations, we suggest that a more productive approach is to generalize the BKST to include such factors. This could take the form of a generalized BKS that includes other conditions, such as pH, water activity, ionic strength, and pressure. We can then calculate the volume of quantiles of such a multi-dimensional distribution. To a limited extent we do this here by calculating areas under quantile curves. This proposed approach may also shed light on the nature of the MTG.
As noted earlier, the BKST provides the range of growth rates and maximum rates possible for familiar life. We do not expect to find any form of life growing above the BKST, but we can discuss what might be expected should this not prove to be correct. We can do this by examining the degree of certainty exhibited by the estimate of the maximum limit at various temperatures. We propose that organisms with growth rates greatly in excess of the ascending curve are unlikely to exist since the estimated maximum growth curve for the ascending curve has a very narrow credible band indicating strong support by the available data. The descending curve has a wider credible band and so is less well defined. This means that if new fast-growing strains are discovered they are more likely to be located above the descending curve than the ascending curve. The wider credible band of the descending curve also includes the peak at T sup, so that further observations also might be expected here.
More specifically, we do not expect new strains to be located in the heavily shaded region shown in Fig. 7. This is the region located above the 99% credible band for the maximum limit. The choice of 99% is arbitrary and could be adjusted higher if required. For example, a novel strain observed with a growth rate of 0.06 at 25°C, shown as ‘A’ in the heavily shaded area in the figure, would be an example of a case that would disprove our theory. A strain observed with a growth rate of 0.1 at 95°C, shown as ‘B’ in the figure, would be another example. These growth rates are not implausible in themselves, but we suggest they are unlikely to occur at these particular temperatures. If they were observed at T sup, then neither would be improbable, but as the temperature shifts to increasingly lower or higher values, so that they fall in the shaded area then they become untenable. Observation of either rate in the shaded area would represent contrary evidence for the existence of the BKST. On the other hand, Baross & Deming (Reference Baross and Deming1983) claimed to have detected life growing at 250°C. As already discussed, this was considered unlikely since biomolecules hydrolyse in water at this temperature (White, Reference White1984), and their observations were later thought to result from particulate matter being mistaken for cells. However, they reported a growth rate of 0.017 (per minute), shown in Fig. 7 as a dashed line. According to the BKST, such a rate is only achievable in the range 21–106°C. Such a claim would definitely be considered as implausible based on the BKST. These are examples of how the BKST may be used as a diagnostic for plausible growth rates, or alternatively, to identify unusual life.

Fig. 7. Growth regions. The observed data are shown as circles. Regions where growth rates exceed the posterior mean for the maximum limit but within the 99% credible band are cross-hatched, while rates that exceed the 99% credible band are shaded using diagonal green lines. The rate obtained by Baross & Deming (Reference Baross and Deming1983) is shown as a dashed line. Strains with T opt ≤ 50 are shown as blue closed circles and those with T opt > 50 are shown as red open circles. Two hypothetical cases discussed in the text are indicated by A and B.
While we do not expect to find any new organism on Earth capable of growing substantially above the BKST, this may not be the case for life elsewhere. If we accept the notion that the BKST arises from a MRS that has been strongly conserved since the LUCA, then none of the descendants of the LUCA will display growth rates greatly in excess of the BKST. However, organisms that do not descend from the LUCA or those with a sufficiently modified or omitted MRS, might do so. The BKST therefore potentially provides a diagnostic for unusual life.
One possibility for unusual life may be synthetic life (Blain & Szostak, Reference Blain and Szostak2014) that is constructed, perhaps deliberately, so as to lack the MRS. If so, there appears no reason why synthetic life that lacks the MRS could not be constructed to achieve growth rates in excess of the BKST maximum limit. This experiment would also provide a direct test for the existence of the MRS.
Life may have appeared more than once on Earth (Cleland & Copley, Reference Cleland and Copley2005; Davies & Lineweaver, Reference Davies and Lineweaver2005). This is considered possible since life probably appeared very quickly on this planet (Des Marais & Walter, Reference Des Marais and Walter1999), almost as soon as the cometary impact of the Archean ceased. If the propensity for biogenesis is as high as this suggests, then life probably appeared during the Archean, but was extirpated by the heavy bombardment. It has also been suggested that these earlier forms of life may have survived in subsurface refugia and still be extant today (Davies & Lineweaver, Reference Davies and Lineweaver2005). Discovery of second biogenesis life would increase the estimated probability that non-terrestrial life exists (Des Marais & Walter, Reference Des Marais and Walter1999; Davies, Reference Davies2012). However, detection of second biogenesis life may be difficult since it would not share the same biological characteristics as ordinary life (Davies et al. Reference Davies, Benner, Cleland, Lineweaver, McKay and Wolfe-Simon2009). However, as with synthetic life, second biogenesis life would not necessarily possess the same MRS as ordinary life, and therefore, would not be expected to conform to the BKST. Naturally evolved organisms from Earth that grow above the BKST maximum limit would therefore be candidates for second biogenesis life.
Early life on Earth may have been thermophilic (Stetter, Reference Stetter2006; Weiss et al. Reference Weiss, Sousa, Mrnjavac, Neukirchen, Roettger, Nelson-Sathi and Martin2016) and, although not all agree (Boussau et al. Reference Boussau, Blanquart, Necsulea, Lartillot and Gouy2008), for illustrative purposes we assume here that early life was thermophilic. If this was the case then an early visitor to Earth might have observed a BKST as shown in Fig. 7 by red open circles; the peak represented by the closed blue circles, and consisting of mesophiles, would not yet have appeared. This implies that the BKST may contain relics of evolution and the two peaks of the BKST could potentially correspond to alternative evolutionary histories. Non-terrestrial life will have evolved separately (excluding panspermia), not be descended from the LUCA, not possess the same MRS, and also not be expected to conform to the BKST. The BKST for non-terrestrial life may similarly include evolutionary information specific to that form of life, and consequently differ in shape and location. It may, for example, be translated along the temperature scale, rise to a different height, or possess fewer or more than two peaks. An extreme example might be life based on a solvent other than water (Bains, Reference Bains2004) for which the BKST would be displaced to much colder or hotter temperatures, but we would still expect it to be bounded below by the bulk properties of that solvent, such as vitrification at low temperatures and by the degradation of biomolecules at high temperatures.
This assumes that the BKST for non-terrestrial life depends on evolutionary contingencies (Des Marais & Walter, Reference Des Marais and Walter1999; Des Marais et al. Reference Des Marais, Harwit, Jucks, Kasting, Lin, Lunine, Schneider, Seager, Traub and Woolf2002), such as protein activity/stability trade-offs (Fields, Reference Fields2001) and is not a universal principle (Pace, Reference Pace2001). If the nature of the MRS is contingent on evolutionary history then we would not expect the same BKST as we observe on Earth for life elsewhere. But if the limit results from some fundamental process, then the BKST may be universal. One possibility for a universal mechanism is hydrogen bonding between water molecules that is known to influence protein folding and hence protein stability (Wiggins, Reference Wiggins2008).
A biosignature is a characteristic produced by the presence of life that may allow for its detection (Des Marais & Walter, Reference Des Marais and Walter1999), perhaps at a remote distance, typically by spectroscopic features (Des Marais et al. Reference Des Marais, Harwit, Jucks, Kasting, Lin, Lunine, Schneider, Seager, Traub and Woolf2002). If a biosignature signal is available that can be plausibly linked to a temperature-dependent process then we suggest examining if it possesses characteristics analogous to the BKST: at least one distinct peak, exponential ascent and descent. A signal with a temperature dependence that is simply exponential with temperature might well arise from an abiogenic mechanism. But a signal that displays both an ascending and a descending curve would be more likely biogenic.
Summary
The BKST describes the distribution of growth rates at which individuals reproduce, populations increase, or cells divide, for life on Earth over the range of temperatures at which life is known to exist. Additional factors may constrain growth at very low or very high temperatures, but we can expect the BKST to indicate the maximum limits for growth rates at all temperatures.
This work has generated a number of novel results. We:
-
1. develop a method to extrapolate trends of quantile curves to obtain estimates of the maximum rate of growth;
-
2. note that the BKST curve overshoots the observed data below 0°C and at very high temperatures;
-
3. suggest that growth above the vitrification temperature and below 0°C is limited by bulk properties of water, such as water activity, and suggest that growth rates at very high temperatures are limited by degradation of biomolecules;
-
4. suggest that growth rates may be complementary to the use of cardinal temperatures in astrobiology;
-
5. argue that since the MRS may determine the BKST, and that the MRS is strongly conserved since the LUCA, that life of an alternative origin would not possess the same MRS and therefore display a different BKST;
-
6. predict no growth will be observed substantially above the maximum rate limit, with some interesting exceptions, including for synthetic life, and life that does not share the same LUCA, such as non-terrestrial life;
-
7. suggest that the BKST may be of use as a diagnostic for unusual life, including second biogenesis life and non-terrestrial life;
-
8. suggest that the BKST may contain relics of evolution or evolutionary adaptations;
-
9. suggest that biogenic and abiogenic biosignatures may be distinguishable using the properties of the BKST.
Acknowledgements
We are grateful to Dr Shane Powell and Professor Philip Boyd for reading the manuscript and for the thoughtful comments by the reviewer.










