


INTRODUCTION
Biogenic substrates—comprising whale and wood falls, sulphide-rich emissions, and methane seeps—are home to a taxonomically and phylogenetically diverse molluscan fauna that includes representatives from almost all the classes (Desbruyères et al., Reference Desbruyères, Sagonzac and Bright2006; Warén, Reference Warén, Bouchet, Le Guyader and Pascal2011). Typically, molluscs that are ecologically endemic to biogenic substrates graze the bacterial film or fungi, filter bacteria, or live symbiotically with bacteria (Wolff, Reference Wolff1979; Marshall, Reference Marshall1985, Reference Marshall1986, Reference Marshall1988; Warén & Bouchet, Reference Warén and Bouchet2001). Carnivorous and predatory molluscs are usually assumed to be mere ‘visitors’ in reducing environments, where they are thought to be attracted by the high biomass of these communities (for review see Sasaki et al., Reference Sasaki, Warén, Kano, Okutani, Fujikura and Kiel2010).
The vast majority of predatory snails in biogenic substrates belong to two neogastropod superfamilies—Conoidea and Buccinoidea. At least one genus of Raphitomidae (Conoidea), Phymorhynchus Dall, 1918, appears to be found preferentially in hydrothermal vent fields and seeps (Warén & Bouchet, Reference Warén and Bouchet1989, Reference Warén and Bouchet1993, Reference Warén and Bouchet2001; Beck, Reference Beck1996), and may be an ecological endemic. The other family of carnivorous gastropods well represented in such reducing environments is the family Buccinidae. The reasons why some buccinids show a preference for biogenic substrates are unclear. The anatomy has been examined (Harasewych & Kantor, 2002) by dissection for a single species from seeps, Buccinum thermophilum, and no morphological characteristic was standing out. In other groups of molluscs, and in particular in mussels from chemosynthetic environments, it has been proposed that several morphological or physiological features, such as the large size of the animal or the presence of endosymbionts, may explain their adaptation to extreme environments (Lallier, Reference Lallier2006). However, recent literature also revealed that a lack of taxonomic knowledge prevents any robust analysis of the evolution of these groups, and in particular the role of the potential adaptations to organic substrates (Génio et al., Reference Génio, Kiel, Cunha, Grahame and Little2012).
At least two genera of deep-water tropical Buccinidae—Eosipho Thiele, 1929, and Manaria E.A. Smith, 1906—are often found on sunken wood (Warén, Reference Warén, Bouchet, Le Guyader and Pascal2011), and several species were described either from sunken wood (Eosipho aldermenensis (Powell, 1971)), or from vents and seeps (Eosipho desbruyeresi Okutani & Ohta, 1993, E. auzendei Warén & Bouchet, Reference Warén and Bouchet2001 and E. canetae (Clench & Aguayo, 1944)). Calliloncha nankaiensis Okutani & Iwasaki, Reference Okutani and Iwasaki2003, was also described from abyssal seeps off Japan (Okutani & Iwasaki, Reference Okutani and Iwasaki2003) and a species of Kryptos has recently been described from methane seeps off Chile (Fraussen & Sellanes, Reference Fraussen and Sellanes2008). The recent finding of living specimens of the monotypic genus Enigmaticolus Fraussen, Reference Fraussen2008—originally described based on empty shells off north-west Madagascar together with bivalves typically associated with methane seeps or hydrothermal vents (Bathymodiolus, Vesicomya and Solemya), adds another taxon to the list of buccinids from reducing environments, and allows a critical evaluation of its relationships to Eosipho. A further two species of Neptunea Röding, 1798, three species of Bayerius Olsson 1971, and at least three species of Buccinum Linnaeus, 1758 are also found ‘near’ seeps or vents (Warén & Bouchet, Reference Warén and Bouchet2001; Harasewych & Kantor, 2002). Juvenile specimens (less than 15 mm) of Aulacofusus hiranoi (Shikama, 1962) and A. coerulescens (Kuroda & Habe in Habe, 1961) were found on pieces of sunken wood in bathyal depths off Japan (Hasegawa, Reference Hasegawa and Fujita2009).
The regular finding of buccinids on sunken wood in tropical deep water suggests that these are more than occasional ‘guests’. However, the phylogeny of the family Buccinidae is poorly understood, and we do not know whether the species occurring in vent, seeps and biogenic substrates form a monophyletic radiation, or whether they represent offshoots of various buccinid lineages. The purpose of the present paper is thus twofold:
-
(1) Describe the anatomical diversity of Buccinidae from sunken wood and seeps as a contribution towards a better understanding of the evolution of this particular fauna;
-
(2) provide, on the basis of molecular phylogenetic methods, the genus-level framework of the ‘Eosipho’ and ‘Manaria’-like species associated with sunken wood and other organic substrates, for a forthcoming species-level revision.
MATERIALS AND METHODS
Material
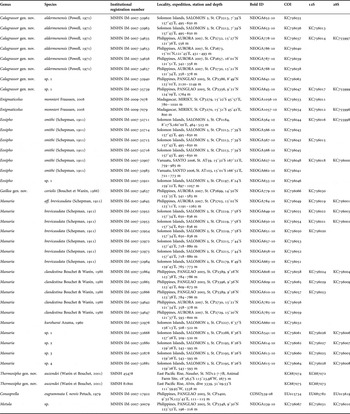
The material that forms the basis for this paper was collected during several deep-sea expeditions conducted by the Muséum National d'Histoire Naturelle (MNHN) and Institut de Recherche pour le Développement (IRD) in the Indo-West Pacific, notably: (1) Philippines. PANGLAO 2005 cruise on board MV ‘DA-BFAR’ associating the University of San Carlos, MNHN (co-Principal Investigator (co-PI) Philippe Bouchet) and the Philippines Bureau of Fisheries and Aquatic Research (BFAR; co-PI Ludivina Labe) (see Richer de Forges et al., 2009)); the ‘AURORA’ 2007 cruise also on board MV ‘DA-BFAR’ associating the National Museum of the Philippines (NMP, co-PI Marivene Manuel), MNHN (co-PI Philippe Bouchet) and ‘BFAR’; (2) Vanuatu. Material from the MNHN–IRD–PNI Santo 2006 expedition (see Bouchet et al., 2011); (3) Solomon Islands. SALOMON 2 2004 cruise (see Bouchet et al., 2008); and (4) Madagascar. MIRIKY 2009 cruise off north-west Madagascar as part of the ‘Our Planet Reviewed’ Mozambique–Madagascar expeditions. Specimens were preserved in 95 or 98% ethanol specifically for molecular analysis by clipping pieces of the head-foot from anaesthetized specimens, thus keeping the shell and the rest of the body intact for morphological analyses. All specimens used for molecular studies (Table 1) were individually catalogued and deposited in the MNHN, and the corresponding sequences are deposited in BOLD (Barcode of Life Datasystem) and GenBank. Two tissue samples of ‘Eosipho’ auzendei from the SMNH were provided by Anders Warén.
Table 1. List of specimens with collecting data and GenBank Accession numbers.
DNA extraction, amplification and sequencing
Genomic DNA was extracted from a piece of foot or from digestive gland tissue, using the 6100 Nucleic Acid Prepstation system (Applied Biosystems, St Aubin, France). Three genes were amplified: the ‘barcoding’ fragment of the Cytochrome Oxidase I (COI) mitochondrial gene, using universal primers LCO1490 and HCO2198 (Folmer et al., Reference Folmer, Black, Hoeh, Lutz and Vrijenhoek1994); a fragment of the mitochondrial rRNA 12S gene, using the primers 12S1 and 12S3 (Simon et al., Reference Simon, Franke, Martin, Hewitt, Johnson and Young1991); and a fragment of the nuclear rRNA 28S gene, using the primers C1 and D2 (Jovelin & Justine, Reference Jovelin and Justine2001). All PCR reactions were performed in 20 µl, containing 3 ng of DNA, 10× reaction buffer containing 15 mM MgCl2, 0.26 mM dNTP, 0.3 µM of each primer, 5% DMSO, 1 mg/ml BSA, and 1 unit of QBiotaq (MPBiomedicals, Illkirch Graffenstaden, France). Amplification consisted of an initial denaturation step at 95°C for 5 min, followed by 35 cycles of denaturation at 95°C for 1 min, annealing at 50°C for the COI gene, 54°C for the 16S and 56°C for the 28S genes for 30 s, followed by extension at 72°C for 30 s. The final extension was at 72°C for 5 min. The PCR products were purified and sequenced by the Eurofins sequencing facility. Both directions were sequenced to confirm accuracy of each sequence.
Phylogenetic analyses
Sequences were manually (COI) or automatically (28S and 12S) aligned using Muscle online (www.ebi.ac.uk/Tools/msa/muscle/). Phylogenetic analyses were performed using MrBayes (Huelsenbeck et al., Reference Huelsenbeck, Ronquist and Hall2001), running two parallel analyses, consisting each of eight Markov chains of 30,000,000 generations with a sampling frequency of one tree each ten thousand generations. The number of swaps was set to 5, and the chain temperature at 0.02. Parameters of the substitution model were estimated during the analysis (6 substitution categories, a gamma-distributed rate variation across sites approximated in four discrete categories and a proportion of invariable sites). A different model of substitution was applied for each codon position of the COI gene. A concatenated dataset of the three genes (COI, 12S and 28S) was built after checking for congruency of the three trees obtained with each gene; five independent partitions were defined (three codon positions of the COI gene, 12S, and 28S). Convergence of each analysis was evaluated using Tracer 1.4.1 (Rambaut & Drummond, Reference Rambaut and Drummond2007) to check that ESS values were all greater than 200 (default burning). Metula was identified as the most closely related genus to the group that included the Eosipho and Manaria group, based on an extended dataset of ~500 COI sequences of Buccinidae from the Indo-Pacific (unpublished data). We thus used a specimen of Metula sp. as closely related outgroup. A second outgroup, Conasprella eugrammata f. nereis Petuch, 1979 was used as distant outgroup to artificially root the tree (Table 1).
RESULTS
Molecular analysis
Separate analyses of CO1 (Figure 1), 12S (Figure 2) and 28S (Figure 3) genes revealed slightly different topologies, but most deep nodes were only weakly supported and no supported incongruences were found. The analysis of the concatenated dataset clearly helped in defining major lineages within the Manaria–Eosipho group and the relationships between them (Figure 4). A first clade was retrieved (posterior probabilities PP = 1) that includes all the Manaria species, of which four revealed by DNA sequences are potentially new species. This clade was also found with the 12S (PP = 0.40) and the 28S (PP = 0.99) genes. In all analyses the species traditionally placed in Eosipho were spread in four deeply divergent lineages. One of them includes E. smithi and an undescribed species as suggested by the COI tree. Because E. smithi is the type species of the genus, the name Eosipho remains attached to this clade. A second lineage corresponds to ‘Eosipho’ auzendei, a species from hydrothermal vents. It is sister-group to seep species, Enigmaticolus monnieri, but this relationship is not supported in any of the trees, and these species differ significantly both conchologically (‘Eosipho’ auzendei has more ovate shell with much shorter spire and lacks distinct spiral cords on the shell base and subsutural ramp) and anatomically (different morphology of proboscis retractors] and more poorly developed valve of Leiblein—see below). Thus, ‘Eosipho’ auzendei cannot be included in Enigmaticolus, and we propose the name Thermosipho gen. nov. A third lineage includes only one species, ‘Eosipho’ coriolis. This species is the sister-group to all the other species, excluding Enigmaticolus monnieri and Thermosipho auzendi. There is no generic name available for this branch and therefore a new genus is proposed, Gaillea gen. nov. The last lineage includes two species in our trees, ‘Eosipho’ aldermenensis and one undescribed species. This clade is a sister group to Manaria in the combined and 12S trees. In any case it cannot be included in Manaria on morphological grounds (shell and radula), and the new genus Calagrassor gen. nov. is proposed for this clade.

Fig. 1. Bayesian phylogenetic tree obtained with the COI gene. Posterior probabilities >0.80 are indicated for each node. Shells of sequenced specimens (except Thermosipho auzendei, for which the holotype is illustrated) are illustrated for comparison.

Fig. 2. Bayesian phylogenetic tree obtained with the 12S gene. Posterior probabilities >0.80 are indicated for each node. Shells of sequenced specimens (except Thermosipho auzendei, for which the holotype is illustrated) are illustrated for comparison.

Fig. 3. Bayesian phylogenetic tree obtained with the 28S gene. Posterior probabilities >0.80 are indicated for each node. Shells of sequenced specimens are illustrated for comparison.

Fig. 4. Bayesian phylogenetic tree obtained with the COI, 12S and 28S genes. Posterior probabilities >0.80 are indicated for each node. Shells of sequenced specimens (except Thermosipho auzendei, for which the holotype is illustrated) are illustrated for comparison.
Taxonomy
Class GASTROPODA Cuvier, 1795
Clade NEOGASTROPODA Wenz, 1938
Family BUCCINIDAE Rafinesque, 1815
Because our dataset is taxonomically focused, and because the suprageneric phylogeny of the Buccinoidea, including Buccinidae, is still far from resolved (Kantor et al., Reference Kantor, Puillandre, Rivasseau and Bouchet2012), we do not attribute any rank or name to the clade that includes the genera under consideration, pending a more robust and comprehensive phylogeny of the group. The species-level taxonomy will be discussed elsewhere (Fraussen & Stahlschmidt, in preparation).
Genus Enigmaticolus Fraussen, Reference Fraussen2008
Enigmaticolus Fraussen, Reference Fraussen2008: 75–77.
TYPE SPECIES
Enigmaticolus monnieri Fraussen, Reference Fraussen2008, by original designation.
DESCRIPTION
Shell large (Figure 5A–F), up to 100 mm, usually thick, solid. Colour brownish, occasionally purplish. Periostracum thick, dark brown, occasionally shiny or with metallic appearance, adherent. Shell broad fusiform, spire moderately high, siphonal canal short, broad, open. Protoconch and upper spire whorls decollate already in early, subadult age. Whorls convex, suture narrow canaliculate, subsutural ramp pronounced, rather broad, concave. Sculpture consisting of prominent, closely spaced, axial ribs spanning suture to suture on upper spire whorls, and gradually becoming obsolete on lower whorls. Spiral sculpture of inconspicuous cords; on siphonal canal, cords prominent, flattened, closely spaced, delimitated by narrow groves. Aperture large, slightly less than 1/2 of shell length, oval. Outer lip thick, slightly recurved outwards in adults, sometimes crenulated in abapical part. Callus narrow, but thick, overlaying parietal region and columella. Canal short, rather broad.

Fig. 5. Enigmaticolus monnieri Fraussen, Reference Fraussen2008: (A–B) holotype, MNHN 20464, SL 63.7 mm; (C) MNHN IM–2009–7078, SL 76.5 mm; (D–F) MNHN IM–2009–7079, SL 99.4 mm.
Operculum thick, corneous, usually dark brown; slightly smaller than aperture, semi-oval, nucleus terminal.
Radula (Figure 6E–F): rachidian teeth with anteriorly strongly arched base and with three closely spaced medium long, nearly equal, cusps. Two lateral cusps more raised over base, forming narrow concavity, possessing median cusp. Lateral teeth with two strong cusps, inner one shorter, but slightly broader than outer one.

Fig. 6. Soft body morphology of Enigmaticolus monnieri Fraussen, Reference Fraussen2008. (A–D) IM–2009–7078: (A) dorsal, (B) lateral, and (C) ventral views of the body; (D) mantle; (E–F) radula (IM–2009–7079). Abbreviations: cme, cut mantle edge; col.m, columellar muscle; ct, ctenidium; dg, digestive gland; eye, eye; hg, hypobranchial gland; ne, nephridium; op, operculum; os, osphradium; per, pericardium; re, rectum; s, siphon; sf, siphonal fold; st, stomach; tn, tentacle.
ANATOMY (ENIGMATICOLUS MONNIERI)
Two specimens of Enigmaticolus monnieri were dissected, from MIRIKY, Station CP3270, 15°31′S 45°42′E, 800 m, IM–2009–7078 (SL 76.5 mm) and from Station CP3279, 15°22′S 45°57′E, 780–1020 m, IM–2009–7079 (SL 99.4 mm).
Soft tissues (Figure 6A–D, based on specimen IM–2009–7078) comprise approximately 3 1/2 whorls. Mantle cavity spans just under 1/2 whorl, kidney 1/4 whorl, digestive gland and gonad just under 2 1/2; whorl. Columellar muscle short, broad, and thick, comprising one whorl, attached to shell at rear of mantle cavity. Foot large, strongly folded, narrowly rectangular (L/W ~2). Body colour yellowish tan, without pigmentation. Head large with long, medium thick tapering tentacles (Figure 6C, tn), without pronounced neck. Eyes large, on distinct rounded lobes at the base of tentacles, rather submerged into lobes. Nephridium brownish, with small and narrow nephridial gland (Figure 6B, ng), which has thin semi-transparent walls. Digestive gland greenish, lobes fused without visible border. Externally, ovary not distinguishable from digestive gland. Operculum thick, brownish, elongate–oval, with terminal abraded nucleus and numerous thickened growth lines.
Mantle cavity (Figure 6D): mantle thick, ctenidium and osphradium hardly visible through its wall. Mantle cavity of medium width (L/W ~1.2), mantle edge slightly thickened, non-serrated. Siphon medium long, free, muscular, extending substantially beyond mantle edge. Siphonal fold at siphon base well developed. Osphradium (Figure 6D, os) small (~0.35× mantle cavity length), narrow, greenish, bipectinate, with thin osphradial nerve. Ctenidium (Figure 6D, ct) large, very wide, spanning about 3/4 of mantle cavity length. Ctenidial lamellae simple triangular in shape, very low (width/height ratio ~5). Hypobranchial gland lacking distinct folds, partially overlaying rectum, covered by thick layer of mucus. Entire mantle covered by thick mucus layer.
Anterior foregut (Figure 7A–B) (based on the specimen IM–2009–7079). Proboscis in inverted position medium long, ~30 mm in length (~30% of SL, 60% of AL), occupying entire rhynchocoel, with slightly folded walls, unpigmented; slightly narrowing towards base of rhynchocoel. Mouth in shape of triangular opening, typical for Buccinidae. Proboscis retractor muscles numerous, powerful, arranged in two lateral symmetrical bundles, extending from mid-length of rhynchodaeum forward along entire anterior half of rhynchodaeum. Free parts of retractors short and attached to roof of anterior haemocoel, bending posteriorly at base and fusing with walls of rhynchodaeum, forming its outer layer. Rhynchodaeum medium thick, slightly thinner in anterior part, its internal wall medium folded.

Fig. 7. Digestive and reproductive systems of Enigmaticolus monnieri Fraussen, Reference Fraussen2008 (IM–2009–7079): (A) organs of anterior foregut, left view, rhynchodaeum (=proboscis sheath) dissected and proboscis exposed; (B) anterior oesophagus with valve of Leiblein, circumoesophageal nerve ring and part of right salivary gland, right view; (C) outer view of the stomach; (D) penis. Abbreviations: aoe, anterior oesophagus; crop, crop; dgd, ducts of the digestive gland; gL, gland of Leiblein; lsd, duct of left salivary gland; lsg, left salivary gland; nr, circumoesophageal nerve ring; poe, posterior oesophagus, pr, proboscis; prr, proboscis retractors; rsg, right salivary gland; vL, valve of Leiblein.
Odontophore large, 75% of proboscis length, occupying most of the proboscis volume, and compressing the anterior oesophagus dorso-ventrally, so that it becomes narrowly ovate. Odontophoral cartilages constituting about half of odontophore length (11 mm), fused in anterior 1/3 of their length and separated posteriorly.
Salivary glands loose, fused without visible border, with chinked surface, creamy white and brownish where they adjoin gland of Leiblein, and covering poorly pronounced valve of Leiblein and larger part of circumoesophageal nerve ring. Salivary ducts rather thin, straight, joining oesophagus shortly after leaving gland, and following between ventral surface of oesophagus and rhynchodaeum, completely concealed by former, hardly discernible in posterior part of inverted proboscis among numerous thin muscle fibres.
Gland of Leiblein large, grey-brown, bulky, broadly tubular and folded anteriorly, where it is more intensively coloured. Folds of gland in anterior part encompassing large right salivary gland. Posterior part of gland laterally compressed and tightly attached to posterior oesophagus by connective tissue fibres. Partitions of gland better seen in posterior part. In anteriormost part, gland narrowing to form short distinct duct, opening into mid-oesophagus significantly posterior to nerve ring. (position marked by arrow on Figure 7A)
Anterior oesophagus rather muscular and attached to rhynchodaeum by numerous muscular fibres, oesophagus completely covering and hiding salivary ducts. Two thick proboscis nerves follow on both sides of thick blood vessel between dorsal side of oesophagus and rhynchodaeum. Valve of Leiblein pyriform, slightly broader than oesophagus, mostly covered by right salivary gland that is tightly attached to valve. Glandular pad seen through walls of valve. Mid-oesophagus rather long, but thin walled. Border between mid- and posterior oesophagus marked by distinct but short duct of gland of Leiblein. Posterior oesophagus flaccid, laterally compressed and forming distinct outgrowth in its posterior part.
Stomach (Figure 7C) not large, spanning ~1/3 whorl, U–shaped, posterior mixing area very short, hardly discernible. Posterior oesophagus and stomach inside lined with orange-brownish epithelium. Posterior oesophagus thick and broad. Posterior duct of digestive gland large, situated at entrance of oesophagus into the stomach. Anterior duct shifted anteriorly, marking transition to intestine. Due to preservation conditions internal epithelium slogged off walls of stomach and it was not possible to study its anatomy in details. No stomach content was found.
Specimen IM–2009–7079 was an adult male, penis long, club-shaped on top with very tiny conical seminal papilla (Figure 7D).
INCLUDED SPECIES
Enigmaticolus monnieri Fraussen Reference Fraussen2008. Type locality: Mozambique Channel, Madagascar, off Tulear, trawled by commercial fishery vessels (shrimpers), 800 m deep.
There are still several undescribed species in the East China Sea (China) and the western and south-western Pacific (Japan, Philippines, New Caledonia, Fiji and Lau Basin), descriptions of which are now in preparation (Fraussen & Stahlschmidt, in preparation).
REMARKS
Enigmaticolus is most similar to Thermosipho gen. nov. and differs by a longer and more pronounced siphonal canal, the presence of spiral sculpture, and a distinctly set-off subsutural ramp, the presence of axial folds in the subsutural area in subadult and adult whorls, and a larger operculum.
In Madagascar, E. monnieri was trawled alive in hauls on the sedimentary cone of the River Betsibuka, with an associated fauna including bathymodiolines, vesicomyids and solemyids, suggesting the presence of seeps.
Genus Thermosipho gen. nov.
(Figure 1(2))
TYPE SPECIES
Eosipho auzendei Warén & Bouchet, Reference Warén and Bouchet2001.
DESCRIPTION
Shell medium to large, up to 70 mm, thin to medium thick, solid. Colour brownish to dark brown. Periostracum thick, orange to dark brown, forming dense axial lamellae, but usually smooth due to abrasion except near suture. Suture deeply impressed, not canaliculate. Shape broadly fusiform, spire moderately short. Protoconch and upper spire whorls decollate already at early subadult age. Whorls weakly convex, without subsutural ramp. Axial folds present only on earliest whorls and absent in most adults and subadults. Spiral sculpture of very weak inconspicuous cords or absent. Aperture large, oval, more than half of shell length (up to 0.7 of SL in juveniles). Outer lip thin to rather thick, slightly reflected outwards in adults. Callus narrow, overlaying parietal region and columella. Canal very short, broad.
Operculum corneous, usually light to dark brown, oval, occupying about half of aperture length, nucleus terminal.
Radula (illustrated by Warén & Bouchet, Reference Warén and Bouchet2001: figure 39A for T. auzendei and by Okutani & Ohta, 1993: figure 5 for T. desbruyeresi): rachidian teeth with base strongly arched anteriorly and with three closely spaced, medium to long, nearly equal, cusps. Two lateral cusps more raised over base, forming narrow concavity, possessing median cusp. Lateral teeth with two strong cusps, inner one shorter, stronger curved and slightly broader than outer one.
ANATOMY (THERMOSIPHO AUZENDEI)
The dissected earlier paratype 2 of Thermosipho auzendei was reexamined. Mantle cavity morphology very similar to Enigmaticolus monnieri, differing in slightly larger ctenidium.
Anterior foregut similar to E. monnieri. Instead of series of lateral proboscis retractors subequal in size, pair of very large more posterior retractors, branching near tip, attached to roof of body haemocoel. Several smaller retractors attached to rhynchodaeum anterior to major retractors. Gland of Leiblein even larger than in E. monieri, very long, broadly tubular and folded several times, forming very voluminous mass which spans on ventral side of rhynchodaeum anteriorly. In anteriormost part gland becomes narrower and opens into mid-oesophagus by narrow duct significantly posterior to nerve ring. Valve of Leiblein very poorly pronounced, hardly discernible from oesophagus.
REMARKS
Thermosipho gen. nov. is most similar to Enigmaticolus. The two genera cluster together in the molecular trees, although without significant support (PP only 0.81 in the three genes tree). They have similar radulae and anatomy (see below). Nevertheless, Thermosipho differs from Enigmaticolus in its more ovate shell with higher last whorl, shorter siphonal canal, absence of distinct spiral cords on the shell base and absence of subsutural ramp.
INCLUDED SPECIES
Thermosipho auzendei (Warén & Bouchet, Reference Warén and Bouchet2001) comb. nov. Type locality: East Pacific Rise, Rehu site, 17°24.85′S, 113°12.15′W, 2578 m, vent.
Thermosipho desbruyeresi (Okutani & Ohta, 1993) comb. nov. (original combination: Eosipho desbruyeresi). Type locality: BIOLAU 89 dive 06, 23°13′S 176°38′W, 1750 m, vent). Synonym: Eosipho desbruyeresi nipponensis Okutani & Fujiwara, 2000. Type locality: Okinawa Trough, Iheya Ridge, North Knoll, Shinkai 2000 Dive 1112, 27°47.180′N 126°54.149′E, 1049 m, vent.
ETYMOLOGY
Thermosipho gen. nov. is named after thermae (Latin), the hot bath of the Romans, in reference to the habitat of the type species associated with hydrothermal vents (black smokers); rendered as thermo—for euphonic reasons; in combination with—sipho, a name that is in use for many cold-water fusiform buccinids.
Genus Eosipho Thiele, 1929
(Figure 1(4), (5))
Sipho (Eosipho) Thiele, 1929: 307.
TYPE SPECIES
Chrysodomus (Sipho) smithi Schepman, 1911, by original designation.
DESCRIPTION
Shell large, up to 75 mm, rather thick in adults, occasionally thin but solid, white. Periostracum thin, olive, often peeling and retains between spiral cords. Shape semi-ovate, rather oviform, broad with rather high spire and short but distinct siphonal canal. Protoconch eroded in all studied specimens, consisting of 1 3/4–2 whorls. Teleoconch whorls poorly convex, without pronounced shoulder. Teleoconch with dominant spiral sculpture. Spiral cords flattened, on later whorls bilirate. Interspaces variable, narrow to broad, usually fine groove. Axial sculpture weak, on upper spire whorls only, or absent by erosion; lower spire whorls and body whorl without traces of axial sculpture. Aperture large, usually about 1/2 of shell length, occasionally larger. Outer lip thick in adults, slightly curling outwards with a row of moderately broad, flattened knobs along edge.
Radula (illustrated by Bouchet & Warén, Reference Bouchet and Warén1986: figure 15): rachidian teeth with rectangular base, shallowly arched anteriorly and with three closely spaced medium long cusps, central slightly longer, emanating from posterior edge. Lateral teeth with two strong cusps, inner narrower and slightly shorter than outer.
INCLUDED SPECIES
Eosipho smithi (Schepman, 1911) (Type locality: Indonesia, north of Pulau Talisei, Celebes Sea, Siboga Station 122, 01°58′N 125°00′E, 1165–1264 m) has a broad Indo-West Pacific distribution from East Africa (Mozambique Channel) in the west, along Indonesia and Philippines to southern Japan, Solomons and Fiji in the east.
According to COI tree, Eosipho includes at least one more unnamed species (Eosipho sp. 1, IM-2007-32921) from lower bathyal depths in the Solomon Islands. The sequenced specimen is immature (shell length 24.7 mm) and axial ribs are present on the last whorl.
Genus Gaillea gen. nov.
(Figure 1(3))
TYPE SPECIES
Eosipho coriolis Bouchet & Warén, Reference Bouchet and Warén1986.
DESCRIPTION
Shell of medium to large size, up to 70 mm, thick and solid. Colour white. Periostracum thin, adhering, light olive, forming irregularly spaced axial lamellae and occasionally short bristles. Shape broad, fusiform, to semi-oval with attenuated, but distinct, siphonal canal. Protoconch usually eroded in both adult and juvenile species; pointed, higher than broad. Teleoconch whorls weakly convex with angulated shoulder, subsutural slope straight. Sculpture with some reticulate pattern; spiral sculpture fine, especially on abapical spire whorls and body whorl, variable, usually consisting of spiral cords separated by deep and narrow interspaces or grooves; along shoulder, a single, more pronounced, primary spiral cord accentuating shoulder angulation. Axial sculpture weak, consisting of low axial ribs along on adapical part of whorls, usually slightly broader, but weaker, on body whorl. Aperture oval to round. Columella gently curved; transition to siphonal canal usually strongly curved. Callus thin, overlaying spiral sculpture and outer shell layer of parietal region, glossy, smooth. Outer lip usually thin, occasionally thick; edge sharp.
Radula (illustrated by Bouchet & Warén, Reference Bouchet and Warén1986: figures. 14, G. engonia; 15, G. coriolis): rachidian teeth with rectangular base, shallowly arched anteriorly, and with three broadly spaced, medium long, cusps, central equal in length to, or slightly longer than, lateral ones, emanating from posterior edge. Lateral teeth with two strong cusps, inner narrower and slightly shorter than outer.
REMARKS
Species belonging to Gaillea gen. nov. are most similar to Eosipho and Calagrassor gen. nov. They differ from Eosipho in slightly carinated whorl profile; finer spiral sculpture consisting of broader spaced and narrower cords alternating in size; presence of finer spiral cords on the subsutural slope and better pronounced axial ribs which are present on the last whorl but absent on the base.
From Calagrassor gen. nov., Gaillea differs by its broader shell, with usually constricted base, thinner spiral cords with broader interspaces, presence of a stronger spiral cord along the shoulder accentuating the shoulder angulation, sharper apex with pointed protoconch, and slightly larger adult size.
INCLUDED SPECIES
Gaillea coriolis (Bouchet & Warén, Reference Bouchet and Warén1986) comb. nov. Type locality: Philippines, northern Mindoro, south-eastern Ambil Island, MUSORSTOM 2, Station 81, 13°34′N 120°31′E, 856–884 m.
Gaillea engonia (Bouchet & Warén, Reference Bouchet and Warén1986) comb. nov. (original combination: Eosipho engonia). Type locality: northern Mozambique Channel, Vauban Station 139, 13°50′S 47°37′E, 850–1125 m.
On the basis of shell shape with angulated shoulder and shell sculpture, Gaillea tosaensis (Okutani & Iwahori, 1992) comb. nov. (original combination: Eosipho tosaensis) is also probably congeneric. Type locality: Tosa Bay, Japan, 780 m.
Finally, Gaillea canetae (Clench & Aguayo, 1944) comb. nov. (original combination: Buccinum canetae) appears to belong to Gaillea based on the shell and radula (illustrated by Warén & Bouchet, Reference Warén and Bouchet2001, figure 39d). Type locality: off La Havana, Cuba, 23°12′30″N 82°12′W, 240–300 fathoms.
ETYMOLOGY
The new genus is named after Françoise Gaill, a long-time companion of P. Bouchet during deep-sea expeditions with CNEXO, and later IFREMER, and in recognition for her interests in chemosynthetic environments and sunken wood.
Genus Calagrassor gen. nov.
(Figure 1(13) (14))
TYPE SPECIES
Cantharus aldermenensis Powell, 1971.
DIAGNOSIS
Shell small to medium in size (adult length from 9 to 45 mm), solid, usually rather thin. Colour white. Periostracum velvety, forming fine axial lamellae or bristles; occasionally smooth, often well adherent to shell. Shape oval with short siphonal canal. Protoconch usually eroded, juveniles with a moderately small, naticoid protoconch, width exceeds height, consisting of 1 1/4 to 1 1/2 smooth, convex whorls. Teleoconch whorls evenly weakly convex. Spiral sculpture dominant, usually of broad, flattened spiral cords separated by deep interspaces, narrower than or equal to cords width. Axial sculpture usually weak, pronounced on adapical spire whorls, usually absent on body whorl, ranging from broad, flattened ribs to narrow and moderately sharp ones. Aperture oval, adapically usually slightly pinched. Columella gently curved. Callus thin, narrow, formed by dissolution of sculpture and outer shell layer of preceding whorl, glossy, smooth. Outer lip usually thin, occasionally thick, edge sharp, occasionally with internal lirae or knobs.
Radula (illustrated by Bouchet & Warén, Reference Bouchet and Warén1986, figure 13): rachidian teeth with rectangular base, shallowly arched anteriorly, with three closely spaced, long cusps, central much longer than lateral ones, emanating from posterior edge. Lateral teeth with two strong, long cusps, inner narrower and much shorter than outer. In C. zephyrus, there is an additional very thin and short intermediate cusp on lateral teeth.
ANATOMY (CALAGRASSOR ALDERMENENSIS)
The anatomy of a specimen collected off Madagascar (MIRIKY, Station CP 3290, 14°47'S 47°26′E, 409–425 m; shell length 19.1 mm) was examined.
Soft tissues (Figure 8C–D) comprise just under 3 whorls. Mantle cavity spans just under 1/2 whorl, kidney 1/4 whorl, digestive gland and gonad just under 1 1/2 whorl. Columellar muscle extending over 1 1/2 whorl, broad and thick, attached to shell posteriorly to rear of mantle cavity, subdivided into three bands, lower one longest. Foot large, contracted, sole broadly oval (L/W ≈1.4). Body colour yellowish tan, unpigmented. Head medium sized, with long, medium thick, cylindrical tentacles, rounded at tip (Figure 8D, tn), without pronounced neck. Eyes large, on distinct rounded lobes at base of tentacles. Nephridium light brownish, with few ramifying trabecules seen through wall, with small and narrow nephridial gland (Figure 9C, ng). Digestive gland light brownish, clearly bilobed; lobes separated by testis. Posterior lobe narrow, spanning about 1/3 whorl, overlaid by testis, which occupies most of visceral mass. Operculum thin, yellow, transparent, elongate-oval, with abraded terminal nucleus and numerous thin growth lines.

Fig. 8. Calagrassor aldermenensis (Powell, 1971): (A–B) live snails on the sunken log, MIRIKY, Station CP 3290; (C) ventral and (D) dorsal views of the body; (E) mantle. Abbreviations: see Figure 6.

Fig. 9. Digestive system of Calagrassor aldermenensis (Powell, 1971): (A) organs of anterior foregut, left view, rhynchodaeum (=proboscis sheath) dissected and proboscis exposed; (B) anterior oesophagus with valve of Leiblein, circumoesophageal nerve ring, right salivary gland and part of gland of Leiblein, right view; (C) visceral mass with the stomach. Abbreviations: adg, anterior digestive gland; anrh, anterior thick portion of rhynchodaeum; cae, caecum of posterior oesophagus; ht, heart; pdg, posterior digestive gland; psrh, posterior thin portion of rhynchodaeum; rm, rhynchodaeum retractor muscle; ts, testis. Other abbreviations see Figure 7.
Mantle cavity (Figure 8E). Mantle thin, ctenidium and osphradium clearly seen through its wall. Mantle cavity nearly square (L/W ~1.1), mantle edge slightly thickened, serrated in correspondence with spiral cords. Siphon not long, free, muscular, only slightly extending beyond mantle edge. Siphonal fold well developed at base of siphon. Osphradium large (~0.5× mantle cavity length), as broad as ctenidium, brownish, bipectinate, with thick osphradial nerve. Ctenidium large, medium wide, spanning about 3/4 of mantle cavity length. Ctenidial lamellae simple, right-angled triangular in shape, very low. Hypobranchial gland lacking distinct folds, partially overlaying rectum, covered by thick layer of mucus. Entire mantle cavity covered with thick mucus layer. Rectum rather short, slightly over half mantle length, terminating in anus without distinct papilla.
Anterior foregut (Figure 9A, B). Proboscis in inverted position folded within the rhynchocoel, long, ~8.5 mm in length (~45% of SL, equals AL), occupying only posterior part of rhynchocoel, with folded walls, unpigmented, of approximately even diameter along its length. Slit-shaped mouth. Rhynchodaeum thick-walled and very narrow in anterior half, and thin-walled, transparent and spacious posteriorly, where folded proboscis contained. (It should be noted that, in several dissected specimens from Vanuatu, the proboscis was not folded within the rhynchodaeum.) Two pairs of proboscis retractor muscles, large and thick, attached approximately to mid-length of rhynchodaeum. Thin flat band-like transparent muscle originating at base of posterior pair of major retractors (Figure 9A, rm), attached to posterior part of the rhynchodaeum, probably folding and contracting rhynchodaeum.
Odontophore minute, about 1.5 mm long (~20% of proboscis length).
Salivary glands creamy white, with slightly chinked surface, fused without visible border, adjoining gland of Leiblein, oesophagus and valve of Leiblein and larger part of circumoesophageal nerve ring. Salivary ducts rather thin, nearly straight, and joining oesophagus in front of valve of Leiblein shortly after leaving it, and then following tightly attached to oesophagus by connective tissue, not seen exteriorly. Gland of Leiblein large, brown, bulky lobular, and voluminous anteriorly, where it is more intensively coloured, encompassing in anterior part most of left salivary gland. Posterior part of gland laterally compressed, with semi-transparent walls and mostly lacking glandular tissue.
Anterior oesophagus rather muscular, narrow, and attached to rhynchodaeum by numerous muscular fibres. Thick proboscis nerve follows between dorsal side of oesophagus and rhynchodaeum. Valve of Leiblein well pronounced, approximately twice broader than anterior oesophagus, long and nearly cylindrical, with mucous pad well seen through its walls. Mid-oesophagus very short, bordered posteriorly by indistinct short duct of gland of Leiblein. Posterior oesophagus flaccid, laterally compressed and forming distinct outgrowth in its posterior part.
Stomach (Figure 9C) not large, spanning ~1/3 whorl, U–shaped, posterior mixing area not discernible. Posterior oesophagus broad, of same diameter as stomach. Posterior duct of digestive gland small, situated at entrance of oesophagus into the stomach. Anterior duct slightly shifted anteriorly. Intestine after leaving stomach becoming much narrower as it follows under the kidney. Due to preservation, internal epithelium slogged off the walls of the stomach and impossible to study in detail.
REMARKS
Species of Calagrassor differ from Gaillea in being more slender, with evenly rounded whorls, without angulation. In general shell shape and sculpture, they are most similar to Eosipho and differ in having a smaller protoconch formed by convex whorls, more convex adapical teleoconch whorls, non subdivided spiral cords, and a simple outer lip.
INCLUDED SPECIES
Calagrassor aldermenensis (Powell, 1971) comb. nov. Type locality: New Zealand, Coromandel, east of Aldermen Islands, 36°58′S 176°06′E, 366–475 m.
Calagrassor poppei (Fraussen, 2001) comb. nov. (original combination: Eosipho poppei). Type locality: Philippines, Balicasag Island, 140–400 m.
Calagrassor tashiensis (Lee & Lan, 2002) comb. nov. (original combination: Colus (Aulacofusus) tashiensis Lee & Lan, 2002). Type locality: north-east Taiwan, around Kue-Shun Islands, 400–500 m.
Calagrassor zephyrus (Fraussen, Sellanes & Stahlschmidt, 2012) comb. nov. (original combination: Eosipho zephyrus Fraussen, Sellanes & Stahlschmidt, 2012). Type locality: southern Chile, off Chiloé, about 42′S, in 500 m.
A further deep-water (1764–2150 m) species from the Philippines (Calagrassor sp. 1 in our analysis) will be named elsewhere (Fraussen & Stahlschmidt, in preparation).
ETYMOLOGY
From the Latin cala, a piece of wood; and grassor, vagabond. Used as a noun in apposition.
REMARKS
Calagrassor aldermenensis is exclusively found on sunken logs, sometimes in very deep holes, that are exposed when the wood is cut (Figure 8A, B).
Genus Manaria E.A. Smith, 1906
(Figure 1 (6–12))
Manaria E.A. Smith 1906: 167.
TYPE SPECIES
Manaria thurstoni E.A. Smith, 1906, by original designation.
DESCRIPTION
Shell medium large to large (adult length from 14 to 80 mm), usually thick and solid. Colour chalky white. Shape fusiform with moderately long siphonal canal. Periostracum velvety, forming with fine axial lamellae or bristles, occasionally smooth, well adherent to shell. Protoconch (often eroded even in young specimens) moderately small, of 1 1/4 to 1 1/2 smooth, convex whorls. Teleoconch whorls convex, occasionally with weak, concave subsutural ramp. Suture deep, rarely shallowly canaliculate. Spiral sculpture dominant, variable, usually of well pronounced cords separated by deep interspaces, up to twice as broad as spiral cords but occasionally restricted to sharp grooves. Axial sculpture variable, usually dominant on adapical whorls, weaker or absent on last whorl, ranging from broad, flattened ribs to narrow and sharp ones. Aperture narrow to oval, usually slightly pinched adapically. Columella gently curved, smooth or with one or more weak knobs; callus thin, formed by dissolution of sculpture and outer shell layer of preceding whorl, glossy, smooth or with one or more weak columellar knobs. Outer lip usually thin, occasionally thick, edge sharp, with internal lirae or knobs.
Operculum corneous, thick, dark to pale brown, semi-ovate, nucleus terminal.
Radula (several species, illustrated by Bouchet & Warén, Reference Bouchet and Warén1986: figures 19–24): rachidian teeth with narrow rectangular base (length may exceed width), shallowly to deeply arched anteriorly and with three closely spaced, medium to long, cusps, occupying most of posterior edge of basal plate. Central cusp very slightly longer than lateral ones. Lateral teeth with two strong short cusps, inner slightly shorter than outer; occasionally there can be a vestigial intermediate cusp.
REMARKS
Manaria differs from other genera under consideration in its generally narrower, fusiform shell, with higher spire and longer siphonal canal, and in well developed axial ribs on all whorls.
SPECIES INCLUDED
Manaria thurstoni E.A. Smith, 1906. Type locality: India, Gulf of Manar, Investigator Station 333, 06°31′N 19°39′E, 730 m.
Manaria brevicaudata (Schepman, 1911) (original combination: Fusus brevicaudatus). Type locality: Flores Sea, Siboga, Station 45, 07°24′S 118°04′E, 470–475 m.
Manaria kuroharai Azuma, 1960. Type locality: Japan, Tosa Bay, Shikoku Island, 180 m.
Manaria lirata Kuroda & Habe in Habe, 1961. Type locality: Japan, Ashizuri-misaki, Kochi Prefecture, Shikoku, 100 m.
Manaria clandestina Bouchet & Warén, Reference Bouchet and Warén1986. Type locality: Philippines, Sibuyan Sea,, Musorstom 2 Station 36, 13°31′N 121°24′E, 570–595 m.
Manaria makassarensis Bouchet & Warén, Reference Bouchet and Warén1986. Type locality: Makassar Strait, Corindon 2, Station 214, 00°31′N 117°50′E, 595 m.
Manaria formosa Bouchet & Warén, Reference Bouchet and Warén1986. Type locality: Mozambique Channel, Vauban, Station 39, 12°46′S 48°10′E, 495–500 m.
Manaria thorybopus (Bouchet & Warén, Reference Bouchet and Warén1986) comb. nov. (original combination: Eosipho thorybopus). Type locality: Mozambique Channel, Vauban, Station 114, 22°15′S 43°04′E, 470–475 m.
Manaria chinoi Fraussen, Reference Fraussen and Hadorn2005. Type locality: Philippines, Mindanao, off Davao, 160 m, and many still unnamed species from the Indo-West Pacific (Fraussen & Stahlschmidt, in preparation).
DISCUSSION
Position of buccinids from sunken wood within Buccinoidea
The dataset analysed has limitations due to the number of taxa included. A number of species already described were not available for molecular analyses, and many more species are still unnamed judging from traditional shell characters. Furthermore, the sequenced material is biased towards the Indo-West Pacific, and none of the Atlantic species attributed to the group (Fraussen & Hadorn, Reference Fraussen and Hadorn2005) was included in the molecular phylogeny. Nevertheless, a number of conclusions can be significantly drawn. The species included in the analysis were previously classified in Manaria and Eosipho, two genera rather loosely defined based on shell characters, but they actually correspond to six deep molecular lineages that we rank as genera. All share rather a similar radular morphology, with bicuspid lateral teeth and a tricuspid rachidian. Although bicuspid lateral teeth are found in other, obviously non related, genera of Buccinidae (e.g. in Phos, Cominella, and several Antarctic genera)—thus suggesting convergent evolution—this similarity of radula among several conchologically dissimilar, but phylogenetically related, genera is remarkable. This suggests that, at least in some Buccinoidea the radula is more conservative in terms of morphological variation than the shell.
The molecular dataset recovers as monophyletic Buccinidae that are known (Thermosipho, Enigmaticolus, some Calagrassor) or suspected (Manaria) to inhabit biogenic substrates. Bayerius and Calliloncha nankaiensis (also known to inhabit biogenic substrates; for review see Warén & Bouchet, Reference Warén and Bouchet2001; Sasaki et al., Reference Sasaki, Okutani and Fujikura2005) may or may not belong to this group. However, several species of Neptunea and Buccinum, also recorded from biogenic substrates certainly do not fall in this clade. This is confirmed by the differences in radulae anatomy (strictly bicuspid lateral teeth in the described genera and tri- or multicuspid in Buccinum and Neptunea, the differences significant enough to attribute genera to different tribes in Buccinidae), as well as unpublished results based on COI sequences. There have thus been several colonization events that brought Buccinidae to live on biogenic substrates, but a single one appears to have diversified into at least six genera which are restricted to such habitats.
Although the clade that includes the genera under consideration is morphologically and, presumably (in the absence of a larger phylogeny of the Buccinidae), molecularly well defined, we abstain from giving it a formal rank and establishing a name for it. Unpublished, and still fragmentary, molecular data on the Buccinidae reveal a complex phylogenetic pattern, in a number of ways contradicting the classification of Buccinoidea adopted by Bouchet & Rocroi (Reference Bouchet and Rocroi2005). We therefore consider it premature to give a name and a rank to the clade.
Anatomy and morphological adaptations
SHELL
One of the remarkable characters of the shells of Thermosipho and Enigmaticolus is the very thick periostracum, especially in the former genus, while the inorganic matrix of the shell is relatively thin for such large sizes. In adults, periostracum is usually smooth due to abrasion, but in the youngest specimens it bears axial lamellae. In species of both genera, protoconch and upper teleoconch whorls are decollated, probably as a result of the dissolution of the shell. A thick periostracum could be a protection of the shells in aggressive environmental conditions in the vicinity of vents and seeps. In other genera, the periostracum is also prominent, but not so thick, and is in fact more similar to other deep water non seep-vent buccinids, suggesting the lack of necessity of protecting the shell from dissolution. Also the protoconchs are often eroded, but upper teleoconch whorls are usually intact.
DIGESTIVE SYSTEM AND FEEDING
As was mentioned in the introduction, the anatomy of a single buccinoid species from biogenic substrates has been examined before this study: Buccinum thermophilum Harasewych & Kantor, 2002. That species was found to be an active predator and opportunistic scavenger with a broad diet, including vestimentiferans, pycnogonids and polychaetes (Martell et al., Reference Martell, Tunnicliffe and Macdonald2002). Its anatomy was typical for Buccinum species (Harasewych & Kantor, 2002), and no anatomical characters were linked to its inhabiting vents environment.
Likewise, the anatomy of Enigmaticolus monnieri and Calagrassor aldermenensis, studied here, is typical for Buccinoidea. Neither their external morphology, nor their digestive system anatomy, suggest any specific adaptation. In Calagrassor aldermenensis, the proboscis is very long and able to coil within the rhynchodaeum, which may be an adaptation for capturing prey hiding in narrow holes and crevices. However, a similar proboscis is found in other Buccinidae from the ambient deep sea, such as in some Aulacofusus and Colus species, as well as in Ancistrolepis (Kantor, Reference Kantor1988; Kosyan & Kantor, Reference Kosyan and Kantor2009). Nothing so far is known about the diet of any of these species. We dissected more than 25 specimens of Calagrassor aldermenensis, but stomach and gut were empty in all of them. One specimen of Enigmaticolus monnieri, when preserved, regurgitated a mass of unidentifiable, partially digested, soft tissues, which may be the tentacles of large sedentary polychaetes; this remains entirely speculative, though, and our attempts to sequence the gut content for the COI gene failed.
So, judging from the foregut anatomy we may suggest that Calagrassor and Enigmaticolus are predators or scavengers, in that respect not differing from other Buccinidae. Apparently, the buccinoid foregut anatomy allows preying on a wide variety of invertebrates without any special morphological adaptations.
Diversification of Buccinidae on biogenic substrates
Three major biogenic substrates (hydrothermal vents, methane seeps and sunken wood) are colonized by Buccinidae. It is rather obvious that Thermosipho and Enigmaticolus are really specialized on vents and seeps, since they are collected in numbers only there and not elsewhere. We have much fewer data about actual occurrences of Buccinidae on sunken wood, and hauling a trawl on the bottom is not the best approach to documenting labile associations. In fact, of all the species analysed herein, only Calagrassor aldermenensis is known to inhabit exclusively sunken logs. It obviously prefers to hide deep inside holes, which would explain why specimens are rarely—if ever—found loose in the bottom residue. By contrast, many other species are often found in the same hauls as sunken wood, but not attached to the logs themselves. For these, there is only indirect evidence of association with wood: the whelks can either have been washed off from the logs during hauling or be free dwellers, and their co-occurrence with sunken wood is fortuitous. Also it is possible that whelks spend only part of their life cycle on wood, being free roamers during most of their life. This is known in Aulacofusus hiranoi and A. coerulescens, the juveniles of which live on sunken wood. It is also interesting to record that traps containing wood and protected by a netting with 3 mm mesh, deployed in deep water off Santo, Vanuatu (Samadi et al., Reference Samadi, Corbari, Lorion, Hourdez, Haga, Dupont, Boisselier and Richer De Forges2010), were recovered with Manaria specimens, as well as Calagrassor aldermenensis (unpublished data), suggesting that the association of Manaria with wood is not just fortuitous.
Several other groups of molluscs (e.g. chitons, Pectinodonta, cocculinids, mytilids, Pholadoidea) also diversified on biogenic substrates, but for many of them we lack the comprehensive molecular phylogenies that are necessary to place the wood-associated taxa in perspective. In mytilids, wood is certainly the ancestral state, followed by several colonization events to other biogenic substrates—whale carcasses, cold seeps or hydrothermal vents (Lorion et al., Reference Lorion, Buge, Cruaud and Samadi2010). The pattern observed in Buccinidae is much cruder, but it is nevertheless noteworthy that species exclusively associated with vents and seeps and species found on or with sunken wood are closely related, although this relationships needs to be thoroughly tested in a larger phylogenetic context, also supporting a non-fortuitous association with sunken wood.
ACKNOWLEDGeMENTS
The work was conducted in Paris by Y.I.K. under the MNHN visiting curatorship programme. This author expresses his thanks to Virginie Héros, Barbara Buge, Philippe Maestrati and Pierre Lozouet for assistance during his stays in Paris. Anders Warén provided samples of tissues of Thermosipho auzendei. The authors express their thanks to anonymous referees for their valuable comments and suggestions.
FINANCIAL SUPPORT
The expeditions that collected the material for this paper were funded by the Total Foundation, Price Albert II of Monaco Foundation, Stavras Niarchos Foundation (PANGLAO 2005, SANTO 2006 and MIRIKY), Lounsbery Foundation (AURORA 2007), and Institut pour le Developpement (SALOMAN 2). The MarBOL project in the MNHN is a joint effort with funding from: (1) the Alfred P. Sloan Foundation, PI D. Steinke, co-PIs P. Bouchet and S. Samadi; (2) the Consortium National de Recherche en Génomique and the Service de Systématique Moléculaire (UMS 2700 CNRS–MNHN), part of the agreement 2005/67 between the Genoscope and MNHN on the project Macrophylogeny of Life directed by G. Lecointre; (3) the ATM Barcode (PIs: S. Samadi and J.N. Labat); (4) the Fondation pour la Recherche sur la Biodiversité; and (5) the EDIT programme. This work was partially supported by the grant of RFBR 11–04–01284–a ‘Evolution of digestive system of carnivorous gastropods: testing of morphologically-based hypotheses by molecular data’ (PI Yu. Kantor).










