


Introduction
Throughout development, our neural and physiological systems are exceptionally responsive to the environment. This responsivity allows us to both learn about and use resources from our surroundings. Dynamic detection and adjustment to ongoing stimulation ultimately influence the functioning of our neural and physiological systems and, consequently, our mental and physical well-being. The caregiving environment plays a particularly important role in the establishing the foundations of these neural and physiological regulatory systems and scaffolding their development over the life span (Shonkoff, Garner, CPACF, & CECAD, Reference Shonkoff and Garner2012). Humans are both physically and emotionally dependent on caregivers, particularly for the first several years of life, which allows infants and young children to safely explore an unpredictable world. In turn, this process guides developing neural and physiological networks to adapt to current conditions. Early caregiving experiences continue to have an impact on development and well-being, in part through the child's neural and physiological regulation (Blair & Raver, Reference Blair and Raver2012). Because children's stress-regulatory systems are still developing, caregivers play an important role in externally regulating their child's stress response and helping them respond adaptively to stress and challenge. Importantly, caregivers’ own physiology has an impact on their ability to provide sensitive and responsive care to their children (Finegood, Blair, Granger, Hibel, & Mills-Koonce, Reference Finegood, Blair, Granger, Hibel and Mills-Koonce2016; Mills-Koonce et al., Reference Mills-Koonce, Propper, Gariepy, Barnett, Moore, Calkins and Cox2009), but the role that caregiver physiology, and specifically caregiver–child physiological synchrony, plays in the development of children's self-regulation is still uncertain.
Research on caregiver–child physiological synchrony has been growing rapidly in recent years, as it is suspected to help explain how caregivers and children interact with and respond to each other, thus affecting children's long-term development. Physiological synchrony has been measured and discussed using various terms (e.g., concordance, attunement, coregulation, coordination) with little consistency or clarity in the construct's exact definition (e.g., Saxbe et al., Reference Saxbe, Margolin, Spies Shapiro, Ramos, Rodriguez and Iturralde2014; Skoranski, Lunkenheimer, & Lucas-Thompson, Reference Skoranski, Lunkenheimer and Lucas-Thompson2017; Suveg et al., Reference Suveg, Braunstein West, Davis, Caughy, Smith and Oshri2019). Generally, the concept is used to refer to the association between some measure of physiological activity in the caregiver and that same measure in the child in response to a task or in a particular context. This includes tasks eliciting a physiological stress response (changes in arousal in response to real or imagined threat; Charmandari, Tsigos, & Chrousos, Reference Charmandari, Tsigos and Chrousos2005) but also reactivity to neutral or positive tasks not intended to elicit a stress response. “Synchrony,” “concordance,” and “attunement” as used currently tend to assume a positive relation between caregiver and child physiological activity (i.e., physiological activity moving up or down for both individuals together) while “negative attunement,” “discordance,” and “asynchrony” are used to refer to a negative association (i.e., physiological activity moving up for one individual and down for the other). While few studies acknowledge it, positive versus negative synchrony may be critical for understanding its function, correlates, and consequences.
The benefits of physiological synchrony for children's development have yet to be explored in detail. Physiological synchrony in romantic couples and therapist–client dyads (see Palumbo et al., Reference Palumbo, Marraccini, Weyandt, Wilder-Smith, McGee, Liu and Goodwin2017 for a review), at least in the autonomic nervous system (ANS), is associated with marital satisfaction, relationship quality, and empathy, though results tend to be mixed. It is possible that parent–child physiological synchrony is beneficial by providing a scaffold from which children can develop independent regulatory skills (Lougheed & Hollenstein, Reference Lougheed and Hollenstein2018; Wass et al., Reference Wass, Smith, Clackson, Gibb, Eitzenberger and Mirza2019). Alternatively, synchronous physiological activity could exaggerate the experience of stress between two individuals (Waters, West, Karnilowicz, & Mendes, Reference Waters, West, Karnilowicz and Mendes2017; Waters, West, & Mendes, Reference Waters, West and Mendes2014). If a child exhibits a physiological stress response, it might not be desirable for a caregiver to have a synchronous stress response, and vice versa.
Physiological synchrony may also function differently and have distinct developmental implications based on a variety of factors. The body's physiological systems work in concert with one another; however, each system in the network has its own niche, with varying patterns of activity, function, and timescale of operation (Joëls & Baram, Reference Joëls and Baram2009). The form and function of each system must be considered when quantifying and interpreting caregiver–child synchrony, because synchrony may have divergent functions across systems. It is also likely that synchrony will have unique implications across developmental periods particularly for the caregiver–child relationship, the function of which changes throughout development. Context may also play a role such that acute stressors or chronically stressful environments may change our interpretation of the function of synchrony and how it manifests in different caregiver–child dyads.
Thus, for research on physiological synchrony to move forward, a clearer definition must be adopted consistently across studies and research groups. Once defined, it must be thoroughly evaluated for its relevance and implications for children's development. Further, variations in patterns of synchrony and implications for child developmental functioning should be considered across physiological systems, developmental periods, and contexts. Others have begun to direct attention to this need (for example, see Bernard, Kashy, Levendosky, Bogat, & Lonstein, Reference Bernard, Kashy, Levendosky, Bogat and Lonstein2017; Davis, West, Bilms, Morelen, & Suveg, Reference Davis, West, Bilms, Morelen and Suveg2018; Palumbo et al., Reference Palumbo, Marraccini, Weyandt, Wilder-Smith, McGee, Liu and Goodwin2017), describing physiological synchrony and highlighting important concerns regarding measurement and statistical approaches. However, a detailed account of developmental processes and contextual influences is essential to fully understand the construct of caregiver–child physiological synchrony.
To begin to address these questions, the author proposes a working definition of physiological synchrony as the dynamic, within-dyad coordination of physiological activity over time between two individuals that is directly tied to an interpersonal process. Further, the author has chosen “synchrony” instead of other terms such as “attunement” or “concordance” because synchrony is the most prevalent term in the literature to date. “Negative synchrony” will be used to represent an inverse association between parent and child physiology (one individual increases while the other decreases) and “asynchrony” will represent the absence of positive or negative synchrony (no association between parent and child physiology). The author has chosen to define synchrony in terms of the direction of the statistical association between a caregiver and child's physiological activity, rather than in terms of its function or its consequences for child development, because there is little consensus about the function of synchrony and its developmental consequences thus far. This review intends to provide some clarity about these open questions.
Moving forward, it will be important for the field to develop common terminology with which to discuss the phenomenon of synchrony with clear definitions for these and other central concepts. For example, it is clear that synchrony in one physiological system can inform the study of synchrony in another system, and measuring synchrony in multiple systems simultaneously would be even more useful. However, to date, terminology and theoretical perspectives seem siloed within physiological systems (e.g., adrenocortical vs. autonomic) and synchrony appears to be defined and operationalized in many different ways. Both caregiver-to-child and child-to-caregiver effects have been examined, but there is not much of a guiding framework for which directionality, if not both, should be expected in particular circumstances. This review will survey the extant research, which has increased exponentially in frequency over the past few years, to interpret findings from a developmental perspective (where possible) and with respect to the definition of synchrony given above. This review will also identify critical gaps in our knowledge and methods and delineate paths for future research that will be necessary to identify relevant developmental and contextual factors.
Objectives
In light of the emerging questions described above, the following review has three objectives. First, it will characterize the state of the literature regarding caregiver–child physiological synchrony and associations with other indices of child development. Where available, relations with desirable developmental functioning (e.g., behavior regulation, reduced psychopathology) will be examined to inform the function and developmental correlates of physiological synchrony. However, so far too few synchrony studies have examined measures of child functioning; more prevalent are associations between physiological synchrony and behavioral measures of caregiving or the parent–child relationship, so these will also be considered. Second, this review will examine whether there are developmental differences in the correlates of physiological synchrony. Finally, where relevant studies exist, this review will explore whether context (e.g., task demands, parent behavior, relationship characteristics), risk (e.g., maltreatment history), and/or stress (e.g., activation of the stress response) influence the association between physiological synchrony and indices of positive development.
While previous reviews have begun to examine similar questions (see Davis et al., Reference Davis, West, Bilms, Morelen and Suveg2018), the authors furthers this discussion with an in-depth examination of patterns across developmental periods and an inclusion of central nervous system (CNS) data that will likely inform our understanding of synchrony in lower level systems. In addition, by physiological synchrony in response to a task and not diurnal/baseline synchrony, the author aims to increase clarity and consistency in the measurement of physiological synchrony to facilitate the integration of study findings. While important, diurnal synchrony likely operates through different mechanisms and should be interpreted separately. Further, by focusing on task-based activity, this review can examine task characteristics (e.g., positive- vs. negative-valence, parent and child behavior) that may affect the display of physiological synchrony or moderate the association between caregiver–child physiological synchrony and other variables. While the role of genetics and heritability of physiological regulation in caregiver–child relationships is an important topic, this is beyond the scope of the present review. This review will focus on the specific role of parasympathetic, sympathetic (and nonspecific autonomic), hypothalamic–pituitary–adrenocortical (HPA), and CNS synchrony. It is acknowledged that there are highly complex within-person interactions between these systems, but few studies measure physiological synchrony across systems. Therefore, findings will be summarized separately for each physiological system and discussed in the context of each system's biological niche.
Methodology
The protocol for this systematic review has been registered with PROSPERO, which is available at http://www.crd.york.ac.uk/PROSPERO/display_record.php?ID=CRD42019123770, registration number CRD42019123770.
Search strategy and eligibility criteria
PubMed and PsycINFO databases were searched on December 6, 2018 for relevant empirical articles using the following search strategy: ((synchrony OR coordination OR concordance OR attunement OR coregulation) AND (caregiver OR parent) AND (child* OR adolesc* OR infan*) AND (biolog* OR physiolog*)). A secondary search was conducted in February 2020 to find relevant articles published between the original search and the date of submission of this article. Details of the article selection process are shown in Figure 1 (following Preferred Reporting Items for Systematic Reviews and Meta-Analyses [PRISMA] guidelines; Moher et al., Reference Moher, Shamseer, Clarke, Ghersi, Liberati, Petticrew, Shekelle and Stewart2015). Duplicates between the two databases were removed, and studies were triaged based on titles/abstracts if they were not an English peer-reviewed empirical article or were clearly off-topic. Then, full text articles were examined and all papers that did not (a) include measures of activity in the parasympathetic nervous system, sympathetic nervous system, HPA axis, or CNS activity in both caregiver and child and (b) directly test the correlation, covariation, or some other direct association between caregiver and child physiology in response to a task were removed. The reference lists of these studies were also examined for relevant manuscripts, resulting in a total of 69 studies. Inclusion decisions were confirmed by a second independent reviewer. Reviewers showed 97.4% agreement, and all discrepancies were resolved through reviewer discussion. Due to the variability in measurement strategies and constructs examined in association with synchrony, eligible papers are qualitatively summarized below.
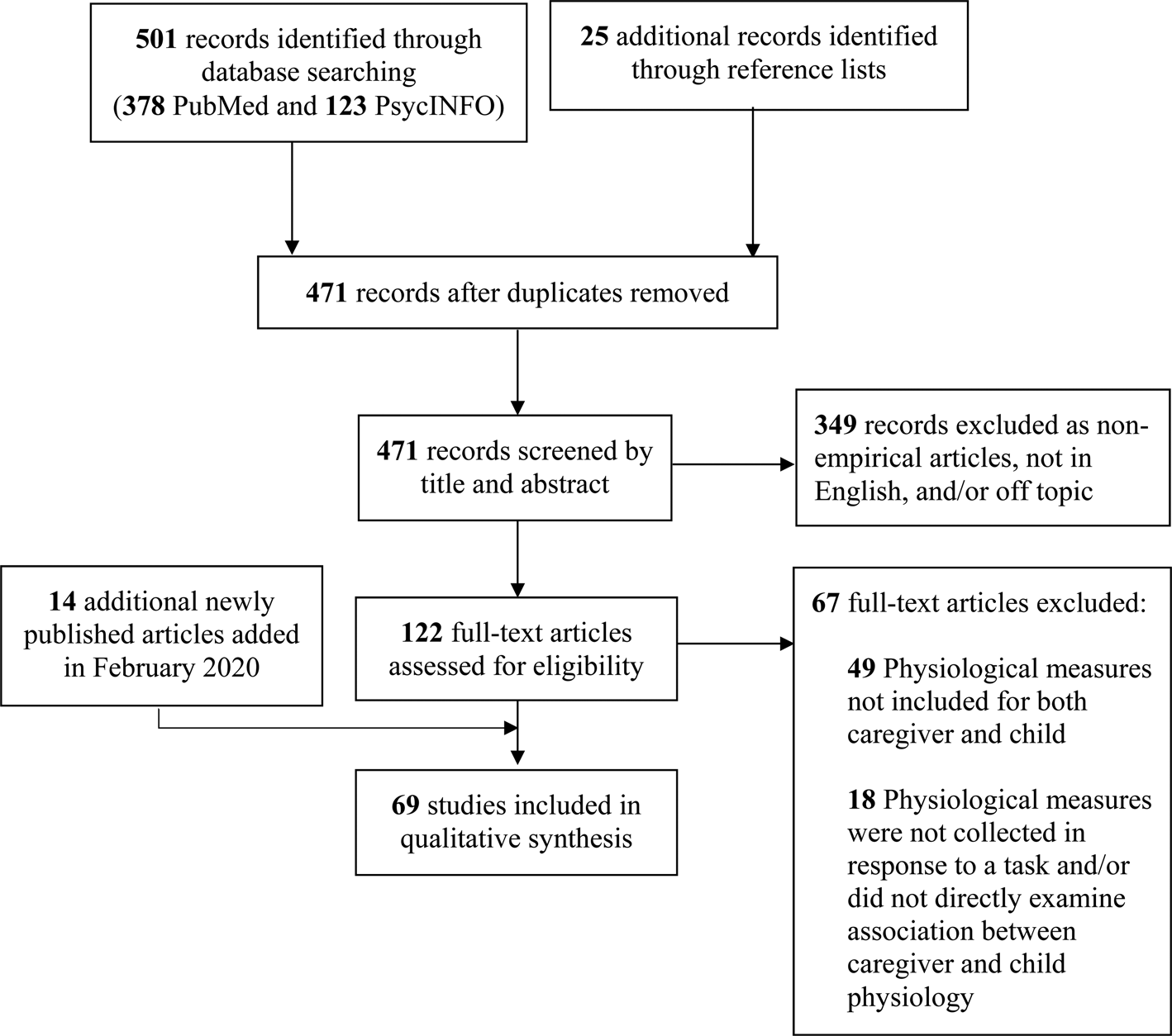
Figure 1. Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) systematic review flow diagram (Moher, Liberati, Tetzlaff, & Altman, Reference Moher, Liberati, Tetzlaff and Altman2009).
This review is organized by physiological system. Within each physiological system, pertinent methodological challenges and statistical modeling approaches were considered. Patterns related to (a) the developmental period of child participants, (b) associations with family/caregiver characteristics (e.g., maltreatment history, caregiving behavior) and/or child characteristics (e.g., child behavior, psychopathological symptoms), and (c) differences across contexts and risk status were also discussed. Selective publication bias and bias within studies was not assessed quantitatively due to the substantial heterogeneity of the selected studies.
Results
Systematic review study characteristics
Of the 69 studies included in this review, only two (one autonomic and one adrenocortical) incorporated multiple measurements of physiological synchrony over time. Sample sizes ranged from 6 to 1,141, with the smallest samples tending to be found in studies involving the brain, though the four smallest samples (N = 6, 10, 11, and 14 dyads) measured autonomic synchrony. Thirty-six studies assessed autonomic synchrony (parasympathetic, sympathetic, or both), while 27 included adrenocortical measures (all salivary cortisol activity), and seven measured CNS activity (typically after an interaction task). Only one study assessed both autonomic and adrenocortical synchrony (Laurent, Ablow, & Measelle, Reference Laurent, Ablow and Measelle2012). Forty-four studies assessed physiological synchrony in relation to some characteristic of the caregiver or the caregiver's behavior during the task and 22 measured the association between synchrony and child functioning, including 13 studies that contained both. The remaining 16 studies did not directly assess either caregiver or child characteristics. See Table 1a–1c for a summary of all 69 studies, categorized by physiological system and roughly in order of child participant age.
Table 1a. Summary of articles assessing caregiver–child autonomic synchrony
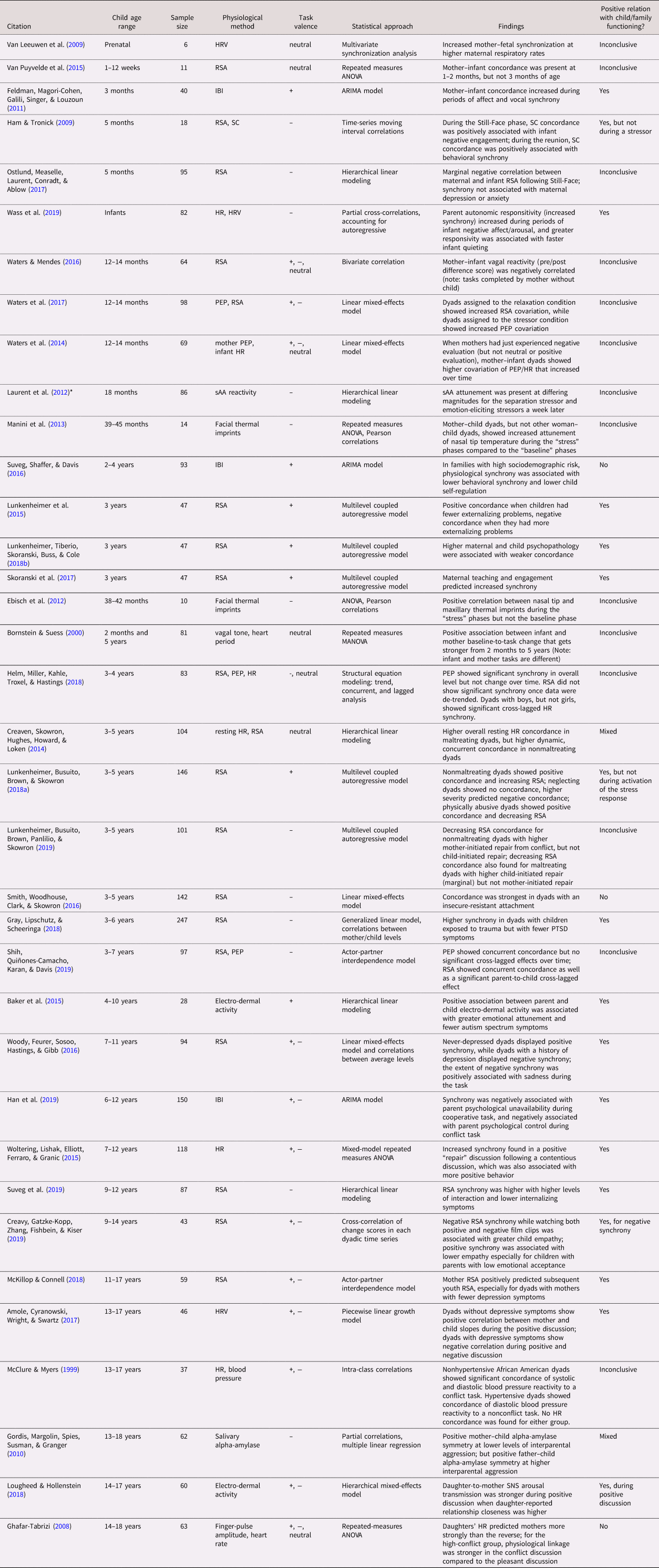
Table 1b. Summary of articles assessing caregiver–child adrenocortical synchrony
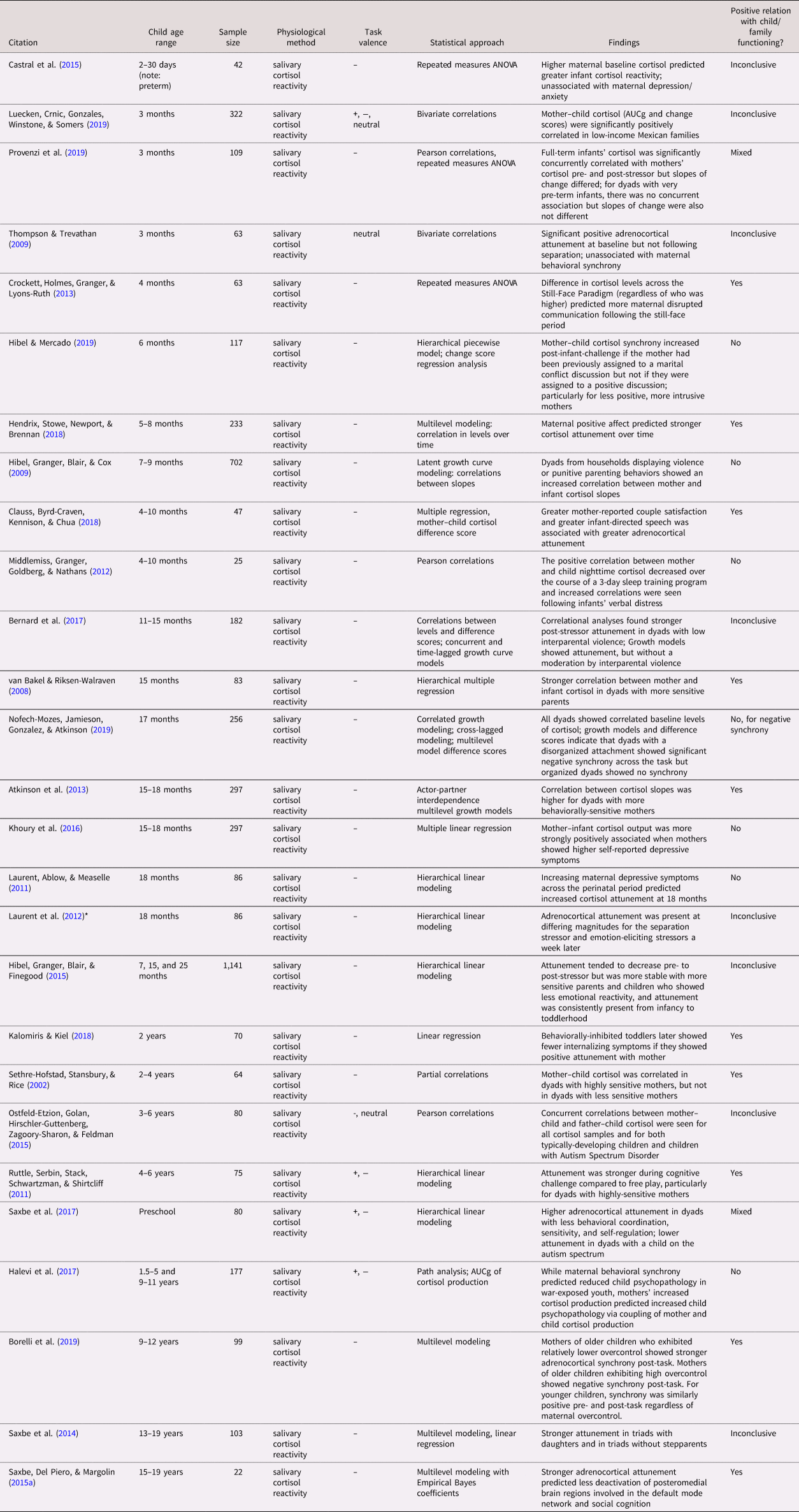
Table 1c. Summary of articles assessing caregiver–child central nervous system synchrony.
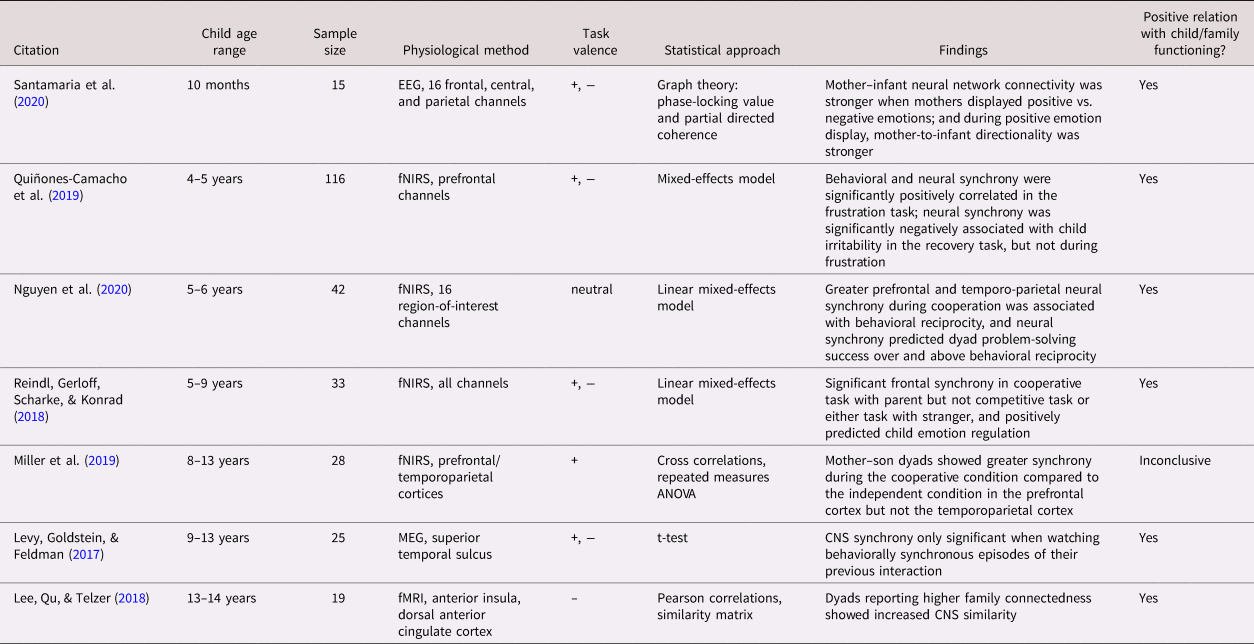
Note. Studies are arranged roughly by participant age within physiological system (a: ANS, b: HPA, c: CNS). Parasympathetic, sympathetic, and nonspecific autonomic measures were combined into the same table because of frequent overlap in studies. Study sample sizes provided refer to the total number of dyads, not individuals, in each study. *Laurent et al., Reference Laurent, Ablow and Measelle2012 is included in both the autonomic and adrenocortical tables (1a, 1b). ANOVA: analysis of variance; ANS: autonomic nervous system; ARIMA: auto-regressive integrated moving average; AUCg: area under the curve with respect to ground; CNS: central nervous system; EEG: electroencephalography; fMRI: functional magnetic resonance imaging; fNIRS: functional near-infrared spectroscopy; HPA: hypothalamic–pituitary–adrenocortical; HR: heart rate; HRV: heart rate variability; IBI: inter-beat interval; MANOVA: multivariate analysis of variance; MEG: magnetoencephalography; PEP: pre-ejection period; PTSD: post-traumatic stress disorder; RSA: respiratory sinus arrhythmia; sAA: salivary alpha-amylase; SC: skin conductance.
Parasympathetic nervous system
Biological processes
A complex network of neural, physiological, and endocrine factors is involved in the coordination of the stress response. Each plays a role in managing a particular aspect of the stress response across contexts and at different timescales. This coordination allows the brain and body to fine-tune responses to different types of stressors in a way that, when working effectively, maximally helps us deal with and recover from stress (Joëls & Baram, Reference Joëls and Baram2009).
On the shortest timescale, the parasympathetic nervous system (PNS) modulates its activity in accordance with changes in the environment. Particularly when in a calm, socially engaged state, a number of organs are under parasympathetic control via the vagus nerve (e.g., heart, pupils, lungs, and stomach) and primarily promote rest-and-repair functions (Porges, Reference Porges2007). Commonly measured by vagal tone (heart rate variability [HRV]; respiratory sinus arrhythmia [RSA]), increased PNS activity reduces heart rate and blood pressure and promotes digestion and pupil constriction. This is achieved via tonic inhibition of the vagus nerve on the sympathetic nervous system (SNS), which innervates the same organs and generates the fight-or-flight response (Porges & Furman, Reference Porges and Furman2012). Then, within seconds of encountering a stressor, the PNS “lifts the vagal brake” on the fight-or-flight response (i.e., vagal withdrawal), which may then result in increased activity in the SNS. After the challenge has been overcome or the stressor has been removed, vagal activity is again increased in order to aid in the termination of the stress response (Ulrich-Lai & Herman, Reference Ulrich-Lai and Herman2009). Thus, the PNS is most active in nonstressful social contexts, and a lack of PNS activity in this context would either indicate a poorly timed, maladaptive stress response or inadequate social engagement with the environment (Hastings et al., Reference Hastings, Nuselovici, Utendale, Coutya, McShane and Sullivan2008; Porges & Furman, Reference Porges and Furman2012). PNS activity, typically a moderate decrease in PNS activity followed by a prompt return to baseline, has been associated with emotional responding and better emotion regulation (Applehans & Luecken, Reference Applehans and Luecken2006; Musser et al., Reference Musser, Backs, Schmitt, Ablow, Measelle and Nigg2012). Conversely, too much activity of the PNS during stress or socio–emotional challenge may represent the body's inability to mount a sufficient response to the stressor, which can also be maladaptive because appropriate biological resources have not been recruited to deal with the situation.
Measurement of caregiver–child parasympathetic synchrony
Several studies have shown that relationship factors and interaction quality impact children's parasympathetic regulation (Hinnant, Erath, & El-Sheikh, Reference Hinnant, Erath and El-Sheikh2015; Kennedy, Rubin, Hastings, & Maisel, Reference Kennedy, Rubin, Hastings and Maisel2004), which can have downstream impacts on children's development (Hinnant et al., Reference Hinnant, Erath and El-Sheikh2015; Sturge-Apple, Davies, Martin, Cicchetti, & Hentges, Reference Sturge-Apple, Davies, Martin, Cicchetti and Hentges2012). Similarly, caregivers’ parasympathetic functioning predicts rates of positive veersus negative parenting behaviors (Mills-Koonce et al., Reference Mills-Koonce, Propper, Gariepy, Barnett, Moore, Calkins and Cox2009). Through the PNS's role in social engagement processes, dyadic synchrony between caregivers’ and children's PNS activity during an interaction may indicate, over and above individual PNS activity, a dyad's ability to positively engage with one another and respond flexibly to environmental changes. The PNS is uniquely suited to identify dynamic, within-dyad patterns of synchrony (rather than between-dyad correlations), both in terms of caregiver-to-child and child-to-caregiver effects (e.g., Lunkenheimer et al., Reference Lunkenheimer, Busuito, Brown and Skowron2018a; Wass et al., Reference Wass, Smith, Clackson, Gibb, Eitzenberger and Mirza2019), due to its short timescale of activity and noninvasive measurement.
This does not imply, however, that there are not significant challenges that need to be considered when measuring parasympathetic synchrony. For example, the appropriate lag time at which synchrony occurs between two interacting partners has not yet been determined. For a physiological system that changes on the order of milliseconds and is commonly measured in 30-second epochs (or, more recently, second-by-second moving intervals; Gates, Gatzke-Kopp, Sandsten, & Blandon, Reference Gates, Gatzke-Kopp, Sandsten and Blandon2015), we must identify the meaningful interval of measurement for dyadic synchrony. The working definition of synchrony provided above requires that synchrony be directly tied to an interpersonal process – the timescale of which is not yet determined. Some have attempted to investigate this issue by examining multiple different lag times in the same study (e.g., Lunkenheimer et al., Reference Lunkenheimer, Busuito, Brown and Skowron2018a; Nofech-Mozes et al., Reference Nofech-Mozes, Jamieson, Gonzalez and Atkinson2019). Researchers should statistically compare multiple epoch lengths and lag times that make sense for the system being measured (e.g., autonomic activity on the order of seconds, HPA on the order of minutes) and choose the form that best fits the data. Over time, we can begin to build a body of work that will hopefully highlight consistencies across studies. These consistencies can then be used to inform best practices for operationalizing synchrony moving forward to ensure that we do not overinterpret biological noise (e.g., respiratory patterns, movement, vocal activity) and measurement error as a meaningful interpersonal process.
In addition, appropriate statistical methods must be used to ensure that we are accurately attributing associations to within- rather than between-dyad change over time. Multilevel modeling techniques are critical for this reason, allowing researchers to directly test within-dyad and between-dyad effects as well as time-variant and time-invariant predictors (Creaven et al., Reference Creaven, Skowron, Hughes, Howard and Loken2014; Davis et al., Reference Davis, West, Bilms, Morelen and Suveg2018). To best approximate physiological synchrony as presently defined, multilevel modeling should “pull out” overall individual- or dyad-level patterns (i.e., de-trending) in order to isolate within-dyad synchrony over time (Curran, Howard, Bainter, Lane, & McGinley, Reference Curran, Howard, Bainter, Lane and McGinley2014). This can be achieved, for example, using multilevel autoregressive models such as autoregressive integrated moving average (ARIMA) models that include random intercepts or control for each individual's mean level of physiological activity (Lunkenheimer et al., Reference Lunkenheimer, Tiberio, Buss, Lucas-Thompson, Boker and Timpe2015). Each specific multilevel modelling method has strengths and different strategies should be used for different questions (for more detail, see Davis et al., Reference Davis, West, Bilms, Morelen and Suveg2018; Helm et al., Reference Helm, Miller, Kahle, Troxel and Hastings2018).
Parasympathetic synchrony across development
Infancy (8 studies)
There are several studies examining caregiver–child parasympathetic synchrony in infancy and even prenatally; however, few examine its associations with caregiving behavior or child functioning. A pair of small-sample studies (N = 6 and 11) conducted a paced breathing task with caregivers and their child either prenatally (Van Leeuwen et al., Reference Van Leeuwen, Geue, Thiel, Cysarz, Lange, Romano and Grönemeyer2009) or within the first 12 weeks of life (Van Puyvelde et al., Reference Van Puyvelde, Loots, Meys, Neyt, Mairesse, Simcock and Pattyn2015) demonstrating that, even at this early stage of development, caregivers and children have the ability to coordinate their parasympathetic activity. Furthermore, synchrony increased or decreased at higher or lower maternal respiratory rates, respectively. However, Van Puyvelde and colleagues also showed that parasympathetic synchrony disappeared by 12 weeks. This was analyzed using repeated measures analysis of variance (rmANOVA), which cannot fully tease apart within-dyad associations from between-dyad differences. Thus, these studies provide only preliminary evidence for the presence of parasympathetic synchrony in the perinatal period that must be corroborated with multilevel modeling techniques.
Later in infancy, vocal and affect synchrony tend to be associated with increased parasympathetic synchrony (Feldman et al., Reference Feldman, Magori-Cohen, Galili, Singer and Louzoun2011). Caregivers may support synchrony via their own arousal levels, as parasympathetic synchrony tends to be higher when the caregiver was assigned to a “relaxation” condition before the interaction but not if the caregiver was assigned to a “stress” condition (Waters et al., Reference Waters, West, Karnilowicz and Mendes2017). Child-driven effects are apparent as well, because if mothers show increased arousal in response to higher infant arousal (reduced HRV, stronger parasympathetic synchrony), infants tend to quiet more quickly (Wass et al., Reference Wass, Smith, Clackson, Gibb, Eitzenberger and Mirza2019). Importantly, though, this was not true if the mother's arousal level was already high, in which case reductions in mother arousal predicted faster infant quieting. Conversely, Ostlund et al. (Reference Ostlund, Measelle, Laurent, Conradt and Ablow2017) found negative parasympathetic synchrony during the reunion phase of the Still-Face Paradigm, when infants tended to show increasing RSA (calming) while mothers showed decreasing RSA (increased arousal). Another study found no parasympathetic synchrony during either phase of the Still-Face Paradigm (Ham & Tronick, Reference Ham and Tronick2009), though the sample size was small (N = 18) and likely did not afford sufficient power to detect synchrony. Thus, positive parasympathetic synchrony may be most adaptive when overall arousal levels are low, and negative parasympathetic synchrony may be more helpful when one or both individual's arousal levels are high; however, future research is needed regarding the developmental correlates of parasympathetic synchrony to support this conclusion.
Another study found a negative bivariate correlation between maternal and infant parasympathetic reactivity while the mother completed social interviews with an experimenter and with the infant on her lap (Waters & Mendes, Reference Waters and Mendes2016; same sample as Waters et al., Reference Waters, West and Mendes2014). However, due to methodological considerations (unaddressed variations in speech rate across tasks that affect respiration and parasympathetic activity; the primary interaction is between the mother and experimenter, not mother and infant), this study was determined not to fit the criteria for measuring physiological synchrony as presently defined.
Toddlerhood and early childhood (12 studies)
Toddlerhood is typically regarded as a developmental period when self-regulation skills develop (Calkins, Reference Calkins, C. A. and C. B.2007; Fox & Calkins, Reference Fox and Calkins2003), and may therefore signal a shift in the quality and importance of physiological synchrony. It is possible that during this period, reliance on the caregiver's physiological regulation is counter-productive to the development of a child's own self-regulation skills. Conversely, synchrony may be an important mechanism through which toddlers develop those skills (e.g., Wass et al., Reference Wass, Smith, Clackson, Gibb, Eitzenberger and Mirza2019). Parasympathetic synchrony has been shown to increase across the preschool period using rmANOVA (Bornstein & Suess, Reference Bornstein and Suess2000), possibly indicating a typical developmental process that promotes parasympathetic synchrony. However, Helm et al. (Reference Helm, Miller, Kahle, Troxel and Hastings2018) found that parasympathetic synchrony disappears in dyads with 3–4-year-olds after de-trending, suggesting different analytic techniques lead to different conclusions. Regardless, several studies that use strong multilevel modeling strategies demonstrate that parasympathetic synchrony in toddlerhood is associated with lower maternal and child psychopathological symptoms (Gray et al., Reference Gray, Lipschutz and Scheeringa2018; Lunkenheimer et al., Reference Lunkenheimer, Tiberio, Buss, Lucas-Thompson, Boker and Timpe2015 & Reference Lunkenheimer, Tiberio, Skoranski, Buss and Cole2018b, same sample), increased maternal teaching and engagement (Skoranski et al., Reference Skoranski, Lunkenheimer and Lucas-Thompson2017; same sample as Lunkenheimer et al., Reference Lunkenheimer, Tiberio, Buss, Lucas-Thompson, Boker and Timpe2015; Reference Lunkenheimer, Tiberio, Skoranski, Buss and Cole2018b), and a lack of a history of maltreatment (Creaven et al., Reference Creaven, Skowron, Hughes, Howard and Loken2014). Shih et al. (Reference Shih, Quiñones-Camacho, Karan and Davis2019) found that parent parasympathetic activity during a frustrating puzzle task positively predicted child parasympathetic activity during recovery, but did not test for associations with behavior or child functioning.
Risk factors in the family and parent–child relationship may moderate this association. For dyads at high socioeconomic risk, behavioral synchrony during a collaborative Etch-a-Sketch™ task and child self-regulation were higher if parasympathetic synchrony was lower (Suveg et al., Reference Suveg, Shaffer and Davis2016). Dyads with an insecure resistant attachment (Smith et al., Reference Smith, Woodhouse, Clark and Skowron2016) and physically abusive dyads (Lunkenheimer et al., Reference Lunkenheimer, Busuito, Brown and Skowron2018a) have shown synchronously decreasing PNS activity (i.e., stress response) to the Strange Situation Procedure and a challenging puzzle task, respectively, while nonmaltreating dyads showed synchronously increasing PNS activity (i.e., social engagement; Lunkenheimer et al., Reference Lunkenheimer, Busuito, Brown and Skowron2018a). The same group found that moments of repair after conflict coincided with decreasing parasympathetic synchrony for both maltreating and nonmaltreating dyads (Lunkenheimer et al., Reference Lunkenheimer, Busuito, Brown, Panlilio and Skowron2019; same sample as Lunkenheimer et al., Reference Lunkenheimer, Busuito, Brown and Skowron2018a). Thus, similar to infancy, parasympathetic synchrony may be particularly helpful during periods of low stress, but not during engagement of the stress response or for dyads with more family-level risk factors. To more directly test this hypothesis, future studies should include both positive and stress-eliciting tasks for dyads with varying caregiver–child relationship characteristics.
Late childhood and adolescence (6 studies)
Adolescence is an important developmental period characterized by large shifts in physiological regulation and emotional well-being (Natsuaki et al., Reference Natsuaki, Klimes-Dougan, Ge, Shirtcliff, Hastings and Zahn-Waxler2009; Spear, Reference Spear2009). Particularly related to the social buffering of HPA reactivity seen among caregivers and their infants, adolescence seems to be the time when caregivers are no longer able to effectively buffer their child's stress response (e.g., Doom, Hostinar, VanZomeren-Dohm, & Gunnar, Reference Doom, Hostinar, VanZomeren-Dohm and Gunnar2015) and overall stress responses tend to increase in magnitude across adolescence (Gunnar, Wewerka, Frenn, Long, & Griggs, Reference Gunnar, Wewerka, Frenn, Long and Griggs2009b). Still, all but one of the studies examining caregiver–adolescent parasympathetic synchrony in this review have found that greater synchrony during both positive and negative discussion tasks is associated with fewer internalizing symptoms and less negative affect for both individuals in the dyad (Amole et al., Reference Amole, Cyranowski, Wright and Swartz2017; Han et al., Reference Han, Gao, Yan, Hu, Zhou and Li2019; McKillop & Connell, Reference McKillop and Connell2018; Suveg et al., Reference Suveg, Braunstein West, Davis, Caughy, Smith and Oshri2019; Woody et al., Reference Woody, Feurer, Sosoo, Hastings and Gibb2016). These studies tended to have relatively larger sample sizes (largest N = 150) and used strong multilevel modelling techniques. Of these studies, the only one (Amole et al., Reference Amole, Cyranowski, Wright and Swartz2017) that described the direction of synchronous physiological activity across the task found synchronous, increasing parasympathetic synchrony for nondepressed, but not depressed, dyads. Therefore, if parasympathetic synchrony is most useful in low stress contexts as it appears in infancy and toddlerhood, this pattern of findings suggests that caregiver–adolescent dyads who are able to maintain parasympathetic synchrony during an interaction are also able to maintain lower overall levels of arousal during discussion.
Conversely, stronger positive synchrony while simultaneously viewing positive and negative film clips was found to be associated with lower child self-reported empathy, especially when parents were rated lower on emotional acceptance (Creavy et al., Reference Creavy, Gatzke-Kopp, Zhang, Fishbein and Kiser2020). While this study also used strong statistical methods assessing within-dyad associations accounting for overall between-dyad differences, simultaneously watching the same film clip does not necessarily support interpersonal interaction and so this study does not fully meet the definition of physiological synchrony proposed here.
Overall, the general body of research on parasympathetic synchrony suggests that it is associated with positive behavioral interaction and supports better child behavioral and emotional regulation, though future longitudinal studies are needed to examine the direction of these associations (synchrony to behavior vs. behavior to synchrony, or both).
Sympathetic nervous system
Biological processes
As coordinated branches of the ANS, the PNS works in tandem with the SNS such that reductions in PNS activity impact SNS activity, and vice versa. This PNS×SNS interaction helps the body quickly calibrate in response to constantly changing environmental circumstances (Porges & Furman, Reference Porges and Furman2012). Vagal withdrawal is not required to see increases in SNS activity, but the PNS and SNS tend to perform inverse functions (Porges, Reference Porges2007; Ulrich-Lai & Herman, Reference Ulrich-Lai and Herman2009). During vagal withdrawal, the SNS increases its influence on its target organs, typically resulting in increased heart rate and blood pressure, shortened pre-ejection period (PEP) of the heart, and dilated pupils and airways (Ulrich-Lai & Herman, Reference Ulrich-Lai and Herman2009). The sympatho-adrenomedullary (SAM) system then releases preganglionic acetylcholine via the splanchnic nerve into the adrenal medulla, stimulating the release of postganglionic epinephrine (and a small amount of norepinephrine) into the bloodstream. Circulating epinephrine enhances the fight-or-flight response through its actions on target organs and a positive feedback loop with the SNS (Gunnar & Quevedo, Reference Gunnar and Quevedo2007). This process mobilizes the body's resources to respond to threat and challenge and sets in motion a number of evolutionarily adaptive metabolic and hormonal changes, particularly for physical and performance stressors.
SNS activity is quantified in a number of ways, including PEP and skin conductance (SC). Epinephrine also stimulates the production of salivary alpha-amylase (sAA) via activation of alpha- and beta-adrenergic receptors, which has been used as a peripheral biomarker of SNS activation, though it may more accurately be a marker of both PNS and SNS activation (Nater & Rohleder, Reference Nater and Rohleder2009). Still, many studies find a positive association between stress and sAA, and sAA seems to reach peak levels a few minutes after stressor onset (Stroud et al., Reference Stroud, Foster, Papandonatos, Handwerger, Granger, Kivlighan and Niaura2009; West, Granger, Kivlighan, Psota, & Hurston, Reference West, Granger, Kivlighan, Psota and Hurston2006), preceding an increase in cortisol production. SNS activity plays a role in the activation of the HPA axis via stimulation of hypothalamic corticotropin releasing hormone (CRH; Engert et al., Reference Engert, Vogel, Efanov, Duchesne, Corbo, Ali and Pruessner2011). This is adaptive in the short term, but prolonged SNS activity can shift the autonomic balance away from parasympathetic regulation, overactivate the HPA axis, and ultimately reduce an individual's ability to regulate and respond to their environment (Del Giudice, Ellis, & Shirtcliff, Reference Del Giudice, Ellis and Shirtcliff2011).
Measurement of caregiver–child sympathetic synchrony
Measures of SNS activity in caregivers and children have typically been used as an indicator of acute stress and arousal. Parenting behavior tends to correlate with children's SNS reactivity to stressors (Oosterman, De Schipper, Fisher, Dozier, & Schuengel, Reference Oosterman, De Schipper, Fisher, Dozier and Schuengel2010) presumably via the impact of parenting on children's ability to regulate their own arousal and/or the caregiver's ability to regulate their children's arousal. It also seems that both caregivers and children can “transmit” their arousal to the other interacting partner (Lougheed & Hollenstein, Reference Lougheed and Hollenstein2018; Waters et al., Reference Waters, West, Karnilowicz and Mendes2017). Thus, sympathetic synchrony may shed light on how dyads reduce or exacerbate one another's arousal, with implications for relationship quality and children's self-regulatory development. It is possible that sympathetic synchrony when individual arousal levels are high reflects a mutual stress response, where one individual's arousal is driving the other's arousal up or both individuals are unable to bring each other's arousal down. Alternatively, sympathetic synchrony during stress may promote empathy and help to mobilize mental resources to overcome the challenge together (Lougheed & Hollenstein, Reference Lougheed and Hollenstein2018).
It is also important to consider the possibility that SNS “synchrony” is actually due to separate, individual responses to an external stimulus that does not fall under the definition of synchrony as a specific interpersonal process. The genetic confound between parents and children in most, if not all, studies of physiological synchrony increases the likelihood that correlations between parent and child physiological reactivity are due to genetic or other shared predispositions rather than the two individuals having an effect on each other in the moment (Wu, Snieder, & de Geus, Reference Wu, Snieder and de Geus2010). This issue likely applies to dyadic synchrony in all physiological systems, but may be particularly pernicious in sympathetic and adrenocortical studies that do not use the appropriate statistical modeling procedures.
Statistically, because SNS activity can be measured on a similar timescale as PNS activity (e.g., PEP, SC), the same modeling techniques can be appropriate (see above for details). However, SNS activity can also be approximated in other ways, for example as sAA in saliva, levels of which fluctuate on a much longer timescale than other more direct measures of SNS activity via the heart. Nonspecific autonomic measures like sAA, heart rate, and thermal facial imprints that constitute a more integrated, distal index of parasympathetic and sympathetic activity may provide a unique perspective of caregiver–child synchrony compared to more direct measures like vagal tone and PEP that more closely represent parasympathetic or sympathetic activity respectively. However, dense sampling of saliva every few minutes (to capture short-term changes in sAA activity) can be disruptive to an interpersonal interaction and still results in a much smaller number of data points per individual relative to heart rate measures that can be collected nonintrusively and continuously throughout a task. Fewer data points per individual reduces the ability to investigate dynamic within-dyad patterns of synchrony compared to between-dyad correlations over time. It is also unlikely that, for example, facial thermal imprints, sAA, and PEP represent the exact same underlying physiological process, reducing our ability to compare across studies. Thus, the findings summarized below will be interpreted in light of their variability in measurement, sample timing, and statistical approach (see Table 1).
Sympathetic and nonspecific autonomic synchrony across development
Infancy (3 studies)
Two studies by the same group found that mother–infant sympathetic synchrony (measured via mother PEP and infant heart rate) was higher after the mother completed tasks (without the infant present) designed to elicit a stress response but not if the mother had completed nonstressful tasks (Waters et al., Reference Waters, West and Mendes2014; Waters et al., Reference Waters, West, Karnilowicz and Mendes2017). The authors believed this to be a “stress contagion” mechanism, where infants are detecting their mothers’ heightened arousal as a signal for potential threat which then results in the activation of the infant's own stress response. These studies did not examine parent or child characteristics as correlates, so it is unclear if “stress contagion” is an adaptive or maladaptive process for children's development. One small sample study (N = 18; Ham & Tronick, Reference Ham and Tronick2009) found that sympathetic synchrony (SC) was associated with infant negative engagement during the still-face phase of the Still Face Paradigm but was associated with behavioral synchrony during the reunion phase. Much more research is needed before conclusions can be drawn about the correlates and consequences of sympathetic synchrony in infancy.
Toddlerhood and early childhood (7 studies)
Of the few studies examining sympathetic synchrony during toddlerhood, one small-sample study (N = 12; Ebisch et al., Reference Ebisch, Aureli, Bafunno, Cardone, Romani and Merla2012) used rmANOVA to show increased sympathetic synchrony (nasal-tip thermal imprint) during a distress task compared to baseline, and this was specific to mother–child dyads and not unrelated woman–child dyads (same sample; Manini et al., Reference Manini, Cardone, Ebisch, Bafunno, Aureli and Merla2013). Using multilevel modeling techniques with relatively larger sample sizes (Ns = 83–104), others found concurrent PEP or heart rate concordance but no association in change over time (Creaven et al., Reference Creaven, Skowron, Hughes, Howard and Loken2014; Helm et al., Reference Helm, Miller, Kahle, Troxel and Hastings2018; Shih et al., Reference Shih, Quiñones-Camacho, Karan and Davis2019), suggesting this is a between- rather than within-dyad phenomenon. One exception (Laurent et al., Reference Laurent, Ablow and Measelle2012, N = 86) found mother–child sAA synchrony during a set of fear- and frustration-eliciting tasks but not during a separation stressor (three saliva samples per task). Neither of these tasks were used in the studies that found no within-individual association, suggesting that sympathetic synchrony may be task-dependent. Certain tasks may be differentially effective in activating the stress response (Dickerson & Kemeny, Reference Dickerson and Kemeny2004; Peters et al., Reference Peters, Godaert, Ballieux, Van Vliet, Willemsen, Sweep and Heijnen1998), which may influence the presence and correlates of sympathetic synchrony. Since none of these studies specified whether dyads showed synchronously increasing or decreasing sympathetic activity, it is unclear whether inconsistent findings are due to differential activation of the stress response. Future studies, preferably using measures that afford denser physiological measurement like SC or PEP, are needed to examine this hypothesis.
Only one sympathetic study examined associations with indices of caregiver behavior and child functioning. Baker et al. (Reference Baker, Fenning, Howland, Baucom, Moffitt and Erath2015) used multilevel modeling to demonstrate that sympathetic synchrony (wrist SC) was positively associated with concurrent emotional attunement during free play and negatively associated with autism spectrum symptoms in dyads with 4- to 10-year-olds. Again, much more (longitudinal) research is needed before conclusions can be drawn about correlates and consequences of sympathetic synchrony in toddlerhood.
Late childhood and adolescence (5 studies)
There is similarly little research examining sympathetic synchrony in adolescence, and the little data that do exist are mixed. The majority also use nonspecific autonomic measures rather than more targeted measures like PEP, which likely contributes to the inconsistency. Two studies, one examining SC and the other heart rate (Lougheed & Hollenstein, Reference Lougheed and Hollenstein2018; Woltering et al., Reference Woltering, Lishak, Elliott, Ferraro and Granic2015), found stronger synchrony in a positive compared to negative discussion, and the magnitude of synchrony was positively associated with the amount of positive behavior during the discussion (Woltering et al., Reference Woltering, Lishak, Elliott, Ferraro and Granic2015) and in mother–adolescent dyads reporting less interparental aggression (Gordis et al., Reference Gordis, Margolin, Spies, Susman and Granger2010). However, for dyads with high levels of overall conflict and, interestingly, hypertension, sympathetic synchrony may be higher during negative compared to positive discussion, (Ghafar-Tabrizi, Reference Ghafar-Tabrizi2008; McClure & Myers, Reference McClure and Myers1999). Gordis et al. (Reference Gordis, Margolin, Spies, Susman and Granger2010) also noted that interparental aggression was positively associated with father–adolescent sympathetic synchrony, while it was negatively associated with mother–adolescent synchrony. Taking these studies together, sympathetic synchrony tends to be positively associated with caregiver–adolescent relationship quality, particularly during positive interactions, though results are mixed. Future studies should attempt to identify potential moderators that may explain these discrepancies.
Hypothalamic–pituitary–adrenocortical axis
Biological processes
On the order of minutes to hours, stressors initiate an increase in HPA activity that results in a relatively gradual increase in the glucocorticoid cortisol throughout the bloodstream, followed by a similarly gradual decrease back to baseline. While the HPA axis is activated by a variety of different stressors, it is most reliably activated during social-evaluative threat (Dickerson & Kemeny, Reference Dickerson and Kemeny2004) and, in children, the threat of caregiver separation (Gunnar, Talge, & Herrera, Reference Gunnar, Talge and Herrera2009a). The adrenal cortex releases the steroid hormone cortisol into the bloodstream in phasic bursts, with peak cortisol levels occurring approximately 20–40 minutes post-stressor (Kirschbaum & Hellhammer, Reference Kirschbaum and Hellhammer1994). This process delays its wide-ranging effect on the brain and body relative to that of the PNS and SNS. Soon after it is released into the bloodstream, cortisol increases in saliva, where it is commonly measured as the end-product of HPA axis activity. Furthermore, the products of HPA activity (including cortisol) contribute to a negative feedback loop such that increased levels of these hormones cause a subsequent downregulation of activity at higher levels of the HPA axis (Ulrich-Lai & Herman, Reference Ulrich-Lai and Herman2009).
Aside from faster, cell membrane-mediated responses, HPA axis activity also initiates long-term changes in gene transcription and epigenetic regulation across a number of systems, including upstream regions of the HPA axis (Strüber, Strüber, & Roth, Reference Strüber, Strüber and Roth2014). Thus, the HPA axis facilitates enduring alterations to system regulation following stress and challenge. These changes might be adaptive, as when systems are altered to better deal with a stressor, should something similar be encountered again. However, sometimes the HPA axis is activated too often or for too long, as with chronic stress and trauma. Prolonged HPA activity increases allostatic load, or the long-term wear and tear that occurs in the brain and body after chronic activation of the stress response (McEwen, Reference McEwen2006), and may result in HPA hypoactivity over time, overcompensating for the initial overactivity (Fries, Hesse, Hellhammer, & Hellhammer, Reference Fries, Hesse, Hellhammer and Hellhammer2005). This is a problem, because the ability to mount a moderate, brief HPA response to stress or challenge is known to be important for healthy functioning (Finsterwald & Alberini, Reference Finsterwald and Alberini2014).
Measurement of caregiver–child adrenocortical synchrony
Caregiver–child adrenocortical synchrony poses the most statistical challenges due to its timescale of activity and methodological limitations. As discussed above, it takes several minutes for a stimulus to ultimately result in changes in the level of salivary cortisol, so measurement of moment-to-moment dynamic synchrony is already limited. Aside from a continuous blood draw, salivary cortisol is the best way to assess short-term changes in HPA activity, resulting in a more delayed representation of HPA activity and a much wider sampling interval compared to autonomic heart rate measures. This affects the type of statistical modeling that can be applied and the sample size required for adequate power to detect meaningful within-dyad effects. Sampling interval may also influence how much of a time-lag should be considered, if any. For such a slow-acting system, concurrent measures of adrenocortical synchrony might be most appropriate to capture the relevant timescale of this interpersonal process, though this has yet to be determined.
An additional challenge is that when examining adrenocortical synchrony during a stressor, the most potent stressors for infants and toddlers involve caregiver separation or disconnection (e.g., Strange Situation Procedure and Still-Face Paradigm; Gunnar et al., Reference Gunnar, Talge and Herrera2009a). By design, this interrupts the interpersonal process that is expected to facilitate physiological synchrony and may affect interpretation of adrenocortical synchrony. It is also important to note that the adrenocortical synchrony studies included below are some of the farthest from statistically quantifying “synchrony” as presently defined. Most use correlational statistical approaches or rmANOVA and generally use other terms like “attunement,” which may hinder the field's ability to define this complex process and develop a common language surrounding it.
Adrenocortical synchrony across development
Infancy (18 studies)
The vast majority of caregiver–child adrenocortical synchrony studies were conducted in infancy, all of which examined dyadic salivary cortisol in response to a stressor (see Table 1). There is a substantial body of literature demonstrating that sensitive, responsive caregivers buffer their children's stress responses particularly in infancy (Gunnar, Brodersen, Nachmias, Buss, & Rigatuso, Reference Gunnar, Brodersen, Nachmias, Buss and Rigatuso1996; Gunnar & Hostinar, Reference Gunnar and Hostinar2015; Luijk et al., Reference Luijk, Saridjan, Tharner, van IJzendoorn, Bakermans-Kranenburg, Jaddoe and Tiemeier2010), and harsh, insensitive, and negative caregivers contribute to general dysregulation of their children's HPA axes (DePasquale, Raby, Hoye, & Dozier, Reference DePasquale, Raby, Hoye and Dozier2018; Dougherty, Klein, Rose, & Laptook, Reference Dougherty, Klein, Rose and Laptook2011). A caregiver's own HPA activity may play a role in Caregiver×Child interaction quality, and adrenocortical synchrony may signify adaptive or maladaptive dyadic coregulation patterns. It seems that infants distress vocalizations can influence caregiver adrenocortical activity and therefore synchrony (Middlemiss et al., Reference Middlemiss, Granger, Goldberg and Nathans2012). The social buffering hypothesis suggests that caregivers may buffer their infants’ stress responses via the effective regulation of their own (Gordon, Zagoory-Sharon, Leckman, & Feldman, Reference Gordon, Zagoory-Sharon, Leckman and Feldman2010; Ham & Tronick, Reference Ham and Tronick2009), though it is yet unclear how that would manifest as adrenocortical synchrony, if at all.
Several primarily large sample studies (Ns = 233–1,141) and two smaller sample studies (N = 47 and 83) have shown a stronger (or less negative) association between caregiver and infant cortisol reactivity when the caregiver is more positive and responsive and report greater marital satisfaction (Atkinson et al., Reference Atkinson, Gonzalez, Kashy, Santo Basile, Masellis, Pereira and Levitan2013; Clauss et al., Reference Clauss, Byrd-Craven, Kennison and Chua2018; Hendrix et al., Reference Hendrix, Stowe, Newport and Brennan2018; Hibel et al., Reference Hibel, Granger, Blair and Finegood2015; Nofech-Mozes et al., Reference Nofech-Mozes, Jamieson, Gonzalez and Atkinson2019; van Bakel & Riksen-Walraven, Reference van Bakel and Riksen-Walraven2008). Other studies (with similarly large samples) have found stronger adrenocortical synchrony in dyads from households with higher rates of violence or punitive/intrusive parenting behaviors (Hibel & Mercado, Reference Hibel and Mercado2019; Hibel et al., Reference Hibel, Granger, Blair and Cox2009). Stronger adrenocortical synchrony during Still-Face has predicted more disrupted maternal communication in the reunion episode of the task (Crockett et al., Reference Crockett, Holmes, Granger and Lyons-Ruth2013). Khoury et al. (Reference Khoury, Gonzalez, Levitan, Masellis, Basile and Atkinson2016; same sample as Atkinson et al., Reference Atkinson, Gonzalez, Kashy, Santo Basile, Masellis, Pereira and Levitan2013) and Laurent et al. (Reference Laurent, Ablow and Measelle2011) both demonstrated a positive association between maternal depressive symptoms and adrenocortical synchrony during the Strange Situation. Other small sample studies (N = 42 and 63) have found no association with caregiver-related factors (Castral et al., Reference Castral, Warnock, Dos Santos, Daré, Moreira, Antonini and Scochi2015; Thompson & Trevathan, Reference Thompson and Trevathan2009). Infant pre-term status and distress during a three-day sleep training program also predicted increased adrenocortical synchrony (Middlemiss et al., Reference Middlemiss, Granger, Goldberg and Nathans2012; Provenzi et al., Reference Provenzi, Giusti, Fumagalli, Frigerio, Morandi, Borgatti and Montirosso2019). Cortisol synchrony was found in a sample of low-income Mexican mother–child dyads (Luecken et al., Reference Luecken, Crnic, Gonzales, Winstone and Somers2019); however, they did not include a higher-income comparison group or examine associations with indices of caregiver or child functioning, impeding the interpretation of these findings.
As mentioned above, different stressors can be differentially effective in activating the HPA axis (Dickerson & Kemeny, Reference Dickerson and Kemeny2004; Gunnar et al., 2009). When only considering studies using well-validated stressors known to elicit a stress response in infants (Strange Situation and Still-Face) and strong multilevel modeling techniques, correlates of adrenocortical synchrony are generally positive (Atkinson et al., Reference Atkinson, Gonzalez, Kashy, Santo Basile, Masellis, Pereira and Levitan2013; Nofech-Mozes et al., Reference Nofech-Mozes, Jamieson, Gonzalez and Atkinson2019; but see also Laurent et al., Reference Laurent, Ablow and Measelle2011). Multilevel modeling techniques tend to be more conservative than correlational techniques in analyzing associations between synchrony and other variables (Bernard et al., Reference Bernard, Kashy, Levendosky, Bogat and Lonstein2017). For example, Bernard et al. (Reference Bernard, Kashy, Levendosky, Bogat and Lonstein2017) found that interparental violence moderated adrenocortical synchrony such that synchrony was stronger in dyads with low interparental violence. No such moderation was found when using multilevel growth curve models, suggesting that the correlational findings described above may be spurious.
In addition, it is possible that the Strange Situation and Still-Face are inappropriate for the study of physiological synchrony because they seem to interrupt Caregiver×Child interaction. However, Laurent et al. (Reference Laurent, Ablow and Measelle2012; same sample as Laurent et al., Reference Laurent, Ablow and Measelle2011), found that adrenocortical synchrony was actually stronger during the Strange Situation compared to a frustration task. Findings using these tasks should be interpreted with caution until future studies can demonstrate the interpersonal mechanisms that may explain the presence of adrenocortical synchrony during the Strange Situation, Still-Face, and similar tasks. Thus, strong conclusions cannot yet be drawn from these studies.
Toddlerhood and early childhood (5 studies)
Surprisingly, the majority of evidence regarding adrenocortical synchrony in toddlerhood suggests that synchrony is associated with more positive parenting and better child functioning. Studies examining cortisol production during both positive- (free play; Ruttle et al., Reference Ruttle, Serbin, Stack, Schwartzman and Shirtcliff2011) and negative-valence tasks (exposure to challenge/novel stimuli; Kalomiris & Kiel, Reference Kalomiris and Kiel2018; Sethre-Hofstad et al., Reference Sethre-Hofstad, Stansbury and Rice2002) using a range of statistical approaches found that stronger adrenocortical synchrony was associated with higher parental sensitivity and reduced child internalizing problems. Further, adrenocortical synchrony was weaker for dyads with children with autism spectrum disorder (ASD; Saxbe et al., Reference Saxbe, Golan, Ostfeld-Etzion, Hirschler-Guttenberg, Zagoory-Sharon and Feldman2017), though they also found a negative association between adrenocortical synchrony and behavioral sensitivity. Importantly, Saxbe et al. (Reference Saxbe, Golan, Ostfeld-Etzion, Hirschler-Guttenberg, Zagoory-Sharon and Feldman2017) used multilevel modeling; another study used a correlational analysis and found no differences in adrenocortical synchrony for dyads with children with or without ASD (Ostfeld-Etzion et al., Reference Ostfeld-Etzion, Golan, Hirschler-Guttenberg, Zagoory-Sharon and Feldman2015). These studies generally support the notion that adrenocortical synchrony is associated with positive indices of children development, though more (longitudinal) research is needed to strengthen this claim for a wider variety of developmental domains.
Late childhood and adolescence (4 studies)
Caregiver adolescent adrenocortical synchrony was shown to be higher in triads with both biological parents compared to stepparents and in triads with daughters instead of sons (Saxbe et al., Reference Saxbe, Margolin, Spies Shapiro, Ramos, Rodriguez and Iturralde2014), though they did not examine associations with indices of family or child functioning. When the same research group asked adolescents to rate their parents’ emotions after a conflict discussion, stronger adrenocortical synchrony during the discussion was associated with less deactivation of posteromedial brain regions involved in the default mode network and social cognition (Saxbe et al., Reference Saxbe, Del Piero and Margolin2015a), suggesting a better ability to reflect on others’ emotions. Others found that adrenocortical synchrony was associated with lower maternal overcontrol during a challenging puzzle task (Borelli et al., Reference Borelli, Shai, Smiley, Boparai, Goldstein, Rasmussen and Granger2019). However, adrenocortical synchrony has also been suggested as a potential mediator for the intergenerational transmission of risk for psychopathology in war-exposed families (Halevi et al., Reference Halevi, Djalovski, Kanat-Maymon, Yirmiya, Zagoory-Sharon, Koren and Feldman2017). Still, the two studies that use multilevel modeling strategies tend to suggest that adrenocortical synchrony is associated with better Parent×Adolescent interaction quality and greater adolescent emotion awareness (Borelli et al., Reference Borelli, Shai, Smiley, Boparai, Goldstein, Rasmussen and Granger2019; Saxbe et al., Reference Saxbe, Del Piero and Margolin2015a). Though no strong conclusions can be drawn from two studies, future research should expand upon these findings and the similarly positive (though preliminary) findings for adrenocortical synchrony across all developmental periods.
The central nervous system and neurotransmitters
Biological processes
A number of neurotransmitter receptors and brain regions exert a top-down influence on autonomic and HPA activity. CRH and glucocorticoid receptors are expressed widely throughout the brain, implicating a range of brain regions in HPA activity and negative feedback regulation. Glucocorticoid receptors (specifically high-affinity mineralocorticoid receptors; MRs) in the hippocampus aid in post-stressor information processing and memory formation (de Kloet, Karst, & Joëls, Reference de Kloet, Karst and Joëls2008; Joëls & Baram, Reference Joëls and Baram2009). Lower-affinity glucocorticoid receptors (GRs) in the hippocampus and hypothalamus are involved in HPA axis negative feedback, suppressing future cortisol production. GRs in the medial and basolateral nuclei of the amygdala assist increase HPA activity, partially via the suppression of the hippocampus (Ulrich-Lai & Herman, Reference Ulrich-Lai and Herman2009). Amygdalar CRH production also rapidly activates autonomic activity through increased epinephrine (Brown, Fisher, Webb, Vale, & Rivier, Reference Brown, Fisher, Webb, Vale and Rivier1985; Yamaguchi-Shima et al., Reference Yamaguchi-Shima, Okada, Shimizu, Usui, Nakamura, Lu and Yokotani2007).
Regions of the medial prefrontal cortex (mPFC) have also been implicated in the regulation of the physiological stress response. Prelimbic regions of the mPFC tend to be associated with inhibition or termination of the HPA stress response, while ventromedial regions have been found to stimulate both autonomic and HPA activity (Hänsel & von Känel, Reference Hänsel and von Känel2008; Ulrich-Lai & Herman, Reference Ulrich-Lai and Herman2009). Relatedly, dorsolateral and medial regions of the PFC have been implicated in caregiver–child behavioral synchrony/cooperation (Reindl et al., Reference Reindl, Gerloff, Scharke and Konrad2018) and prosociality in unfamiliar female college student dyads (Hu, Hu, Li, Pan, & Cheng, Reference Hu, Hu, Li, Pan and Cheng2017) suggesting a role in dyadic interaction, though their role in the regulation of the stress response is not fully elucidated. The anterior cingulate cortex (ACC), involved in processing emotions and social salience (Atzil, Hendler, & Feldman, Reference Atzil, Hendler and Feldman2013), shows extensive functional connectivity with the mPFC (Etkin, Egner, & Kalisch, Reference Etkin, Egner and Kalisch2011). Activity in this region, specifically the dorsal ACC, is associated with maternal behavioral synchrony when watching clips of synchronous and asynchronous interactions (Atzil et al., Reference Atzil, Hendler and Feldman2013) and has been found to play a role in ongoing behavioral adaptation to changing environmental circumstances (Sheth et al., Reference Sheth, Mian, Patel, Asaad, Williams, Dougherty and Eskandar2012). These functions are likely critical for maintaining behavioral synchrony and may also be crucial to understanding the function of physiological synchrony.
Lower brain regions in the temporal area and regions of the brain stem are primarily involved in autonomic regulation. In concert with the central nucleus of the amygdala, a primitive region of the brain stem called the periaqueductal gray (PAG) initiates flight–flight–freeze behaviors associated with SNS activity (Porges, Reference Porges2007; Ulrich-Lai & Herman, Reference Ulrich-Lai and Herman2009). Temporal regions including the fusiform gyrus and the superior temporal sulcus (STS) serve to detect threat such that, in the absence of threat, these regions have inhibitory connections with the amygdala which reduces fight-or-flight behavior and increases social behavior (likely reflecting a preponderance of parasympathetic, rather than sympathetic, autonomic activity; Porges, Reference Porges2007). Interestingly, activity in regions like the STS and other temporo-parietal regions is associated with behavioral synchrony for parent–child dyads (Levy et al., Reference Levy, Goldstein and Feldman2017), hinting at their potential importance for caregiver–child physiological synchrony.
Measurement of caregiver–child central nervous system synchrony
There is still considerable debate among neuroscientists about how to contextualize an individual's brain activity within their environment and more tangible aspects of mental functioning, so it is no surprise that dyadic CNS synchrony compounds those difficulties. The limitations of what can be done in a magnetic resonance imaging (MRI) scanner reduce the ecological validity of interpersonal processes that are foundational to the study of caregiver–child synchrony. Some have improvised by instead conducting MRI scans while an individual watches a video of their own pre-recorded dyadic interaction, but this poses interpretive challenges of what the CNS activity actually means contextually. With regard to the proposed definition of synchrony, CNS “synchrony” when two individuals separately watch their pre-recorded interaction may represent two individuals processing the same external stimuli, rather than an interpersonal process. Furthermore, MRI scanning limits the age range of child participants due to movement artifacts, particularly in toddlers, which is likely an important developmental period to examine the neural bases of caregiver–child synchrony and child functioning.
More portable measurement methods like electroencephalography (EEG) and functional near-infrared spectroscopy (fNIRS) may balance the limitations of MRI by increasing ecological validity, at the expense of spatial precision. EEG and fNIRS use a constellation of electrodes or optodes placed on the scalp to detect electrical or hemodynamic fluctuations in brain tissue, respectively. These methods can be incorporated more naturally in studies involving dyadic interaction compared to methods like MRI and magnetoencephalography (MEG) and, due to their high temporal precision, should hypothetically allow for the use of similar statistical methods as those described for autonomic synchrony (see above for details). EEG and fNIRS cannot achieve the same specificity in targeting precise cortical, and especially subcortical, regions as with MRI, but they should still be leveraged to understand what parent–child inter-brain synchrony looks like, how it manifests and unfolds, as well as the general regions in which synchrony is associated with dyadic behavior and child functioning (for better or worse).
Central nervous system synchrony (7 studies)
There are only seven studies included in this review that examine caregiver–child CNS synchrony which does not allow us to sufficiently examine differences across development, so these studies have been combined into one section. Nevertheless, these studies have the potential to shed light on the neurobiological underpinnings of synchrony. Interpersonal coordination of brain regions already known to be involved in the regulation of autonomic and adrenocortical synchrony may provide a much deeper understanding of how synchrony manifests in real time.
First, when mothers were instructed to display positive emotion towards an object in front of her infant, global neural synchrony (EEG) was higher and also showed stronger mother-to-infant directionality compared to when mothers showed negative emotion towards an object (Santamaria et al., Reference Santamaria, Noreika, Georgieva, Clackson, Wass and Leong2020). These data may have interesting implications for understanding how infants learn about the world around them and begin to participate in social exchanges. In addition, fNIRS data suggest the presence of synchrony in the dorsolateral prefrontal and frontopolar cortices of children with their caregivers during a cooperation task, but not a competition task or either task with a stranger (Reindl et al., Reference Reindl, Gerloff, Scharke and Konrad2018) and regions of the prefrontal cortex showed increased synchrony in mother–son dyads when completing a task cooperatively compared to when they completed it independently (Miller et al., Reference Miller, Vrtička, Cui, Shrestha, Hosseini, Baker and Reiss2019). Infants and young children who show prefrontal and temporo-parietal synchrony with their parent (fNIRS) tend to show more behavioral synchrony, less irritability, and more problem-solving success (Nguyen et al., Reference Nguyen, Schleihauf, Kayhan, Matthes, Vrtička and Hoehl2020; Quiñones-Camacho et al., Reference Quiñones-Camacho, Fishburn, Camacho, Hlutkowsky, Huppert, Wakschlag and Perlman2019). These prefrontal regions have already been implicated in the negative feedback regulation of the HPA axis and autonomic reactivity (Ulrich-Lai & Herman, Reference Ulrich-Lai and Herman2009). Therefore, prefrontal synchrony may play a role in establishing adrenocortical and autonomic synchrony, though this has yet to be examined.
A region-of-interest analysis of similarity in emotion-related brain regions (including the ACC) found that mother–adolescent dyads had higher similarity following an adolescent stressor – and the mother's viewing of the stressor – if mothers and adolescents both reported higher family connectedness (Lee et al., Reference Lee, Qu and Telzer2018). Caregiver–child dyads watching video-recorded vignettes of their own discussions have also shown increased synchrony in the STS when they re-watched periods of behavioral synchrony compared to asynchronous moments (Levy et al., Reference Levy, Goldstein and Feldman2017). Because the STS is involved in threat detection and inhibition of the amygdala, synchrony in this region may reflect a critical stress-buffering process in supportive caregiver–child dyads that extends to adolescence. These findings are particularly encouraging because, if caregivers can still play a role in regulating their children's brain activity in adolescence, it may be possible to harness this process to support adolescent well-being during a period of heightened risk for stress dysregulation and mental health problems. Nevertheless, these two studies used functional magnetic resonance imaging (fMRI) and MEG, respectively, while individuals watched a pre-recorded interaction (Levy et al., Reference Levy, Goldstein and Feldman2017) or an adolescent stressor (Lee et al., Reference Lee, Qu and Telzer2018). These methods allowed them to identify regions of interest for caregiver–child synchrony that would be difficult to identify with other methods like EEG and fNIRS, but they limit the conclusions that can be made about CNS synchrony. Instead, these studies should be considered preliminary data for future CNS synchrony studies.
Several brain regions described above have direct or indirect influences on PNS, SNS, and HPA activity. CNS synchrony using portable measures like EEG and fNIRS may be perfectly situated to provide valuable information about how physiological synchrony operates in real time and the specific neural mechanisms associated with it. The data that do exist seem to uniformly suggest positive implications for children's emotional development. Further, though it does not eliminate the possibility, the differentiation between caregiver–child and stranger–child dyads and cooperative versus independent tasks (Miller et al., Reference Miller, Vrtička, Cui, Shrestha, Hosseini, Baker and Reiss2019; Reindl et al., Reference Reindl, Gerloff, Scharke and Konrad2018) demonstrates that parallel processing of the same external stimuli likely does not fully explain the apparent presence of CNS “synchrony.” It is therefore surprising that there are not more studies of this kind examining the phenomena of neural and physiological synchrony. Future research should incorporate these modalities in an ecologically valid way in order to better understand how behavioral and physiological synchrony are related and what implications it has for children's self-regulatory and mental health outcomes.
Discussion
The objectives of this review were to (a) characterize the state of the literature regarding caregiver–child physiological synchrony and its associations with other indices of child functioning, (b) examine developmental differences in the correlates of synchrony, and (c) explore whether context, risk, and/or stress influence patterns of synchrony. First, there is no evidence that synchrony is universally positive for child functioning across systems and in all situations. The most salient take-away message should be that vastly more research is needed to understand this complex phenomenon. When separating by physiological system and developmental period, there was generally a small number of studies to draw from, with few exceptions (e.g., parasympathetic synchrony in toddlerhood, adrenocortical synchrony in infancy). Many studies simply stated whether caregiver–child physiology was synchronous or not; only two studies measured synchrony over multiple time points (Bornstein & Suess, Reference Bornstein and Suess2000; Hibel et al., Reference Hibel, Granger, Blair and Finegood2015). Only one measured both autonomic and adrenocortical synchrony (Laurent et al., Reference Laurent, Ablow and Measelle2012), though many assessed a combination of autonomic measures. Variations in the stability of synchrony over time, and the coordination across systems within dyads, may better contextualize physiological synchrony and the function it serves for children's self-regulatory development.
Statistical approaches and methodology are lacking, particularly for sympathetic and adrenocortical studies, which could have serious implications for our understanding of the construct of physiological synchrony and its correlates (Bernard et al., Reference Bernard, Kashy, Levendosky, Bogat and Lonstein2017; Helm et al., Reference Helm, Miller, Kahle, Troxel and Hastings2018). Many studies could not tease apart between- versus within-dyad effects, which precludes the ability to interpret the data as specifically “physiological synchrony” rather than just two individuals’ physiological activity that tends to be higher or lower than others.’ Few studies describe the individual physiological trajectories that underlie dyadic synchrony. Positive synchrony as presently defined can reflect both mutually increasing and mutually decreasing individual trajectories, which could substantially impact interpretation of study findings (e.g., Lunkenheimer et al., Reference Lunkenheimer, Busuito, Brown and Skowron2018a).
Several studies measured physiological activity without a direct interpersonal interaction, which should not be classified as synchrony according to the definition put forward in this review. When physiological activity is measured in an interpersonal context, correlates of, for example, parent behavior are not always measured concurrently (e.g., parental sensitivity during free play predicting adrenocortical attunement during a stress task; Hibel et al., Reference Hibel, Granger, Blair and Finegood2015). These data can still represent physiological synchrony, though parent behavior not measured concurrent to synchrony should be interpreted as an index of broader relationship quality rather than an assumption about how the parent acted at the time synchrony was being measured. Finally, there is great potential in portable modalities for assessing CNS synchrony in more ecologically valid ways (e.g., EEG and fNIRS) to improve our understanding of physiological synchrony more broadly, but to date have been considerably underutilized in the field.
Despite these limitations in the extant literature, several patterns have become clear. Regarding parasympathetic synchrony, when overall arousal levels are low (i.e., the stress response is not activated), synchrony during an interaction tends to be associated with better Caregiver×Child interaction quality and fewer caregiver and child psychopathological symptoms across all developmental periods examined. Parasympathetic regulation is associated with improved emotion regulation and less psychopathology (Musser et al., Reference Musser, Backs, Schmitt, Ablow, Measelle and Nigg2012). Thus, in normative-risk contexts, caregivers may be able to promote better emotion regulation and prevent psychopathology in their children via stronger parasympathetic synchrony. Importantly, however, synchrony was not associated with better interaction quality or child functioning for families at high socioeconomic risk, a history of maltreatment, or dyads with insecure attachment. It is possible that, in circumstances of elevated risk, synchrony may be an additional risk factor in the intergenerational transmission of the effects of stress and trauma (Thomas, Letourneau, Campbell, Giesbrecht, & Apron Study Team, Reference Thomas, Letourneau, Campbell and Giesbrecht2018).
A similar pattern was suggested regarding sympathetic synchrony, particularly in adolescence. Specifically, it seems that sympathetic synchrony was associated with better caregiver–child relationship quality, though studies are sparse and results were generally mixed. Thus, few conclusions can be drawn from the literature at this time. Much more research is needed examining sympathetic synchrony in diverse populations with varying levels of risk, during tasks that do and do not tend to elicit a stress response, as well as concurrent associations with interaction behavior and longitudinal associations with child and adolescent functioning.
Adrenocortical synchrony studies in toddlerhood tended to show a positive association with parental sensitivity and child functioning across multiple types of tasks. This may indicate a protective effect of adrenocortical synchrony during a developmental period marked by the final stages of attachment formation (Perry, Blair, & Sullivan, Reference Perry, Blair and Sullivan2017) and substantial self-regulatory development (Bernier, Beauchamp, Carlson, & Lalonde, Reference Bernier, Beauchamp, Carlson and Lalonde2015; Blair & Raver, Reference Blair and Raver2012). For example, adrenocortical synchrony may allow children to use their caregiver's physiological regulation as a scaffold for developing their own self-regulation skills. This may occur via subtle behavioral cues from both the caregiver and child that signal threat or distress (or the lack thereof) which, when detected, subsequently alter the other person's behavior and physiology accordingly (e.g., an infant eliciting comfort by displaying distress, a caregiver signaling threat or stress via facial cues). However, future studies must test this via real-time variation in the coordination of both adrenocortical activity and dyadic behavior simultaneously.
Despite the consistency of these studies, the majority will need to be replicated with more sophisticated multilevel modeling strategies (Bernard et al., Reference Bernard, Kashy, Levendosky, Bogat and Lonstein2017). Adrenocortical synchrony studies in infancy and late childhood/adolescence were inconsistent in their relation to caregiver and child characteristics, though patterns became clearer when only considering studies that used gold-standard stressor tasks (Strange Situation, Still-Face) and strong multilevel modelling techniques. Studies with the strongest methodological and statistical approaches tended to show positive associations between adrenocortical synchrony and parenting behaviors. These discrepancies should not be ignored, however, especially because positive findings in infancy and adolescence are based on normative risk populations (Atkinson et al., Reference Atkinson, Gonzalez, Kashy, Santo Basile, Masellis, Pereira and Levitan2013; Borelli et al., Reference Borelli, Shai, Smiley, Boparai, Goldstein, Rasmussen and Granger2019; Nofech-Mozes et al., Reference Nofech-Mozes, Jamieson, Gonzalez and Atkinson2019). Findings could vastly differ for higher-risk populations, as seen with parasympathetic synchrony, though this remains to be examined in future studies.
The small but quickly expanding body of research examining CNS synchrony is surprisingly consistent. All CNS synchrony studies included in this review were published in 2017 or later, and generally suggest that CNS synchrony, particularly in emotion-related brain regions, is associated with caregiver–child behavioral synchrony and child emotion regulation and awareness. The strength of study designs were variable and few studies examined caregiver or child correlates of CNS synchrony, but the results point to logical regions of interest for future synchrony studies. These regions, and how they are connected to autonomic and adrenocortical activity and synchrony, may be especially informative for how physiological synchrony manifests in caregiver–child dyads – a question still elusive to the field.
Limitations and future directions
Future studies should work to gain a more nuanced understanding of the factors associated with the presence and consequences of physiological synchrony and whether there are functional differences across physiological systems. Primarily, more research should be devoted to examining associations with positive/negative behavior during interactions as well as longitudinal child outcomes like executive functioning, socioemotional competence, mental health, and academic success. In addition, the majority of synchrony research has been conducted in toddlerhood/early childhood and using parasympathetic measures. Future studies should investigate synchrony across developmental stages and multiple physiological systems, particularly with longitudinal samples to investigate within-dyad changes, or stability, in synchrony over time. In order to tease apart patterns of synchrony in the context of chronic adversity (vs. normative risk contexts) and during an acute stress response (vs. calm, neutral, positive situations), future studies could employ a 2 × 2 design of samples with exposure to low versus high chronic stress/adversity and tasks that do or do not intend to elicit a stress response. This would allow us to more fully elucidate the circumstances under which physiological synchrony is a positive developmental process as a function of current context and long-term exposure to adversity. Studies should also consider the impact of gender (e.g., same-sex vs. opposite-sex dyads; Davis et al., Reference Davis, West, Bilms, Morelen and Suveg2018; Helm et al., Reference Helm, Miller, Kahle, Troxel and Hastings2018; Miller et al., Reference Miller, Vrtička, Cui, Shrestha, Hosseini, Baker and Reiss2019; Saxbe et al., 2015), socioeconomic status, race (e.g. how stress exposure impacts physiological attunement within different races), as well as cultural differences in levels of synchrony and/or the developmental correlates of synchrony.
It will be important to use multilevel modeling techniques that appropriately account for the dynamic, dyadic nature of the data, accurately separate between- from within-dyad effects, and otherwise reduce error associated with inappropriate statistical methods. Statistical methods that do not target within-dyad variation (i.e., regression) or measure dynamic changes over time (i.e., pre-/posttask difference scores) do not measure synchrony as presently defined and may result in inaccurate interpretation of the data (Bernard et al., Reference Bernard, Kashy, Levendosky, Bogat and Lonstein2017; Helm et al., Reference Helm, Miller, Kahle, Troxel and Hastings2018). Techniques like randomly re-pairing individuals into unfamiliar pairs (e.g., Saxbe et al., 2015), can be used to examine whether synchrony is truly an interpersonal process, rather than two individual processes. While every study cannot explicitly prove whether observed physiological synchrony is the result of an interpersonal process, efforts to rule out alternative explanations (e.g., two people processing the same stimulus in parallel, genetic inheritance of stress reactivity profiles) will be invaluable for understanding the conditions under which true, interpersonal physiological synchrony is observed. It is also possible to experimentally manipulate behavioral synchrony and examine resulting changes in physiological synchrony, as has been done with unfamiliar adult pairs (Danyluck & Page-Gould, Reference Danyluck and Page-Gould2018; Hove & Risen, Reference Hove and Risen2009; Hu et al., Reference Hu, Hu, Li, Pan and Cheng2017; Szymanski, Müller, Brick, von Oertzen, & Lindenberger, Reference Szymanski, Müller, Brick, von Oertzen and Lindenberger2017). In addition, while a discussion of diurnal physiological synchrony was outside the scope of this review, diurnal synchrony may be uniquely useful to understanding dyadic coregulation processes, especially for systems that operate over longer timescales and have systematic diurnal patterns (e.g., HPA activity; Pratt et al., Reference Pratt, Apter-Levi, Vakart, Kanat-Maymon, Zagoory-Sharon and Feldman2017). Some work on diurnal synchrony has already been done (e.g., Hibel, Mercado, & Valentino, Reference Hibel, Mercado and Valentino2019; Merwin, Smith, Kushner, Lemay Jr, & Dougherty, Reference Merwin, Smith, Kushner, Lemay and Dougherty2017; Williams et al., Reference Williams, Cash, Daup, Geronimi, Sephton and Woodruff-Borden2013), and future work should expand on these studies in addition to task-related synchrony.
Future research must also investigate the appropriate time-lag (if any) that should be used when measuring synchrony in different physiological systems, as well as which methods and analyses are appropriate for which research questions. Measures like salivary cortisol and sAA are very distal to the actual process researchers are intending to assess compared to, for example, measures of neural activity in regions directly upstream of the HPA axis or direct innervation of the heart from the SNS. This has implications for study design, sampling interval, data analysis, and interpretation. Knowledge of the biological and behavioral mechanisms underlying synchrony will provide critical guidance for addressing these methodological, statistical, and conceptual challenges. The working definition of physiological synchrony provided above (the dynamic, within-dyad coordination of physiological activity over time between two individuals that is directly tied to an interpersonal process) should be used to scaffold improvement of our methodological and statistical tools so that we can better operationalize and understand physiological synchrony using a common linguistic and analytic framework.
Conclusions
Overall, physiological synchrony research has developed rapidly, particularly over the past three to five years. Substantial knowledge has been gained, but there is much more that needs to be done to more fully understand the phenomenon of dyadic physiological synchrony. With continuous improvements in methodological, technological, and statistical approaches, our understanding of physiological synchrony can become much more nuanced. By understanding the predictors and outcomes associated with physiological synchrony across systems, we can more fully understand the mechanisms of adaptive and maladaptive caregiver–child relationships. This understanding will inform ways to improve caregiver–child relationship quality most effectively and help develop interventions aiming to reduce the intergenerational transmission of stress and trauma via the promotion of better self- and dyadic-regulation across development.
Acknowledgments
The author thanks Samantha Gardow, for providing the second independent review of study eligibility. Thanks also go to Drs. Megan Gunnar and Ann Masten for their input to improve the quality of this manuscript. Systematic review registration number: CRD42019123770.
Funding Statement
This research was supported by NIMH training grant T32 MH015755 to the sole author. The content is solely the responsibility of the author and does not represent the views of the National Institutes of Health.
Conflicts of Interest
None.




